
Functional and Structural Changes in the Corticospinal Tract of Streptozotocin-Induced Diabetic Rats


Abstract
:1. Introduction
2. Results
2.1. Model Animals
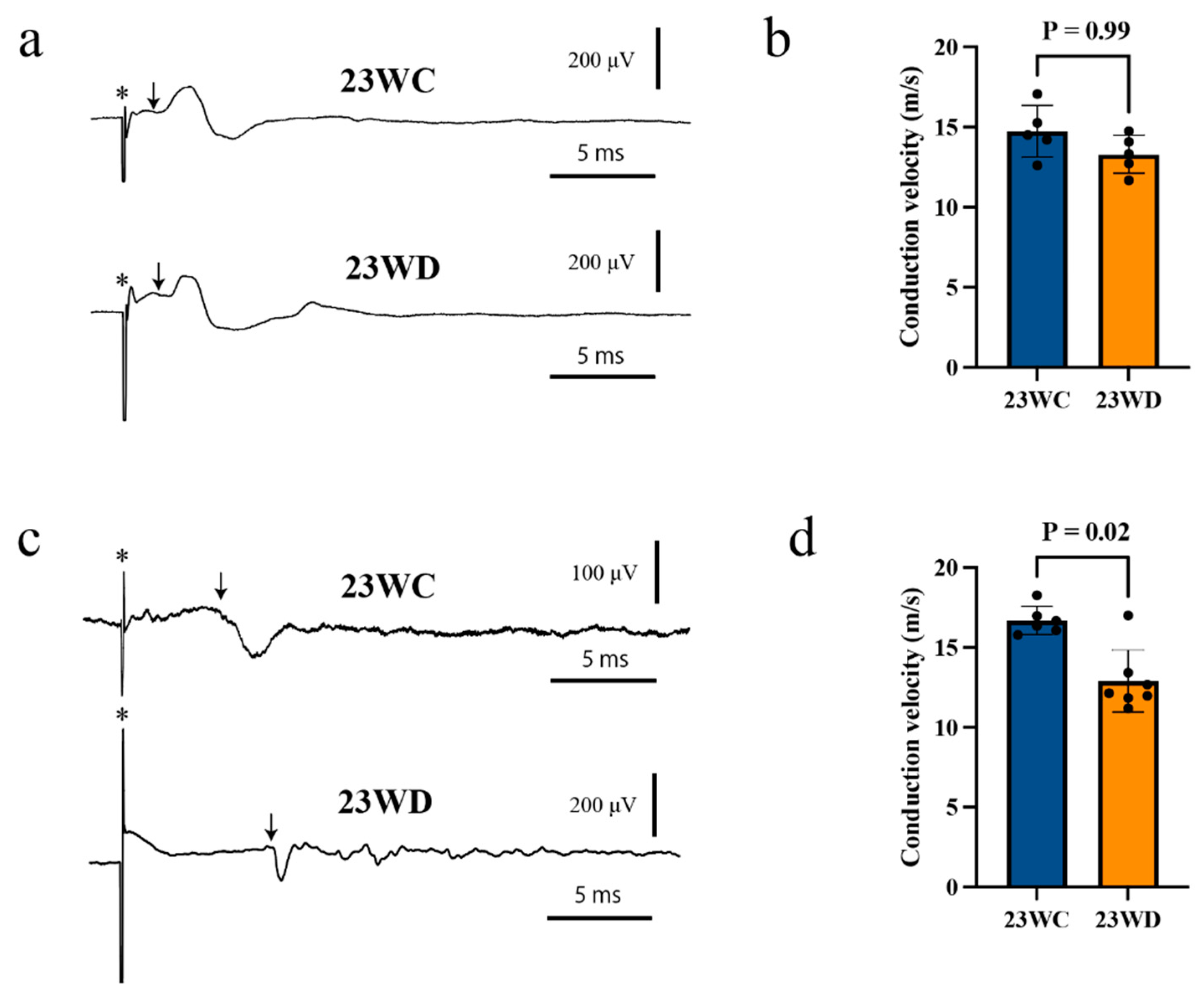
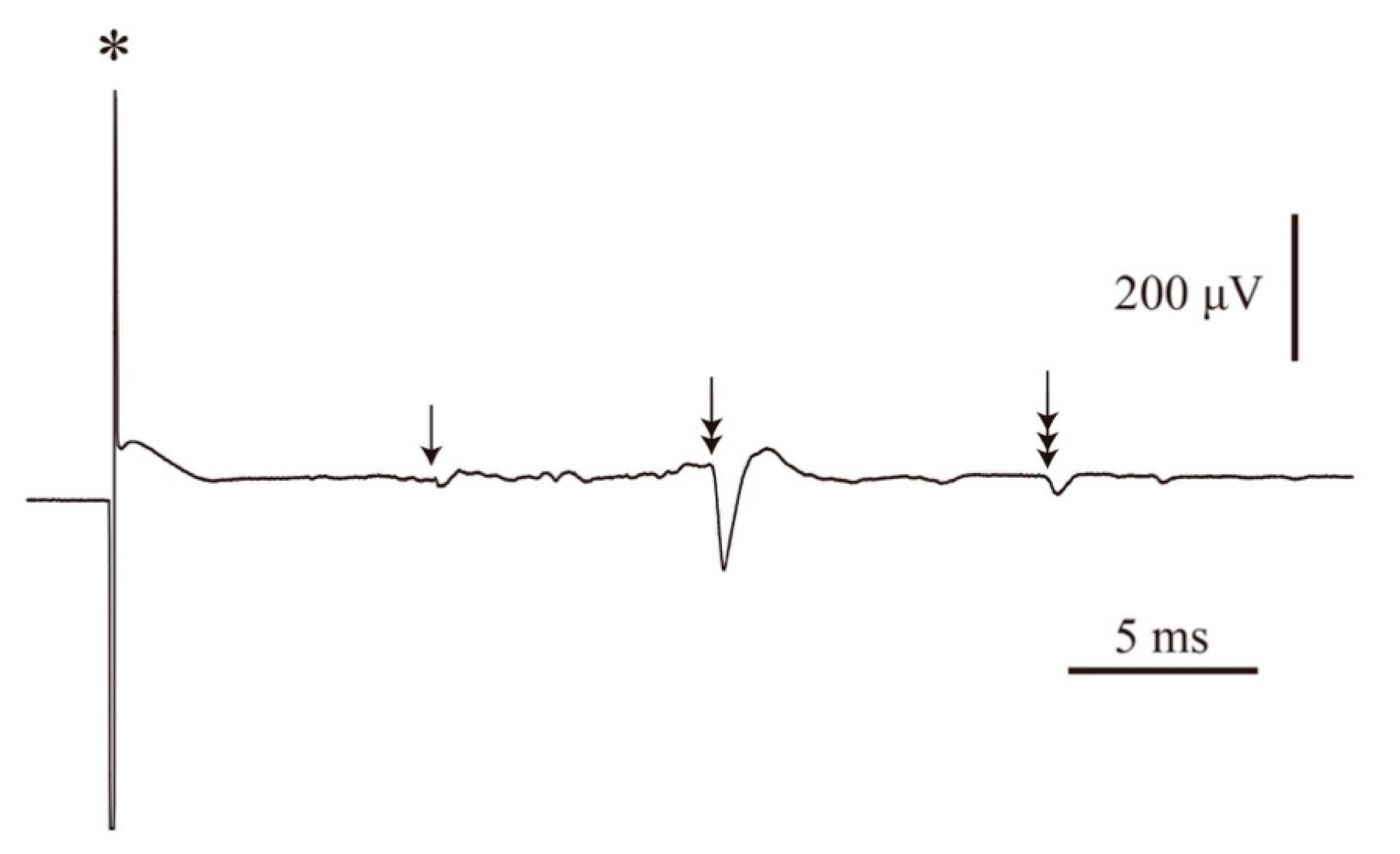
2.2. Conduction Velocities of the CST and Motor Nerve
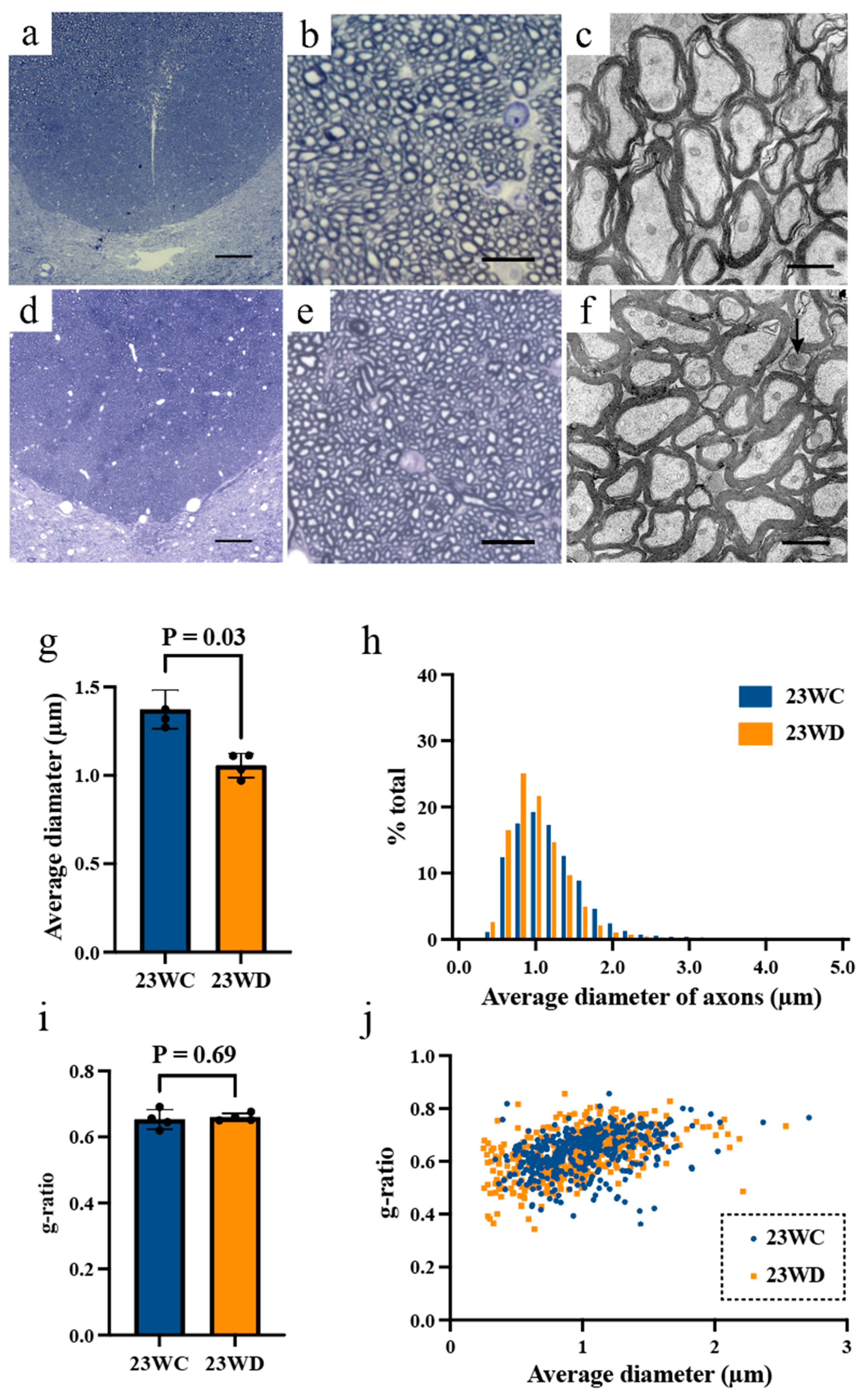
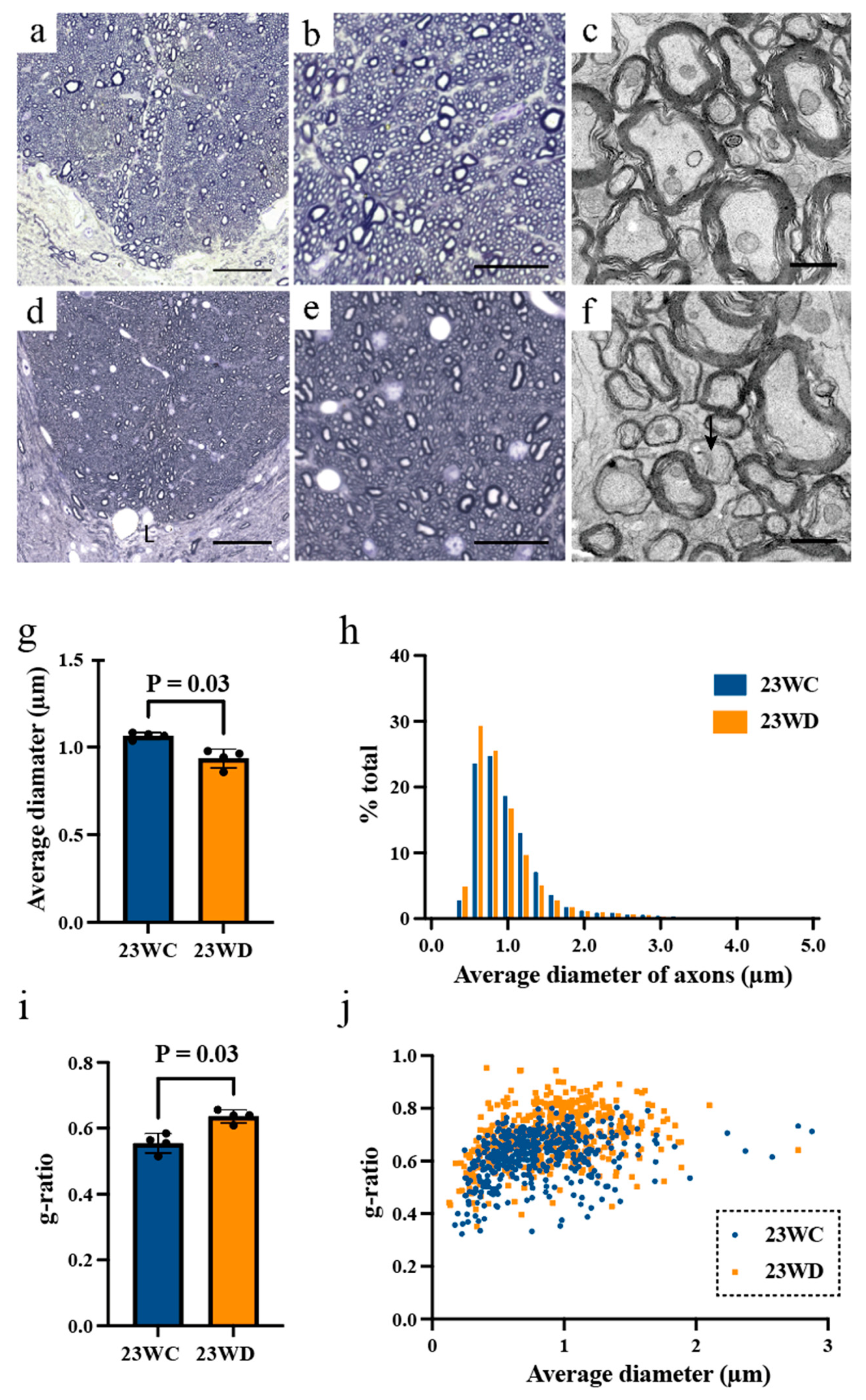
2.3. Morphological Analysis of Tract Fibers at the Base of the Dorsal Funiculus
3. Discussion
4. Materials and Methods
4.1. Induction of Experimental Diabetes (Diabetic Animal Model)
4.2. Measurement of the Conduction Velocity of CST Fibers and Motor Nerve
4.3. Morphological Analysis
4.4. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Maurer, M.S.; Burcham, J.; Cheng, H. Diabetes mellitus is associated with an increased risk of falls in elderly residents of a long-term care facility. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1157–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijzerman, T.H.; Schaper, N.C.; Melai, T.; Meijie, K.; Willems, P.J.B.; Savelberg, H.H.C.M. Lower extremity muscle strength is reduced in people with type 2 diabetes, with and without polyneuropathy, and is associated with impaired mobility and reduced quality of life. Diabetes Res. Clin. Pract. 2012, 95, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allet, L.; Armand, S.; Golay, A.; Monnin, D.; Bie, R.A.; Burin, E.D. Gait characteristics of diabetic patients: A systematic review. Diabetes Metab. Res. Reve. 2008, 24, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.J.; Handsaker, J.C.; Bowling, F.L.; Boulton, A.J.M.; Reeves, N.D. Diabetic peripheral neuropathy compromises balance during daily activities. Diabetes Care 2015, 38, 1116–1122. [Google Scholar] [CrossRef] [Green Version]
- Andersen, H. Motor function in diabetic neuropathy. Acta Neurol. Scand. 1999, 100, 211–220. [Google Scholar] [CrossRef]
- Ferris, J.K.; Inglis, J.T.; Madden, K.M.; Boyd, L.A. Brain and Body: A Review of Central Nervous System Contributions to Movement Impairments in Diabetes. Diabetes 2020, 69, 3–11. [Google Scholar] [CrossRef]
- Muramatsu, K. Diabetes Mellitus-Related Dysfunction of the Motor System. Int. J. Mol. Sci. 2020, 21, 7485. [Google Scholar] [CrossRef]
- Muramatsu, K.; Ikutomo, M.; Tamaki, T.; Shimo, S.; Niwa, M. Effect of streptozotocin-induced diabetes on motor representations in the motor cortex and corticospinal tract in rats. Brain Res. 2018, 1680, 115–126. [Google Scholar] [CrossRef]
- Madsen, J.G.; Østergaard, J.A.; Andersen, H.; Pedersen, M. Attenuation of Cortically Evoked Motor-Neuron Potential in Streptozotocin-Induced Diabetic Rats: A Study about the Effect of Diabetes upon Cortical-Initiated Movement. Biomed. Res. Int. 2020, 2020, 1942534. [Google Scholar] [CrossRef] [Green Version]
- van Duinkerken, E.; Schoonheim, M.M.; IJzerman, R.G.; Klein, M.; Ryan, C.M.; Moll, A.C.; Snoek, F.J.; Barkhof, F.; Diamant, M.; Pouwels, P.J.W. Diffusion tensor imaging in type 1 diabetes: Decreased white matter integrity relates to cognitive functions. Diabetologia 2012, 55, 1218–1220. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Sui, Y.; Xu, Z.; Zhang, Q.; Karaman, M.M.; Cai, K.; Anderson, T.M.; Zhu, W.; Wang, J.; Zhou, X.J. Diffusion Tensor Imaging Study on White Matter Abnormalities in Patients with Type 2 Diabetes Using Tract-Based Spatial Statistics. AJNR Am. J. Neuroradiol. 2016, 37, 1462–1469. [Google Scholar] [CrossRef] [Green Version]
- Van Bloemendaal, L.; Ijzerman, R.G.; Jennifer, S.; Barkhof, F.; Diamant, M.; Veltman, D.J.; van Duinkerken, E. Alterations in white matter volume and integrity in obesity and type 2 diabetes. Metab. Brain Dis. 2016, 31, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Abbruzzese, G.; Schenone, A.; Scramuzza, G.; Caponnetto, C.; Gasparetto, B.; Adezati, L.; Abbruzzese, M.; Viviani, G.L. Impairment of central motor conduction in diabetic patients. Electroencephalogr. Clin. Neurophysiol. 1993, 89, 335–340. [Google Scholar] [CrossRef]
- Dolu, H.; Ulas, U.H.; Bolu, E.; Ozkardes, A.; Odabasi, Z.; Ozata, M.; Vural, O. Evaluation of central neuropathy in type II diabetes mellitus by multimodal evoked potentials. Acta Neurologica Belgica 2003, 103, 206–211. [Google Scholar] [PubMed]
- Goldenberg, Z.; Kucera, P.; Brezinova, M.; Kurca, E.; Barak, L.; Traubner, P. Clinically unapparent central motor pathways lesion in patients with type I diabetes mellitus. A transcranial magnetic stimulation study. Bratisl Lek Listy 2004, 105, 400–403. [Google Scholar] [PubMed]
- Lemon, R.N. Descending pathways in motor control. Annu. Rev. Neurosci. 2008, 31, 195–218. [Google Scholar] [CrossRef] [Green Version]
- Saliani, A.; Perraud, B.; Duval, T.; Stikov, N.; Rossignol, S.; Cohen-Adad, J. Axon and Myelin Morphology in Animal and Human Spinal Cord. Front. Neuroanat. 2017, 11, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brösamle, C.; SAMLE, C.; Schwab, M.E. Cells of origin, course, and termination patterns of the ventral, uncrossed component of the mature rat corticospinal tract. J. Comp. Neurol. 1997, 386, 293–303. [Google Scholar] [CrossRef]
- Kimura, J.; Yamada, T.; Nelson, P.S. Distal slowing of motor nerve conduction velocity in diabetic polyneuropathy. J. Neurol. Sci. 1979, 42, 291–302. [Google Scholar] [CrossRef]
- Mediratta, N.K.; Nicoll, J.A. Conduction velocities of corticospinal axons in the rat studied by recording cortical antidromic responses. J. Physiol. 1983, 336, 545–561. [Google Scholar] [CrossRef] [Green Version]
- Chomiak, T.; Hu, B. What is the optimal value of the g-ratio for myelinated fibers in the rat CNS? A theoretical approach. PLoS ONE 2009, 4, e7754. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.Y.; Bujalka, H.; Emery, B. Axoglial interactions in myelin plasticity: Evaluating the relationship between neuronal activity and oligodendrocyte dynamics. Glia 2019, 67, 2038–2049. [Google Scholar] [CrossRef]
- Muramatsu, K.; Niwa, M.; Nagai, M.; Kamimura, T.; Sasaki, S.-I.; Ishiguro, T. The size of motoneurons of the gastrocnemius muscle in rats with diabetes. Neurosci. Lett. 2012, 531, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, K.; Niwa, M.; Tamaki, T.; Ikutomo, M.; Masu, Y.; Hasegawa, T.; Shimo, S.; Sasaki, S.-I. Effect of streptozotocin-induced diabetes on motoneurons and muscle spindles in rats. Neurosci. Res. 2017, 115, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sathornsumetee, S.; McGavern, D.B.; Ure, D.R.; Rodriguez, M. Quantitative ultrastructural analysis of a single spinal cord demyelinated lesion predicts total lesion load, axonal loss, and neurological dysfunction in a murine model of multiple sclerosis. Am. J. Pathol. 2000, 157, 1365–1376. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.J.; Bennett, B.J. Topographic and quantitative description of rat dorsal column fibres arising from the lumbar dorsal roots. J. Anat. 1987, 153, 203–215. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 23WC (n = 6) | 23WD (n = 7) | p-Value | |
|---|---|---|---|
| Body weight (g) | 449.7 ± 16.6 | 282.0 ± 6.0 | <0.0001 |
| Blood sugar (mg/dL) | 152.7 ± 16.3 | 465.4 ± 46.2 | <0.0001 |
| Conduction velocity of the CST (m/s) | |||
| Stimulation: Cervical | 14.7 ± 1.6 | 14.3 ± 1.2 | 0.99 |
| Stimulation: Lumbar | 16.7 ± 0. 9 | 12.9 ± 1.9 | 0.02 |
| MNCVs (m/s) | 56.3 ± 5.7 | 39.9 ± 8.2 | <0.01 |
| 23WC (n = 4) | 23WD (n = 4) | p-Value | |
|---|---|---|---|
| Body weight (g) | 449.8 ± 24.5 | 277.5 ± 8.2 | <0.0001 |
| Blood sugar (mg/dL) | 145.3 ± 13.6 | 463.1 ± 45.3 | <0.0001 |
| Average diameter of axon (μm) | |||
| Cervical | 1.37 ± 0.11 | 1.06 ± 0.07 | 0.03 |
| Lumbar | 1.07 ± 0.02 | 0.94 ± 0.05 | 0.03 |
| Cross-sectional area of axon (μm2) | |||
| Cervical | 1.12 ± 0.20 | 0.90 ± 0.09 | 0.06 |
| Lumbar | 1.04 ± 0.04 | 0.78 ± 0.11 | 0.03 |
| Perimeter of axon (μm) | |||
| Cervical | 4.38 ± 0.51 | 3.78 ± 0.26 | 0.06 |
| Lumbar | 3.96 ± 0.20 | 3.31 ± 0.28 | 0.03 |
| G-ratio | |||
| Cervical | 0.65 ± 0.03 | 0.66 ± 0.01 | 0.69 |
| Lumbar | 0.62 ± 0.05 | 0.72 ± 0.03 | 0.03 |
| Myelin thickness (μm) | |||
| Cervical | 0.54 ± 0.05 | 0.38 ± 0.01 | 0.03 |
| Lumbar | 0.42 ± 0.03 | 0.33 ± 0.04 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muramatsu, K.; Shimo, S.; Tamaki, T.; Ikutomo, M.; Niwa, M. Functional and Structural Changes in the Corticospinal Tract of Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2021, 22, 10123. https://doi.org/10.3390/ijms221810123
Muramatsu K, Shimo S, Tamaki T, Ikutomo M, Niwa M. Functional and Structural Changes in the Corticospinal Tract of Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences. 2021; 22(18):10123. https://doi.org/10.3390/ijms221810123
Chicago/Turabian StyleMuramatsu, Ken, Satoshi Shimo, Toru Tamaki, Masako Ikutomo, and Masatoshi Niwa. 2021. "Functional and Structural Changes in the Corticospinal Tract of Streptozotocin-Induced Diabetic Rats" International Journal of Molecular Sciences 22, no. 18: 10123. https://doi.org/10.3390/ijms221810123
APA StyleMuramatsu, K., Shimo, S., Tamaki, T., Ikutomo, M., & Niwa, M. (2021). Functional and Structural Changes in the Corticospinal Tract of Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences, 22(18), 10123. https://doi.org/10.3390/ijms221810123