The Yin and Yang of Autosomal Recessive Primary Microcephaly Genes: Insights from Neurogenesis and Carcinogenesis


Abstract
:1. Introduction
2. MCPH Gene Deficiency Leads to Neurogenesis Defects
3. Some MCPH Genes Can Be Considered as Potential Cancer Biomarkers
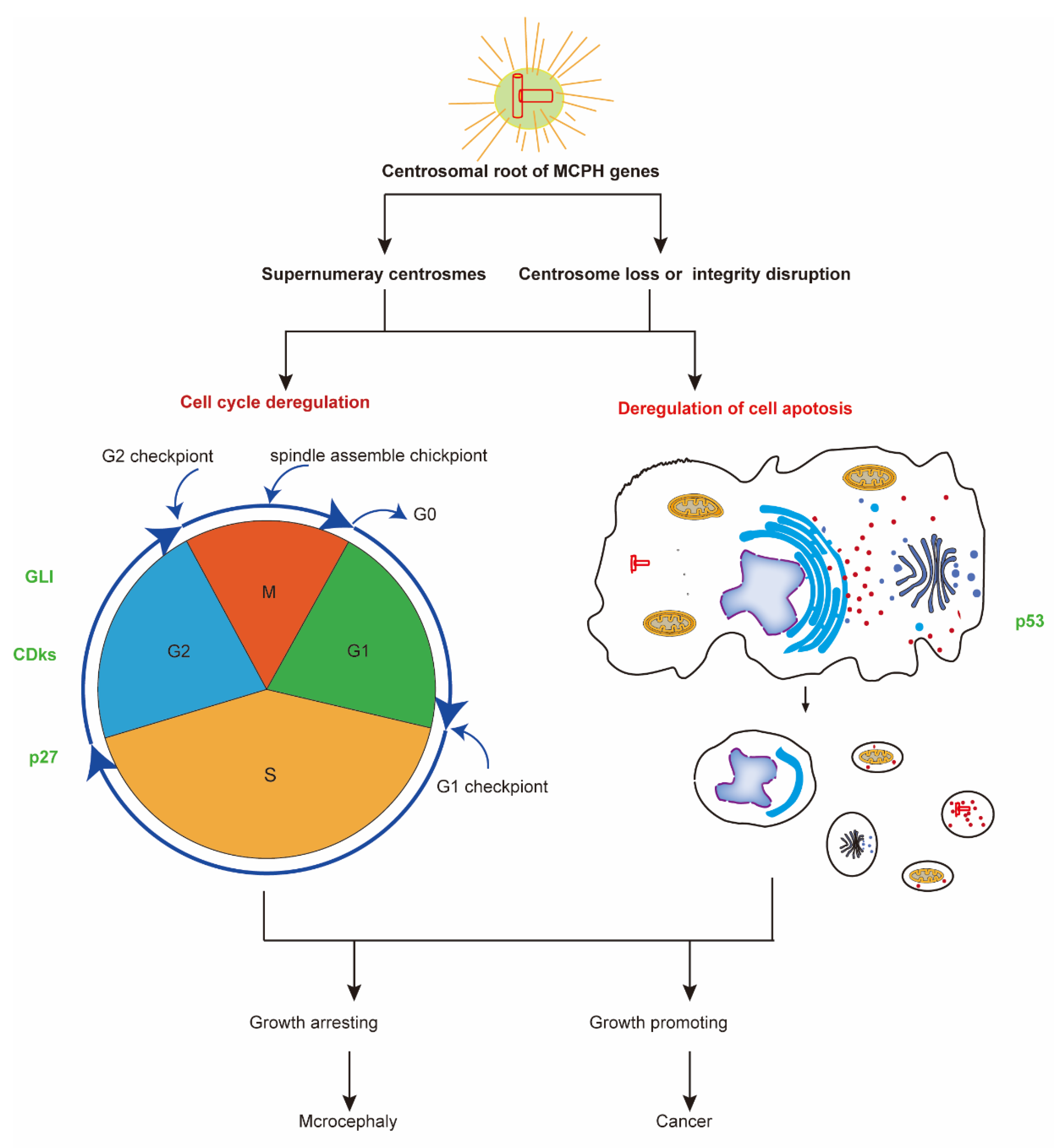
4. The Centrosomal Root of MCPH Genes
5. MCPH Gene Regulate Neurogenesis and Carcinogenesis through Regulation of Cell Cycle and Cell Division
6. MCPH Gene Regulate Neurogenesis and Carcinogenesis via Cell Apoptosis Regulation
7. The Molecular Regulators of MCPH Genes during Neurogenesis and Carcinogenesis
8. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MCPH | MCPH Autosomal recessive primary microcephaly |
| NSC | Neural stem cell |
| CSCs | Cancer stem cells |
| NPCs | Neural progenitor cells |
| KO | Knockout |
| BrdU | 5-bromo-2′-deoxyuridine |
| CCK-8 | Cell counting kit-8 |
| EdU | 5-ethynyl-2′-deoxyuridine |
| FACS | Fluorescent-activated cell sorting |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| TUNEL | Terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling |
| HC | Head circumference |
| GC | Gastric cancer |
| OC | Ovarian cancer |
| CRCs | Colorectal cancers |
| CC | Colon cancer |
| PCa | Prostate cancer |
| CDKs | Cyclin-dependent kinases |
| GLI | Glioblastoma protein |
| FOXM1 | Forkhead box M1 |
| TP53 | The p53 gene |
| pY15-Cdk1 | Tyr 15-phosphorylated Cdk1 |
| Cdk1b | Cyclin-dependent kinase inhibitor 1B |
| Shh | Sonic hedgehog |
| SUFU | Suppressor of fused |
References
- Taverna, E.; Gotz, M.; Huttner, W.B. The cell biology of neurogenesis: Toward an understanding of the development and evolution of the neocortex. Annu. Rev. Cell. Dev. Biol. 2014, 30, 465–502. [Google Scholar] [CrossRef] [PubMed]
- Land, H.; Parada, L.F.; Weinberg, R.A. Cellular oncogenes and multistep carcinogenesis. Science 1983, 222, 771–778. [Google Scholar] [CrossRef]
- Toh, T.B.; Lim, J.J.; Chow, E.K. Epigenetics in cancer stem cells. Mol. Cancer 2017, 16, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, J.M.; Jordan, C.T. The increasing complexity of the cancer stem cell paradigm. Science 2009, 324, 1670–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, D.; Bae, B.I.; Walsh, C.A. The Genetics of Primary Microcephaly. Annu. Rev. Genom. Hum. Genet. 2018, 19, 177–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, B.; Christian, K.M.; He, C.; Jin, P.; Ming, G.L.; Song, H. Epigenetic mechanisms in neurogenesis. Nat. Rev. Neurosci. 2016, 17, 537–549. [Google Scholar] [CrossRef]
- Baylin, S.B. Stem cells, cancer, and epigenetics. In StemBook; Harvard Stem Cell Insititute: Cambridge, MA, USA, 2009. [Google Scholar] [CrossRef]
- Zarco, N.; Norton, E.; Quinones-Hinojosa, A.; Guerrero-Cazares, H. Overlapping migratory mechanisms between neural progenitor cells and brain tumor stem cells. Cell. Mol. Life Sci. 2019, 76, 3553–3570. [Google Scholar] [CrossRef]
- Pallavicini, G.; Berto, G.E.; Di Cunto, F. Precision Revisited: Targeting Microcephaly Kinases in Brain Tumors. Int. J. Mol. Sci. 2019, 20, 2098. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, S.; Ahmad, W.; Hassan, M.J. Autosomal Recessive Primary Microcephaly (MCPH): Clinical manifestations, genetic heterogeneity and mutation continuum. Orphanet J. Rare Dis. 2011, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Eyaid, W.; Mochida, G.H.; Al-Moayyad, F.; Bodell, A.; Woods, C.G.; Walsh, C.A. ASPM mutations identified in patients with primary microcephaly and seizures. J. Med. Genet. 2005, 42, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Faheem, M.; Naseer, M.I.; Rasool, M.; Chaudhary, A.G.; Kumosani, T.A.; Ilyas, A.M.; Pushparaj, P.; Ahmed, F.; Algahtani, H.A.; Al-Qahtani, M.H.; et al. Molecular genetics of human primary microcephaly: An overview. BMC Med. Genom. 2015, 8, S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.J.; Zhou, X.K.; Xu, D. Update on autosomal recessive primary microcephaly (MCPH)-associated proteins. Yi Chuan Hered. 2019, 41, 905–918. [Google Scholar]
- Manzini, M.C.; Walsh, C.A. What disorders of cortical development tell us about the cortex: One plus one does not always make two. Curr. Opin. Genet. Dev. 2011, 21, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Q.; Li, W.; Lai, X.; Chen, H.; Huang, L.; Kang, Z.; Li, K.; Ren, J.; Lin, X.; Zheng, H.; et al. TALEN-based generation of a cynomolgus monkey disease model for human microcephaly. Cell Res. 2016, 26, 1048–1061. [Google Scholar] [CrossRef] [PubMed]
- Gruber, R.; Zhou, Z.W.; Sukchev, M.; Joerss, T.; Frappart, P.O.; Wang, Z.Q. MCPH1 regulates the neuroprogenitor division mode by coupling the centrosomal cycle with mitotic entry through the Chk1-Cdc25 pathway. Nat. Cell Biol. 2011, 13, 1325–1334. [Google Scholar] [CrossRef]
- Jayaraman, D.; Kodani, A.; Gonzalez, D.M.; Mancias, J.D.; Mochida, G.H.; Vagnoni, C.; Johnson, J.; Krogan, N.; Harper, J.W.; Reiter, J.F.; et al. Microcephaly proteins Wdr62 and Aspm define a mother centriole complex regulating centriole biogenesis, apical complex, and cell fate. Neuron 2016, 92, 813–828. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zhang, F.; Wang, Y.; Sun, Y.; Xu, Z. Microcephaly-associated protein WDR62 regulates neurogenesis through JNK1 in the developing neocortex. Cell Rep. 2014, 6, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Florio, M.; Huttner, W.B. Neural progenitors, neurogenesis and the evolution of the neocortex. Development 2014, 141, 2182–2194. [Google Scholar] [CrossRef] [Green Version]
- Fish, J.L.; Kosodo, Y.; Enard, W.; Paabo, S.; Huttner, W.B. Aspm specifically maintains symmetric proliferative divisions of neuroepithelial cells. Proc. Natl. Acad. Sci. USA 2006, 103, 10438–10443. [Google Scholar] [CrossRef] [Green Version]
- Buchman, J.J.; Durak, O.; Tsai, L.H. ASPM regulates Wnt signaling pathway activity in the developing brain. Genes Dev. 2011, 25, 1909–1914. [Google Scholar] [CrossRef] [Green Version]
- Fujikura, K.; Setsu, T.; Tanigaki, K.; Abe, T.; Kiyonari, H.; Terashima, T.; Sakisaka, T. Kif14 mutation causes severe brain malformation and hypomyelination. PLoS ONE 2013, 8, e53490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragich, J.M.; Kuwajima, T.; Hirose-Ikeda, M.; Yoon, M.S.; Eenjes, E.; Bosco, J.R.; Fox, L.M.; Lystad, A.H.; Oo, T.F.; Yarygina, O.; et al. Autophagy linked FYVE (Alfy/WDFY3) is required for establishing neuronal connectivity in the mammalian brain. eLife 2016, 5, e14810. [Google Scholar]
- Frantzi, M.; Zoidakis, J.; Papadopoulos, T.; Zurbig, P.; Katafigiotis, I.; Stravodimos, K.; Lazaris, A.; Giannopoulou, I.; Ploumidis, A.; Mischak, H.; et al. IMAC fractionation in combination with LC-MS reveals H2B and NIF-1 peptides as potential bladder cancer biomarkers. J. Proteome Res. 2013, 12, 3969–3979. [Google Scholar] [CrossRef]
- Shan, L.; Zhao, M.; Lu, Y.; Ning, H.; Yang, S.; Song, Y.; Chai, W.; Shi, X. CENPE promotes lung adenocarcinoma proliferation and is directly regulated by FOXM1. Int. J. Oncol. 2019, 55, 257–266. [Google Scholar] [PubMed]
- Wu, Z.; Zhu, X.; Xu, W.; Zhang, Y.; Chen, L.; Qiu, F.; Zhang, B.; Wu, L.; Peng, Z.; Tang, H. Up-regulation of CIT promotes the growth of colon cancer cells. Oncotarget 2017, 8, 71954–71964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, B.; Du, J.; Song, D.F.; Ren, J.C.; Feng, Y. Dysregulation of NCAPG, KNL1, miR-148a-3p, miR-193b-3p, and miR-1179 may contribute to the progression of gastric cancer. Biol. Res. 2018, 51, 44. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.C.; Xiao, Y.; Yan, W.G.; Ji, Z.G.; Zheng, G.Y. The human oncogene SCL/TAL1 interrupting locus (STIL) promotes tumor growth through MAPK/ERK, PI3K/Akt and AMPK pathways in prostate cancer. Gene 2019, 686, 220–227. [Google Scholar] [CrossRef]
- Ganapathi Sankaran, D.; Stemm-Wolf, A.J.; Pearson, C.G. CEP135 isoform dysregulation promotes centrosome amplification in breast cancer cells. Mol. Biol. Cell 2019, 30, 1230–1244. [Google Scholar] [CrossRef]
- Mi, Y.; Yu, M.; Zhang, L.; Sun, C.; Wei, B.; Ding, W.; Zhu, Y.; Tang, J.; Xia, G.; Zhu, L. COPB2 is upregulated in prostate cancer and regulates PC-3 cell proliferation, Cell Cycle, and Apoptosis. Arch. Med. Res. 2016, 47, 411–418. [Google Scholar] [CrossRef]
- Yin, L.; Jiang, L.P.; Shen, Q.S.; Xiong, Q.X.; Zhuo, X.; Zhang, L.L.; Yu, H.J.; Guo, X.; Luo, Y.; Dong, J.; et al. NCAPH plays important roles in human colon cancer. Cell Death Dis. 2017, 8, e2680. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, Y.; Dou, Z.; Jiang, H.; Wang, Y.; Gao, X.; Xin, X. Knockdown of STIL suppresses the progression of gastric cancer by down-regulating the IGF-1/PI3K/AKT pathway. J. Cell. Mol. Med. 2019, 23. [Google Scholar] [CrossRef]
- Pallavicini, G.; Sgro, F.; Garello, F.; Falcone, M.; Bitonto, V.; Berto, G.E.; Bianchi, F.T.; Gai, M.; Chiotto, A.M.A.; Filippi, M.; et al. Inactivation of citron kinase inhibits medulloblastoma progression by inducing apoptosis and cell senescence. Cancer Res. 2018, 78, 4599–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinmura, K.; Kato, H.; Kawanishi, Y.; Igarashi, H.; Inoue, Y.; Yoshimura, K.; Nakamura, S.; Fujita, H.; Funai, K.; Tanahashi, M.; et al. WDR62 overexpression is associated with a poor prognosis in patients with lung adenocarcinoma. Mol. Carcinog. 2017, 56, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, Q.; Luh, F.; Jin, B.; Liu, X. Overexpression of the ASPM gene is associated with aggressiveness and poor outcome in bladder cancer. Oncol. Lett. 2019, 17, 1865–1876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, Y.; Liang, P.; Zhang, Z.; Guo, X.; Xia, L.; Zhao, Y.; Shu, X.S.; Sun, S.; Ying, Y.; et al. Overexpression of a novel candidate oncogene KIF14 correlates with tumor progression and poor prognosis in prostate cancer. Oncotarget 2017, 8, 45459–45469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Liu, Y.; Feng, W.; Lei, L.; Du, Y.; Wu, J.; Wang, S. NUP37, a positive regulator of YAP/TEAD signaling, promotes the progression of hepatocellular carcinoma. Oncotarget 2017, 8, 98004–98013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, F.; Hu, J.; Xu, Z.; Tan, J.; Tang, H. Overexpression of NCAPH is upregulated and predicts a poor prognosis in prostate cancer. Oncol. Lett. 2019, 17, 5768–5776. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, F.; Zhang, C.; Ren, M.; Kuang, M.; Xiao, T.; Di, X.; Feng, L.; Fu, L.; Cheng, S. Non-SMC condensin I complex subunit D2 is a prognostic factor in triple-negative breast cancer for the ability to promote cell cycle and enhance invasion. Am. J. Pathol. 2020, 190, 37–47. [Google Scholar] [CrossRef]
- Mai, L.; Yi, F.; Gou, X.; Zhang, J.; Wang, C.; Liu, G.; Bu, Y.; Yuan, C.; Deng, L.; Song, F. The overexpression of MCPH1 inhibits cell growth through regulating cell cycle-related proteins and activating cytochrome c-caspase 3 signaling in cervical cancer. Mol. Cell. Biochem. 2014, 392, 95–107. [Google Scholar] [CrossRef]
- Spinola, M.; Falvella, F.S.; Colombo, F.; Sullivan, J.P.; Shames, D.S.; Girard, L.; Spessotto, P.; Minna, J.D.; Dragani, T.A. MFSD2A is a novel lung tumor suppressor gene modulating cell cycle and matrix attachment. Mol. Cancer 2010, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Liu, W.; Liu, X.; Ai, Q.; Yu, J. Overexpression of MCPH1 inhibits the migration and invasion of lung cancer cells. Oncotargets Ther. 2018, 11, 3111–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T.; Akutsu, S.N.; Fukumitsu, A.; Morino, H.; Masatsuna, Y.; Hosoba, K.; Kawakami, H.; Yamamoto, T.; Shimizu, K.; Ohashi, H.; et al. PLK1-mediated phosphorylation of WDR62/MCPH2 ensures proper mitotic spindle orientation. Hum. Mol. Genet. 2017, 26, 4429–4440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, S.; Tao, Y.; Huang, J.; Zhang, S.; Shen, L.; Yang, H.; Pei, H.; Zhong, M.; Zhang, G.; Liu, T.; et al. WD40 repeat-containing 62 overexpression as a novel indicator of poor prognosis for human gastric cancer. Eur. J. Cancer 2013, 49, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Alshawaf, A.J.; Antonic, A.; Skafidas, E.; Ng, D.C.; Dottori, M. WDR62 regulates early neural and glial progenitor specification of human pluripotent stem cells. Stem Cells Int. 2017, 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tian, Y.; Yu, J.J.; He, J.; Luo, J.; Zhang, S.; Tang, C.E.; Tao, Y.M. Overexpression of WDR62 is associated with centrosome amplification in human ovarian cancer. J. Ovarian Res. 2013, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.F.; Zhang, Y.; Wilde, J.; Hansen, K.C.; Lai, F.; Niswander, L. Microcephaly disease gene Wdr62 regulates mitotic progression of embryonic neural stem cells and brain size. Nat. Commun. 2014, 5, 3885. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Yao, M.; Wang, Y.; Yuan, L.; Hoeck, J.D.; Yu, J.; Liu, L.; Yeap, Y.Y.C.; Zhang, W.; Zhang, F.; et al. MEKK3 coordinates with FBW7 to regulate WDR62 stability and neurogenesis. PLoS Biol. 2018, 16, e2006613. [Google Scholar] [CrossRef]
- Lizarraga, S.B.; Margossian, S.P.; Harris, M.H.; Campagna, D.R.; Han, A.P.; Blevins, S.; Mudbhary, R.; Barker, J.E.; Walsh, C.A.; Fleming, M.D. Cdk5rap2 regulates centrosome function and chromosome segregation in neuronal progenitors. Development 2010, 137, 1907–1917. [Google Scholar] [CrossRef]
- Walz, C.; Curtis, C.; Schnittger, S.; Schultheis, B.; Metzgeroth, G.; Schoch, C.; Lengfelder, E.; Erben, P.; Muller, M.C.; Haferlach, T.; et al. Transient response to imatinib in a chronic eosinophilic leukemia associated with ins(9;4)(q33;q12q25) and a CDK5RAP2-PDGFRA fusion gene. Genes Chromosomes Cancer 2006, 45, 950–956. [Google Scholar] [CrossRef]
- Buchman, J.J.; Tseng, H.C.; Zhou, Y.; Frank, C.L.; Xie, Z.; Tsai, L.H. Cdk5rap2 interacts with pericentrin to maintain the neural progenitor pool in the developing neocortex. Neuron 2010, 66, 386–402. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Zhao, Y.; Liu, Y.; Cai, B.; Dong, N.; Li, B. Effect of KNL1 on the proliferation and apoptosis of colorectal cancer cells. Technol. Cancer Res. Treat. 2019, 18. [Google Scholar] [CrossRef] [Green Version]
- Ogi, H.; Nitta, N.; Tando, S.; Fujimori, A.; Aoki, I.; Fushiki, S.; Itoh, K. Longitudinal diffusion tensor imaging revealed nerve fiber alterations in Aspm mutated microcephaly model mice. Neuroscience 2018, 371, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, A.; Itoh, K.; Goto, S.; Hirakawa, H.; Wang, B.; Kokubo, T.; Kito, S.; Tsukamoto, S.; Fushiki, S. Disruption of Aspm causes microcephaly with abnormal neuronal differentiation. Brain Dev. 2014, 36, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Liao, W.Y.; Chan, T.S.; Chen, W.Y.; Lee, C.T.; Shan, Y.S.; Huang, P.J.; Hou, Y.C.; Li, C.R.; Tsai, K.K. The differential distributions of ASPM isoforms and their roles in Wnt signaling, cell cycle progression, and pancreatic cancer prognosis. J. Pathol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Wu, Q.; Sun, L.; Pan, N.C.; Wang, X. Cenpj regulates cilia disassembly and neurogenesis in the developing mouse cortex. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 1994–2010. [Google Scholar] [CrossRef]
- Garcez, P.P.; Diaz-Alonso, J.; Crespo-Enriquez, I.; Castro, D.; Bell, D.; Guillemot, F. Cenpj/CPAP regulates progenitor divisions and neuronal migration in the cerebral cortex downstream of Ascl1. Nat. Commun. 2015, 6, 6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottee, M.A.; Muschalik, N.; Wong, Y.L.; Johnson, C.M.; Johnson, S.; Andreeva, A.; Oegema, K.; Lea, S.M.; Raff, J.W.; van Breugel, M. Crystal structures of the CPAP/STIL complex reveal its role in centriole assembly and human microcephaly. eLife 2013, 2, e01071. [Google Scholar] [CrossRef]
- Arquint, C.; Nigg, E.A. The PLK4-STIL-SAS-6 module at the core of centriole duplication. Biochem. Soc. Trans. 2016, 44, 1253–1263. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Wu, C.T.; Tang, C.C.; Lin, Y.N.; Wang, W.J.; Tang, T.K. Human microcephaly protein RTTN interacts with STIL and is required to build full-length centrioles. Nat. Commun. 2017, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, D.; Mani, S.; Passemard, S.; Gressens, P.; El Ghouzzi, V. STIL balancing primary microcephaly and cancer. Cell Death Dis. 2018, 9, 65. [Google Scholar] [CrossRef]
- Rabinowicz, N.; Mangala, L.S.; Brown, K.R.; Checa-Rodriguez, C.; Castiel, A.; Moskovich, O.; Zarfati, G.; Trakhtenbrot, L.; Levy-Barda, A.; Jiang, D.; et al. Targeting the centriolar replication factor STIL synergizes with DNA damaging agents for treatment of ovarian cancer. Oncotarget 2017, 8, 27380–27392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.S.; Baig, S.M.; Neumann, S.; Nurnberg, G.; Farooq, M.; Ahmad, I.; Alef, T.; Hennies, H.C.; Technau, M.; Altmuller, J.; et al. A truncating mutation of CEP135 causes primary microcephaly and disturbed centrosomal function. Am. J. Hum. Genet. 2012, 90, 871–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Ramdas Nair, A.; Cabernard, C. The centriolar protein Bld10/Cep135 is required to establish centrosome asymmetry in Drosophila neuroblasts. Curr. Biol. CB 2014, 24, 1548–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.J.; Baltus, A.E.; Mathew, R.S.; Murphy, E.A.; Evrony, G.D.; Gonzalez, D.M.; Wang, E.P.; Marshall-Walker, C.A.; Barry, B.J.; Murn, J.; et al. Microcephaly gene links trithorax and REST/NRSF to control neural stem cell proliferation and differentiation. Cell 2012, 151, 1097–1112. [Google Scholar] [CrossRef] [Green Version]
- Hasenpusch-Theil, K.; West, S.; Kelman, A.; Kozic, Z.; Horrocks, S.; McMahon, A.P.; Price, D.J.; Mason, J.O.; Theil, T. Gli3 controls the onset of cortical neurogenesis by regulating the radial glial cell cycle through Cdk6 expression. Development 2018, 145, dev163147. [Google Scholar]
- Dall’Acqua, A.; Sonego, M.; Pellizzari, I.; Pellarin, I.; Canzonieri, V.; D’Andrea, S.; Benevol, S.; Sorio, R.; Giorda, G.; Califano, D.; et al. CDK6 protects epithelial ovarian cancer from platinum-induced death via FOXO3 regulation. EMBO Mol. Med. 2017, 9, 1415–1433. [Google Scholar] [CrossRef]
- Caron, N.; Genin, E.C.; Marlier, Q.; Verteneuil, S.; Beukelaers, P.; Morel, L.; Hu, M.G.; Hinds, P.W.; Nguyen, L.; Vandenbosch, R.; et al. Proliferation of hippocampal progenitors relies on p27-dependent regulation of Cdk6 kinase activity. Cell. Mol. Life Sci. 2018, 75, 3817–3827. [Google Scholar] [CrossRef]
- Bellail, A.C.; Olson, J.J.; Hao, C. SUMO1 modification stabilizes CDK6 protein and drives the cell cycle and glioblastoma progression. Nat. Commun. 2014, 5, 4234. [Google Scholar] [CrossRef]
- Luyckx, E.; Van Leuven, W.; Andre, D.; Quarta, A.; Reekmans, K.; Fransen, E.; Moens, L.; Hankeln, T.; Ponsaerts, P.; Dewilde, S. Loss of neuroglobin expression alters Cdkn1a/Cdk6-expression resulting in increased proliferation of neural stem cells. Stem Cells Dev. 2018, 27, 378–390. [Google Scholar] [CrossRef]
- Kollmann, K.; Heller, G.; Schneckenleithner, C.; Warsch, W.; Scheicher, R.; Ott, R.G.; Schafer, M.; Fajmann, S.; Schlederer, M.; Schiefer, A.I.; et al. A kinase-independent function of CDK6 links the cell cycle to tumor angiogenesis. Cancer Cell 2013, 24, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Jena, N.; Sheng, J.; Hu, J.K.; Li, W.; Zhou, W.; Lee, G.; Tsichlis, N.; Pathak, A.; Brown, N.; Deshpande, A.; et al. CDK6-mediated repression of CD25 is required for induction and maintenance of Notch1-induced T-cell acute lymphoblastic leukemia. Leukemia 2016, 30, 1033–1043. [Google Scholar] [CrossRef] [Green Version]
- Shinmura, K.; Kato, H.; Kawanishi, Y.; Nagura, K.; Kamo, T.; Okubo, Y.; Inoue, Y.; Kurabe, N.; Du, C.; Iwaizumi, M.; et al. SASS6 overexpression is associated with mitotic chromosomal abnormalities and a poor prognosis in patients with colorectal cancer. Oncol. Rep. 2015, 34, 727–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Zvi, A.; Lacoste, B.; Kur, E.; Andreone, B.J.; Mayshar, Y.; Yan, H.; Gu, C. Mfsd2a is critical for the formation and function of the blood-brain barrier. Nature 2014, 509, 507–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Jaiswal, M.; Charng, W.L.; Gambin, T.; Karaca, E.; Mirzaa, G.; Wiszniewski, W.; Sandoval, H.; Haelterman, N.A.; Xiong, B.; et al. A drosophila genetic resource of mutants to study mechanisms underlying human genetic diseases. Cell 2014, 159, 200–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgro, F.; Bianchi, F.T.; Falcone, M.; Pallavicini, G.; Gai, M.; Chiotto, A.M.; Berto, G.E.; Turco, E.; Chang, Y.J.; Huttner, W.B.; et al. Tissue-specific control of midbody microtubule stability by Citron kinase through modulation of TUBB3 phosphorylation. Cell Death Differ. 2016, 23, 801–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.T.; Tocco, C.; Pallavicini, G.; Liu, Y.; Verni, F.; Merigliano, C.; Bonaccorsi, S.; El-Assawy, N.; Priano, L.; Gai, M.; et al. Citron kinase deficiency leads to chromosomal instability and TP53-sensitive microcephaly. Cell Rep. 2017, 18, 1674–1686. [Google Scholar] [CrossRef]
- Li, W.; Ma, S.; Bai, X.; Pan, W.; Ai, L.; Tan, W. Long noncoding RNA WDFY3-AS2 suppresses tumor progression by acting as a competing endogenous RNA of microRNA-18a in ovarian cancer. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef]
- Le Duc, D.; Giulivi, C.; Hiatt, S.M.; Napoli, E.; Panoutsopoulos, A.; Harlan De Crescenzo, A.; Kotzaeridou, U.; Syrbe, S.; Anagnostou, E.; Azage, M.; et al. Pathogenic WDFY3 variants cause neurodevelopmental disorders and opposing effects on brain size. Brain A J. Neurol. 2019, 142, 2617–2630. [Google Scholar] [CrossRef]
- Napoli, E.; Song, G.; Panoutsopoulos, A.; Halmai, J.; Levenson, R.; Zarbalis, K.S.; Giulivi, C.; Riyadh, M.A.; Kaushik, G. Beyond autophagy: A novel role for autism-linked Wdfy3 in brain mitophagy. Sci. Rep. 2018, 8, 11348. [Google Scholar] [CrossRef] [Green Version]
- DiStasio, A.; Driver, A.; Sund, K.; Donlin, M.; Muraleedharan, R.M.; Pooya, S.; Kline-Fath, B.; Kaufman, K.M.; Prows, C.A.; Schorry, E.; et al. Copb2 is essential for embryogenesis and hypomorphic mutations cause human microcephaly. Hum. Mol. Genet. 2017, 26, 4836–4848. [Google Scholar] [CrossRef] [Green Version]
- Mi, Y.; Sun, C.; Wei, B.; Sun, F.; Guo, Y.; Hu, Q.; Ding, W.; Zhu, L.; Xia, G. Coatomer subunit beta 2 (COPB2), identified by label-free quantitative proteomics, regulates cell proliferation and apoptosis in human prostate carcinoma cells. Biochem. Biophys. Res. Commun. 2018, 495, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Wang, J.; Li, W.; Fan, W.; Wang, L.; Mao, Y.; Yang, S.; Liu, S.; Xu, J.; Lv, Z.; et al. COPB2 promotes cell proliferation and tumorigenesis through up-regulating YAP1 expression in lung adenocarcinoma cells. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 103, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.S.; Liu, C.H.; Liu, Z.; Zhu, C.L.; Huang, Q. Downregulation of COPB2 by RNAi inhibits growth of human cholangiocellular carcinoma cells. Eur. Rev. Med Pharmacol. Sci. 2018, 22, 985–992. [Google Scholar]
- Wang, Y.; Chai, Z.; Wang, M.; Jin, Y.; Yang, A.; Li, M. COPB2 suppresses cell proliferation and induces cell cycle arrest in human colon cancer by regulating cell cycle-related proteins. Exp. Ther. Med. 2018, 15, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Li, H.; Zhang, X.; Wang, J.; Qiang, Y.; Ye, X.; Li, Q.; Guan, Q.; Zhou, Y. Silencing of COPB2 inhibits the proliferation of gastric cancer cells and induces apoptosis via suppression of the RTK signaling pathway. Int. J. Oncol. 2019, 54, 1195–1208. [Google Scholar] [CrossRef]
- Theriault, B.L.; Basavarajappa, H.D.; Lim, H.; Pajovic, S.; Gallie, B.L.; Corson, T.W. Transcriptional and epigenetic regulation of KIF14 overexpression in ovarian cancer. PLoS ONE 2014, 9, e91540. [Google Scholar] [CrossRef]
- Xu, H.; Choe, C.; Shin, S.H.; Park, S.W.; Kim, H.S.; Jung, S.H.; Yim, S.H.; Kim, T.M.; Chung, Y.J. Silencing of KIF14 interferes with cell cycle progression and cytokinesis by blocking the p27(Kip1) ubiquitination pathway in hepatocellular carcinoma. Exp. Mol. Med. 2014, 46, e97. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.A.; Murray, J.E.; Carroll, P.; Leitch, A.; Mackenzie, K.J.; Halachev, M.; Fetit, A.E.; Keith, C.; Bicknell, L.S.; Fluteau, A.; et al. Mutations in genes encoding condensin complex proteins cause microcephaly through decatenation failure at mitosis. Genes Dev. 2016, 30, 2158–2172. [Google Scholar] [CrossRef] [Green Version]
- Perez, Y.; Bar-Yaacov, R.; Kadir, R.; Wormser, O.; Shelef, I.; Birk, O.S.; Flusser, H.; Birnbaum, R.Y. Mutations in the microtubule-associated protein MAP11 (C7orf43) cause microcephaly in humans and zebrafish. Brain 2019, 142, 574–585. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A.D.; Feldman, J.L. Microtubule-organizing centers: From the centrosome to non-centrosomal sites. Curr. Opin. Cell Biol. 2017, 44, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Marthiens, V.; Rujano, M.A.; Pennetier, C.; Tessier, S.; Paul-Gilloteaux, P.; Basto, R. Centrosome amplification causes microcephaly. Nat. Cell Biol. 2013, 15, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Nigg, E.A.; Raff, J.W. Centrioles, centrosomes, and cilia in health and disease. Cell 2009, 139, 663–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Gao, H.; Lin, S.Y.; Peng, G.; Huang, X.; Zhang, P.; Goss, J.A.; Brunicardi, F.C.; Multani, A.S.; Chang, S.; et al. BRIT1/MCPH1 is essential for mitotic and meiotic recombination DNA repair and maintaining genomic stability in mice. PLoS Genet 2010, 6, e1000826. [Google Scholar] [CrossRef] [Green Version]
- Barr, A.R.; Kilmartin, J.V.; Gergely, F. CDK5RAP2 functions in centrosome to spindle pole attachment and DNA damage response. J. Cell Biol. 2010, 189, 23–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letard, P.; Drunat, S.; Vial, Y.; Duerinckx, S.; Ernault, A.; Amram, D.; Arpin, S.; Bertoli, M.; Busa, T.; Ceulemans, B.; et al. Autosomal recessive primary microcephaly due to ASPM mutations: An update. Hum. Mutat. 2018, 39, 319–332. [Google Scholar] [CrossRef]
- Okamoto, N.; Kohmoto, T.; Naruto, T.; Masuda, K.; Imoto, I. Primary microcephaly caused by novel compound heterozygous mutations in ASPM. Hum. Genome Var. 2018, 5, 18015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, D.; Lv, S.; Wang, H.; Zhong, X.; Liu, B.; Wang, B.; Liao, J.; Li, J.; Pfeifer, G.P.; et al. CDK5RAP2 is required for spindle checkpoint function. Cell Cycle 2009, 8, 1206–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Chang, C.W.; Hsu, W.B.; Tang, C.J.; Lin, Y.N.; Chou, E.J.; Wu, C.T.; Tang, T.K. Human microcephaly protein CEP135 binds to hSAS-6 and CPAP, and is required for centriole assembly. EMBO J. 2013, 32, 1141–1154. [Google Scholar] [CrossRef] [Green Version]
- Cizmecioglu, O.; Arnold, M.; Bahtz, R.; Settele, F.; Ehret, L.; Haselmann-Weiss, U.; Antony, C.; Hoffmann, I. Cep152 acts as a scaffold for recruitment of Plk4 and CPAP to the centrosome. J. Cell Biol. 2010, 191, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.S.; Baig, S.M.; Neumann, S.; Peche, V.S.; Szczepanski, S.; Nurnberg, G.; Tariq, M.; Jameel, M.; Khan, T.N.; Fatima, A.; et al. CDK6 associates with the centrosome during mitosis and is mutated in a large Pakistani family with primary microcephaly. Hum. Mol. Genet. 2013, 22, 5199–5214. [Google Scholar] [CrossRef]
- Strnad, P.; Leidel, S.; Vinogradova, T.; Euteneuer, U.; Khodjakov, A.; Gonczy, P. Regulated HsSAS-6 levels ensure formation of a single procentriole per centriole during the centrosome duplication cycle. Dev. Cell 2007, 13, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.S.; Bakker, B.; Boeckx, B.; Moyett, J.; Lu, J.; Vitre, B.; Spierings, D.C.; Lansdorp, P.M.; Cleveland, D.W.; Lambrechts, D.; et al. Centrosome amplification is sufficient to promote spontaneous tumorigenesis in mammals. Dev. Cell 2017, 40, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arquint, C.; Nigg, E.A. STIL microcephaly mutations interfere with APC/C-mediated degradation and cause centriole amplification. Curr. Biol. 2014, 24, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megraw, T.L.; Sharkey, J.T.; Nowakowski, R.S. Cdk5rap2 exposes the centrosomal root of microcephaly syndromes. Trends Cell Biol. 2011, 21, 470–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sercin, O.; Larsimont, J.C.; Karambelas, A.E.; Marthiens, V.; Moers, V.; Boeckx, B.; Le Mercier, M.; Lambrechts, D.; Basto, R.; Blanpain, C. Transient PLK4 overexpression accelerates tumorigenesis in p53-deficient epidermis. Nat. Cell Biol. 2016, 18, 100–110. [Google Scholar] [CrossRef]
- Insolera, R.; Bazzi, H.; Shao, W.; Anderson, K.V.; Shi, S.H. Cortical neurogenesis in the absence of centrioles. Nat. Neurosci. 2014, 17, 1528–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Nagle, R.B.; Knudsen, B.S.; Cress, A.E.; Rogers, G.C. Centrosome loss results in an unstable genome and malignant prostate tumors. Oncogene 2020, 39, 399–413. [Google Scholar] [CrossRef]
- Doxsey, S.; Zimmerman, W.; Mikule, K. Centrosome control of the cell cycle. Trends Cell Biol. 2005, 15, 303–311. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, E.; Dai, H.; Hu, R.; Liang, Y.; Li, K.; Wang, G.; Peng, G.; Lin, S.Y. BRIT1 regulates p53 stability and functions as a tumor suppressor in breast cancer. Carcinogenesis 2013, 34, 2271–2280. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wo, D.; Ma, E.; Yan, H.; Peng, J.; Zhu, W.; Fang, Y.; Ren, D.N. Deletion of low-density lipoprotein-related receptor 5 inhibits liver Cancer cell proliferation via destabilizing Nucleoporin 37. Cell Commun. Signal. 2019, 17, 174. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q. The centrosome-centered cell-brain in apoptosis. Med. Hypotheses 2003, 61, 126–132. [Google Scholar] [CrossRef]
- Zhou, Z.W.; Tapias, A.; Bruhn, C.; Gruber, R.; Sukchev, M.; Wang, Z.Q. DNA damage response in microcephaly development of MCPH1 mouse model. DNA Repair 2013, 12, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.E.; Garcia, I.; Crowther, A.J.; Li, S.; Stewart, A.; Liu, H.; Lough, K.J.; O’Neill, S.; Veleta, K.; Oyarzabal, E.A.; et al. Aspm sustains postnatal cerebellar neurogenesis and medulloblastoma growth in mice. Development 2015, 142, 3921–3932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erez, A.; Castiel, A.; Trakhtenbrot, L.; Perelman, M.; Rosenthal, E.; Goldstein, I.; Stettner, N.; Harmelin, A.; Eldar-Finkelman, H.; Campaner, S.; et al. The SIL gene is essential for mitotic entry and survival of cancer cells. Cancer Res. 2007, 67, 4022–4027. [Google Scholar] [CrossRef] [Green Version]
- Alves, C.L.; Elias, D.; Lyng, M.; Bak, M.; Kirkegaard, T.; Lykkesfeldt, A.E.; Ditzel, H.J. High CDK6 protects cells from fulvestrant-mediated apoptosis and is a predictor of resistance to fulvestrant in estrogen receptor-positive metastatic breast cancer. Clin. Cancer Res. 2016, 22, 5514–5526. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.; Yim, E.K.; Dai, H.; Jackson, A.P.; Burgt, I.; Pan, M.R.; Hu, R.; Li, K.; Lin, S.Y. BRIT1/MCPH1 links chromatin remodelling to DNA damage response. Nat. Cell Biol. 2009, 11, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Bai, Y.; Li, Y.; Liu, X.; Tan, T.; Meng, S.; He, W.; Wu, X.; Dong, Z. Overexpression of MCPH1 inhibits uncontrolled cell growth by promoting cell apoptosis and arresting the cell cycle in S and G2/M phase in lung cancer cells. Oncol. Lett. 2016, 11, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Lim, N.R.; Shohayeb, B.; Zaytseva, O.; Mitchell, N.; Millard, S.S.; Ng, D.C.H.; Quinn, L.M. Glial-specific functions of microcephaly protein WDR62 and interaction with the mitotic kinase AURKA are essential for Drosophila brain growth. Stem Cell Rep. 2017, 9, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Rosales, J.L.; Rattner, J.B.; Lee, K.Y. The primary microcephaly 3 (MCPH3) interacting protein, p35 and its catalytic subunit, Cdk5, are centrosomal proteins. Cell Cycle 2010, 9, 618–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukumaran, S.K.; Stumpf, M.; Salamon, S.; Ahmad, I.; Bhattacharya, K.; Fischer, S.; Muller, R.; Altmuller, J.; Budde, B.; Thiele, H.; et al. CDK5RAP2 interaction with components of the Hippo signaling pathway may play a role in primary microcephaly. Mol. Genet. Genom. 2017, 292, 365–383. [Google Scholar] [CrossRef] [Green Version]
- Genin, A.; Desir, J.; Lambert, N.; Biervliet, M.; Van Der Aa, N.; Pierquin, G.; Killian, A.; Tosi, M.; Urbina, M.; Lefort, A.; et al. Kinetochore KMN network gene CASC5 mutated in primary microcephaly. Hum. Mol. Genet 2012, 21, 5306–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urata, Y.N.; Takeshita, F.; Tanaka, H.; Ochiya, T.; Takimoto, M. Targeted Knockdown of the kinetochore protein D40/Knl-1 inhibits human cancer in a p53 status-independent manner. Sci. Rep. 2015, 5, 13676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.Y.; Hsu, C.C.; Wang, T.Y.; Li, C.R.; Hou, Y.C.; Chu, J.M.; Lee, C.T.; Liu, M.S.; Su, J.J.; Jian, K.Y.; et al. A gene expression signature of epithelial tubulogenesis and a role for ASPM in pancreatic tumor progression. Gastroenterology 2013, 145, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Pai, V.C.; Hsu, C.C.; Chan, T.S.; Liao, W.Y.; Chuu, C.P.; Chen, W.Y.; Li, C.R.; Lin, C.Y.; Huang, S.P.; Chen, L.T.; et al. ASPM promotes prostate cancer stemness and progression by augmenting Wnt-Dvl-3-beta-catenin signaling. Oncogene 2019, 38, 1340–1353. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.J.; Xu, C.; Ping, G.Q.; Song, G.X.; Zhang, W.M.; Wang, C.; Zhang, Z.H. Correlation analysis of PD-L1 expression and prognosis in triple-negative breast cancers. Zhonghua Bing Li Xue Za Zhi Chin. J. Pathol. 2017, 46, 822–826. [Google Scholar]
- Izraeli, S.; Lowe, L.A.; Bertness, V.L.; Good, D.J.; Dorward, D.W.; Kirsch, I.R.; Kuehn, M.R. The SIL gene is required for mouse embryonic axial development and left-right specification. Nature 1999, 399, 691–694. [Google Scholar] [CrossRef]
- Kasai, K.; Inaguma, S.; Yoneyama, A.; Yoshikawa, K.; Ikeda, H. SCL/TAL1 interrupting locus derepresses GLI1 from the negative control of suppressor-of-fused in pancreatic cancer cell. Cancer Res. 2008, 68, 7723–7729. [Google Scholar] [CrossRef] [Green Version]
- Raleigh, D.R.; Choksi, P.K.; Krup, A.L.; Mayer, W.; Santos, N.; Reiter, J.F. Hedgehog signaling drives medulloblastoma growth via CDK6. J. Clin. Investig. 2018, 128, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Turrion, M.J.; Rodriguez-Martin, I.; Lopez-Bellido, R.; Rodriguez, R.E. Whole-genome expression profile in zebrafish embryos after chronic exposure to morphine: Identification of new genes associated with neuronal function and mu opioid receptor expression. BMC Genom. 2014, 15, 874. [Google Scholar] [CrossRef] [Green Version]
- Singel, S.M.; Cornelius, C.; Zaganjor, E.; Batten, K.; Sarode, V.R.; Buckley, D.L.; Peng, Y.; John, G.B.; Li, H.C.; Sadeghi, N.; et al. KIF14 promotes AKT phosphorylation and contributes to chemoresistance in triple-negative breast cancer. Neoplasia 2014, 16, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Bellutti, F.; Tigan, A.S.; Nebenfuehr, S.; Dolezal, M.; Zojer, M.; Grausenburger, R.; Hartenberger, S.; Kollmann, S.; Doma, E.; Prchal-Murphy, M.; et al. CDK6 antagonizes p53-induced responses during tumorigenesis. Cancer Discov. 2018, 8, 884–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, M.; Kuriyama, R.; Trimborn, M.; Keifenheim, D.; Canuelo, A.; Sanchez, A.; Clarke, D.J.; Marchal, J.A. MCPH1, mutated in primary microcephaly, is required for efficient chromosome alignment during mitosis. Sci. Rep. 2017, 7, 13019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villavicencio, E.H.; Walterhouse, D.O.; Iannaccone, P.M. The sonic hedgehog-patched-gli pathway in human development and disease. Am. J. Hum. Genet. 2000, 67, 1047–1054. [Google Scholar] [CrossRef]
- Armas-Lopez, L.; Zuniga, J.; Arrieta, O.; Avila-Moreno, F. The Hedgehog-GLI pathway in embryonic development and cancer: Implications for pulmonary oncology therapy. Oncotarget 2017, 8, 60684–60703. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Li, C.; Qin, C.F.; Xu, Z. Update on the animal Models and underlying mechanisms for ZIKV-induced microcephaly. Annu. Rev. Virol. 2019, 6, 459–479. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, J.; Ye, Q.; Ma, F.; Zhu, Q.; Wu, Y.; Shan, C.; Xie, X.; Li, D.; Zhan, X.; et al. Treatment of human glioblastoma with a live attenuated Zika virus vaccine candidate. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Gorman, M.J.; McKenzie, L.D.; Chai, J.N.; Hubert, C.G.; Prager, B.C.; Fernandez, E.; Richner, J.M.; Zhang, R.; Shan, C.; et al. Zika virus has oncolytic activity against glioblastoma stem cells. J. Exp. Med. 2017, 214, 2843–2857. [Google Scholar] [CrossRef] [Green Version]
- Lubin, J.A.; Zhang, R.R.; Kuo, J.S. Zika virus has oncolytic activity against glioblastoma stem cells. Neurosurgery 2018, 82, E113–E114. [Google Scholar] [CrossRef] [Green Version]




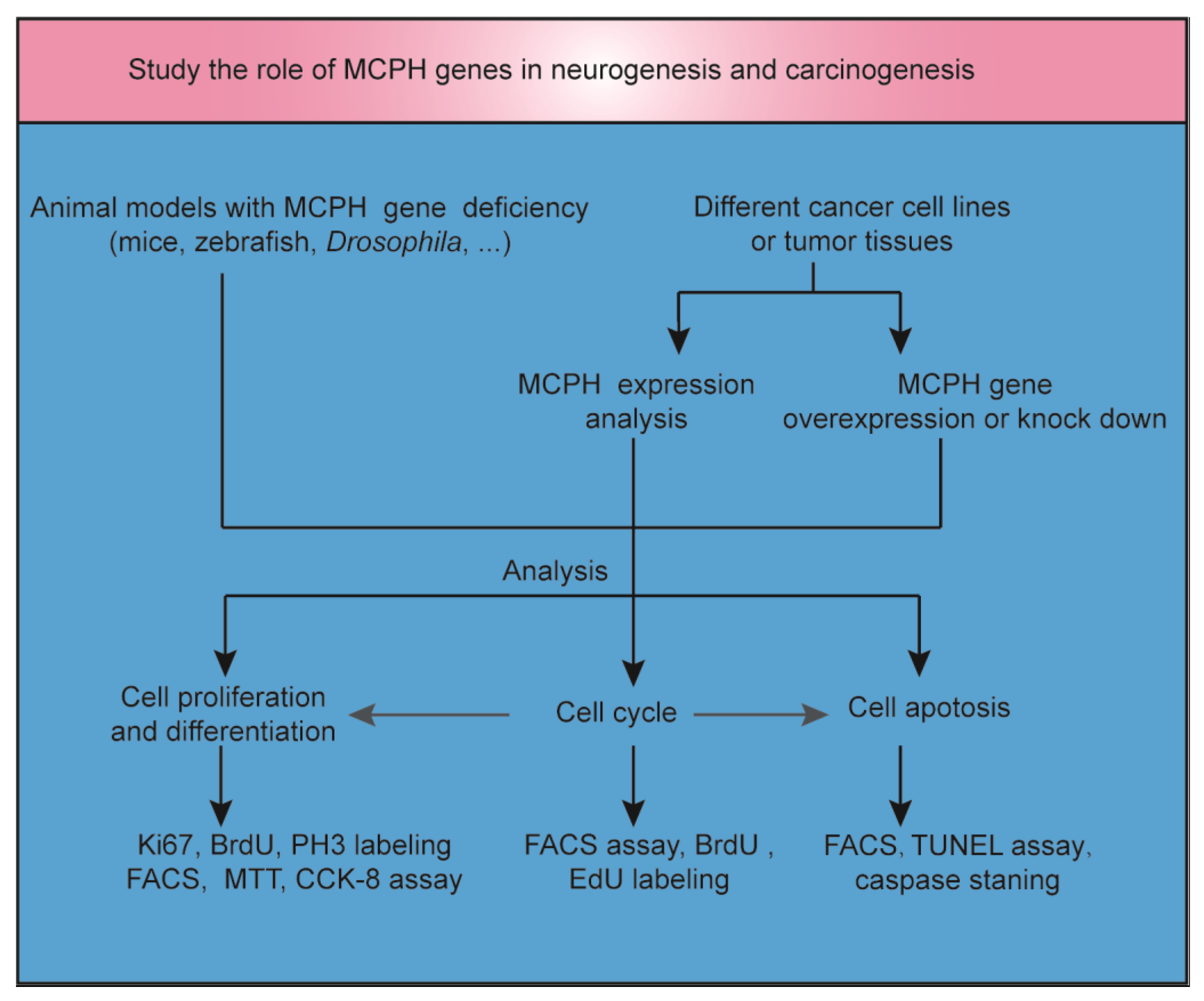{kind=link}
{kind=link}
{kind=link}
| Gene | Role in/Effect on Neurogenesis | Ref. | Role in/Effect on Carcinogenesis | Ref. |
|---|---|---|---|---|
| MCPH1 (BRIT1) | Premature neurogenic production | [16] | Cell cycle and apoptosis (cervical cancer) | [40] |
| Reductions in head circumference, premature chromosome condensation, and hypoplasia of the corpus callosum | [15] | Migration and invasion (lung cancer) | [42] | |
| MCPH2 (WDR62) | Mitotic spindle orientation | [43] | Cell cycle (gastric cancer, GC) | [44] |
| Regulating intermediate neural and glial progenitors | [45] | Centrosome amplification (ovarian cancer, OC) | [46] | |
| Cell cycle, centriole biogenesis, and mitotic spindle orientation | [17] | Cell growth (lung adenocarcinoma) | [34] | |
| Regulating neural stem cell division | [47,48] | |||
| MCPH3 (CDK5RAP2) | Reduced neurons and neural progenitors, premature cell cycle exit, and increased cell death | [49] | N/A (Leukemia) | [50] |
| Premature neuronal differentiation | [51] | |||
| MCPH4 (CASC5) | N/A | Cell proliferation and apoptosis (colorectal cancers, CRCs) | [52] | |
| Nuclear division of cells (GC) | [27] | |||
| MCPH5 (ASPM) | Defects in cortical layers IV, V, and VI formation and imbalance of horizontal and vertical neurites | [53] | Differentiation and metastasis (bladder cancer) | [35] |
| Abnormal proliferation and differentiation of nerve stem/progenitor cells | [54] | Cell cycle progression (pancreatic cancer prognosis) | [55] | |
| MCPH6 (CENPJ) | Cell proliferation, cell apoptosis | [56] | N/A | |
| Centrosome generation and microtubule stability, progenitor division, and neuronal migration | [57] | |||
| MCPH7 (STIL) | Centriole duplication | [58,59] | Cell proliferation, cell cycle G2/M phase and apoptosis (GC) | [32] |
| Centromere assembly | [60] | Cell proliferation and apoptosis (prostate cancer, PCa) | [28] | |
| Cell cycle and chromosomal segregation | [61] | DNA repair (OC) | [62] | |
| MCPH8 (CEP135) | Assembly of centrosome and microtubule | [63] | Centriole duplication (breast cancer) | [29] |
| Establishment of centrosome asymmetry | [64] | |||
| MCPH10 (ZNF335) | Neural progenitor self-renewal, neurogenesis, and neuronal differentiation | [65] | tumor progression (Bladder cancer) | [24] |
| MCPH12 (CDK6) | Regulating radial glial cells G1 and S phases | [66] | DNA damage response, apoptosis (epithelial ovarian cancer) | [67] |
| Positively regulates the proliferation of hippocampal progenitors | [68] | Cell cycle (glioblastoma) | [69] | |
| Proliferation of neural stem cells | [70] | Cell cycle, cell proliferation and angiogenesis (hematopoietic malignancies) | [71] | |
| Cell cycle and apoptosis (T-cell acute lymphoblastic leukemia) | [72] | |||
| MCPH13 (CENPE) | N/A | Cell G2/M phase and proliferation (lung cancer) | [25] | |
| MCPH14 (SAS6) | N/A | Centrosome amplification, mitotic abnormality (CRCs) | [73] | |
| MCPH15 (MFSD2A) | Blood–brain barrier disruption | [74] | Cell cycle (G1 phase) and matrix attachment (lung cancer) | [41] |
| MCPH16 (ANKLE2) | Reduced cell proliferation | [75] | N/A | |
| MCPH17 (CIT) | Apoptosis of neuronal progenitors [76,77] | Cell cycle and apoptosis (colon cancer, CC) | [26] | |
| DNA damage, proliferation, cell senescence and apoptosis (medulloblastoma) | [33] | |||
| MCPH18 (WDFY3) | Perinatal lethality, telencephalic junction, axonal connectivity defect, and localization of glial guidepost cells | [23] | Cell proliferation, migration, invasion, and epithelial-to-mesenchymal transition (OC) | [78] |
| Neurodevelopmental delay, intellectual disability, macrocephaly, and psychiatric disorders | [79] | |||
| Distribution in neuronal projection and axon guidance | [80] | |||
| MCPH19 (COPB2) | Increased apoptosis in the brain and slow growth of neurospheres | [81] | Cell cycle, apoptosis and proliferation (PCa) | [30,82] |
| Cell proliferation and apoptosis (lung adenocarcinoma) | [83] | |||
| Cell G1 phase, proliferation and apoptosis (cholangiocellular cancers) | [84] | |||
| Suppresses cell proliferation and induces cell G0/G1 or S phase arrest (CC) | [85] | |||
| Cell proliferation and apoptosis (GC) | [86] | |||
| MCPH20 (KIF14) | Flat head, motor impairment, growth retardation, decreased cell proliferation, and cell death | [22] | Induced cell cycle arrest and apoptosis (PCa) | [36] |
| Regulation of the expression of transcription factors (SP1, YY1) (ovarian cancers) | [87] | |||
| Cell cycle and proliferation (hepatocellular carcinoma) | [88] | |||
| MCPH21 (NCAPD2) | Reduced cortex | [89] | Promote cell cycle and enhance [39] invasion (Triple-negative breast cancer) | |
| MCPH23 (NCAPH) | N/A | Cell cycle (G2/M phase), cells proliferation, migration and apoptosis (CC) | [31] | |
| Cell cycle (PCa) | [38] | |||
| MCPH24 (NUP37) | N/A | Cell growth, migration and invasion (hepatocellular carcinoma) | [37] | |
| MCPH25 (MAP11) | Microcephaly, decreased neuronal proliferation, a reduction in white matter, and hypoplasia of corpus callosum | [90] | N/A | |
| Gene | Neurogenesis | Ref | Carcinogenesis | Ref. |
|---|---|---|---|---|
| MCPH1 (BRIT1) | CHK1, CDC25, CDK1 | [16] | SLUG, CDK1, p53, CDH1, MDM2, SNAIL | [42] |
| p53 | [110] | |||
| CyclinA2, CDK2, CDC25C-cyclinB, CDC2, p53, BCL-2, Bax, Cytochrome c, Caspase-3, PARP-1 | [40] | |||
| MCPH2 (WDR62) | JNK | [18] | N/A | |
| AURKA | [119] | |||
| PLK1 | [43] | |||
| MCPH3 (CDK5RAP2) | p35 | [120] | N/A | |
| MST1, TAZ (Hippo) | [121] | |||
| MCPH4 (CASC5) | BUB1, BUBR, ZWINT-1 | [122] | BUB1 | [52] |
| miR-193b-3p | [27] | |||
| p53 | [123] | |||
| MCPH5 (ASPM) | Wnt p53 | [21] [114] | Wnt | [55,124,125] |
| MCPH6 (CENPJ) | KIF2A | [56] | N/A | |
| ASCL1 | [57] | |||
| MCPH7 (STIL) | CPAP | [58] | Casepase-3/7, MAPK/ERK, PI3K/AKT, AMPK | [28] |
| RTTN | [60] | PD-L1 | [126] | |
| CDK1 | [104] | IGF-1/PI3K/Akt | [32] | |
| GLI1(Shh) | [127] | GLI1(Shh) | [128] | |
| MCPH10 (ZNF335) | REST/NRSF | [65] | N/A | |
| MCPH12 (CDK6) | GLI3 (Shh) | [66] | FOXO3 | [67] |
| Neuroglobin | [70] | GLI2 (Shh) | [129] | |
| p27 | [68] | p16INK4a, VEGF-A | [71] | |
| CD25, Notch | [72] | |||
| SUMO1 | [69] | |||
| MCPH13 (CENPE) | N/A | FOXM1 | [25] | |
| MCPH17 (CIT) | Tubulin β-III | [76] | p53 | [26] |
| Trp53 | [77] | Tp53, Tp73 | [33] | |
| MCPH18 (WDFY3) | Wnt | [79] | miR-18a, RORA | [78] |
| MCPH19 (COPB2) | Camk1γb | [130] | p21, WAF1/CIP1, P27 KIP1, CDK2, CDK4, Cyclin D1 | [82] |
| YAP1 (Hippo) | [83] | |||
| RTK | [86] | |||
| MCPH20 (KIF14) | N/A | p27 (KIP1), SKP2, CKS1 | [88] | |
| GADD45A, GADD45B, P21, PIDD, SHISA5, p53, Hippo | [36] | |||
| AKT | [131] | |||
| SP1, YY1 | [87] | |||
| MCPH21 (NCAPD2) | N/A | p53 | [39] | |
| MCPH24 (NUP37) | N/A | YAP (Hippo) | [37] | |
| MCPH25 (MAP11) | PLK1 | [90] | N/A | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Zhi, Y.; Yu, J.; Xu, D. The Yin and Yang of Autosomal Recessive Primary Microcephaly Genes: Insights from Neurogenesis and Carcinogenesis. Int. J. Mol. Sci. 2020, 21, 1691. https://doi.org/10.3390/ijms21051691
Zhou X, Zhi Y, Yu J, Xu D. The Yin and Yang of Autosomal Recessive Primary Microcephaly Genes: Insights from Neurogenesis and Carcinogenesis. International Journal of Molecular Sciences. 2020; 21(5):1691. https://doi.org/10.3390/ijms21051691
Chicago/Turabian StyleZhou, Xiaokun, Yiqiang Zhi, Jurui Yu, and Dan Xu. 2020. "The Yin and Yang of Autosomal Recessive Primary Microcephaly Genes: Insights from Neurogenesis and Carcinogenesis" International Journal of Molecular Sciences 21, no. 5: 1691. https://doi.org/10.3390/ijms21051691
APA StyleZhou, X., Zhi, Y., Yu, J., & Xu, D. (2020). The Yin and Yang of Autosomal Recessive Primary Microcephaly Genes: Insights from Neurogenesis and Carcinogenesis. International Journal of Molecular Sciences, 21(5), 1691. https://doi.org/10.3390/ijms21051691