Efficient Cardiac Differentiation of Human Amniotic Fluid-Derived Stem Cells into Induced Pluripotent Stem Cells and Their Potential Immune Privilege

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characterization of hPSCs with and without Cardiac Differentiation
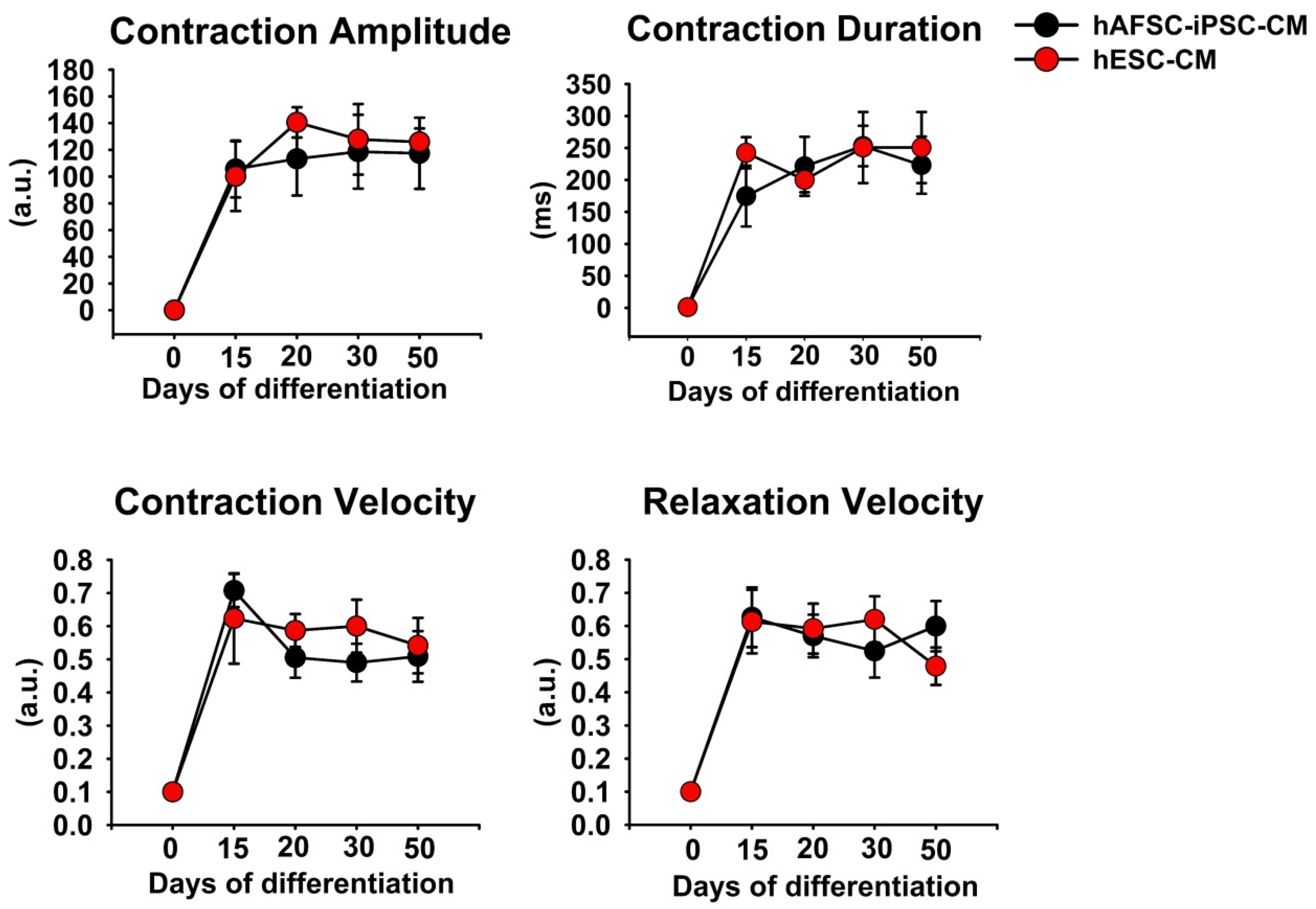
2.2. Contractility of hPSC-CMs
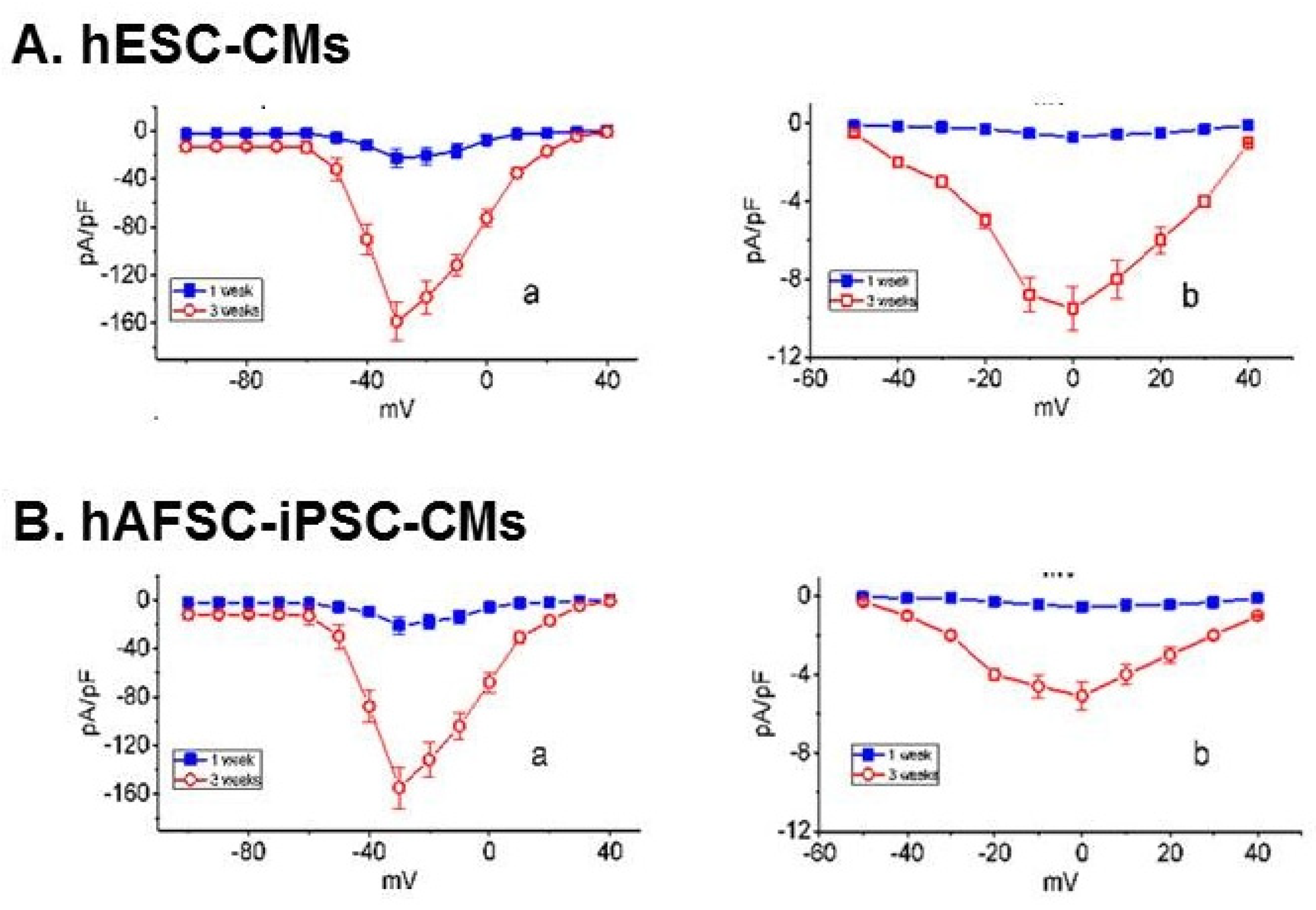
2.3. Characterization of the electrophysiological properties of hAFSC-iPSC-CMs and hESC-CMs
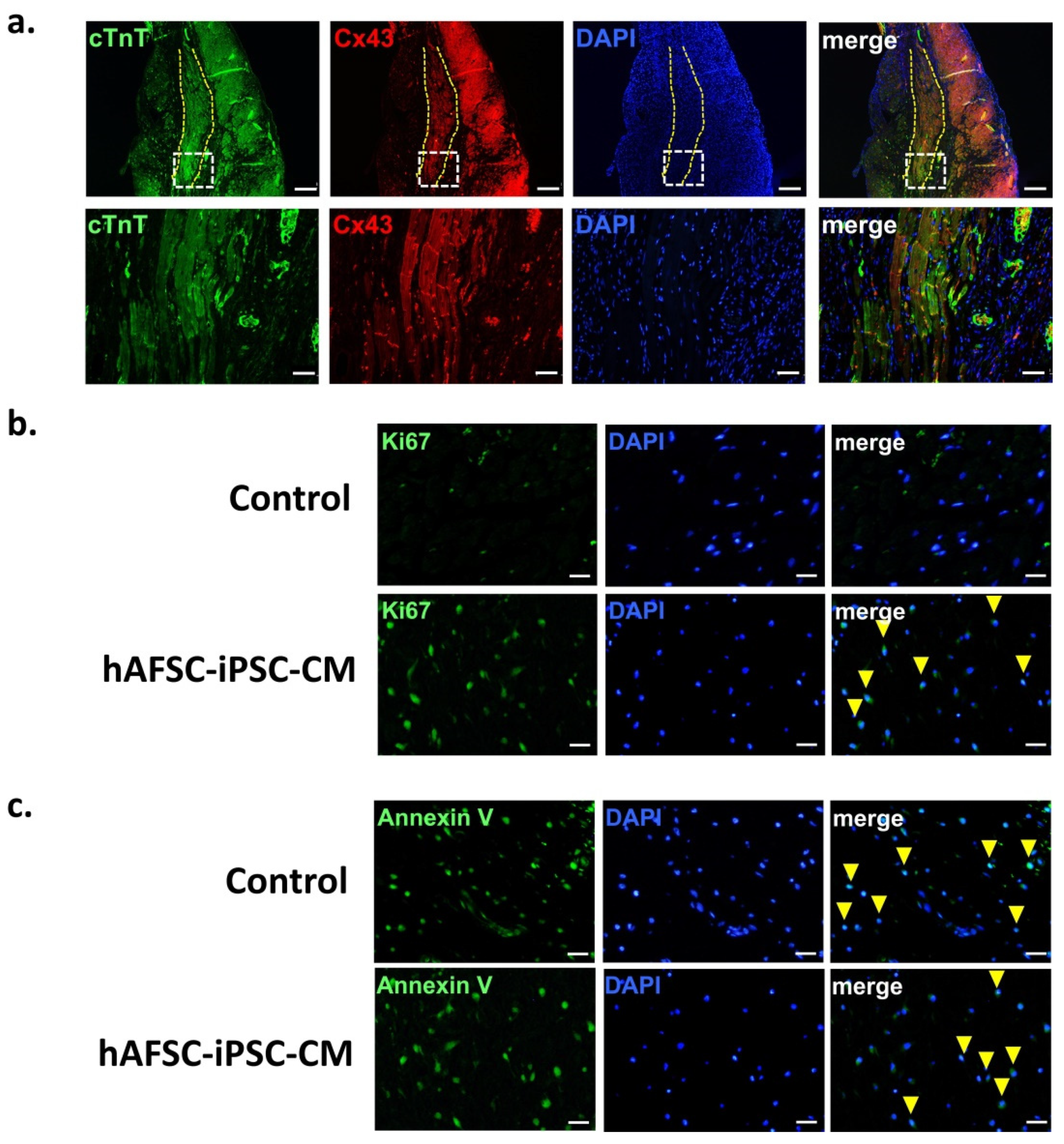
2.4. Therapeutic Effects of hAFSC-iPSC-CMs on Post-Myocardial Infarction (MI) Left Ventricular (LV) Function
3. Discussion
4. Materials and Methods
4.1. hPSCs Culture and Cardiac Differentiation
4.2. Characterization of Differentiated Cardiomyocytes
4.2.1. Immunofluorescent Staining
4.2.2. Flow Cytometry
4.2.3. Contractility Measurements of hAFSC-iPSC-CMs and hESC-CMs
4.3. Electrophysiological Measurements
4.4. Electrophysiological Data Analysis
4.5. Rat Myocardial Infarction Model
4.6. Cells Preparation and Intramyocardial Transplantation
4.7. Cardiac Function Evaluation
4.8. Histology and Immunohistochemistry
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CsA | cyclosporine A |
| cTnT | cardiac troponin T |
| FS | fractional shortening |
| hAFSC | human amniotic fluid-derived stem cell |
| hAFSC-iPSC | human amniotic fluid-derived stem cell induced pluripotent stem cell |
| hAFSC-iPSC-CM | human amniotic fluid-derived stem cell induced pluripotent stem cell derived cardiomyocyte |
| hESC | human embryonic stem cell |
| HLA | human leukocyte antigen |
| hPSC | human pluripotent stem cell |
| hPSC-CM | human pluripotent stem cell derived cardiomyocyte |
| ICa,L | L-type Ca2+ current |
| INa | Na+ current |
| iPSC | induced pluripotent stem cell |
| I-R | ischemia-reperfusion |
| LAD | left anterior descending coronary artery |
| LV | left ventricular |
| LVEDD | left ventricular end-diastolic diameter |
| LVEDS | left ventricular end-diastolic volume |
| LVEF | left ventricular ejection fraction |
| LVESD | left ventricular end-systolic diameter |
| LVESV | left ventricular end-systolic volume |
| MHC | major histocompatibility complex |
| MI | myocardial infarction |
| OCT-4 | octamer-binding transcription factor 4 |
| SSEA4 | Nanog and stage-specific embryonic antigen-4 |
References
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Fernandes, S.; Zhu, W.Z.; Filice, D.; Muskheli, V.; Kim, J.; Palpant, N.J.; Gantz, J.; Moyes, K.W.; Reinecke, H.; et al. Human ES-cell-derived cardiomyocytes electrically couple and suppress arrhythmias in injured hearts. Nature 2012, 489, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.; Yang, X.; Don, C.W.; Minami, E.; Liu, Y.W.; Weyers, J.J.; Mahoney, W.M.; Van Biber, B.; Cook, S.M.; Palpant, N.J.; et al. Human embryonic-stem-cell-derived cardiomyocytes regenerate non-human primate hearts. Nature 2014, 510, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.W.; Chen, B.; Yang, X.; Fugate, J.A.; Kalucki, F.A.; Futakuchi-Tsuchida, A.; Couture, L.; Vogel, K.W.; Astley, C.A.; Baldessari, A.; et al. Human embryonic stem cell-derived cardiomyocytes restore function in infarcted hearts of non-human primates. Nat. Biotechnol. 2018, 36, 597–605. [Google Scholar] [CrossRef]
- Caspi, O.; Huber, I.; Kehat, I.; Habib, M.; Arbel, G.; Gepstein, A.; Yankelson, L.; Aronson, D.; Beyar, R.; Gepstein, L. Transplantation of human embryonic stem cell-derived cardiomyocytes improves myocardial performance in infarcted rat hearts. J. Am. Coll. Cardiol. 2007, 50, 1884–1893. [Google Scholar] [CrossRef]
- van Laake, L.W.; Passier, R.; Doevendans, P.A.; Mummery, C.L. Human embryonic stem cell-derived cardiomyocytes and cardiac repair in rodents. Circ. Res. 2008, 102, 1008–1010. [Google Scholar] [CrossRef] [Green Version]
- Faiella, W.; Atoui, R. Therapeutic use of stem cells for cardiovascular disease. Clin. Transl. Med. 2016, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Romagnuolo, R.; Masoudpour, H.; Porta-Sanchez, A.; Qiang, B.; Barry, J.; Laskary, A.; Qi, X.; Masse, S.; Magtibay, K.; Kawajiri, H.; et al. Human Embryonic Stem Cell-Derived Cardiomyocytes Regenerate the Infarcted Pig Heart but Induce Ventricular Tachyarrhythmias. Stem Cell Reports 2019, 12, 967–981. [Google Scholar] [CrossRef] [Green Version]
- Reske, A.; Reske, A.; Metze, M. Complications of immunosuppressive agents therapy in transplant patients. Minerva Anestesiol. 2015, 81, 1244–1261. [Google Scholar]
- Katabathina, V.; Menias, C.O.; Pickhardt, P.; Lubner, M.; Prasad, S.R. Complications of Immunosuppressive Therapy in Solid Organ Transplantation. Radiol. Clin. North Am. 2016, 54, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Gruttadauria, S.; di Francesco, F.; Pagano, D.; Vizzini, G.; Cintorino, D.; Spada, M.; Tuzzolino, F.; Gridelli, B. Complications in immunosuppressive therapy of liver transplant recipients. J. Surg. Res. 2011, 168, e137–e142. [Google Scholar] [CrossRef] [PubMed]
- De Coppi, P.; Bartsch, G., Jr.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of amniotic stem cell lines with potential for therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.S.; Lee, J.L.; Chang, Y.J.; Hwang, S.M. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum. Reprod. 2004, 19, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Roan, J.N.; Wang, S.P.; Hwang, S.M.; Tsai, M.S.; Chen, J.H.; Hsieh, P.C. Xenografted human amniotic fluid-derived stem cell as a cell source in therapeutic angiogenesis. Int. J. Cardiol. 2013, 168, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.S.; Hwang, S.M.; Chen, K.D.; Lee, Y.S.; Hsu, L.W.; Chang, Y.J.; Wang, C.N.; Peng, H.H.; Chang, Y.L.; Chao, A.S.; et al. Functional network analysis of the transcriptomes of mesenchymal stem cells derived from amniotic fluid, amniotic membrane, cord blood, and bone marrow. Stem Cells 2007, 25, 2511–2523. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Fang, Y.H.; Su, C.T.; Hwang, S.M.; Liu, P.Y.; Wu, S.N. The biochemical and electrophysiological profiles of amniotic fluid-derived stem cells following Wnt signaling modulation cardiac differentiation. Cell Death Discov. 2019, 5, 59. [Google Scholar] [CrossRef]
- Antonucci, I.; Pantalone, A.; Tete, S.; Salini, V.; Borlongan, C.V.; Hess, D.; Stuppia, L. Amniotic fluid stem cells: A promising therapeutic resource for cell-based regenerative therapy. Curr. Pharm. Des. 2012, 18, 1846–1863. [Google Scholar] [CrossRef] [Green Version]
- Sala, L.; van Meer, B.J.; Tertoolen, L.G.J.; Bakkers, J.; Bellin, M.; Davis, R.P.; Denning, C.; Dieben, M.A.E.; Eschenhagen, T.; Giacomelli, E.; et al. MUSCLEMOTION: A Versatile Open Software Tool to Quantify Cardiomyocyte and Cardiac Muscle Contraction In Vitro and In Vivo. Circ. Res. 2018, 122, e5–e16. [Google Scholar] [CrossRef]
- Shiba, Y.; Gomibuchi, T.; Seto, T.; Wada, Y.; Ichimura, H.; Tanaka, Y.; Ogasawara, T.; Okada, K.; Shiba, N.; Sakamoto, K.; et al. Allogeneic transplantation of iPS cell-derived cardiomyocytes regenerates primate hearts. Nature 2016, 538, 388–391. [Google Scholar] [CrossRef]
- Ye, L.; Chang, Y.H.; Xiong, Q.; Zhang, P.; Zhang, L.; Somasundaram, P.; Lepley, M.; Swingen, C.; Su, L.; Wendel, J.S.; et al. Cardiac repair in a porcine model of acute myocardial infarction with human induced pluripotent stem cell-derived cardiovascular cells. Cell Stem Cell 2014, 15, 750–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menasche, P. Cell therapy trials for heart regeneration—lessons learned and future directions. Nat. Rev. Cardiol. 2018, 15, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Palmquist-Gomes, P.; Perez-Pomares, J.M.; Guadix, J.A. Cell-based therapies for the treatment of myocardial infarction: Lessons from cardiac regeneration and repair mechanisms in non-human vertebrates. Heart Fail Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Menasche, P.; Vanneaux, V. Stem cells for the treatment of heart failure. Curr. Res. Transl. Med. 2016, 64, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Neofytou, E.; O’Brien, C.G.; Couture, L.A.; Wu, J.C. Hurdles to clinical translation of human induced pluripotent stem cells. J. Clin. Invest. 2015, 125, 2551–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikonomopoulos, A.; Kitani, T.; Wu, J.C. Pluripotent stem cell-derived cardiomyocytes as a platform for cell therapy applications: Progress and hurdles for clinical translation. Mol. Ther. 2018, 26, 1624–1634. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Zhang, Z.N.; Rong, Z.; Xu, Y. Immunogenicity of induced pluripotent stem cells. Nature 2011, 474, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, W.; Fu, X.; Xu, Y. The Immunogenicity and immune tolerance of pluripotent stem cell derivatives. Front. Immunol. 2017, 8, 645. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Wu, Q.; Ni, C.; Zhang, P.; Zhong, Z.; Wu, Y.; Wang, Y.; Xu, Y.; Kong, M.; Cheng, H.; et al. Lack of remuscularization following transplantation of human embryonic stem cell-dervied cardiovascular progenitor cells in infarcted nonhuman primates. Circ. Res. 2018, 122, 958–969. [Google Scholar] [CrossRef]
- Tachibana, A.; Santoso, M.R.; Mahmoudi, M.; Shukla, P.; Wang, L.; Bennett, M.; Goldstone, A.B.; Wang, M.; Fukushi, M.; Ebert, A.D.; et al. Paracrine effects of the pluripotent stem cell-derived cardiac myocytes salvage the injured myocardium. Circ. Res. 2017, 121, e22–e36. [Google Scholar] [CrossRef]
- Wu, S.N.; Chern, J.H.; Shen, S.; Chen, H.H.; Hsu, Y.T.; Lee, C.C.; Chan, M.H.; Lai, M.C.; Shie, F.S. Stimulatory actions of a novel thiourea derivative on large-conductance, calcium-activated potassium channels. J. Cell Physiol. 2017, 232, 3409–3421. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Wu, Y.H.; Chen, B.S.; Lo, Y.C.; Liu, Y.C. Underlying mechanism of actions of tefluthrin, a pyrethroid insecticide, on voltage-gated ion currents and on action currents in pituitary tumor (GH3) cells and GnRH-secreting (GT1-7) neurons. Toxicology 2009, 258, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.C.; Tseng, Y.T.; Liu, C.M.; Wu, B.N.; Wu, S.N. Actions of KMUP-1, a xanthine and piperazine derivative, on voltage-gated Na(+) and Ca(2+) -activated K(+) currents in GH3 pituitary tumour cells. Br. J. Pharmacol. 2015, 172, 5110–5122. [Google Scholar] [CrossRef] [PubMed]
- Perkins, K.L. Cell-attached voltage-clamp and current-clamp recording and stimulation techniques in brain slices. J. Neurosci. Methods 2006, 154, 1–18. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic / Cardiac Parameters | Sham (n = 4) | Vehicle-Injected Group (Control; n = 6) | hAFSC-iPSC-CM Transplanted Group (n = 6) | |
|---|---|---|---|---|
| Age (weeks) | 12.0 ± 0.2 | 12.0 ± 0.5 | 12.0 ± 0.3 | |
| Body weight (g) | 379.3 ± 9.8 | 358.1 ± 6.6 | 361.3 ± 7.2 | |
| LVFS (%) | Pre-MI | 40.7 ± 3.7 | 33.9 ± 3.3 | 35.8 ± 3.4 |
| Post-MI 2 days | 35.9 ± 2.0 | 15.8 ± 3.4 | 24.1 ± 1.4 | |
| Post-MI 4 weeks | 45.2 ± 5.0 | 17.8 ± 2.5 | 34.4 ± 1.4 | |
| LVEF (%) | Pre-MI | 70.9 ± 4.0 | 67.2 ± 1.4 | 74.7 ± 2.0 |
| Post-MI 2 days | 70.1 ± 2.4 | 48.6 ± 1.9 | 56.2 ± 1.1 | |
| Post-MI 4 weeks | 74.6 ± 2.3 | 51.0 ± 0.7 | 65.6 ± 1.1 | |
| ∆ LVEF (%), between post-MI 2 days and 4 weeks | N/A | 2.4 ± 1.0 | 9.5 ± 0.6 | |
| Infarct size (% LV) | N/A | 13.3 ± 4.1 | 11.2 ± 0.8 | |
| Graft size (% infarct) | N/A | N/A | 9.1 ± 1.6 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.-H.; Wang, S.P.H.; Gao, Z.-H.; Wu, S.-N.; Chang, H.-Y.; Yang, P.-J.; Liu, P.-Y.; Liu, Y.-W. Efficient Cardiac Differentiation of Human Amniotic Fluid-Derived Stem Cells into Induced Pluripotent Stem Cells and Their Potential Immune Privilege. Int. J. Mol. Sci. 2020, 21, 2359. https://doi.org/10.3390/ijms21072359
Fang Y-H, Wang SPH, Gao Z-H, Wu S-N, Chang H-Y, Yang P-J, Liu P-Y, Liu Y-W. Efficient Cardiac Differentiation of Human Amniotic Fluid-Derived Stem Cells into Induced Pluripotent Stem Cells and Their Potential Immune Privilege. International Journal of Molecular Sciences. 2020; 21(7):2359. https://doi.org/10.3390/ijms21072359
Chicago/Turabian StyleFang, Yi-Hsien, Saprina P.H. Wang, Zi-Han Gao, Sheng-Nan Wu, Hsien-Yuan Chang, Pei-Jung Yang, Ping-Yen Liu, and Yen-Wen Liu. 2020. "Efficient Cardiac Differentiation of Human Amniotic Fluid-Derived Stem Cells into Induced Pluripotent Stem Cells and Their Potential Immune Privilege" International Journal of Molecular Sciences 21, no. 7: 2359. https://doi.org/10.3390/ijms21072359
APA StyleFang, Y. -H., Wang, S. P. H., Gao, Z. -H., Wu, S. -N., Chang, H. -Y., Yang, P. -J., Liu, P. -Y., & Liu, Y. -W. (2020). Efficient Cardiac Differentiation of Human Amniotic Fluid-Derived Stem Cells into Induced Pluripotent Stem Cells and Their Potential Immune Privilege. International Journal of Molecular Sciences, 21(7), 2359. https://doi.org/10.3390/ijms21072359