Kandelia candel Thioredoxin f Confers Osmotic Stress Tolerance in Transgenic Tobacco

 ,
,
Abstract
:1. Introduction
2. Results
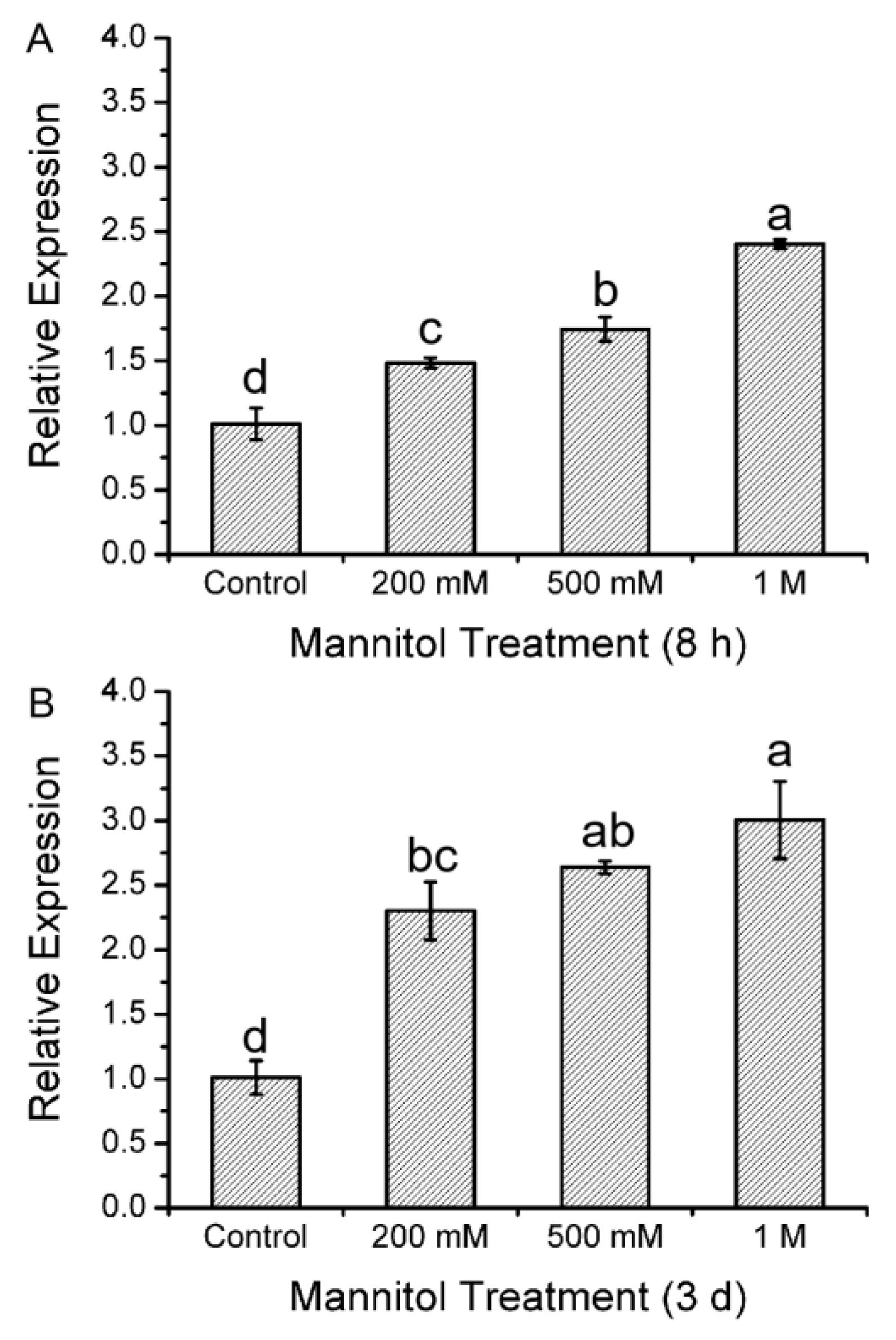
2.1. Expression Profile of KcTrxf Following Exposure to Mannitol in Kandelia Candel Leaves
2.2. KcTrxf Cloning and Sequence Analysis
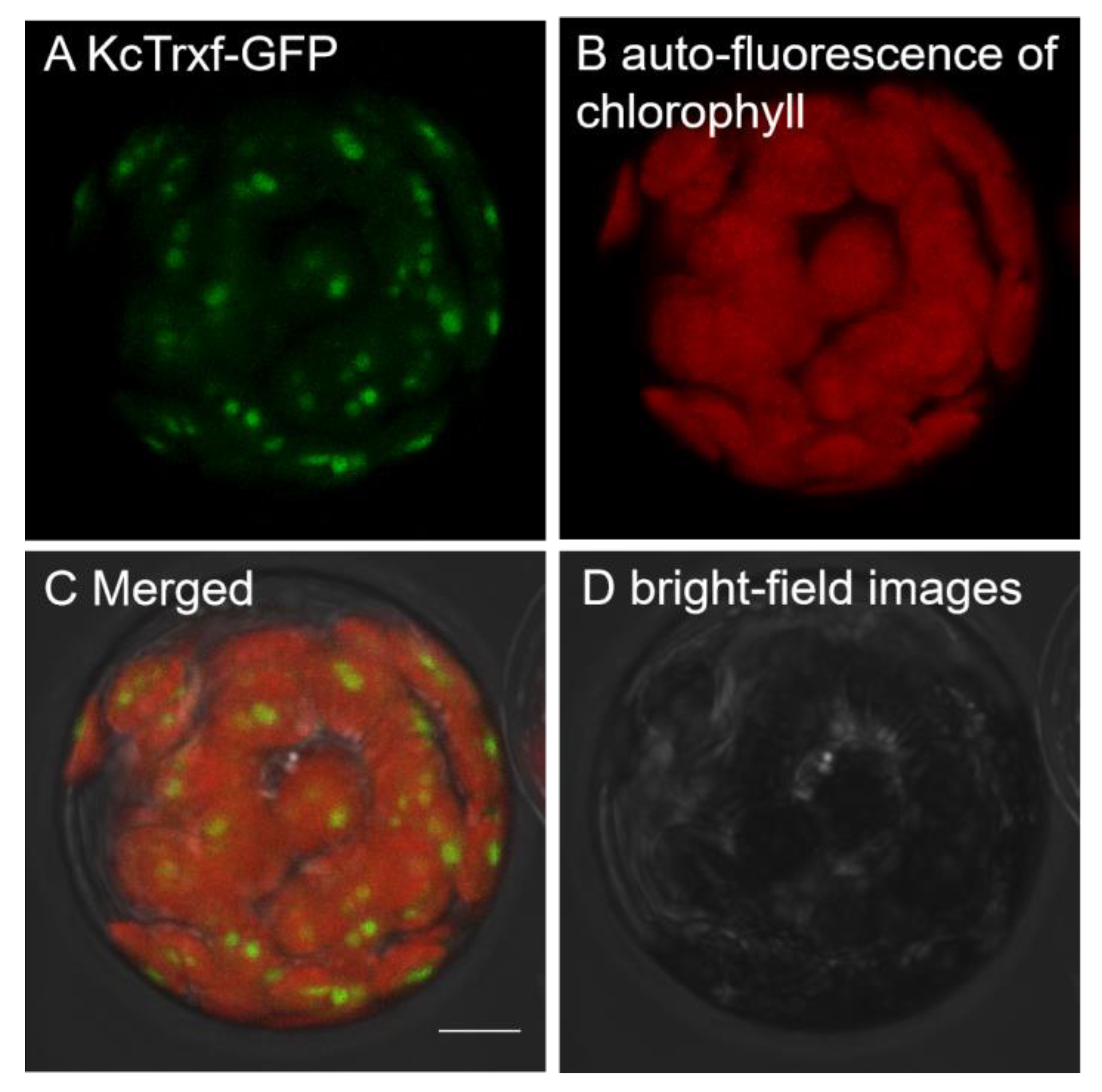
2.3. Subcellular Localization of KcTrxf
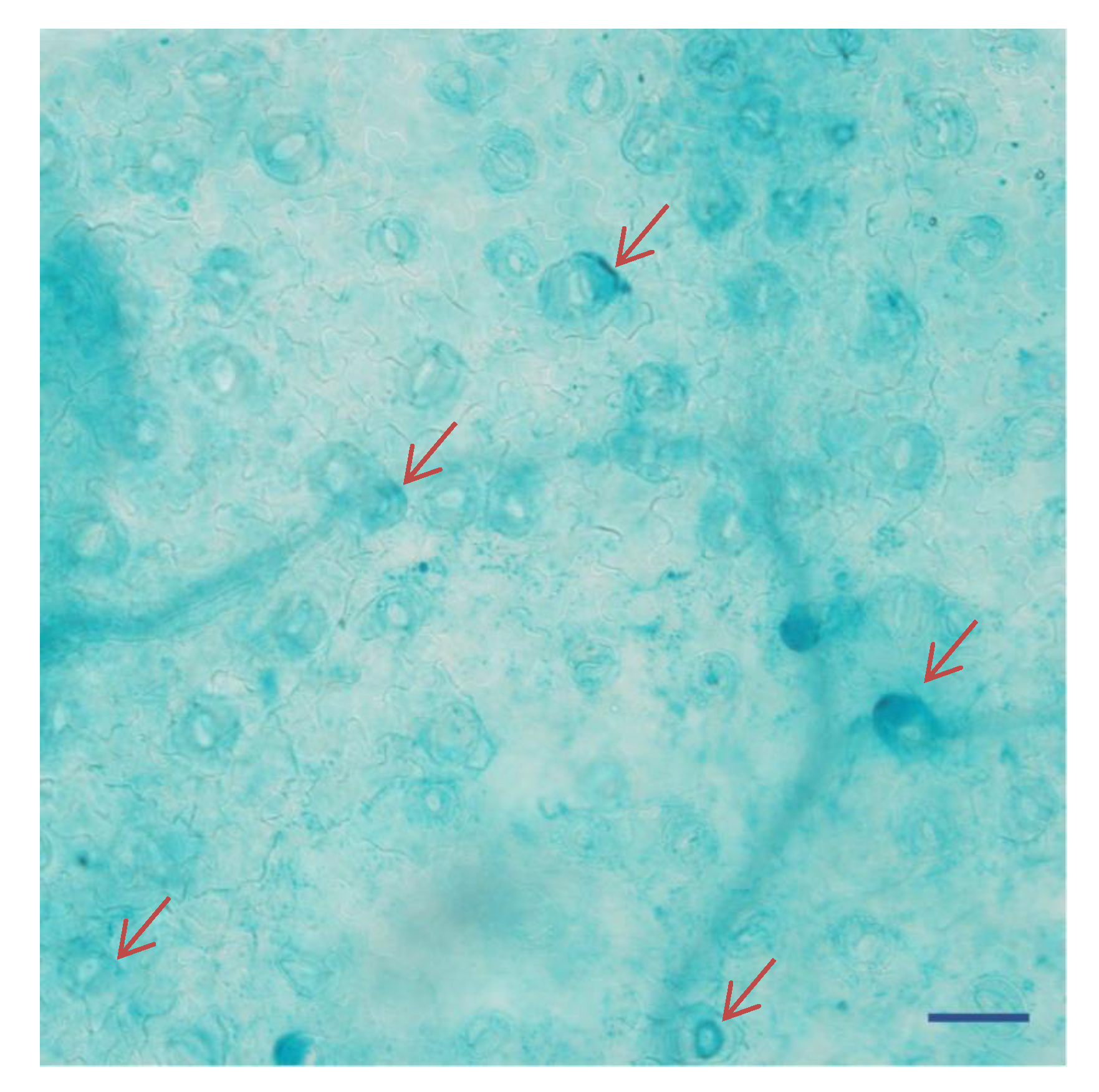
2.4. KcTrxf Promoter:GUS Fusion in Transgenic Arabidopsis Plants
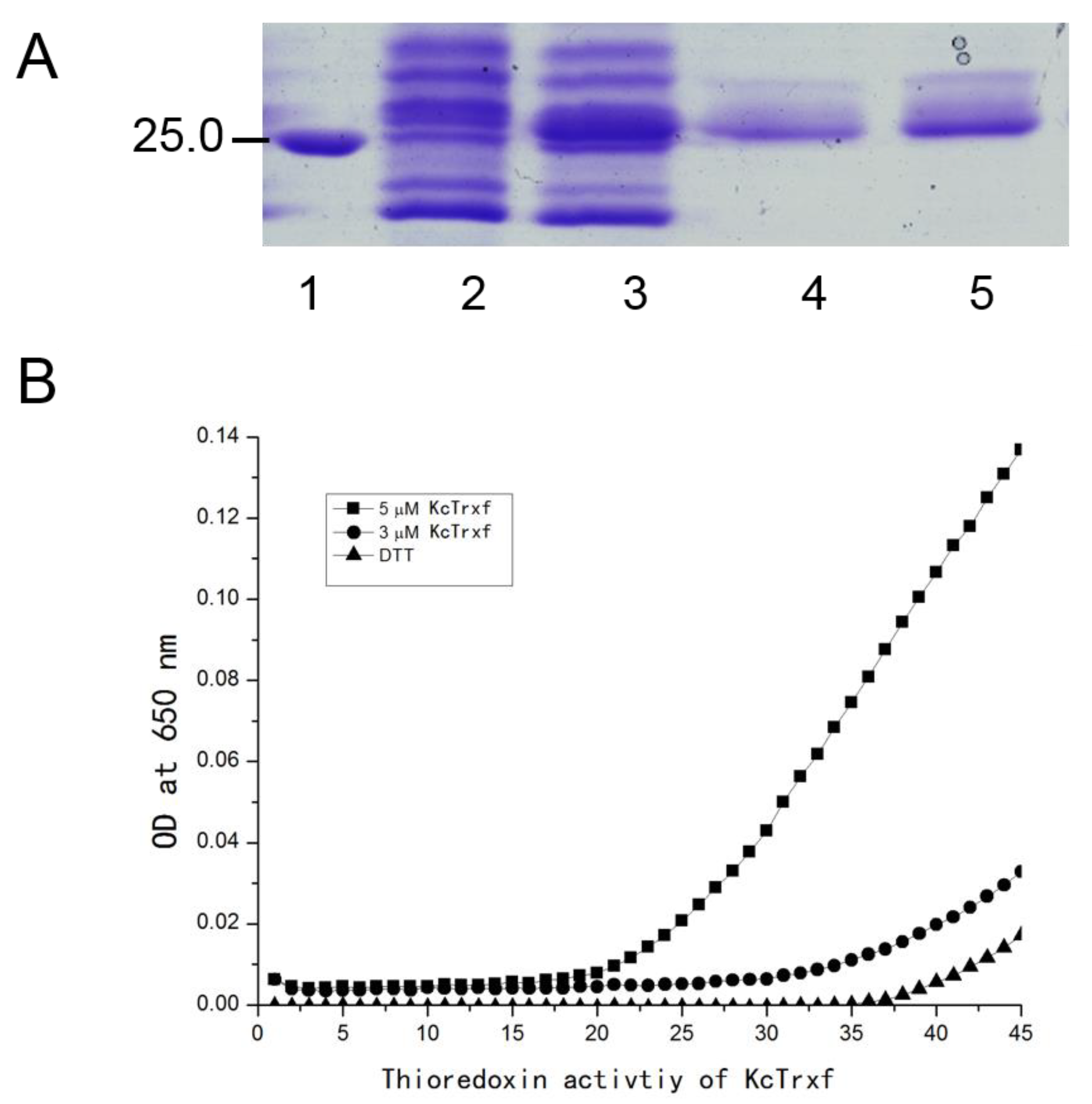
2.5. Purification of Recombinant KcTrxf Protein and Trx Activity
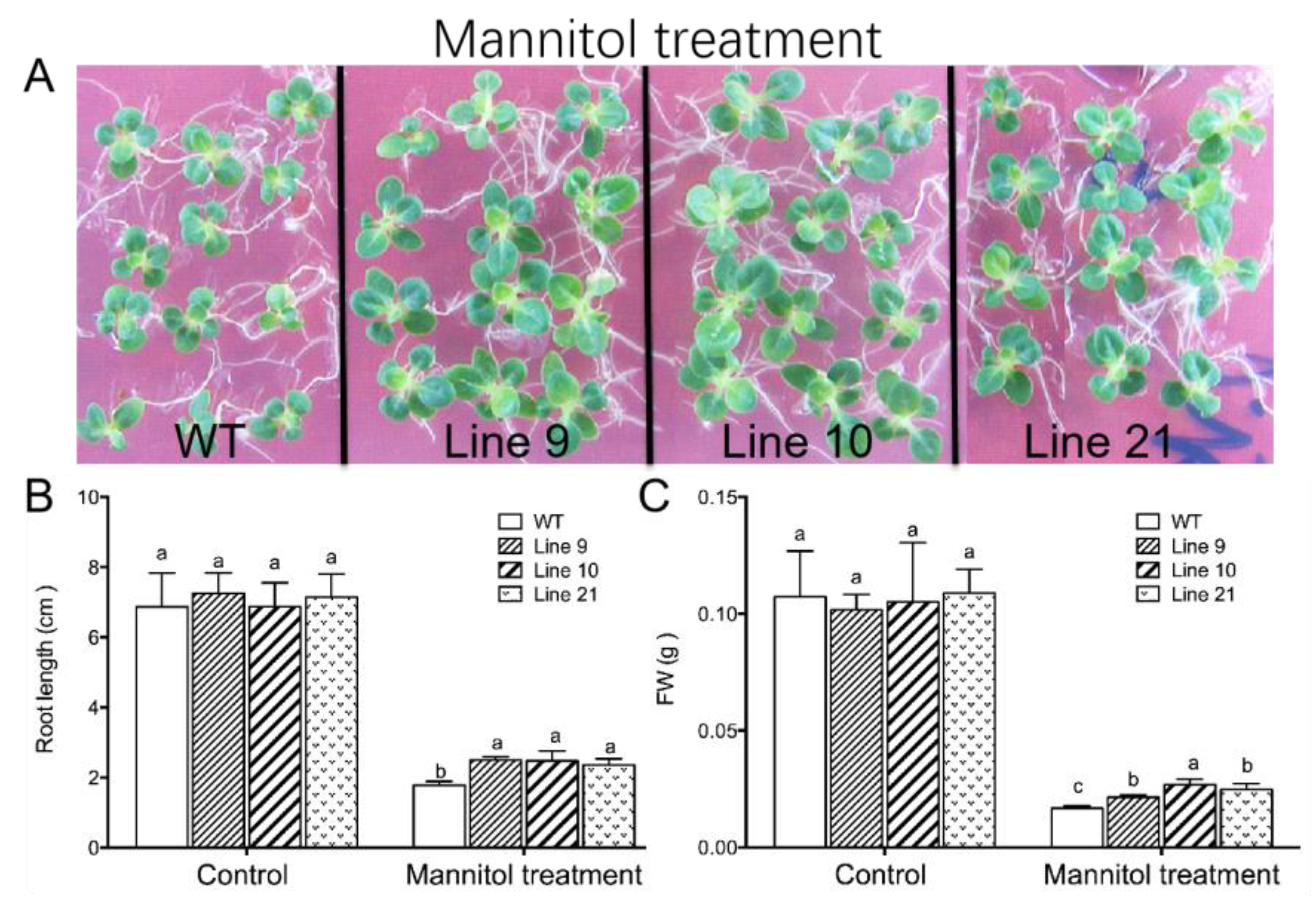
2.6. Overexpression of KcTrxf in Tobacco and Phenotype Tests
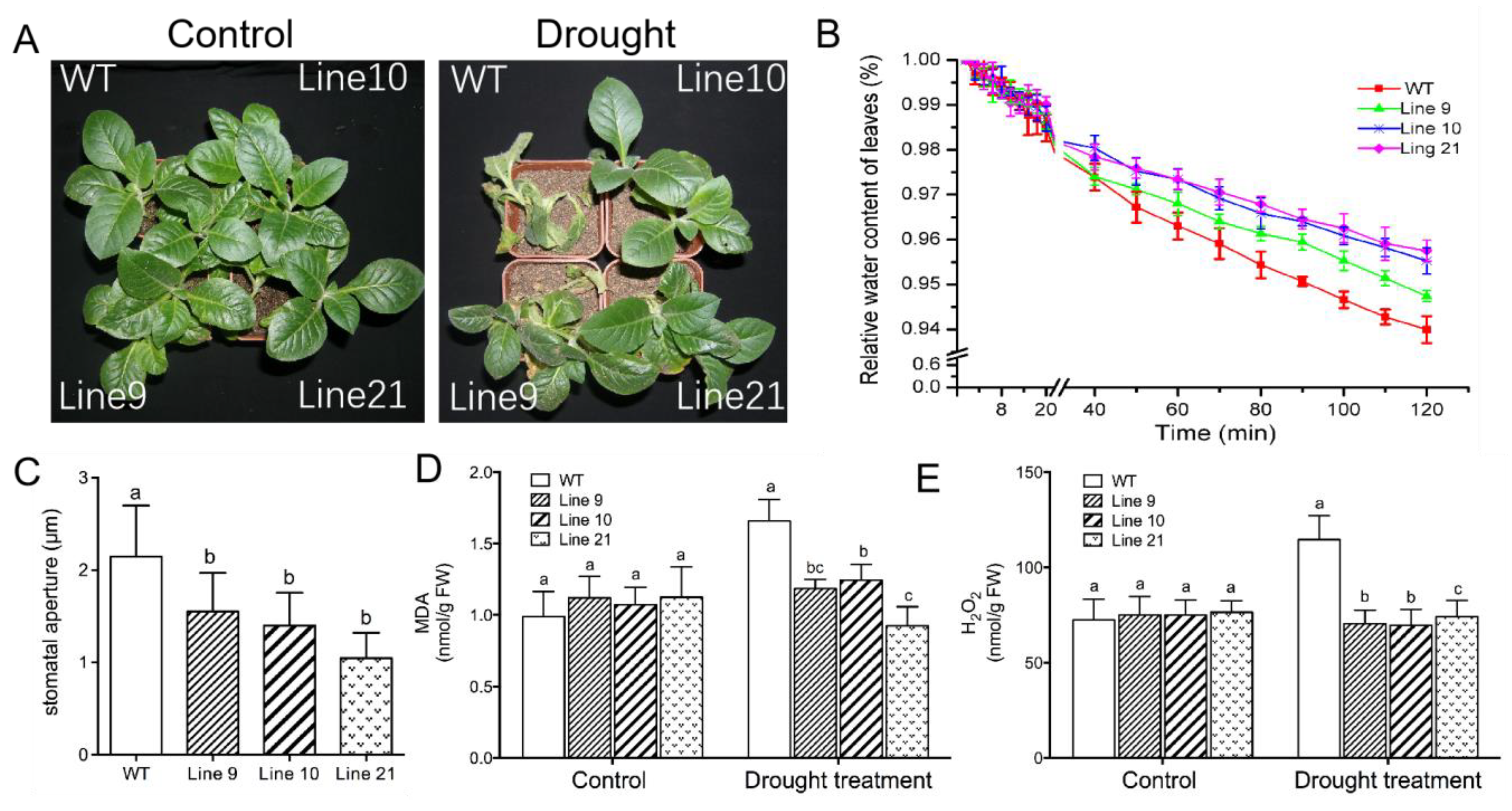
2.7. KcTrxf Overexpression Increases Water-Retaining Capacity and Stomatal Sensitivity to ABA
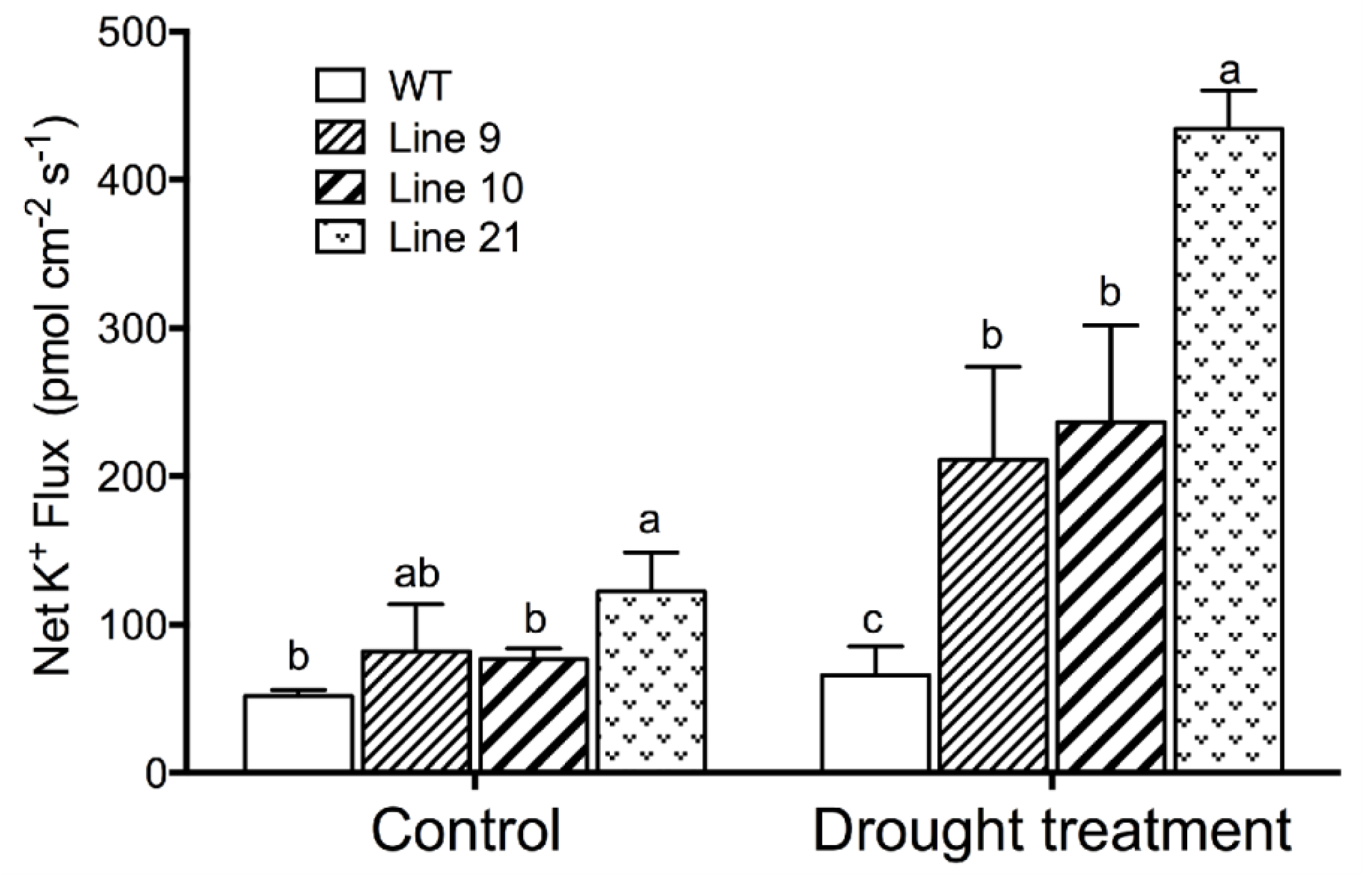
2.8. Drought-Induced K+ Fluxes in Guard Cells
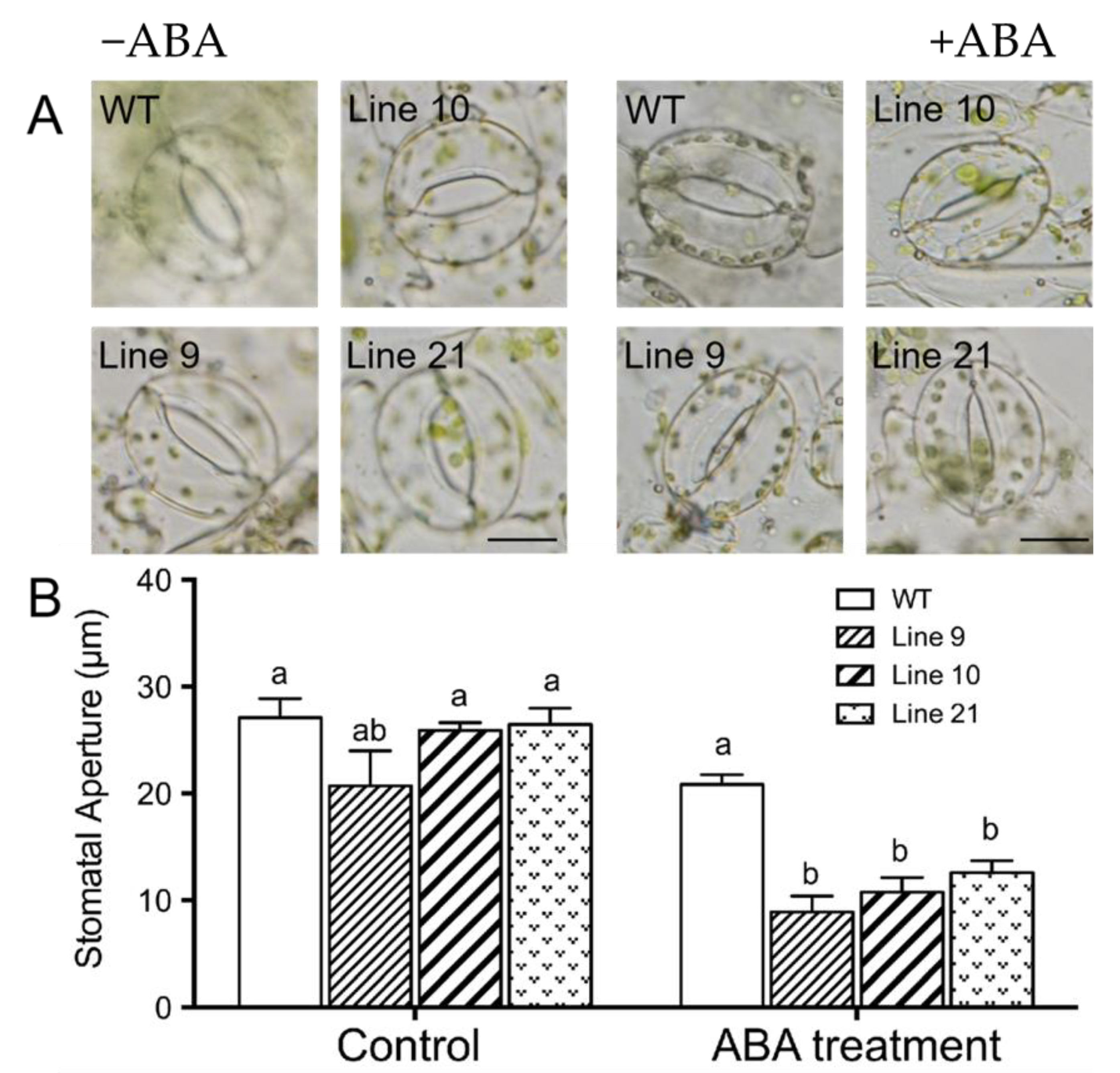
2.9. KcTrxf Overexpression Increases Stomatal Sensitivity to ABA
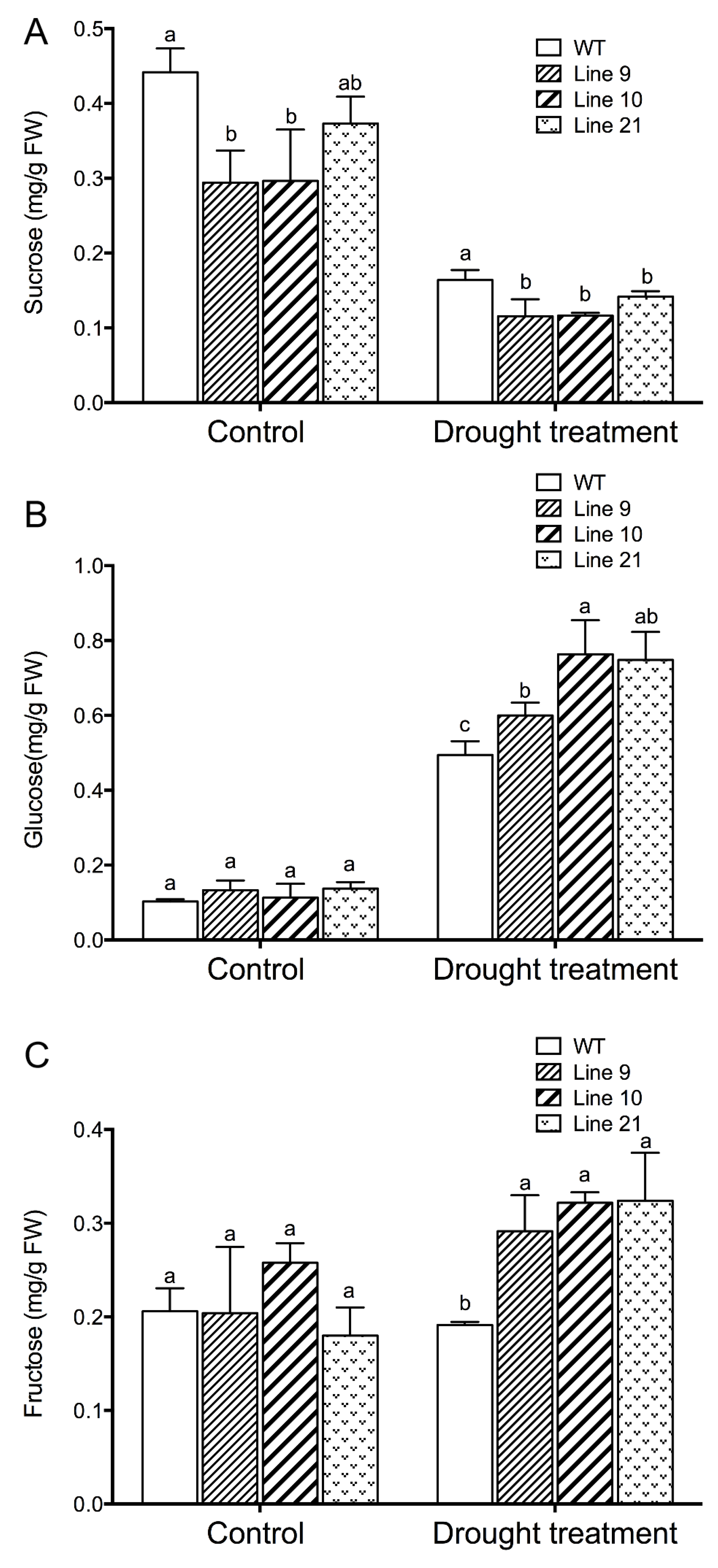
2.10. KcTrxf Increases Glucose and Fructose but Decreased Sucrose under Drought
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Full-Length KcTrxf Gene Cloning and Sequence Analysis
4.3. Subcellular Localization Analysis
4.4. Construction of KcTrxf-pro::GUS and Transformation to Arabidopsis
4.5. KcTrxf Expression and Purification of Recombinant Protein
4.6. Trxf Activity Assay
4.7. KcTrxf Transformation in Tobacco
4.8. Real-Time Quantitative PCR
4.9. Phenotype Tests to Assess Osmotic and Drought Tolerance
4.10. Determination of MDA, H2O2, GSH, and GSSG
4.11. Water-Retaining Capacity
4.12. Stomatal Aperture Measurement
4.13. K+ Flux in Guard Cells
4.14. Soluble Sugar Measurement
4.15. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| DTNB | 5, 5-dithiobis-2-nitrobenzoic acid |
| EDTA | ethylenediaminetetraacetic acid |
| GR | glutathione reductase |
| GSH | reduced glutathione |
| GSSG | oxidized glutathione |
| H2O2 | hydrogen peroxide |
| MDA | malondialdehyde |
| MDAR | monodehydroascorbate reductase |
| MS | Murashige–Skoog medium |
| MES | 2-morpholinoethanesulfonic acid |
| NADPH | reduced nicotinamide adenine dinucleotide phosphate |
| NMT | noninvasive micro-test technique |
| NPTs | nonprotein thiols |
| ORF | open reading frame |
| RT-qPCR | real-time quantitative PCR |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| PM | plasma membrane |
| TRX | thioredoxin |
| TBA | thiobarbituric acid |
| TCA | trichloroacetic acid |
| WRC | water-retaining capacity |
References
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Polle, A.; Chen, S.; Eckert, C.; Harfouche, A. Engineering drought resistance in forest trees. Front. Plant Sci. 2019, 9, 1875. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Thomas, K.; Riglingh, A.; Breshearsi, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Reyer, C.P.O.; Brouwers, N.; Rammig, A.; Brook, B.W.; Epila, J.; Grant, R.F.; Holmgren, M.; Langerwisch1, F.; Leuzinger, S.; Lucht, W.; et al. Forest resilience and tipping points at different spatio-temporal scales: Approaches and challenges. J. Ecol. 2015, 103, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Alonzo, R.; Lopez-Portillo, J.; Moctezuma, C.; Bartlett, M.K.; Sack, L. Osmotic and hydraulic adjustment of mangrove saplings to extreme salinity. Tree Physiol. 2016, 36, 1562–1572. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, R.; Liu, T.; Zheng, H. Solute accumulation and osmotic adjustment characteristics of the mangrove Avicennia marina under NaCl-induced salinity stress. Bot. Mar. 2011, 54, 335–341. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Mittra, B. Effects of salt on growth, ion accumulation, photosynthesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees 2004, 18, 167–174. [Google Scholar] [CrossRef]
- Oh, D.H.; Dassanayake, M.; Bohnert, H.J.; Cheeseman, J.M. Life at the extreme: Lessons from the genome. Genome Biol. 2012, 13, 241. [Google Scholar] [CrossRef]
- Li, N.; Chen, S.; Zhou, X.; Li, C.; Shao, J.; Wang, R.; Fritz, E.; Hüttermann, A.; Polle, A. Effect of NaCl on photosynthesis, salt accumulation and ion compartmentation in two mangrove species, Kandelia candel and Bruguiera gymnorhiza. Aquat. Bot. 2008, 88, 303–310. [Google Scholar] [CrossRef]
- Lu, Y.; Li, N.; Sun, J.; Hou, P.; Jing, X.; Zhu, H.; Deng, S.; Han, Y.; Huang, X.; Ma, X.; et al. Exogenous hydrogen peroxide, nitric oxide and calcium mediate root ion fluxes in two non-secretor mangrove species subjected to NaCl stress. Tree Physiol. 2013, 33, 81–95. [Google Scholar] [CrossRef]
- Lang, T.; Sun, H.; Li, N.; Lu, Y.; Shen, Z.; Jing, X.; Xiang, M.; Shen, X.; Chen, S. Multiple signaling networks of extracellular ATP, hydrogen peroxide, calcium, and nitric oxide in the mediation of root ion fluxes in secretor and non-secretor mangroves under salt stress. Aquat. Bot. 2014, 119, 33–43. [Google Scholar] [CrossRef]
- Jing, X.; Hou, P.; Lu, Y.; Deng, S.; Li, N.; Zhao, R.; Sun, J.; Wang, Y.; Han, Y.; Lang, T.; et al. Overexpression of copper/zinc superoxide dismutase from mangrove Kandelia candel in tobacco enhances salinity tolerance by the reduction of reactive oxygen species in chloroplast. Front. Plant Sci. 2015, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, X.; Sun, Y.; Xiang, M.; Qian, Z.; Lang, T.; Zhao, R.; Shen, X.; Chen, S. Overexpression of KcTrxf in tobacco enhances salt tolerance through the regulation of ROS homeostasis under NaCl stress. J. Beijing For. Univ. 2015, 37, 17–26. [Google Scholar]
- Li, N.; Zhou, X.; Wang, R.; Li, J.; Lu, C.; Chen, S. Salt compartmentation and antioxidant defense in roots and leaves of two non-salt secretor mangroves under salt stress. In Mangrove Ecosystem Ecology and Function; Sahadev, S., Ed.; IntechOpen Limited: London, UK, 2018; pp. 81–104. [Google Scholar]
- Geigenberger, P.; Thormalen, I.; Daloso, D.M.; Fernie, A.R. The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef]
- Polle, A. Dissecting the superoxide dismutase-ascorbate-glutathione pathway by metabolic modeling: Computer analysis as a step towards flux analysis. Plant Physiol. 2001, 126, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Bartoli, C.G.; Buet, A.; Grozeff, G.G.; Galatro, A.; Simontacchi, M. Ascorbate-glutathione cycle and abiotic stress tolerance in plants. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M., Munné-Bosch, S., Burritt, D., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 177–200. [Google Scholar]
- Yan, Y.; Pan, C.; Du, Y.; Li, D.; Liu, W. Exogenous salicylic acid regulates reactive oxygen species metabolism and ascorbate–glutathione cycle in Nitraria tangutorum Bobr. under salinity stress. Physiol. Mol. Biol. Plants 2018, 24, 577–589. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant. J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Fujita, M. Drought stress responses in plants, oxidative stress, and antioxidant defense. In Climate Change and Plant Abiotic Stress Tolerance; Narendra, T., Sarvajeet, S.G., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2013; pp. 209–250. [Google Scholar]
- Sadeghi, F.; Samsampour, D.; Seyahooei, M.A.; Bagheri, A.; Soltani, J. Fungal endophytes alleviate drought-induced oxidative stress in mandarin (Citrus reticulata L.): Toward regulating the ascorbate–glutathione cycle. Sci. Hortic. 2020, 261, 108991. [Google Scholar] [CrossRef]
- Buckley, T.N. The control of stomata by water balance. New Phytol. 2005, 168, 275–292. [Google Scholar] [CrossRef]
- Daszkowska-Golec, A.; Szarejko, I. Open or close the gate-stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodribb, T.J.; Holbrook, N.M. Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol. 2003, 132, 2166–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, W.L.; Fernie, A.R.; Nunes-Nesi, A. Control of stomatal aperture: A renaissance of the old guard. Plant Signal Behav. 2011, 6, 1305–1311. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wang, S.; Altman, A.; Hüttermann, A. Genotypic variation in drought tolerance of poplar in relation to abscisic acid. Tree Physiol. 1997, 17, 797–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Li, J.; Wang, S.; Hüttermann, A.; Altman, A. Salt, nutrient uptake and transport and ABA of Populus euphratica; a hybrid in response to increasing soil NaCl. Trees 2001, 15, 186–194. [Google Scholar] [CrossRef]
- Chen, S.; Li, J.; Wang, T.; Wang, S.; Polle, A.; Hüttermann, A. Osmotic stress and ion-specific effects on xylem abscisic acid and the relevance to salinity tolerance in poplar. J. Plant Growth Regul. 2002, 21, 224–233. [Google Scholar] [CrossRef]
- Chang, Y.; Chen, S.; Yin, W.; Wang, R.; Liu, Y.; Shi, Y.; Shen, Y.; Li, Y.; Jiang, J.; Liu, Y. Growth, gas exchange, abscisic acid, and calmodulin response to salt stress in three poplars. J. Integr. Plant Biol. 2006, 48, 286–293. [Google Scholar] [CrossRef]
- Pantin, F.; Monnet, F.; Jannaud, D.; Costa, J.M.; Renaud, J.; Muller, B.; Genty, B. The dual effect of abscisic acid on stomata. New Phytol. 2013, 197, 65–72. [Google Scholar] [CrossRef]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.Y.; Cutler, S.R.; Sheen, J.; Rodriguez, P.L.; Zhu, J.K. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R. Ion channels in plants. Physiol. Rev. 2012, 92, 1777–1811. [Google Scholar] [CrossRef]
- Homann, U.; Thiel, G. The number of K+ channels in the plasma membrane of guard cell protoplasts changes in parallel with the surface area. Proc. Natl. Acad. Sci. USA 2002, 99, 10215–10220. [Google Scholar] [CrossRef] [Green Version]
- Prodhan, M.Y.; Munemasa, S.; Nahar, M.N.E.N.; Nakamura, Y.; Murata, Y. Guard cell salicylic acid signaling is integrated into abscisic acid signaling via the Ca2+/CPK-dependent pathway. Plant Physiol. 2018, 178, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, J.I.; Allen, G.J.; Hugouvieux, V.; Kwak, J.M.; Waner, D. Guard cell signal transduction. Annu. Rev. Plant Biol. 2001, 52, 627–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Dios Barajas-Lopez, J.; Serrato, A.J.; Olmedilla, A.; Chueca, A.; Sahrawy, M. Localization in roots and flowers of pea chloroplastic thioredoxin f and thioredoxin m proteins reveals new roles in nonphotosynthetic organs. Plant Physiol. 2007, 145, 946–960. [Google Scholar] [CrossRef] [Green Version]
- Janz, D.; Polle, A. Harnessing salt for woody biomass production. Tree Physiol. 2012, 32, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogeat-Triboulot, M.B.; Brosché, M.; Renaut, J.; Jouve, L.; Thiec, D.L.; Fayyaz, P.; Vinocur, B.; Witters, E.; Laukens, K.; Teichmann, T.; et al. Gradual soil water depletion results in reversible changes of gene expression, protein profiles, ecophysiology, and growth performance in Populus euphratica, a poplar growing in arid regions. Plant Physiol. 2007, 143, 876–892. [Google Scholar] [CrossRef] [Green Version]
- Brosché, M.; Vinocur, B.; Alatalo, E.R.; Lamminmäki, A.; Teichmann, T.; Ottow, E.A.; Djilianov, D.; Afif, D.; Bogeat-Triboulot, M.B.; Altman, A.; et al. Gene expression and metabolite profiling of Populus euphratica growing in the Negev desert. Genome Biol. 2005, 6, R101. [Google Scholar] [CrossRef] [Green Version]
- Ottow, E.A.; Brinker, M.; Teichmann, T.; Fritz, E.; Kaiser, W.; Brosché, M.; Kangasjärvi, J.; Jiang, X.; Polle, A. Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress. Plant Physiol. 2005, 139, 1762–1772. [Google Scholar] [CrossRef] [Green Version]
- Dluzniewska, P.; Gessler, A.; Dietrich, H.; Schnitzler, J.-P.; Teuber, M.; Rennenberg, H. Nitrogen uptake and metabolism in Populus × canescens as affected by salinity. New Phytol. 2007, 173, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Ehlting, B.; Dluzniewska, P.; Dietrich, H.; Selle, A.; Teuber, M.; Hänsch, R.; Nehls, U.; Polle, A.; Schnitzler, J.P.; Rennenberg, H.; et al. Interaction of nitrogen nutrition and salinity in Grey poplar (Populus tremula × alba). Plant Cell Environ. 2007, 30, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Pallara, G.; Giovannelli, A.; Traversi, M.L.; Camussi, A.; Racchi, M.L. Effect of water deficit on expression of stress-related genes in the cambial region of two contrasting poplar clones. J. Plant Growth Regul. 2012, 31, 102–112. [Google Scholar] [CrossRef]
- Sanz-Barrio, R.; Corral-Martinez, P.; Ancin, M.; Segui-Simarro, J.M.; Farran, I. Overexpression of plastidial thioredoxin f leads to enhanced starch accumulation in tobacco leaves. Plant Biotechnol. J. 2013, 11, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Thormählen, I.; Ruber, J.; von Roepenack-Lahaye, E.; Ehrlich, S.M.; Massot, V.; Hümmer, C.; Tezycka, J.; Issakidis-Bourguet, E.; Geigenberger, P. Inactivation of thioredoxin f1 leads to decreased light activation of ADP-glucose pyrophosphorylase and altered diurnal starch turnover in leaves of Arabidopsis plants. Plant Cell Environ. 2013, 36, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Sparla, F.; Costa, A.; Lo Schiavo, F.; Pupillo, P.; Paolo, T. Redox regulation of a novel plastid-targeted-amylase of Arabidopsis. Plant Physiol. 2006, 141, 840–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valerio, C.; Costa, A.; Marri, L.; Issakidis-Bourguet, E.; Pupillo, P.; Trost, P.; Sparla, F. Thioredoxin-regulated β-amylase (BAM1) triggers diurnal starch degradation in guard cells, and in mesophyll cells under osmotic stress. J. Exp. Bot. 2011, 62, 545–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seung, D.; Thalmann, M.; Sparla, F.; Abou Hachem, M.; Lee, S.K.; Issakidis-Bourguet, E.; Svensson, B.; Zeeman, S.C.; Santelia, D. Arabidopsis thaliana AMY3 is a unique redox-regulated chloroplastic α-amylase. J. Biol. Chem. 2013, 288, 33620–33633. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; Von Heijne, G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, B.B.; Balmer, Y. Redox regulation: A broadening horizon. Annu. Rev. Plant Biol. 2005, 56, 187–220. [Google Scholar] [CrossRef]
- Schürmann, P.; Buchanan, B.B. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid. Redox Signal. 2008, 10, 1235–1274. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Siala, W.; Bashandy, T.; Riondet, C.; Vignols, F.; Reichheld, J.P. Glutaredoxins and thioredoxins in plants. Biochim. Biophys. Acta 2008, 1783, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin catalyzes the reduction of insulin disulfides by dithiothreitol and dihydrolipoamide. J. Biol. Chem. 1979, 254, 9627–9632. [Google Scholar]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Daloso, D.M.; Dos Anjos, L.; Fernie, A.R. Roles of sucrose in guard cell regulation. New Phytol. 2016, 211, 809–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef]
- Lemaire, S.D.; Michelet, L.; Zaffagnini, M.; Massot, V.; Issakidis-Bourguet, E. Thioredoxins in chloroplasts. Curr. Genet. 2007, 51, 343–365. [Google Scholar] [CrossRef]
- Lindahl, M.; Kieselbach, T. Disulfide proteomes and interactions with thioredoxin on the track towards understanding redox regulation in chloroplasts and cyanobacteria. J. Proteomics. 2009, 72, 416–438. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- de Dios Barajas-Lopez, J.; Tezycka, J.; Travaglia, C.N.; Serrato, A.J.; Chueca, A.; Thormählen, I.; Geigenberger, P.; Sahrawy, M. Expression of the chloroplast thioredoxins f and m is linked to short-term changes in the sugar and thiol status in leaves of Pisum sativum. J. Exp. Bot. 2012, 63, 4887–4900. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.; Sun, J.; Zhao, R.; Ding, M.; Zhang, Y.; Sun, Y.; Wang, W.; Tan, Y.; Liu, D.; Ma, X.; et al. Populus euphratica APYRASE2 enhances cold tolerance by modulating vesicular trafficking and extracellular ATP in Arabidopsis plants. Plant Physiol. 2015, 169, 530–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.G.; Chen, Y. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Biotechniques 2007, 43, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Yao, J.; Sun, J.; Chang, L.; Wang, S.; Ding, M.; Qian, Z.; Zhang, H.; Zhao, N.; Sa, G.; et al. Populus euphratica HSF binds the promoter of WRKY1 to enhance salt tolerance. Plant Sci. 2015, 235, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Han, Y.; Wang, W.; Sun, J.; Ding, M.; Zhao, R.; Deng, S.; Wang, F.; Hu, Y.; Wang, Y.; Lu, Y.; et al. Populus euphratica XTH overexpression enhances salinity tolerance by the development of leaf succulence in transgenic tobacco plants. J. Exp. Bot. 2013, 64, 4225–4238. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Dai, S.; Wang, R.; Chen, S.; Li, N.; Zhou, X.; Lu, C.; Shen, X.; Zheng, X.; Hu, Z.; et al. Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol. 2009, 29, 1175–1186. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Roscher, A.; Ratcliffe, R.G.; Kruger, N.J. Fructose 2,6-bisphosphate activates pyrophosphate: Fructose-6-phosphate 1-phosphotransferase and increases triose phosphate to hexose phosphate cycling in heterotrophic cells. Planta 2001, 212, 250–263. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | Control | Drought | |
|---|---|---|---|
| GSH | WT | 28.01 ± 0.16a | 24.34 ± 0.71a |
| Line 9 | 16.50 ± 0.38c | 17.68 ± 0.17b | |
| Line 10 | 15.57 ± 0.65c | 15.38 ± 0.72b | |
| Line 21 | 20.87 ± 0.38b | 15.91 ± 0.24b | |
| GSSG | WT | 16.48 ± 0.73b | 21.40 ± 0.92a |
| Line 9 | 18.87 ± 0.44ab | 21.97 ± 0.87a | |
| Line 10 | 18.42 ± 0.68ab | 21.55 ± 0.11a | |
| Line 21 | 19.51 ± 0.77a | 20.11 ± 0.71a | |
| GSSG/GSH | WT | 0.59 ± 0.02c | 0.88 ± 0.02b |
| Line 9 | 1.14 ± 0.03a | 1.24 ± 0.04a | |
| Line 10 | 1.18 ± 0.05a | 1.40 ± 0.06a | |
| Line 21 | 0.93 ± 0.04b | 1.26 ± 0.05a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, X.; Yao, J.; Ma, X.; Zhang, Y.; Sun, Y.; Xiang, M.; Hou, P.; Li, N.; Zhao, R.; Li, J.; et al. Kandelia candel Thioredoxin f Confers Osmotic Stress Tolerance in Transgenic Tobacco. Int. J. Mol. Sci. 2020, 21, 3335. https://doi.org/10.3390/ijms21093335
Jing X, Yao J, Ma X, Zhang Y, Sun Y, Xiang M, Hou P, Li N, Zhao R, Li J, et al. Kandelia candel Thioredoxin f Confers Osmotic Stress Tolerance in Transgenic Tobacco. International Journal of Molecular Sciences. 2020; 21(9):3335. https://doi.org/10.3390/ijms21093335
Chicago/Turabian StyleJing, Xiaoshu, Jun Yao, Xujun Ma, Yanli Zhang, Yuanling Sun, Min Xiang, Peichen Hou, Niya Li, Rui Zhao, Jinke Li, and et al. 2020. "Kandelia candel Thioredoxin f Confers Osmotic Stress Tolerance in Transgenic Tobacco" International Journal of Molecular Sciences 21, no. 9: 3335. https://doi.org/10.3390/ijms21093335
APA StyleJing, X., Yao, J., Ma, X., Zhang, Y., Sun, Y., Xiang, M., Hou, P., Li, N., Zhao, R., Li, J., Zhou, X., & Chen, S. (2020). Kandelia candel Thioredoxin f Confers Osmotic Stress Tolerance in Transgenic Tobacco. International Journal of Molecular Sciences, 21(9), 3335. https://doi.org/10.3390/ijms21093335