Analysis of Differentially Expressed Transcripts in Apolygus lucorum (Meyer-Dür) Exposed to Different Temperature Coefficient Insecticides


Abstract
:1. Introduction
2. Results
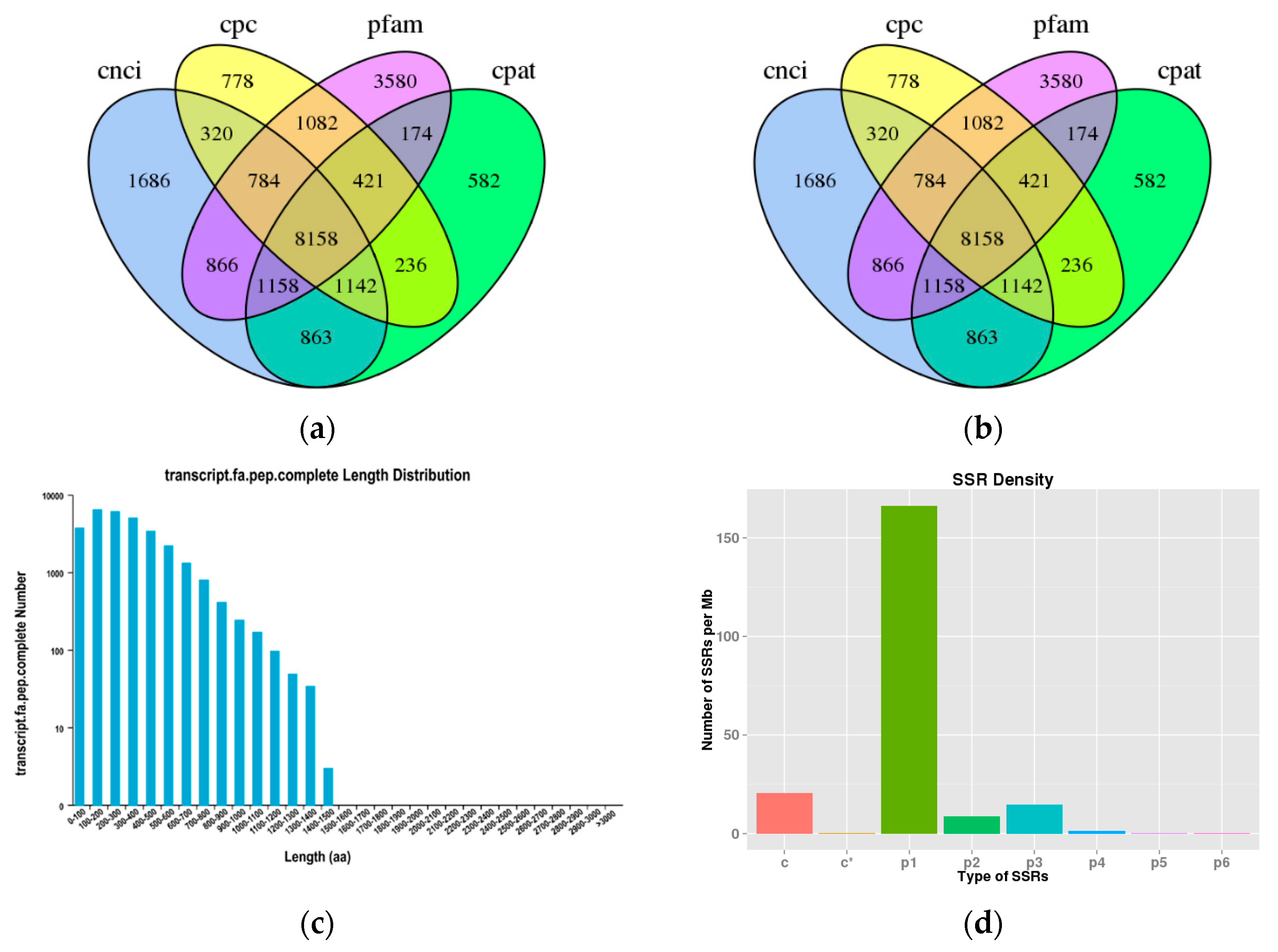
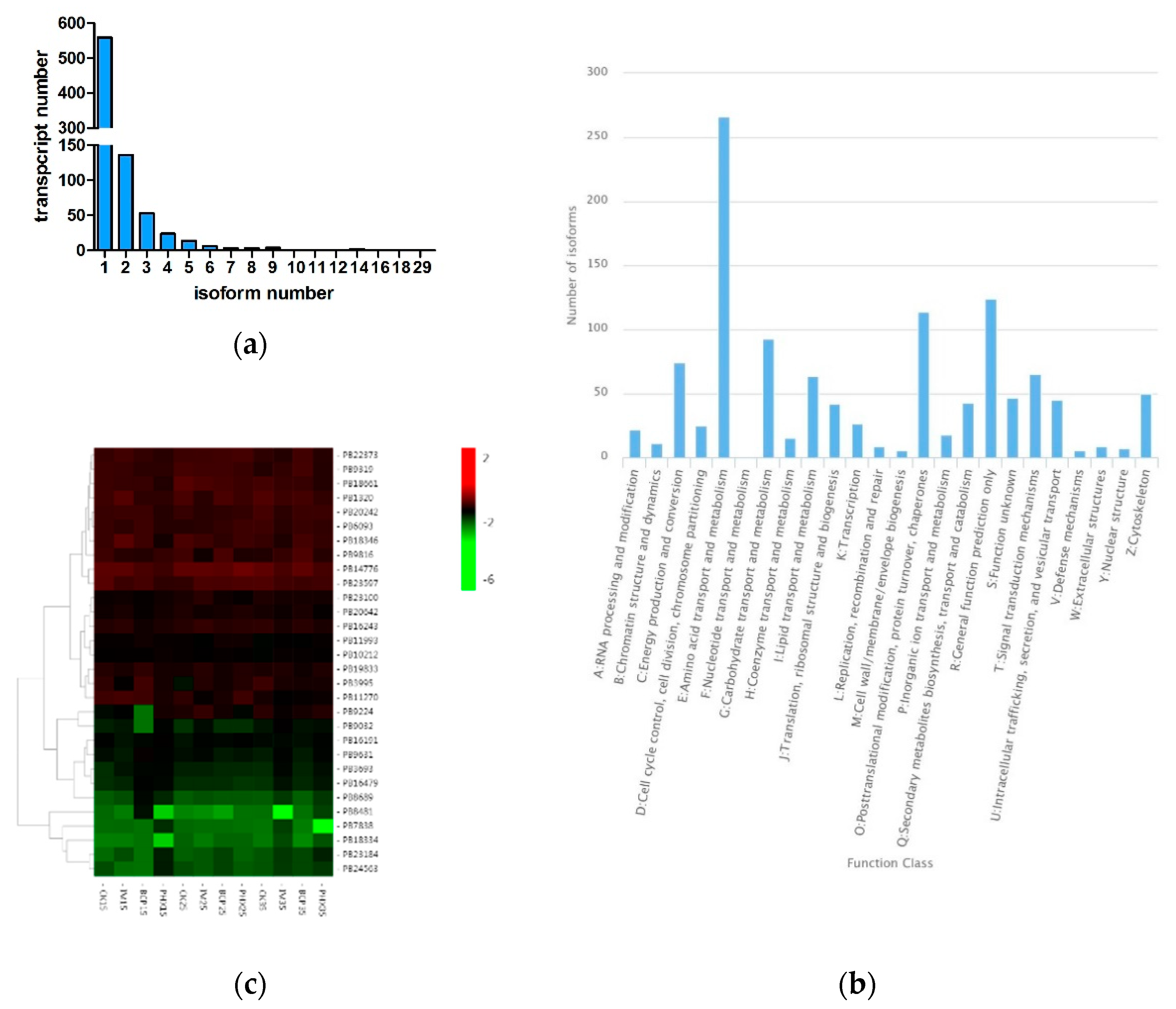
2.1. De Novo Assembly, Functional Annotation and Transcripts Analysis
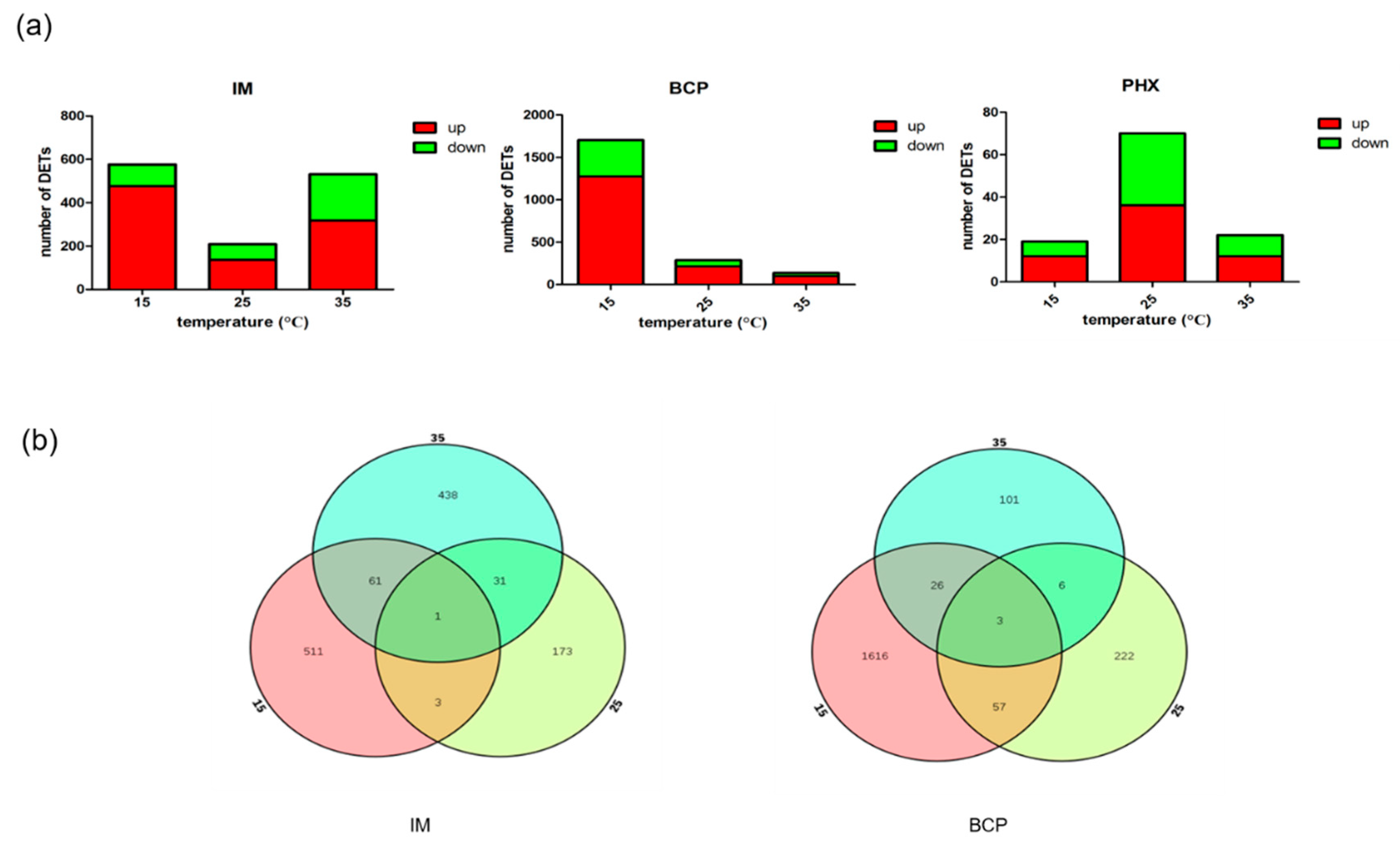
2.2. Identification of Differentially Expressed Transcripts and qRT-PCR Validation
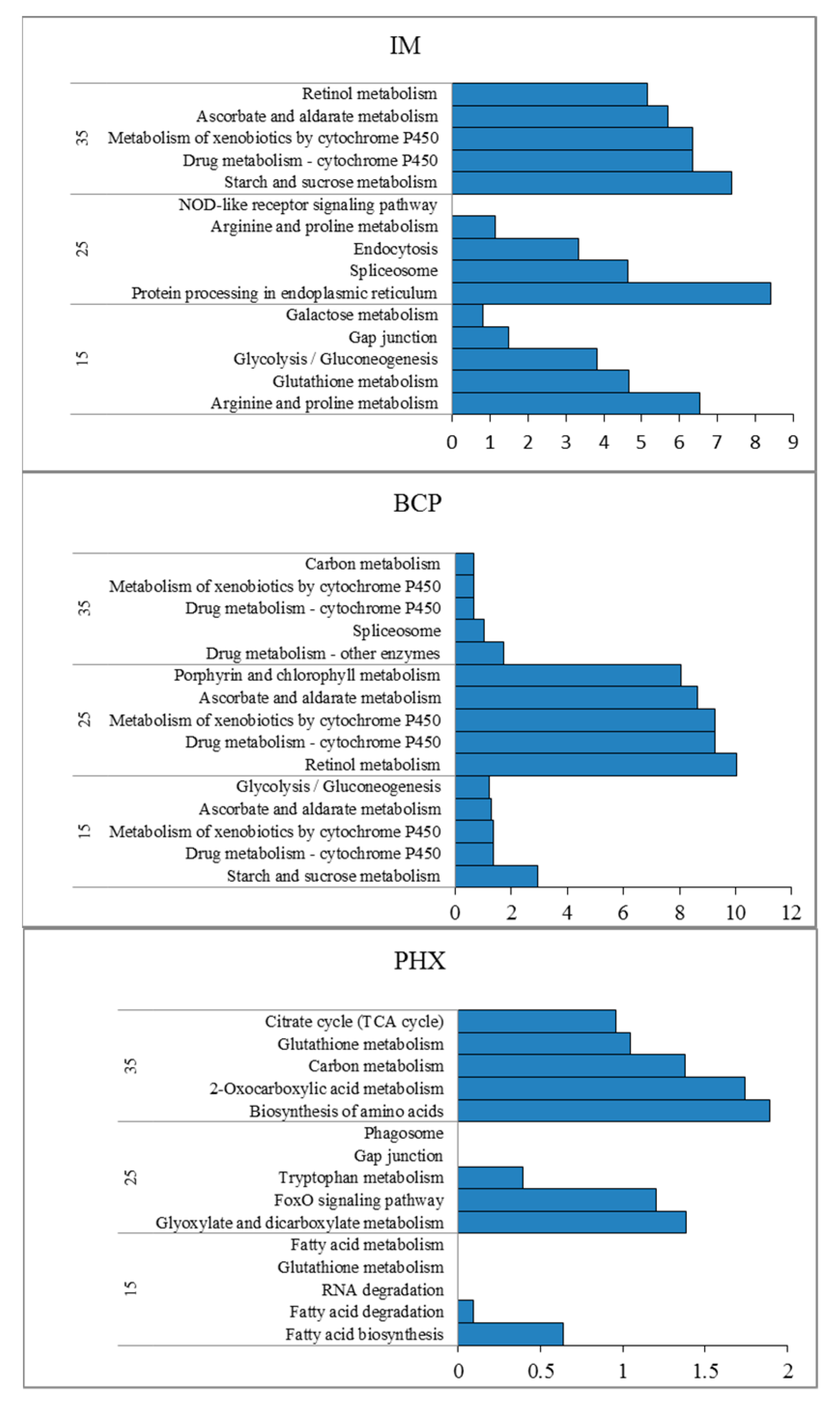
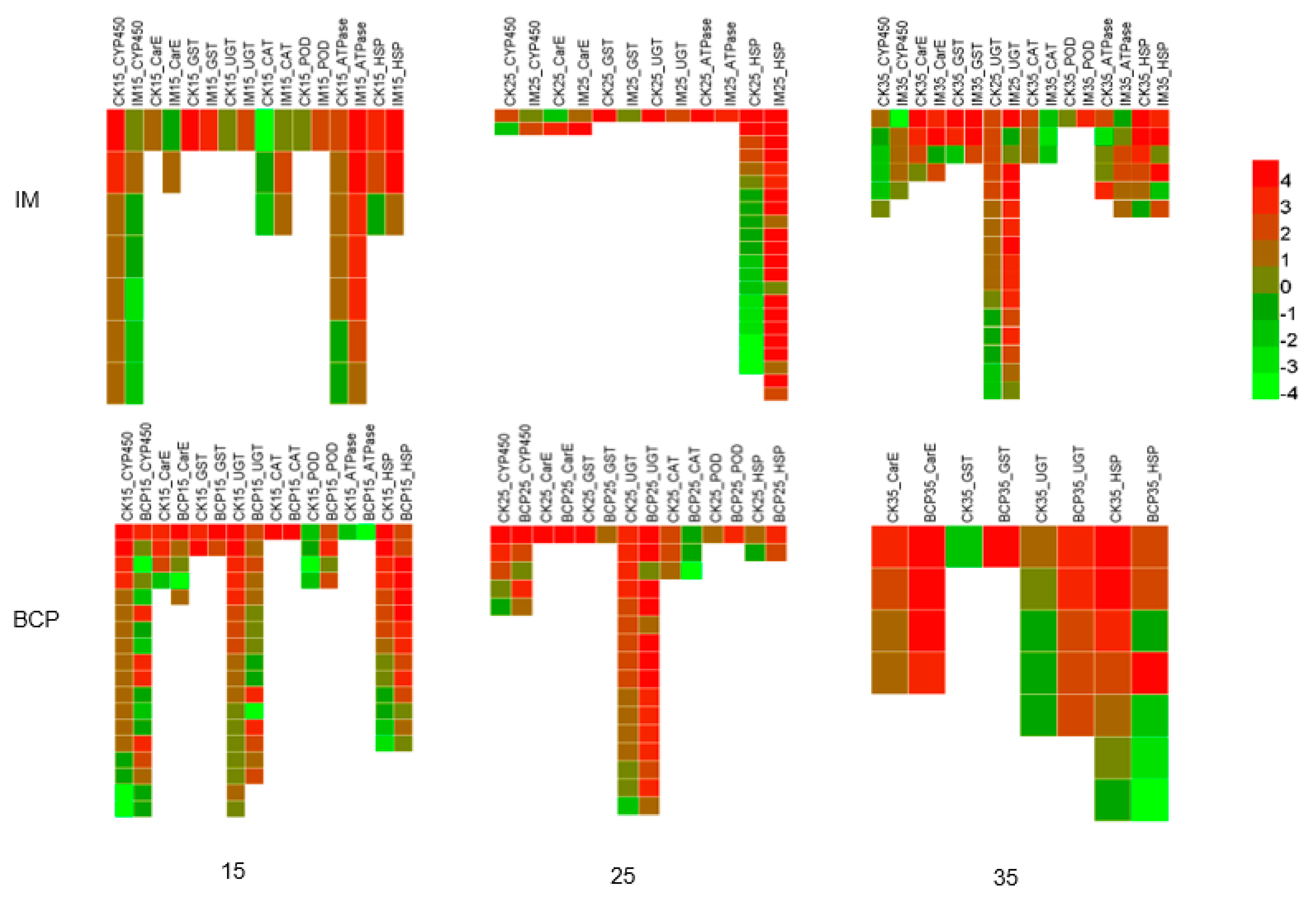
2.3. Pattern Analysis of Temperature Effect Related Transcripts
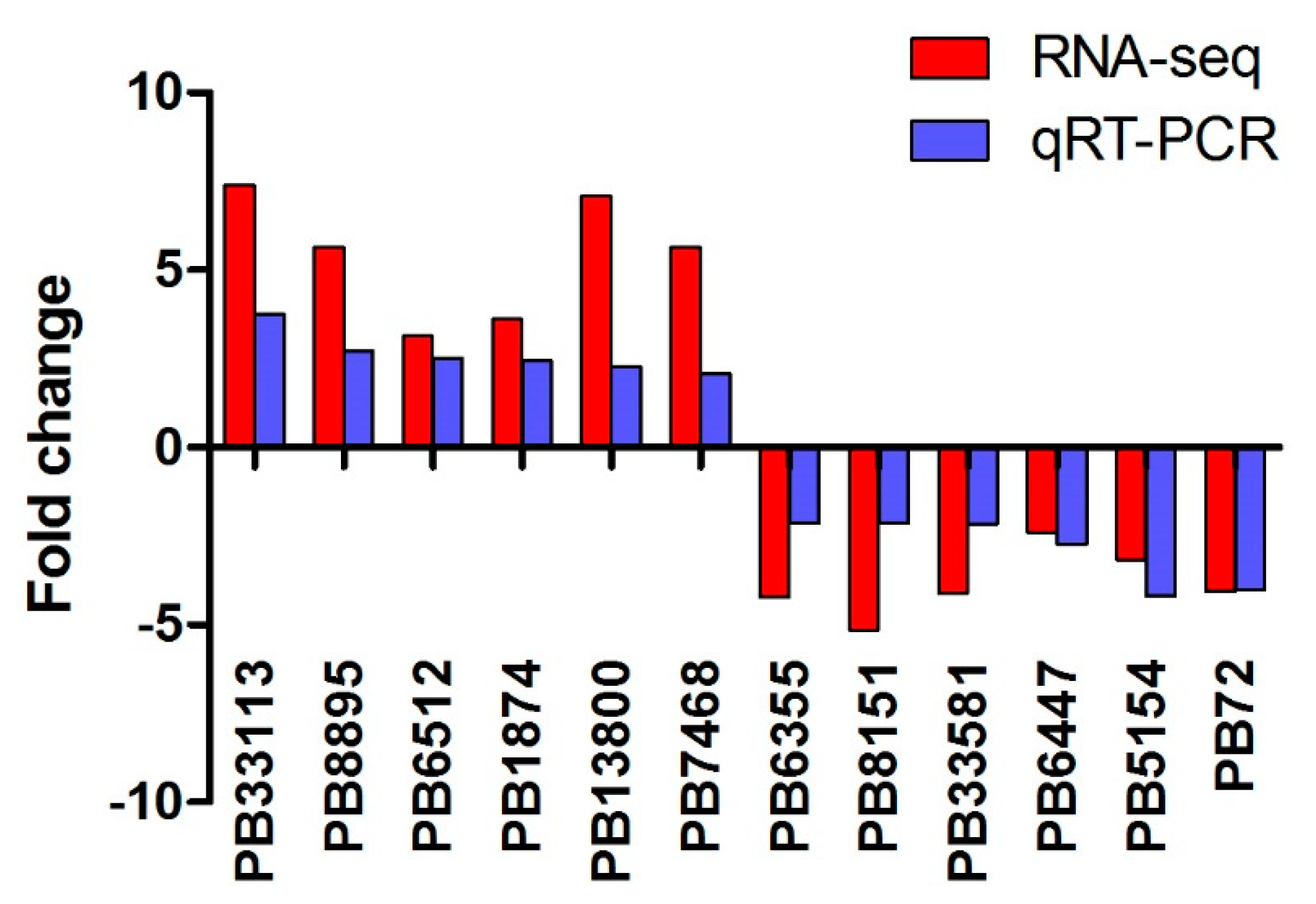
2.4. Validation of RNA-seq by Quantitative Reverse-Transcription PCR
3. Discussion
4. Material and Methods
4.1. Insects Rearing and RNA Sample Preparation
4.2. Library Preparation and SMRT Sequencing
4.3. Next-Generation Library Preparation and Sequencing
4.4. Illumina and Pacbio Data Analysis
4.5. Structure Analysis of Transcripts
4.6. Functional Annotation and Enrichment Analysis
4.7. Accession Number
4.8. Quantitative Real-Time PCR Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| IM | Imidacloprid |
| BCP | Beta-cypermethrin |
| PHX | Phoxim |
| DET | Differentially expressed transcript |
| TC | Temperature coefficient |
| TOI | Toxicity of insecticides |
| nTCIs | Negative temperature insecticides |
| pTCIs | Positive temperature insecticides |
| neTCIs | No effect temperature insecticides |
References
- DeLorenzo, M.E.; Serrano, L.; Chung, K.W.; Hoguet, J.; Key, P.B. Effects of the insecticide permethrin on three life stages of the grass shrimp, Palaemonetes pugio. Ecotoxicol. Environ. Saf. 2006, 64, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Musser, F.R.; Shelton, A.M. The influence of post-exposure temperature on the toxicity of insecticides to Ostrinia nubilalis (Lepidoptera: Crambidae). Pest. Manag. Sci. 2005, 61, 508–510. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Shour, M.H.; Wellemeyer, E.G. Temperature-Toxicity relationships of pyrethroids on three lepidopterans. J. Econ. Entomol. 1982, 75, 643–646. [Google Scholar] [CrossRef]
- Sparks, T.C.; Pavloff, A.M.; Rose, R.L.; Clower, D.F. Temperature-Toxicity relationships of pyrethroids on Heliothis virescens (F.) (Lepidoptera: Noctuidae) and Anthonomus grandis grandis Boheman (Coleoptera: Curculionidae). J. Econ. Entomol. 1983, 76, 243–246. [Google Scholar] [CrossRef]
- Li, H.P.; Feng, T.; Liang, P.; Shi, X.Y.; Gao, X.W.; Jiang, H. Effect of temperature on toxicity of pyrethroids and endosulfan, activity of mitochondrial Na. Pestic. Biochem. Physiol. 2006, 86, 151–156. [Google Scholar] [CrossRef]
- Mao, K.K.; Jin, R.H.; Li, W.H.; Ren, Z.J.; Qin, X.Y.; He, S.; Li, J.H.; Wan, H. The influence of temperature on the toxicity of insecticides to Nilaparvata lugens (Stål). Pestic. Biochem. Physiol. 2019, 156, 80–86. [Google Scholar] [CrossRef]
- IUPS. Glossary of terms for thermal physiology. Pflug. Arch. Eur. J. Phy. 2001, 51, 245–280. [Google Scholar]
- Crawshaw, L.I.; Christopher, J. Gordon, Review of: Temperature and Toxicology: An Integrative, Comparative, and Environmental Approach. J. Therm. Biol. 2005, 30, 628–629. [Google Scholar] [CrossRef]
- Norment, B.R.; Chambeas, H.W. Temperature Relationships in Organophosphorus Poisoning in Boll Weevils12. J. Econ. Entomol. 1970, 63, 502–504. [Google Scholar] [CrossRef]
- Chalfant, R.B. Cabbage Looper: Effect of Temperature on Toxicity of Insecticides in the Laboratory1. J. Econ. Entomol. 1973, 66, 339–341. [Google Scholar] [CrossRef]
- Satpute, N.S.; Deshmukh, S.D.; Rao, N.G.V.; Tikar, S.N.; Moharil, M.P.; Nimbalkar, S.A. Temperature-Dependent Variation in Toxicity of Insecticides Against Earias vitella (Lepidoptera: Noctuidae). J. Econ. Entomol. 2007, 100, 357–360. [Google Scholar] [CrossRef]
- Wang, K. Toxicity of Imidacloprid on Six Aphids and Temperature Effects. Pesticides 1999, 7, 15–16. [Google Scholar]
- Ma, Y.H.; Gao, Z.L.; Dang, Z.H.; Li, Y.F.; Pan, W.L. Effect of temperature on the toxicity of several insecticides to Apolygus lucorum (Heteroptera: Miridae). J. Pestic. Sci. 2012, 37, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Mansoor, M.M.; Afzal, M.; Raza, A.B.M.; Akram, Z.; Waqar, A.; Afzal, M.B.S. Post-exposure temperature influence on the toxicity of conventional and new chemistry insecticides to green lacewing Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). Saudi J. Biol. Sci. 2015, 22, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.G. Effects of temperature on insecticide toxicity. Rev. Pestic. Toxicol. 1995, 3, 111–135. [Google Scholar]
- Lu, Y.H.; Wu, K.M. Biology and Control of Cotton Mirids; Golden Shield Press: Beijing, China, 2008. [Google Scholar]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Xia, B.; Li, P.; Feng, H.Q.; Wyckhuys, K.A.G.; Guo, Y.Y. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China. Science 2010, 328, 1151–1154. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.H.; Qiu, F.; Feng, H.Q.; Li, H.B.; Yang, Z.C.; Wyckhuys, K.A.G.; Wu, K.M. Species composition and seasonal abundance of pestiferous plant bugs (Hemiptera: Miridae) on Bt Cotton in China. Crop. Prot. 2008, 27, 465–472. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Lu, Y.H.; Zeng, J. Forecast, and Management of Mirid Bugs in Multiple Agroecosystems of China; China Agriculture Press: Beijing, China, 2015. [Google Scholar]
- Zhang, Z.Q.; Wang, Y.; Zhao, Y.H.; Li, B.X.; Lin, J.; Zhang, X.F.; Liu, F.; Mu, W. Nitenpyram seed treatment effectively controls against the mirid bug Apolygus lucorum in cotton seedlings. Sci. Rep. 2017, 7, 8573. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Dang, Z.H.; An, J.J.; Gao, Z.L.; Pan, W.L.; Li, Y.F. Temperature effect of different mechanism insecticides on Apolgus lucorum meyer-dür(hemiptera: Miridae). Chin. Agric. Sci. Bull. 2016, 32, 70–75. [Google Scholar]
- Liu, J.; Lincoln, T.R.; An, J.J.; Gao, Z.L.; Dang, Z.H.; Pan, W.L.; Li, Y.F. The Joint Toxicity of Different Temperature Coefficient Insecticides on Apolygus lucorum (Hemiptera: Miridae). J. Econ. Entomol. 2016, 109, 1846–1852. [Google Scholar] [CrossRef]
- Scott, J.G.; Georghiou, G.P. Influence of temperature on knockdown, toxicity, and resistance to pyrethroids in the house fly, Musca domestica. Pestic. Biochem. Physiol. 1984, 21, 53–62. [Google Scholar] [CrossRef]
- Laetz, C.A.; Baldwin, D.H.; Hebert, V.R.; Stark, J.D.; Scholz, N.L. Elevated temperatures increase the toxicity of pesticide mixtures to juvenile coho salmon. Aquat. Toxicol. 2014, 146, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Weston, D.P.; You, J.; Harwood, A.D.; Lydy, M.J. Whole sediment toxicity identification evaluation tools for pyrethroid insecticides: III. Temperature manipulation. Environ. Toxicol. Chem. 2009, 28, 173–180. [Google Scholar] [CrossRef]
- Feng, H.Z.; Wang, L.; Liu, Y.H.; He, L.; Li, M.; Lu, W.C.; Xue, C.H. Molecular Characterization and Expression of a Heat Shock Protein Gene (HSP90) from the Carmine Spider Mite, Tetranychus cinnabarinus (Boisduval). J. Insect Sci. 2010, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.H.; Wang, C.Z.; Kang, L. Cloning and expression of five heat shock protein genes in relation to cold hardening and development in the leafminer, Liriomyza sativa. J. Insect Physiol. 2009, 55, 279–285. [Google Scholar] [CrossRef]
- Sun, Y.; Sheng, Y.; Bai, L.X.; Zhang, Y.J.; Xiao, Y.F.; Xiao, L.B.; Tan, Y.G.; Shen, Y.M. Characterizing heat shock protein 90 gene of Apolygus lucorum (Meyer-Dür) and its expression in response to different temperature and pesticide stresses. Cell Stress Chaperon. 2014, 19, 725–739. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Huang, L.H.; Zhang, A.B. High-throughput transcriptome sequencing technology and its applications in Lepidoptera. Acta Entomol. Sin. 2014, 57, 991–1000. [Google Scholar]
- Yuan, H.B.; Ding, Y.X.; Gu, S.H.; Sun, L.; Zhu, X.Q.; Liu, H.W.; Dhiloo, K.H.; Zhang, Y.J.; Guo, Y.Y. Molecular characterization and expression profiling of odorant-binding proteins in Apolygus lucorum. PLoS ONE 2015, 10, e0140562. [Google Scholar] [CrossRef] [Green Version]
- An, X.K.; Sun, L.; Liu, H.W.; Liu, D.F.; Ding, Y.X.; Li, L.M.; Zhang, Y.J.; Guo, Y.Y. Identification and expression analysis of an olfactory receptor gene family in green plant bug Apolygus lucorum (Meyer-Dür). Sci. Rep. 2016, 6, 37870. [Google Scholar] [CrossRef] [Green Version]
- Cao, D.P.; Liu, Y.; Wei, J.J.; Li, J.H.; Wang, G.R. A de novo transcriptomic analysis to reveal functional genes in Apolygus lucorum. Insect Sci. 2016, 23, 2–14. [Google Scholar] [CrossRef]
- Sharon, D.; Tilgner, H.; Grubert, F.; Snyder, M. A single-molecule long-read survey of the human transcriptome. Nat. Biotechnol. 2013, 31, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bracht, J.R.; Goldman, A.D.; Dolzhenko, E.; Clay, D.M.; Swart, E.C.; Perlman, D.H.; Doak, T.G.; Stuart, A.; Amemiya, C.T.; et al. The architecture of a scrambled genome reveals massive levels of genomic rearrangement during development. Cell 2014, 158, 1187–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Liu, J.X.; Wang, X.S.; Wei, Z.W. Full-length RNA sequencing reveals unique transcriptome composition in bermudagrass. Plant. Physiol. Biochem. 2018, 132, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Tetreau, G.; Stalinski, R.; David, J.P.; Després, L. Increase In Larval Gut Proteolytic Activities And Bti Resistance In The Dengue Fever Mosquito. Arch. Insect Biochem. Physiol. 2013, 82, 71–83. [Google Scholar] [CrossRef]
- Boina, D.R.; Onagbola, E.O.; Salyani, M.; Stelinski, L.L. Influence of Posttreatment Temperature on the Toxicity of Insecticides Against Diaphorina citri (Hemiptera: Psyllidae). J. Econ. Entomol. 2009, 102, 685–691. [Google Scholar] [CrossRef]
- DAS, M.; McINTOSH, A.H. Effect of time and temperature on toxicity of insecticides to insects. Ann. Appl. Biol. 1961, 49, 267–289. [Google Scholar] [CrossRef]
- Wang, C.M.; Narahashi, T.; Scuka, M. Mechanism of negative temperature coefficient of nerve blocking action of allethrin. J. Pharmacol. Exp. Ther. 1972, 182, 442–453. [Google Scholar]
- Glunt, K.D.; Oliver, S.V.; Hunt, R.H.; Paaijmans, K.P. The impact of temperature on insecticide toxicity against the malaria vectors Anopheles arabiensis and Anopheles funestus. Malar. J. 2018, 17, 131. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.F.; Chai, C.L.; Zhang, Z.; Liu, Z.H.; Dai, F.Y.; Lu, C.; Xiang, Z.H. The UDP-glucosyltransferase multigene family in Bombyx mori. BMC Genom. 2008, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.J.; Wang, Z.B.; Li, C.F.; Liu, J.L.; Zeng, X.N. UDP-Glycosyltransferases are involved in imidacloprid resistance in the Asian citrus psyllid, Diaphorina citri (Hemiptera: Lividae). Pestic. Biochem. Physiol. 2019, 154, 23–31. [Google Scholar] [CrossRef]
- Xu, J.; Strange, J.P.; Welker, D.L.; James, R.R. Detoxification and stress response genes expressed in a western North American bumble bee, Bombus huntii (Hymenoptera: Apidae). BMC Genom. 2013, 14, 874. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rojas, J.C.; Robles-Bermudez, A.; Cambero-Campos, O.J.; Carvajal-Cazola, C.R.; Pena-Sandoval, G.R. Metabolic resistance to pesticides. Rev. BIO CIENCIAS 2017, 6, 1–16. [Google Scholar]
- Johnston, J.A.; Ward, C.L.; Kopito, R.R. Aggresomes: A cellular response to misfolded proteins. J. Cell Biol. 1998, 143, 1883–1898. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Liu, X.Q.; Xiao, P.A.; Tang, G.H.; Liu, S.H.; Lou, B.H.; Wang, J.J.; Jiang, H.B. Comparative transcriptome analysis reveals differentially expressed genes in the Asian citrus psyllid (Diaphorina citri) upon heat shock. Comp. Biochem. Physiol. Part. D Genom. Proteom. 2019, 30, 256–261. [Google Scholar] [CrossRef]
- Gu, Z.Y.; Sun, S.S.; Wang, Y.H.; Wang, B.B.; Xie, Y.; Ma, L.; Wang, J.M.; Shen, W.D.; Li, B. Transcriptional characteristics of gene expression in the midgut of domestic silkworms (Bombyx mori) exposed to phoxim. Pestic. Biochem. Physiol. 2013, 105, 36–43. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cDNA Size | Reads of Insert | Number of Filtered Short Reads | Number of Non-Full-Length Reads | Number of Full-Length Reads | Number of Full-Length Non-Chimeric Reads | Average Full-Length Non-Chimeric Read Length | Full-Length Percentage (FL%) |
|---|---|---|---|---|---|---|---|
| 1–2 K | 201,132 | 32,182 | 86,205 | 82,745 | 82,235 | 1475 | 41.14% |
| 2–3 K | 217,069 | 14,850 | 84,544 | 117,675 | 117,094 | 2392 | 54.21% |
| 3–6 K | 88,975 | 3374 | 41,772 | 43,829 | 43,797 | 3542 | 49.26% |
| All | 507,176 | 50,406 | 212,521 | 244,249 | 243,126 | 2288 | 48.16% |
| Pesticide | Temperature | GO_ID | GO_Term | p_adj |
|---|---|---|---|---|
| IM | 15 | GO:0008238 | exopeptidase activity | 1.94 × 10−5 |
| 25 | GO:0008238 | exopeptidase activity | 8.25 × 10−5 | |
| GO:0000062 | fatty-acyl-CoA binding | 2.42 × 10−2 | ||
| 35 | GO:0003844 | 1,4-alpha-glucan branching enzyme activity | 3.53 × 10−4 | |
| GO:0043169 | cation binding | 6.95 × 10−3 | ||
| GO:0009982 | pseudouridine synthase activity | 1.29 × 10−2 | ||
| GO:0016208 | AMP binding | 2.68 × 10−2 | ||
| GO:0016831 | carboxy-lyase activity | 3.47 × 10−2 | ||
| BCP | 15 | GO:0008238 | exopeptidase activity | 8.26 × 10−3 |
| GO:0004368 | glycerol-3-phosphate dehydrogenase activity | 4.42 × 10−2 | ||
| 25 | GO:0042302 | structural constituent of cuticle | 0 | |
| 35 | GO:0016798 | hydrolase activity, acting on glycosyl bonds | 4.22 × 10−7 | |
| GO:0004650 | polygalacturonase activity | 6.40 × 10−6 | ||
| GO:0051287 | NAD binding | 2.71 × 10−2 | ||
| GO:0004617 | phosphoglycerate dehydrogenase activity | 3.64 × 10−2 | ||
| PHX | 15 | GO:0004726 | non-membrane spanning protein tyrosine phosphatase activity | 1.61 × 10−2 |
| GO:0004467 | long-chain fatty acid-CoA ligase activity | 2.42 × 10−2 | ||
| 25 | GO:0042302 | structural constituent of cuticle | 4.66 × 10−3 | |
| GO:0008469 | histone-arginine N-methyltransferase activity | 3.42 × 10−2 | ||
| 35 | GO:0000287 | magnesium ion binding | 1.53 × 10−4 | |
| GO:0004450 | isocitrate dehydrogenase (NADP+) activity | 1.62 × 10−4 | ||
| GO:0008012 | structural constituent of adult chitin-based cuticle | 8.06 × 10−3 | ||
| GO:0051287 | NAD binding | 9.15 × 10−3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, J.; Liu, C.; Dou, Y.; Gao, Z.; Dang, Z.; Yan, X.; Pan, W.; Li, Y. Analysis of Differentially Expressed Transcripts in Apolygus lucorum (Meyer-Dür) Exposed to Different Temperature Coefficient Insecticides. Int. J. Mol. Sci. 2020, 21, 658. https://doi.org/10.3390/ijms21020658
An J, Liu C, Dou Y, Gao Z, Dang Z, Yan X, Pan W, Li Y. Analysis of Differentially Expressed Transcripts in Apolygus lucorum (Meyer-Dür) Exposed to Different Temperature Coefficient Insecticides. International Journal of Molecular Sciences. 2020; 21(2):658. https://doi.org/10.3390/ijms21020658
Chicago/Turabian StyleAn, Jingjie, Chang Liu, Ya’nan Dou, Zhanlin Gao, Zhihong Dang, Xiu Yan, Wenliang Pan, and Yaofa Li. 2020. "Analysis of Differentially Expressed Transcripts in Apolygus lucorum (Meyer-Dür) Exposed to Different Temperature Coefficient Insecticides" International Journal of Molecular Sciences 21, no. 2: 658. https://doi.org/10.3390/ijms21020658
APA StyleAn, J., Liu, C., Dou, Y., Gao, Z., Dang, Z., Yan, X., Pan, W., & Li, Y. (2020). Analysis of Differentially Expressed Transcripts in Apolygus lucorum (Meyer-Dür) Exposed to Different Temperature Coefficient Insecticides. International Journal of Molecular Sciences, 21(2), 658. https://doi.org/10.3390/ijms21020658