Extracellular Vesicles Mediated Early Embryo–Maternal Interactions



Abstract
:1. Introduction
2. Embryo–Maternal Interactions Mediated by Embryotropins
3. Extracellular Vesicles Derived from Female Reproductive Tract and Embryo
4. Embryo–Maternal Interactions through Oviductal EVs
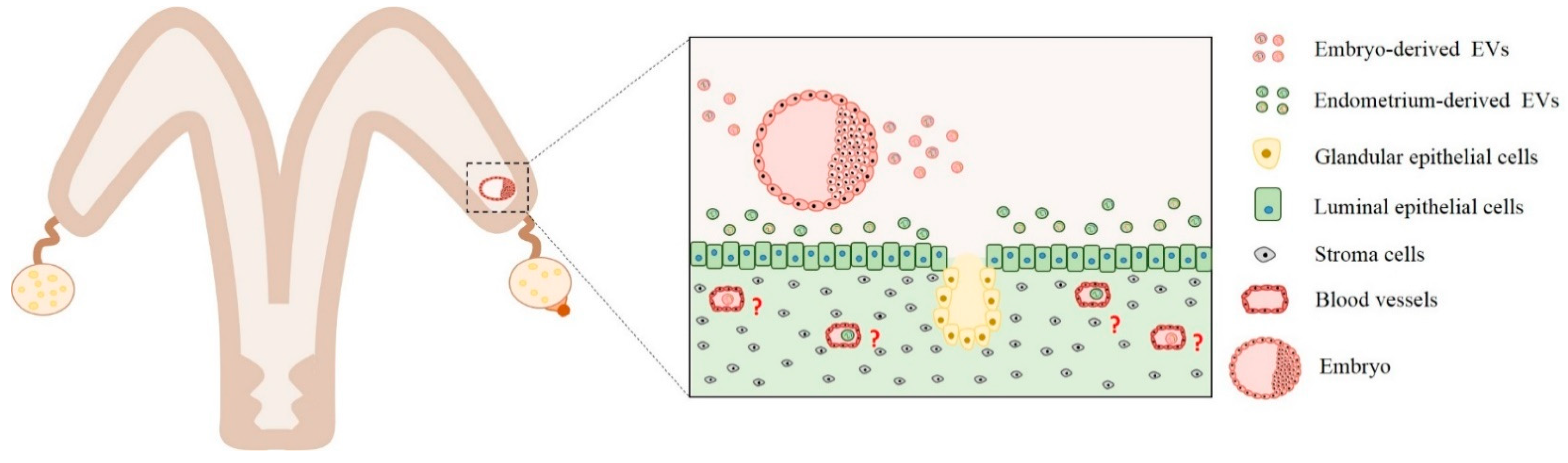
5. Embryo–Maternal Interactions between Uterus and Embryo Mediated by EVs
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holm, P.; Callesen, H. In vivo versus in vitro produced bovine ova: Similarities and differences relevant for practical application. Reprod. Nutr. Dev. 1998, 38, 579–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betteridge, K.J.; Fléchon, J.E. The anatomy and physiology of pre-attachment bovine embryos. Theriogenology 1988, 29, 155–187. [Google Scholar] [CrossRef]
- Memili, E.; Dominko, T.; First, N.L. Onset of transcription in bovine oocytes and preimplantation embryos. Mol. Reprod. Dev. 1998, 51, 36–41. [Google Scholar] [CrossRef]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Dossary, A.A.; Strehler, E.E.; Martin-DeLeon, P.A. Expression and Secretion of Plasma Membrane Ca2+-ATPase 4a (PMCA4a) during Murine Estrus: Association with Oviductal Exosomes and Uptake in Sperm. PLoS ONE 2013, 8, e80181. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.M.; Wang, H.S.; Lee, C.L.; Lee, J.D.; Huang, H.Y.; Chang, F.H.; Lee, J.F.; Soong, Y.K. Insulin-like growth factor-binding proteins produced by Vero cells, human oviductal cells and human endometrial cells, and the role of insulin-like growth factor-binding protein-3 in mouse embryo co-culture systems. Hum. Reprod. 1996, 11, 1281–1286. [Google Scholar] [CrossRef] [Green Version]
- Lopera-Vásquez, R.; Hamdi, M.; Fernandez-Fuertes, B.; Maillo, V.; Beltrán-Breña, P.; Calle, A.; Redruello, A.; López-Martín, S.; Gutierrez-Adán, A.; Yañez-Mó, M.; et al. Extracellular Vesicles from BOEC in In Vitro Embryo Development and Quality. PLoS ONE 2016, 11, e0148083. [Google Scholar] [CrossRef]
- Leese, H.J.; Hugentobler, S.A.; Gray, S.M.; Morris, D.G.; Sturmey, R.G.; Whitear, S.-L.; Sreenan, J.M. Female reproductive tract fluids: Composition, mechanism of formation and potential role in the developmental origins of health and disease. Reprod. Fertil. Dev. 2007, 20, 1–8. [Google Scholar] [CrossRef]
- Spencer, T.E. Early pregnancy: Concepts, challenges, and potential solutions. Anim. Front. 2013, 3, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Guillomot, M. Cellular interactions during implantation in domestic ruminants. J. Reprod. Fertil. Suppl. 1995, 49, 39–51. [Google Scholar] [CrossRef]
- Forde, N.; Lonergan, P. Transcriptomic Analysis of the Bovine Endometrium: What is Required to Establish Uterine Receptivity to Implantation in Cattle ? J. Reprod. Dev. 2012, 58, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, Q.; Wang, H.; Duan, E. Uterine Fluid in Pregnancy: A Biological and Clinical Outlook. Trends Mol. Med. 2017, 23, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.H.; Rome, S.; Jalabert, A.; Forterre, A.; Singh, H.; Hincks, C.L.; Salamonsen, L.A. Endometrial Exosomes/Microvesicles in the Uterine Microenvironment: A New Paradigm for Embryo-Endometrial Cross Talk at Implantation. PLoS ONE 2013, 8, e58502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannetta, D.; Dragovic, R.; Alyahyaei, Z.; Southcombe, J. Extracellular vesicles and reproduction-promotion of successful pregnancy. Cell. Mol. Immunol. 2014, 11, 548–563. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Bazer, F.W. Temporal and spatial alterations in uterine estrogen receptor and progesterone receptor gene expression during the estrous cycle and early pregnancy in the ewe. Biol. Reprod. 1995, 53, 1527–1543. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.M.; Chen, Y.; Ezashi, T.; Walker, A.M. Interferons and the maternal–conceptus dialog in mammals. Semin. Cell Dev. Biol. 2008, 19, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Imakawa, K.; Anthony, R.V.; Kazemi, M.; Marotti, K.R.; Polites, H.G.; Roberts, R.M. Interferon-like sequence of ovine trophoblast protein secreted by embryonic trophectoderm. Nature 1987, 330, 377–379. [Google Scholar] [CrossRef]
- Bazer, F.W.; Ott, T.L.; Spencer, T.E. Pregnancy recognition in ruminants, pigs and horses: Signals from the trophoblast. Theriogenology 1994, 41, 79–94. [Google Scholar] [CrossRef]
- Ticconi, C.; Zicari, A.; Belmonte, A.; Realacci, M.; Rao, C.V.; Piccione, E. Pregnancy-Promoting Actions of HCG in Human Myometrium and Fetal Membranes. Placenta 2007, 28, S137–S143. [Google Scholar] [CrossRef]
- Mellisho, E.A.; Velásquez, A.E.; Nuñez, M.J.; Cabezas, J.G.; Cueto, J.A.; Fader, C.; Castro, F.O.; Rodríguez-Álvarez, L. Identification and characteristics of extracellular vesicles from bovine blastocysts produced in vitro. PLoS ONE 2017, 12, e0178306. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; Spencer, T.E. Extracellular Vesicles Originate from the Conceptus and Uterus During Early Pregnancy in Sheep. Biol. Reprod. 2016, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Kusama, K.; Bai, R.; Sakurai, T.; Isuzugawa, K.; Godkin, J.D.; Suda, Y.; Imakawa, K. Induction of IFNT-Stimulated Genes by Conceptus-Derived Exosomes during the Attachment Period. PLoS ONE 2016, 11, e0158278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, É.; Pap, E.; Kittel, Á.; et al. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cell. Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [Green Version]
- Da Silveira, J.C.; Veeramachaneni, D.N.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain miRNAs and Proteins: A Possible New Form of Cell Communication Within the Ovarian Follicle. Biol. Reprod. 2012, 86. [Google Scholar] [CrossRef]
- Andrade, G.; Meirelles, F.; Perecin, F.; da Silveira, J. Cellular and extracellular vesicular origins of miRNAs within the bovine ovarian follicle. Reprod. Domest. Anim. 2017, 52, 1036–1045. [Google Scholar] [CrossRef]
- Greening, D.W.; Nguyen, H.P.T.; Elgass, K.; Simpson, R.J.; Salamonsen, L.A. Human Endometrial Exosomes Contain Hormone-Specific Cargo Modulating Trophoblast Adhesive Capacity: Insights into Endometrial-Embryo Interactions. Biol. Reprod. 2016, 94, 38. [Google Scholar] [CrossRef]
- Kropp, J.; Salih, S.M.; Khatib, H. Expression of microRNAs in bovine and human pre-implantation embryo culture media. Front. Genet. 2014, 5, 91. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.; Lotvall, J. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Jensen, S.S.; Lim, J.W. Proteomic profiling of exosomes: Current perspectives. Proteomics 2008, 8, 4083–4099. [Google Scholar] [CrossRef]
- Subra, C.; Laulagnier, K.; Perret, B.; Record, M. Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies. Biochimie 2007, 89, 205–212. [Google Scholar] [CrossRef]
- Saadeldin, I.M.; Oh, H.J.; Lee, B.C. Embryonic–maternal cross-talk via exosomes: Potential implications. Stem Cells Cloning Adv. Appl. 2015, 8, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Wydooghe, E.; Vandaele, L.; Heras, S.; De Sutter, P.; Deforce, D.; Peelman, L.; De Schauwer, C.; Van Soom, A. Autocrine embryotropins revisited: How do embryos communicate with each other in vitro when cultured in groups? Biol. Rev. 2017, 92, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.M.; Ealy, A.D.; Alexenko, A.P.; Han, C.S.; Ezashi, T. Trophoblast Interferons. Placenta 1999, 20, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Austin, K.J.; Carr, A.L.; Pru, J.K.; Hearne, C.E.; George, E.L.; Belden, E.L.; Hansen, T.R. Localization of ISG15 and Conjugated Proteins in Bovine Endometrium Using Immunohistochemistry and Electron Microscopy. Endocrinology 2004, 145, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.F.; Henkes, L.E.; Ashley, R.L.; Purcell, S.H.; Smirnova, N.P.; Veeramachaneni, D.N.R.; Anthony, R.V.; Hansen, T.R. Expression of Interferon (IFN)-Stimulated Genes in Extrauterine Tissues during Early Pregnancy in Sheep Is the Consequence of Endocrine IFN-τ Release from the Uterine Vein. Endocrinology 2008, 149, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Antoniazzi, A.Q.; Webb, B.T.; Romero, J.J.; Ashley, R.L.; Smirnova, N.P.; Henkes, L.E.; Bott, R.C.; Oliveira, J.F.; Niswender, G.D.; Bazer, F.W.; et al. Endocrine delivery of interferon tau protects the corpus luteum from prostaglandin F2 alpha-induced luteolysis in ewes. Biol Reprod 2013, 88, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridi, A.; Bertolin, K.; Rissi, V.B.; Mujica, L.K.S.; Glanzner, W.G.; Macedo, M.P.d.; Comim, F.V.; Gonçalves, P.B.D.; Antoniazzi, A.Q. Parthenogenetic bovine embryos secrete type I interferon capable of stimulating ISG15 in luteal cell culture. Anim. Reprod. 2018, 15, 1268–1277. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Austin, K.J.; Rempel, L.A.; Hansen, T.R. Low blood ISG15 mRNA and progesterone levels are predictive of non-pregnant dairy cows. J. Endocrinol. 2006, 191, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Meyerholz, M.M.; Mense, K.; Knaack, H.; Sandra, O.; Schmicke, M. Pregnancy-Induced ISG-15 and MX-1 Gene Expression is Detected in the Liver of Holstein–Friesian Heifers During Late Peri-Implantation Period. Reprod. Domest. Anim. 2016, 51, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Sponchiado, M.; Gomes, N.S.; Fontes, P.K.; Martins, T.; del Collado, M.; Pastore, A.d.A.; Pugliesi, G.; Nogueira, M.F.G.; Binelli, M. Pre-hatching embryo-dependent and -independent programming of endometrial function in cattle. PLoS ONE 2017, 12, e0175954. [Google Scholar] [CrossRef] [PubMed]
- Sponchiado, M.; Gonella-Diaza, A.M.; Rocha, C.C.; Turco, E.G.L.; Pugliesi, G.; Leroy, J.L.M.R.; Binelli, M. The pre-hatching bovine embryo transforms the uterine luminal metabolite composition in vivo. Sci. Rep. 2019, 9, 8354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebington, C.; Doherty, F.J.; Fleming, S.D. Ubiquitin cross-reactive protein gene expression is increased in decidualized endometrial stromal cells at the initiation of pregnancy. Mol. Hum. Reprod. 1999, 5, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Austin, K.J.; Bany, B.M.; Belden, E.L.; Rempel, L.A.; Cross, J.C.; Hansen, T.R. Interferon-Stimulated Gene-15 (Isg15) Expression Is Up-Regulated in the Mouse Uterus in Response to the Implanting Conceptus. Endocrinology 2003, 144, 3107–3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashley, R.L.; Henkes, L.E.; Bouma, G.J.; Pru, J.K.; Hansen, T.R. Deletion of the Isg15 Gene Results in Up-Regulation of Decidual Cell Survival Genes and Down-Regulation of Adhesion Genes: Implication for Regulation by IL-1β. Endocrinology 2010, 151, 4527–4536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Forde, N.; Dorniak, P.; Hansen, T.R.; Romero, J.J.; Lonergan, P. Conceptus-derived prostaglandins regulate gene expression in the endometrium prior to pregnancy recognition in ruminants. Reproduction 2013, 146, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, T.; Gillio-Meina, C.; Phang, S. Prostaglandins and the initiation of blastocyst implantation and decidualization. Reproduction 2007, 134, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Ottobre, J.S.; Vincent, D.L.; Silvia, W.J.; Inskeep, E.K. Aspects of regulation of uterine secretion of prostaglandins during the oestrous cycle and early pregnancy. Anim. Reprod. Sci. 1984, 7, 75–100. [Google Scholar] [CrossRef]
- Weber, J.A.; Freeman, D.A.; Vanderwall, D.K.; Woods, G.L. Prostaglandin E2 Secretion by Oviductal Transport-Stage Equine Embryos1. Biol. Reprod. 1991, 45, 540–543. [Google Scholar] [CrossRef]
- Weber, J.A.; Freeman, D.A.; Vanderwall, D.K.; Woods, G.L. Prostaglandin E2 Hastens Oviductal Transport of Equine Embryos1. Biol. Reprod. 1991, 45, 544–546. [Google Scholar] [CrossRef]
- Woclawek-Potocka, I.; Brzezicka, E.; Skarzynski, D.J. Lysophosphatic Acid Modulates Prostaglandin Secretion in the Bovine Endometrial Cells Differently on Days 8-10 of the Estrous Cycle and Early Pregnancy. J. Reprod. Dev. 2009, 55, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.M.; Adam, M.; Hammond, J.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [PubMed]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569. [Google Scholar] [CrossRef]
- Morelli, A.E.; Larregina, A.T.; Shufesky, W.J.; Sullivan, M.L.G.; Stolz, D.B.; Papworth, G.D.; Zahorchak, A.F.; Logar, A.J.; Wang, Z.; Watkins, S.C.; et al. Endocytosis, intracellular sorting, and processing of exosomes by dendritic cells. Blood 2004, 104, 3257–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, E.; Guérin, C.; Hogg, N.; Amigorena, S.; Théry, C. CD8+ Dendritic Cells Use LFA-1 to Capture MHC-Peptide Complexes from Exosomes In Vivo. J. Immunol. 2007, 179, 1489–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleys, E.R.; Halleran, J.L.; McWhorter, E.; Hergenreder, J.; Enriquez, V.A.; da Silveira, J.C.; Bruemmer, J.E.; Winger, Q.A.; Bouma, G.J. Identification of microRNAs in exosomes isolated from serum and umbilical cord blood, as well as placentomes of gestational day 90 pregnant sheep. Mol. Reprod. Dev. 2014, 81, 983–993. [Google Scholar] [CrossRef]
- Street, J.M.; Barran, P.E.; Mackay, C.L.; Weidt, S.; Balmforth, C.; Walsh, T.S.; Chalmers, R.T.A.; Webb, D.J.; Dear, J.W. Identification and proteomic profiling of exosomes in human cerebrospinal fluid. J. Transl. Med. 2012, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prunotto, M.; Farina, A.; Lane, L.; Pernin, A.; Schifferli, J.; Hochstrasser, D.F.; Lescuyer, P.; Moll, S. Proteomic analysis of podocyte exosome-enriched fraction from normal human urine. J. Proteom. 2013, 82, 193–229. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Buzás, E.I.; Bemis, L.T.; Bora, A.; Lässer, C.; Lötvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J. Extracell Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- De Ávila, A.C.F.C.M.; Bridi, A.; Andrade, G.M.; del Collado, M.; Sangalli, J.R.; Nociti, R.P.; da Silva Junior, W.A.; Bastien, A.; Robert, C.; Meirelles, F.V.; et al. Estrous cycle impacts miRNA content in extracellular vesicles that modulate bovine cumulus cell transcripts during in vitro maturation. Biol. Reprod. 2019. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.M.; Bomfim, M.M.; del Collado, M.; Meirelles, F.V.; Perecin, F.; da Silveira, J.C. Oxygen tension modulates extracellular vesicles and its miRNA contents in bovine embryo culture medium. Mol. Reprod. Dev. 2019, 86, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Qing, S.; Liu, R.; Qin, H.; Wang, W.; Qiao, F.; Ge, H.; Liu, J.; Zhang, Y.; Cui, W.; et al. Effects of embryo-derived exosomes on the development of bovine cloned embryos. PLoS ONE 2017, 12, e0174535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange-Consiglio, A.; Perrini, C.; Albini, G.; Modina, S.; Lodde, V.; Orsini, E.; Esposti, P.; Cremonesi, F. Oviductal microvesicles and their effect on in vitro maturation of canine oocytes. Reproduction 2017, 154, 167. [Google Scholar] [CrossRef]
- Saadeldin, I.M.; Kim, S.J.; Choi, Y.B.; Lee, B.C. Improvement of Cloned Embryos Development by Co-Culturing with Parthenotes: A Possible Role of Exosomes/Microvesicles for Embryos Paracrine Communication. Cell. Reprogramming 2014, 16, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Almiñana, C.; Corbin, E.; Tsikis, G.; Alcântara-Neto, A.S.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct extracellular vesicles protein content and their role during oviduct–embryo cross-talk. Reproduction 2017, 154, 253. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramírez, M.Á.; Yáñez-Mó, M.; Rizos, D. Effect of bovine oviductal extracellular vesicles on embryo development and quality in vitro. Reproduction 2017, 153, 461. [Google Scholar] [CrossRef] [Green Version]
- Qu, P.; Zhao, Y.; Wang, R.; Zhang, Y.; Li, L.; Fan, J.; Liu, E. Extracellular vesicles derived from donor oviduct fluid improved birth rates after embryo transfer in mice. Reprod. Fertil. Dev. 2019, 31, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-González, I.; Xu, J.; Wang, X.; Burghardt, R.C.; Dunlap, K.A.; Bazer, F.W. Exosomes, endogenous retroviruses and toll-like receptors: Pregnancy recognition in ewes. Reproduction 2015, 149, 281–291. [Google Scholar] [CrossRef]
- Kusama, K.; Nakamura, K.; Bai, R.; Nagaoka, K.; Sakurai, T.; Imakawa, K. Intrauterine exosomes are required for bovine conceptus implantation. Biochem. Biophys. Res. Commun. 2018, 495, 1370–1375. [Google Scholar] [CrossRef]
- Burns, G.W.; Brooks, K.E.; O’Neil, E.V.; Hagen, D.E.; Behura, S.K.; Spencer, T.E. Progesterone effects on extracellular vesicles in the sheep uterus†. Biol. Reprod. 2018, 98, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Bidarimath, M.; Khalaj, K.; Kridli, R.T.; Kan, F.W.K.; Koti, M.; Tayade, C. Extracellular vesicle mediated intercellular communication at the porcine maternal-fetal interface: A new paradigm for conceptus-endometrial cross-talk. Sci. Rep. 2017, 7, 40476. [Google Scholar] [CrossRef]
- Brower, L.K.; Anderson, E. Cytological Events Associated with the Secretory Process in the Rabbit Oviduct1. Biol. Reprod. 1969, 1, 130–148. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.W.; Plante, C.; Allan King, W.; Hansen, P.J.; Betteridge, K.J.; Suarez, S.S. Fertilizing Capacity of Bovine Sperm may be Maintained by Binding to Oviductal Epithelial Cells1. Biol. Reprod. 1991, 44, 102–107. [Google Scholar] [CrossRef]
- Almiñana, C.; Bauersachs, S. Extracellular Vesicles in the Oviduct: Progress, Challenges and Implications for the Reproductive Success. Bioengineering 2019, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Della-Giustina, D.; Denny, M. Ectopic pregnancy. Emerg. Med. Clin. N. Am. 2003, 21, 565–584. [Google Scholar] [CrossRef]
- Murray, H.; Baakdah, H.; Bardell, T.; Tulandi, T. Diagnosis and treatment of ectopic pregnancy. CMAJ 2005, 173, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Pisarska, M.D.; Carson, S.A.; Buster, J.E. Ectopic pregnancy. Lancet 1998, 351, 1115–1120. [Google Scholar] [CrossRef]
- Brennan, D.F. Ectopic Pregnancy—Part I: Clinical and Laboratory Diagnosis. Acad. Emerg. Med. 1995, 2, 1081–1089. [Google Scholar] [CrossRef]
- ACOG Practice Bulletin No. 94: Medical management of ectopic pregnancy. Obstet. Gynecol. 2008, 111, 1479–1485.
- Zhao, Z.; Zhao, Q.; Warrick, J.; Lockwood, C.M.; Woodworth, A.; Moley, K.H.; Gronowski, A.M. Circulating MicroRNA miR-323-3p as a Biomarker of Ectopic Pregnancy. Clin. Chem. 2012, 58, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, D.; Barratt, C.L.R.; Björndahl, L.; de Jager, C.; Jequier, A.M.; Muller, C.H. What should it take to describe a substance or product as ‘sperm-safe’. Hum. Reprod. Update 2013, 19, i1–i45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazer, F.W.; Wu, G.; Johnson, G.A.; Kim, J.; Song, G. Uterine Histotroph and Conceptus Development: Select Nutrients and Secreted Phosphoprotein 1 Affect Mechanistic Target of Rapamycin Cell Signaling in Ewes1. Biol. Reprod. 2011, 85, 1094–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Bazer, F.W. Biology of progesterone action during pregnancy recognition and maintenance of pregnancy. Front. Biosci. 2002, 7, d1879–d1898. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.A.; Bazer, F.W.; Jaeger, L.A.; Ka, H.; Garlow, J.E.; Pfarrer, C.; Spencer, T.E.; Burghardt, R.C. Muc-1, Integrin, and Osteopontin Expression During the Implantation Cascade in Sheep. Biol. Reprod. 2001, 65, 820–828. [Google Scholar] [CrossRef] [Green Version]
- Ginther, O.J. INTERNAL REGULATION OF PHYSIOLOGICAL PROCESSES THROUGH LOCAL VENOARTERIAL PATHWAYS: A REVIEW. J. Anim. Sci. 1974, 39, 550–564. [Google Scholar] [CrossRef]
- Klohonatz, K.M.; Cameron, A.D.; Hergenreder, J.R.; da Silveira, J.C.; Belk, A.D.; Veeramachaneni, D.N.R.; Bouma, G.J.; Bruemmer, J.E. Circulating miRNAs as Potential Alternative Cell Signaling Associated with Maternal Recognition of Pregnancy in the Mare1. Biol. Reprod. 2016, 95, 124. [Google Scholar] [CrossRef]
- Burghardt, R.C.; Burghardt, J.R.; Taylor, J.D.; Reeder, A.T.; Nguen, B.T.; Spencer, T.E.; Bayless, K.J.; Johnson, G.A. Enhanced focal adhesion assembly reflects increased mechanosensation and mechanotransduction at maternal–conceptus interface and uterine wall during ovine pregnancy. Reproduction 2009, 137, 567–582. [Google Scholar] [CrossRef] [Green Version]
- Pohler, K.G.; Green, J.A.; Moley, L.A.; Gunewardena, S.; Hung, W.-T.; Payton, R.R.; Hong, X.; Christenson, L.K.; Geary, T.W.; Smith, M.F. Circulating microRNA as candidates for early embryonic viability in cattle. Mol. Reprod. Dev. 2017, 84, 731–743. [Google Scholar] [CrossRef]
- De Bem, T.H.C.; da Silveira, J.C.; Sampaio, R.V.; Sangalli, J.R.; Oliveira, M.L.F.; Ferreira, R.M.; Silva, L.A.; Perecin, F.; King, W.A.; Meirelles, F.V.; et al. Low levels of exosomal-miRNAs in maternal blood are associated with early pregnancy loss in cloned cattle. Sci. Rep. 2017, 7, 14319. [Google Scholar] [CrossRef]
- Markkandan, K.; Ahn, K.; Lee, D.J.; Kim, T.I.; Dang, C.; Hong, S.-E.; Yoon, H.-B.; Lim, H.-J.; Hong, C.P. Profiling and identification of pregnancy-associated circulating microRNAs in dairy cattle. Genes Genom. 2018, 40, 1111–1117. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Topic | Findings | Reference |
|---|---|---|
| EVs derived from ovarian follicles |
| Da Silveira et al., 2012 [24] |
| ||
| EVs derived from in vitro produced embryos |
| Mellisho et al., 2017 [20] |
| Saadeldin et al., 2014 [65] | |
| Qu et al., 2017 [63] | |
| Oviductal EVs |
| Lange-Consiglio et al., 2017 [64] |
| Al-Dossary et al., 2013 [5] | |
| Lopera-Vásquez et al., 2016; Almiñana et al., 2017; Lopera-Vasquez et al., 2017 [7,66,67] | |
| Qu et al., 2019 [68] | |
| Embryonic and uterine EVs |
| Burns et al., 2016 [21] |
| Ruiz-González et al., 2015 [69] | |
| Nakamura et al., 2016; Kusama et al., 2018 [22,70] | |
| Nakamura et al., 2016 [22] | |
| Burns et al., 2018 [71] | |
| ||
| Bidarimath et al., 2017 [72] |
| Species | Period of Pregnancy | Circulating Exosomal miRNAs in Pregnant Female | Pathways Modulated by miRNAs | Reference |
|---|---|---|---|---|
| Equine | Day 9 | Down-regulated: eca-miR-27a, eca-miR-29c, eca-miR-101, and eca-miR-486-5p | Extracellular matrix (ECM)–receptor interaction Proteoglycans in cancer TGF-beta Focal adhesion pathway | Klohonatz et al., 2016 [87] |
| Day 11 | Down-regulated: eca-miR-195 Up-regulated: eca-miR-767-5p | Regulating pluripotency of stem cells Fatty acid biosynthesis FoxO Focal adhesion pathway | ||
| Day 13 | Down-regulated: eca-miR-188-5p, eca-miR-653, eca-miR-874, and eca-miR-140-3p Up-regulated: eca-miR-30c and eca-miR-323-5p | Glutamatergic synapse Regulation of actin cytoskeleton Long-term potentiation Focal adhesion pathway | ||
| Bovine | Day 17 | Down-regulated: bta-let-7c, bta-mir-100, bta-mir-101-1, bta-mir-101-2, bta-mir-106b, bta-mir-125b-2, bta-mir-127, bta-mir-141, bta-mir-143, bta-mir-148a, bta-mir-15a, bta-mir-15b, bta-mir-16a, bta-mir-16b, bta-mir-1839, bta-mir-199a-1, bta-mir-199b, bta-mir-2415, bta-mir-25, bta-mir-331, bta-mir-339b, bta-mir-3596, bta-mir-3604-1, bta-mir-409b, bta-mir-451, bta-mir-652, bta-mir-99a | Cancer Connective tissue disorders Organismal injury and abnormalities Reproductive system disease Endocrine disorders | Pohler et al., 2017 [89] |
| Bovine | Day 21 | Highly abundant: bta-mir-15a, bta-mir-221, bta-mir-425-5p, bta-mir-101, bta-mir-93, bta-mir-106a, bta-mir-106b, bta-mir-22-5p, bta-mir-652, bta-mir-103, bta-mir-138, bta-mir-193b, bta-let-7f, bta-let-7g, bta-mir-15b, bta-let-7e, bta-let-7a-5p, bta-let-7d, bta-mir-660, bta-mir-29a, bta-mir-30d, bta-mir-497, bta-mir-148b, bta-mir-361, bta-mir-30a-5p, bta-mir-29c, bta-mir-29d-3p | Wnt TGF-beta Renal cell carcinoma Melanoma Colorectal cancer Glioma p53 Pancreatic cancer; Bladder cancer Dorsoventral axis formation | De Bem et al., 2017 [90] |
| Bovine | Day 30 | Down-regulated: bta-miR-193b, bta-miR-197, bta-miR-339a, bta-miR-326, bta-miR-484, bta-miR-486, bta-miR-423-3p, bta-miR-92a Up-regulated: bta-miR-146b, bta-miR-27b, bta-miR-26b, bta-miR-200a, bta-miR-450b, bta-miR-199c, bta-miR-194, bta-miR-6119-3p, bta-miR-7, bta-miR-199a-3p, bta-miR-574, bta-miR-215, bta-miR-148a, bta-let-7a-3p, bta-miR-21-5p, bta-miR-126-5p, bta-miR-148b, bta-miR-143, bta-miR-1246, bta-miR-192, bta-miR-98 | Membrane trafficking Chromosome and associated proteins Exosome G protein-coupled receptors Transcription factors Ubiquitin system Olfactory transduction Transporters Protein kinases Cytoskeleton proteins Cell adhesion molecules and their ligands Protein phosphatase and associated proteins Peptidases Pathway in cancer Messenger RNA Biogenesis CD molecules PI3K–Akt signaling pathway Human papillomavirus infection Mitochondrial biogenesis Spliceosome | Markkandan et al., 2018 [91] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bridi, A.; Perecin, F.; Silveira, J.C.d. Extracellular Vesicles Mediated Early Embryo–Maternal Interactions. Int. J. Mol. Sci. 2020, 21, 1163. https://doi.org/10.3390/ijms21031163
Bridi A, Perecin F, Silveira JCd. Extracellular Vesicles Mediated Early Embryo–Maternal Interactions. International Journal of Molecular Sciences. 2020; 21(3):1163. https://doi.org/10.3390/ijms21031163
Chicago/Turabian StyleBridi, Alessandra, Felipe Perecin, and Juliano Coelho da Silveira. 2020. "Extracellular Vesicles Mediated Early Embryo–Maternal Interactions" International Journal of Molecular Sciences 21, no. 3: 1163. https://doi.org/10.3390/ijms21031163
APA StyleBridi, A., Perecin, F., & Silveira, J. C. d. (2020). Extracellular Vesicles Mediated Early Embryo–Maternal Interactions. International Journal of Molecular Sciences, 21(3), 1163. https://doi.org/10.3390/ijms21031163