Effect of Iron Source and Medium pH on Growth and Development of Sorbus commixta In Vitro



Abstract
:1. Introduction
2. Results
2.1. Morphology and Growth Parameters Analyses
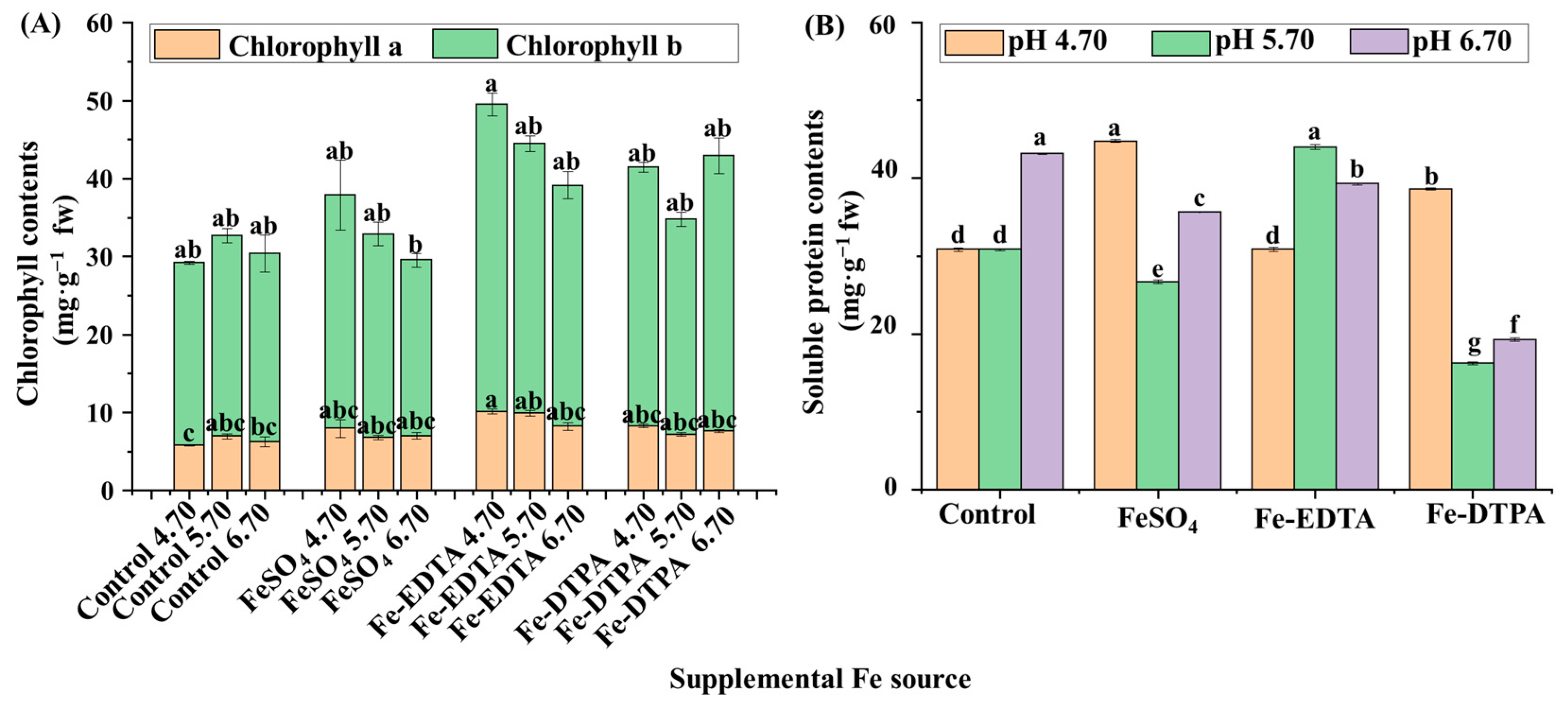
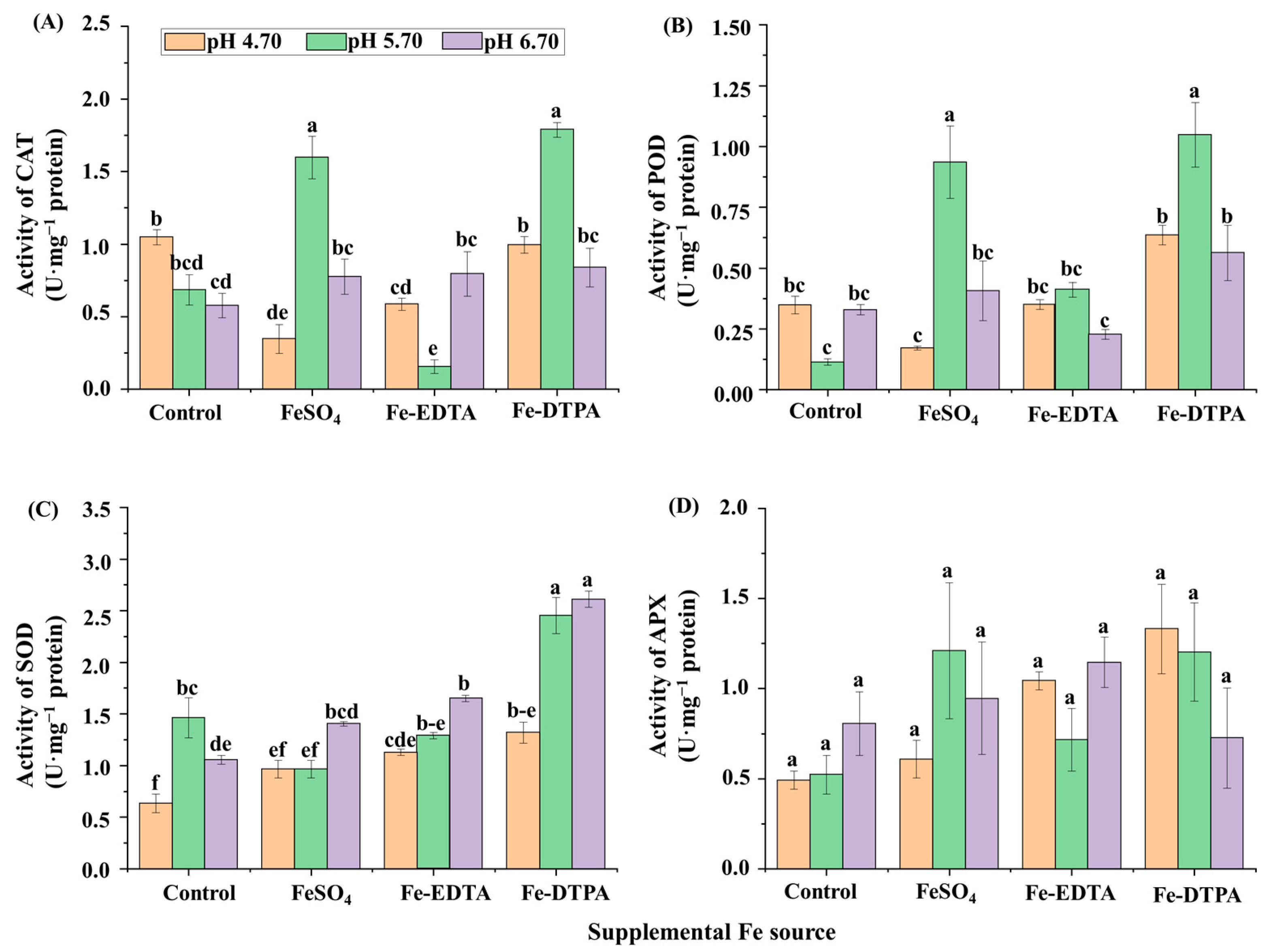
2.2. Chlorophyll Contents, Soluble Proteins, and Antioxidant Enzyme Activities
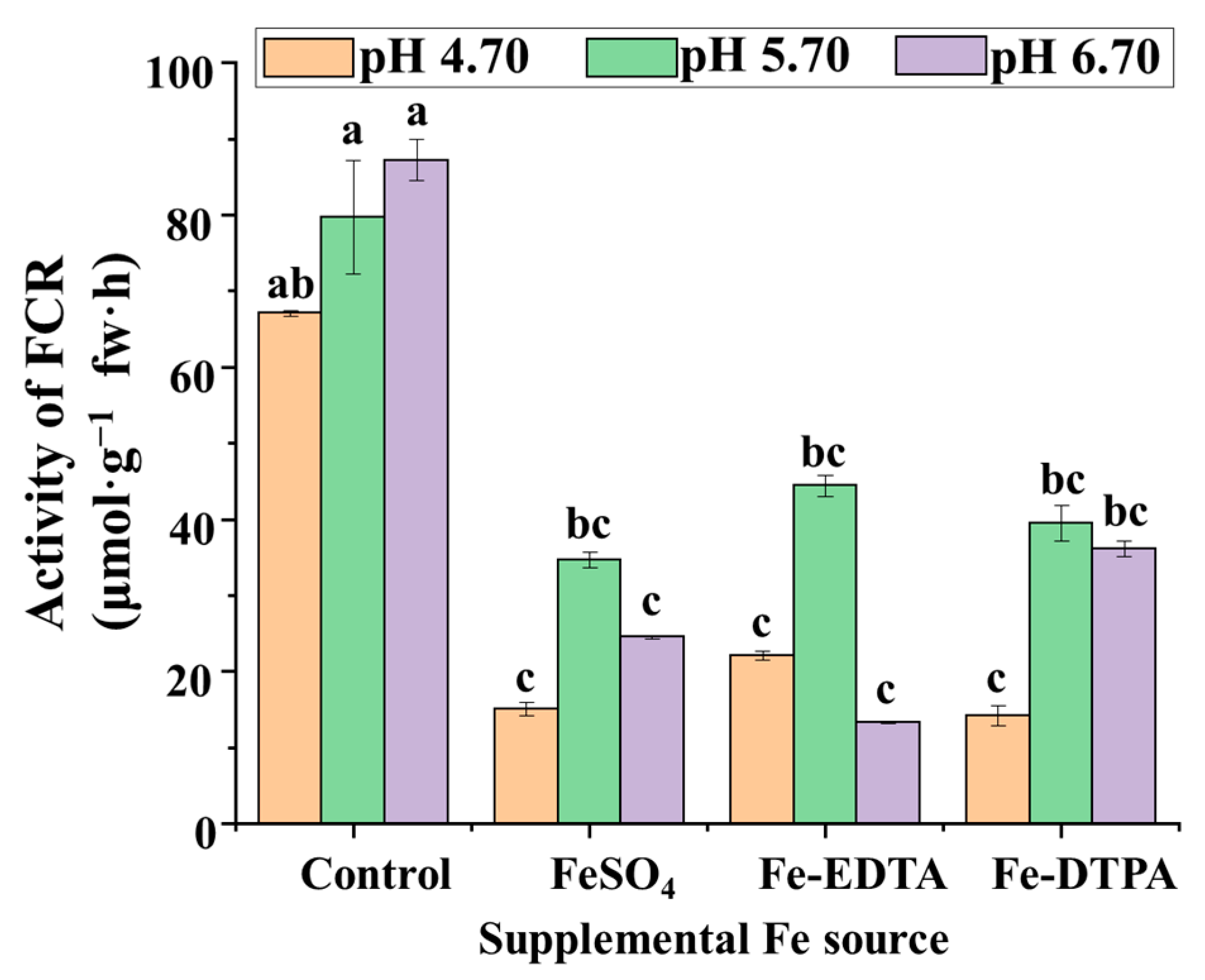
2.3. FCR Activities
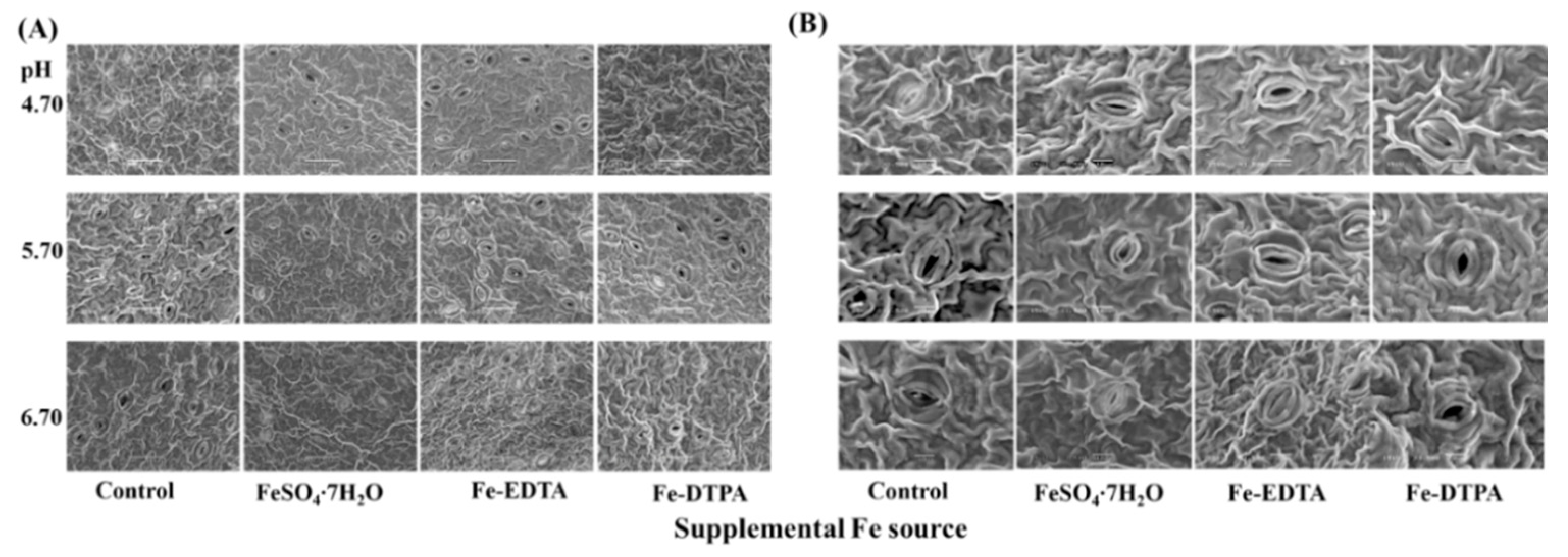
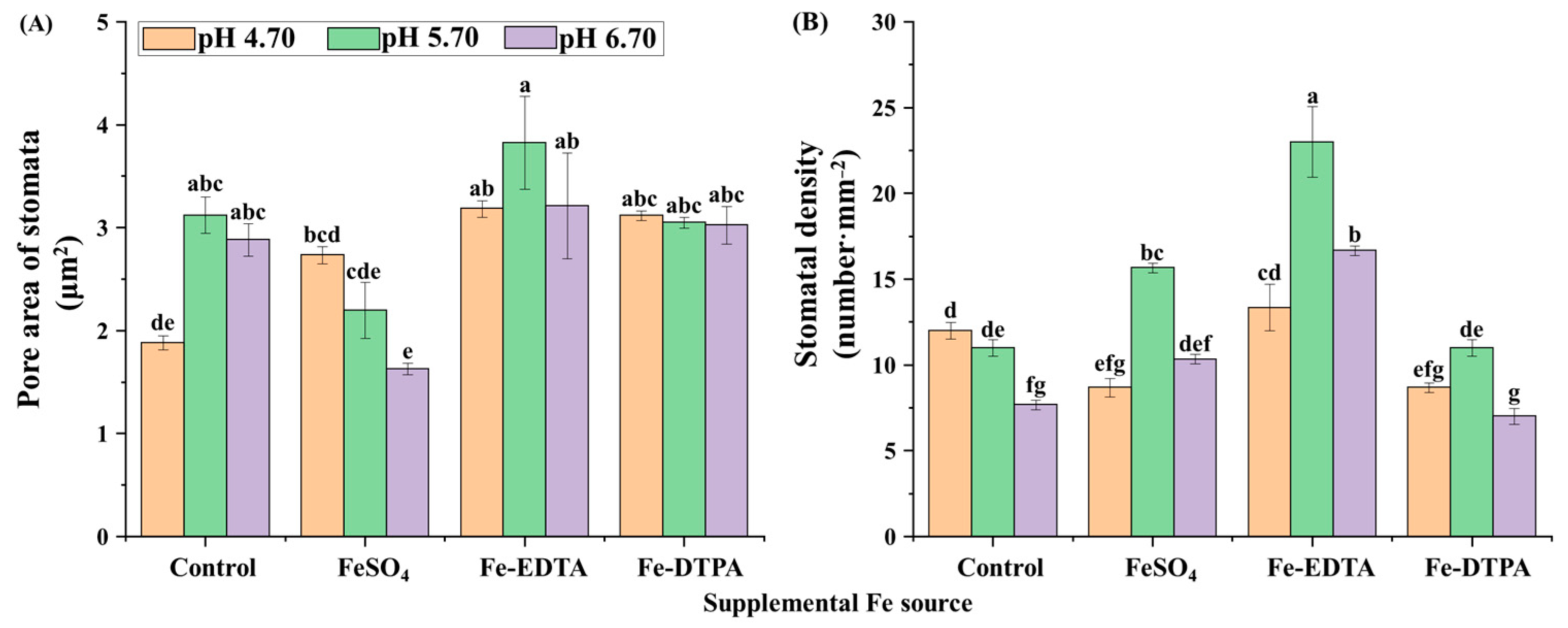
2.4. Analysis of Stomata
2.5. Macro- and Micro-Nutrients Content
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Culture Conditions
4.2. Iron and pH Treatments
4.3. Measurement of the Growth Characteristics
4.4. Chlorophyll Content
4.5. Total Soluble Proteins and Antioxidant Enzyme Activities
4.6. Activity of FCR
4.7. Scanning Electron Microscopic (SEM) Analysis of Stomata
4.8. Determination of Macro- and Micro-Nutrient Contents Using Inductively Coupled Plasma Spectrometer
4.9. Data Collection and Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huh, M.-K.; Kim, S.-H.; Park, S.-H. Phylogenetic study of genus Sorbus in Korea by internal transcribed spacer sequence (ITS). J. Life Sci. 2007, 17, 1610–1615. [Google Scholar] [CrossRef] [Green Version]
- Jeong, B.R.; Sivanesan, I. Micropropagation of Sorbus commixta HEDL. Propag. Ornamrntal Plants 2015, 15, 142–146. [Google Scholar]
- Yu, T.; Lee, Y.; Cho, J. Inhibitory effect of Sorbus commixta extract on lipopolysaccharide-induced pro-inflammatory events in macrophages. J. Med. Plants Res. 2009, 3, 600–607. [Google Scholar]
- Bae, K. The Medicinal Plants of Korea; Kyo-Hak Publishing Co.: Seoul, Korea, 2000; p. 260. [Google Scholar]
- Yagihashi, T.; Hayashida, M.; Miyamoto, T. Effects of bird ingestion on seed germination of Sorbus commixta. Oecologia 1998, 114, 209–212. [Google Scholar] [CrossRef]
- Incesu, M.; Yeşiloğlu, T.; Cimen, B.; Yilmaz, B. Influences of different iron levels on plant growth and photosynthesis of W. Murcott mandarin grafted on two rootstocks under high pH conditions. Turk. J. Agric. For. 2015, 39, 838–844. [Google Scholar] [CrossRef]
- Graham, R.D.; Welch, R.M. Plant food micronutrient composition and human nutrition. Commun. Soil Sci. Plant Anal. 2000, 31, 1627–1640. [Google Scholar] [CrossRef]
- Rajniak, J.; Giehl, R.F.; Chang, E.; Murgia, I.; von Wirén, N.; Sattely, E.S. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat. Chem. Biol. 2018, 14, 442–450. [Google Scholar] [CrossRef]
- Lindsay, W. Chemical reactions in soils that affect iron availability to plants. A quantative approach. In Iron Nutrition in Soils and Plants; Springer: Berlin/Heidelberg, Germany, 1995; pp. 7–14. [Google Scholar]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.-F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H.; Römheld, V.; Kissel, M. Different strategies in higher plants in mobilization and uptake of iron. J. Plant Nutr. 1986, 9, 695–713. [Google Scholar] [CrossRef]
- Smith, B.R.; Cheng, L. Fe-EDDHA Alleviates Chlorosis in Concord’Grapevines Grown at High pH. HortScience 2006, 41, 1498–1501. [Google Scholar] [CrossRef]
- Roosta, H.R. Interaction between water alkalinity and nutrient solution pH on the vegetative growth, chlorophyll fluorescence and leaf magnesium, iron, manganese, and zinc concentrations in lettuce. J. Plant Nutr. 2011, 34, 717–731. [Google Scholar] [CrossRef]
- Kalbasl, M.; Manuchehri, N.; Filsoof, F. Local acidification of soil as a means to alleviate iron chlorosis in quince orchards. J. Plant Nutr. 1986, 9, 1001–1007. [Google Scholar] [CrossRef]
- Hutchinson, T. Lime chlorosis as a factor in seedling establishment on calcareous soil: II. The development of leaf water deficits in plants showing lime-chlorosisi. New Phytol. 1970, 69, 143–157. [Google Scholar] [CrossRef]
- Priyadarshini, P.; Chitdeshwari, T.; Sudhalakshmi, C. A Iron Availability in Calcareous and Non Calcareous Soils as Influenced by Various Sources and Levels of Iron. Madras Agric. J. 2019, 106, 4–6. [Google Scholar] [CrossRef]
- Boamponsem, G.A.; Leung, D.W.; Lister, C. Relationships among iron deficit-induced potato callus growth inhibition, Fe distribution, chlorosis, and oxidative stress amplified by reduced antioxidative enzyme activities. Plant Cell Tissue Organ Cult. 2018, 132, 393–412. [Google Scholar] [CrossRef]
- Eddings, J.L.; Brown, A. Absorption and translocation of foliar-applied iron. Plant Physiol. 1967, 42, 15–19. [Google Scholar] [CrossRef]
- Deo, P.C.; Dugdale, B.; Harding, R.M.; Kato, M.; Dale, J. In vitro micro propagation of Nicotiana benthamiana via axillary shoots. S. Pac. J. Nat. Appl. Sci. 2014, 32, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Roosta, H.R.; Pourebrahimi, M.; Hamidpour, M. Effects of bicarbonate and different Fe sources on vegetative growth and physiological characteristics of bell pepper (Capsicum annuum L.) plants in hydroponic system. J. Plant Nutr. 2015, 38, 397–416. [Google Scholar] [CrossRef]
- Lindsay, W. Chemical Equilibria in Soils; Blackburn Press: Caldwell, NJ, USA, 1979. [Google Scholar]
- Boxma, R. Effect of pH on the behaviour of various iron chelates in sphagnum (moss) peat. Commun. Soil Sci. Plant Anal. 1981, 12, 755–763. [Google Scholar] [CrossRef]
- Kobayashi, N.I.; Ogura, T.; Takagi, K.; Sugita, R.; Suzuki, H.; Iwata, R.; Nakanishi, T.M.; Tanoi, K. Magnesium deficiency damages the youngest mature leaf in rice through tissue-specific iron toxicity. Plant Soil 2018, 428, 137–152. [Google Scholar] [CrossRef]
- Barton, L.L.; Abadia, J. Iron Nutrition in Plants and Rhizospheric Microorganisms; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Terry, N.; Low, G. Leaf chlorophyll content and its relation to the intracellular localization of iron. J. Plant Nutr. 1982, 5, 301–310. [Google Scholar] [CrossRef]
- Bolle-Jones, E.; Notton, B. The relative proportions of the chloroplast pigments as influenced by different levels of iron and potassium supply. Plant Soil 1953, 5, 87–100. [Google Scholar] [CrossRef]
- Morales, F.; Abadía, A.; Abadía, J. Characterization of the xanthophyll cycle and other photosynthetic pigment changes induced by iron deficiency in sugar beet (Beta vulgaris L.). Plant Physiol. 1990, 94, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lysenko, E.A.; Klaus, A.A.; Kartashov, A.V.; Kusnetsov, V.V. Specificity of Cd, Cu, and Fe effects on barley growth, metal contents in leaves and chloroplasts, and activities of photosystem I and photosystem II. Plant Physiol. Biochem. 2020, 147, 191–204. [Google Scholar] [CrossRef]
- Nemati Lafmejani, Z.; Jafari, A.A.; Moradi, P.; Ladan Moghadam, A. Impact of foliar application of iron-chelate and Iron nano Particles on some morpho-physiological traits and rssential oil composition of peppermint (Mentha piperita L.). J. Essent. Oil Bear. Plants 2018, 21, 1374–1384. [Google Scholar] [CrossRef]
- Mann, A.; Singh, A.; Oza, S.; Goswami, N.; Mehta, D.; Chaudhari, V. Effect of iron source on iron deficiency induced chlorosis in groundnut. Legume Res. Int. J. 2017, 40, 241–249. [Google Scholar] [CrossRef]
- Albano, J.P.; Miller, W.B.; Halbrooks, M.C. Iron toxicity stress causes bronze speckle, a specific physiological disorder of marigold (Tagetes erecta L.). J. Am. Soc. Hortic. Sci. 1996, 121, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Dobermann, A. Rice: Nutrient Disorders & Nutrient Management; International Rice Research Institute: Los Banos, Philippines, 2000. [Google Scholar]
- Sahrawat, K.Á. Iron toxicity in wetland rice and the role of other nutrients. J. Plant Nutr. 2005, 27, 1471–1504. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Polle, A. Dissecting the superoxide dismutase-ascorbate-glutathione-pathway in chloroplasts by metabolic modeling. Computer simulations as a step towards flux analysis. Plant Physiol. 2001, 126, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Bowler, C.; Slooten, L.; Vandenbranden, S.; De Rycke, R.; Botterman, J.; Sybesma, C.; Van Montagu, M.; Inzé, D. Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J. 1991, 10, 1723–1732. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-G.; Jung, S. Differential antioxidant mechanisms of rice plants in response to oxyfluorfen and paraquat. Weed Turfgrass Sci. 2013, 2, 254–259. [Google Scholar] [CrossRef]
- Molassiotis, A.; Tanou, G.; Diamantidis, G.; Patakas, A.; Therios, I. Effects of 4-month Fe deficiency exposure on Fe reduction mechanism, photosynthetic gas exchange, chlorophyll fluorescence and antioxidant defense in two peach rootstocks differing in Fe deficiency tolerance. J. Plant Physiol. 2006, 163, 176–185. [Google Scholar] [CrossRef]
- Hell, R.; Stephan, U.W. Iron uptake, trafficking and homeostasis in plants. Planta 2003, 216, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Zouari, M.; Gogorcena, Y.; Lucena, J.J.; Abadía, J. Effects of cadmium and lead on ferric chelate reductase activities in sugar beet roots. Plant Physiol. Biochem. 2003, 41, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-R.; Oh, M.-M.; Park, S.-A. Ferric-chelate reductase activity is a limiting factor in iron uptake in spinach and kale roots. Hortic. Environ. Biotechnol. 2016, 57, 462–469. [Google Scholar] [CrossRef]
- Pestana, M.; Gama, F.; Saavedra, T.; De Varennes, A.; Correia, P. The root ferric-chelate reductase of Ceratonia siliqua (L.) and Poncirus trifoliata (L.) Raf. responds differently to a low level of iron. Sci. Hortic. 2012, 135, 65–67. [Google Scholar] [CrossRef]
- Shi, G.; Sun, L.; Wang, X.; Liu, C. Leaf responses to iron nutrition and low cadmium in peanut: Anatomical properties in relation to gas exchange. Plant Soil 2014, 375, 99–111. [Google Scholar] [CrossRef]
- Soundararajan, P.; Sivanesan, I.; Jo, E.H.; Jeong, B.R. Silicon promotes shoot proliferation and shoot growth of Salvia splendens under salt stress in vitro. Hortic. Environ. Biotechnol. 2013, 54, 311–318. [Google Scholar] [CrossRef]
- Silva, A.L.E.; de Lima Terceiro, L.E.; de Lima, M.F.; Costa-Silva, R.; Dos Santos, E.A.; Agra, M.d.F. Leaf and stem micromorphology of Byrsonima sericea DC. by light and scanning electron microscopy. Microsc. Res. Tech. 2020, 83, 287–296. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hao, W.; Manivannan, A.; Yuze, C.; Jeong, B.R. Effect of Different Cultivation Systems on the Accumulation of Nutrients and Phytochemicals in Ligularia fischeri. Hortic. Plant J. 2018, 4, 24–29. [Google Scholar]
- Garnier, E.; Laurent, G. Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species. New Phytol. 1994, 128, 725–736. [Google Scholar] [CrossRef]
- González, L.; González-Vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Springer: Berlin/Heidelberg, Germany, 2001; pp. 207–212. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soundararajan, P.; Manivannan, A.; Ko, C.H.; Park, J.E.; Jeong, B.R. Evaluation of relative toxicity caused by deicing agents on photosynthesis, redox homeostasis, and the osmoregulatory system in creeper-type plants. Hortic. Environ. Biotechnol. 2019, 60, 175–186. [Google Scholar] [CrossRef]
- Soundararajan, P.; Manivannan, A.; Park, Y.G.; Muneer, S.; Jeong, B.R. Silicon alleviates salt stress by modulating antioxidant enzyme activities in Dianthus caryophyllus ‘Tula’. Hortic. Environ. Biotechnol. 2015, 56, 233–239. [Google Scholar] [CrossRef]
- Bienfait, H.F.; van den Briel, W.; Mesland-Mul, N.T. Free space iron pools in roots: Generation and mobilization. Plant Physiol. 1985, 78, 596–600. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Yu, Q.; Xu, C.; Li, J.; Qin, G. Rapid estimation of stomatal density and stomatal area of plant leaves based on object-oriented classification and its ecological trade-off strategy analysis. Forests 2018, 9, 616. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Dotson, P. The use of microwave muffle furnace for dry ashing plant tissue samples. Commun. Soil Sci. Plant Anal. 1994, 25, 1321–1327. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fe Source (A) | pH (B) | Number of Leaves | Number of Roots | Shoot Length (cm) | Root Length (cm) | Stem Length (cm) | Stem Diameter (mm) | Leaf Area (cm2) | Fresh Weight (mg) | Dry Weight (mg) | Relative Water Content (%) | Water Content (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |||||||||||
| Control | 4.70 | 4 b–d z | 1 cd | 2.3 b–e | 2.3 cd | 1.7 a | 2.24 ab | 2.99 de | 119.0 b–e | 17.2 bc | 33.6 bc | 2.7 bc | 63.70 a | 75.51 a |
| 5.70 | 2 d | 1 cd | 2.3 b–e | 2.2 cd | 1.5 a | 2.08 ab | 2.60 de | 157.7 b–d | 14.9 bc | 40.8 bc | 1.3 c | 79.36 a | 67.14 a | |
| 6.70 | 2 d | 0 d | 1.4 e | 0.0 d | 0.7 b | 1.63 ab | 1.88 e | 55.4 e | 0.0 c | 22.0 c | 0.0 c | 63.94 a | 64.73 a | |
| FeSO4 | 4.70 | 6 ab | 1 cd | 3.1 a–c | 2.8 cd | 1.5 a | 2.07 ab | 5.79 b–e | 182.1 bc | 51.7 bc | 47.9 ab | 1.7 c | 57.31 a | 74.79 a |
| 5.70 | 7 a | 4 ab | 3.5 ab | 10.6 a | 1.4 a | 2.34 a | 8.74 ab | 297.3 a | 102.3 b | 65.1 a | 12.0 b | 66.01 a | 75.87 a | |
| 6.70 | 5 a–c | 2 b–d | 3.1 a–c | 6.7 a–c | 1.7 a | 1.74 ab | 8.43 a–c | 197.7 b | 72.2 bc | 32.6 bc | 7.8 bc | 92.28 a | 77.31 a | |
| Fe-EDTA | 4.70 | 5 a–c | 2 b–d | 3.3 ab | 6.3 a–c | 1.5 a | 1.99 ab | 6.80 a–d | 100.4 c–e | 90.1 bc | 27.5 bc | 7.8 bc | 55.81 a | 61.81 a |
| 5.70 | 5 a–c | 4 a | 3.9 a | 8.1 ab | 1.3 a | 2.28 a | 10.54 a | 296.1 a | 225.5 a | 71.6 a | 19.4 a | 65.34 a | 75.58 a | |
| 6.70 | 4 b–d | 2 cd | 3.0 a–d | 2.9 cd | 1.3 a | 1.63 ab | 5.00 b–e | 120.0 b–e | 42.7 bc | 33.2 bc | 4.5 bc | 62.53 a | 72.42 a | |
| Fe-DTPA | 4.70 | 2 d | 0 d | 1.5 de | 0.0 d | 1.5 a | 1.19 b | 2.22 e | 70.9 de | 0.0 c | 24.5 bc | 0.0 c | 66.09 a | 62.37 a |
| 5.70 | 3 dc | 2 dc | 1.7 c–e | 0.9 d | 1.5 a | 1.80 ab | 2.48 e | 104.7 b–e | 4.5 bc | 37.2 bc | 1.0 c | 73.42 a | 60.62 a | |
| 6.70 | 4 b–d | 3 a–c | 2.6 a–e | 4.5 b–d | 1.4 a | 1.74 ab | 4.40 c–e | 103.7 b–e | 49.0 bc | 31.1 bc | 5.3 bc | 56.50 a | 73.99 a | |
| F-test y | A | *** | ** | *** | *** | NS | NS | *** | *** | *** | * | *** | NS | NS |
| B | ** | *** | NS | NS | NS | NS | NS | *** | NS | ** | NS | NS | NS | |
| A × B | *** | *** | NS | ** | NS | NS | NS | * | ** | * | ** | NS | NS | |
| Fe Source (A) | pH (B) | L* | a* | b* |
|---|---|---|---|---|
| Control | 4.70 | 72 b z | −21 d | 51 bc |
| 5.70 | 50 c | −14 c | 28 fg | |
| 6.70 | 22 fg | −24 a | 27 g | |
| FeSO4 | 4.70 | 35 e | −31 f | 34 de |
| 5.70 | 27 fg | −27 ef | 33 ef | |
| 6.70 | 80 a | −14 c | 60 a | |
| Fe-EDTA | 4.70 | 39 de | −25 e | 39 d |
| 5.70 | 42 de | −12 c | 25 gh | |
| 6.70 | 73 ab | −13 c | 52 b | |
| Fe-DTPA | 4.70 | 19 g | 9 b | 14 i |
| 5.70 | 23 fg | 12 b | 20 h | |
| 6.70 | 46 cd | −27 ef | 46 c | |
| F-test y | A | *** | *** | *** |
| B | *** | *** | *** | |
| A × B | *** | *** | *** |
| Fe Source (A) | pH (B) | K | Ca | Mg | Zn | Mn | Fe | Cu | B | Si | P | S |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 4.70 | 279.27 a z | 132.20 b | 41.59 b | 1.16 b | 1.08 b | 1.75 i | 0.29 a | 0.32 e | 1.16 e | 117.53 b | 28.99 c |
| 5.70 | 212.03 d | 120.27 c | 31.17 f | 1.03 c | 0.68 f | 1.78 i | 0.23 b | 0.37 b | 1.62 c | 102.43 c | 23.24 f | |
| 6.70 | 159.40 h | 100.00 f | 27.57 i | 0.98 e | 0.52 h | 1.55 j | 0.18 cd | 0.34 d | 1.00 f | 92.43 e | 18.03 h | |
| FeSO4 | 4.70 | 252.00 b | 113.33 d | 32.47 e | 1.01 d | 0.89 d | 4.44 e | 0.28 a | 0.35 cd | 1.43 d | 99.98 cd | 25.96 d |
| 5.70 | 234.47 c | 94.24 g | 27.72 i | 1.37 a | 1.35 a | 3.54 f | 0.17 e | 0.61 a | 2.61 a | 77.56 gh | 30.36 b | |
| 6.70 | 189.40 f | 87.51 h | 29.69 g | 0.63 k | 0.58 g | 2.06 h | 0.17 e | 0.26 h | 2.55 b | 76.66 h | 18.59 h | |
| Fe-EDTA | 4.70 | 236.93 c | 99.46 f | 34.84 d | 0.82 g | 0.83 e | 6.62 d | 0.19 c | 0.27 g | 0.91 g | 101.70 c | 24.88 e |
| 5.70 | 278.60 a | 145.27 a | 46.27 a | 0.89 f | 1.02 c | 3.00 g | 0.17 e | 0.37 b | 0.77 i | 125.10 a | 31.56 a | |
| 6.70 | 214.20 d | 130.20 b | 36.78 c | 0.71 i | 1.10 b | 1.86 i | 0.13 f | 0.35 c | 0.82 h | 101.13 c | 31.51 a | |
| Fe-DTPA | 4.70 | 176.97 g | 77.17 j | 20.41 k | 0.62 k | 0.54 h | 18.61 a | 0.18 de | 0.19 i | 0.69 j | 79.62 g | 21.11 g |
| 5.70 | 202.47 e | 104.77 e | 28.56 h | 0.67 j | 0.69 f | 13.69 b | 0.13 f | 0.30 f | 0.56 k | 88.72 f | 25.06 e | |
| 6.70 | 186.27 f | 83.03 i | 24.21 j | 0.80 h | 0.52 h | 9.80 c | 0.10 g | 0.25 h | 0.92 g | 97.73 d | 18.19 h | |
| F-test y | A | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| B | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| A × B | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, J.; Park, Y.G.; Guo, G.; Jeong, B.R. Effect of Iron Source and Medium pH on Growth and Development of Sorbus commixta In Vitro. Int. J. Mol. Sci. 2021, 22, 133. https://doi.org/10.3390/ijms22010133
Xiao J, Park YG, Guo G, Jeong BR. Effect of Iron Source and Medium pH on Growth and Development of Sorbus commixta In Vitro. International Journal of Molecular Sciences. 2021; 22(1):133. https://doi.org/10.3390/ijms22010133
Chicago/Turabian StyleXiao, Jie, Yoo Gyeong Park, Ge Guo, and Byoung Ryong Jeong. 2021. "Effect of Iron Source and Medium pH on Growth and Development of Sorbus commixta In Vitro" International Journal of Molecular Sciences 22, no. 1: 133. https://doi.org/10.3390/ijms22010133
APA StyleXiao, J., Park, Y. G., Guo, G., & Jeong, B. R. (2021). Effect of Iron Source and Medium pH on Growth and Development of Sorbus commixta In Vitro. International Journal of Molecular Sciences, 22(1), 133. https://doi.org/10.3390/ijms22010133