
The Anti-Atherosclerotic Action of FFAR4 Agonist TUG-891 in ApoE–Knockout Mice Is Associated with Increased Macrophage Polarization towards M2 Phenotype

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Body Weight and Plasma Levels of Lipids
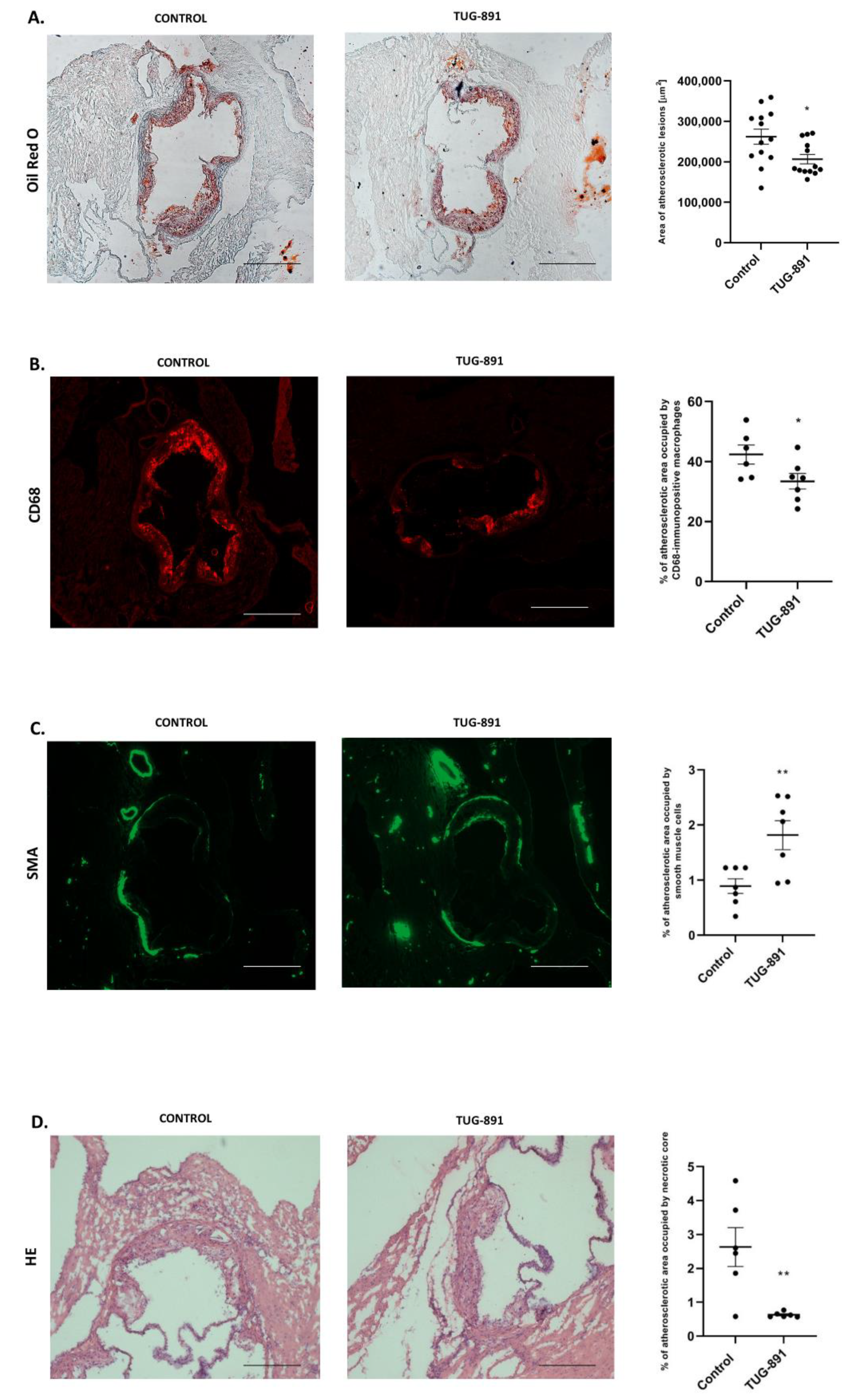
2.2. Effects of TUG-891 on Atherosclerosis
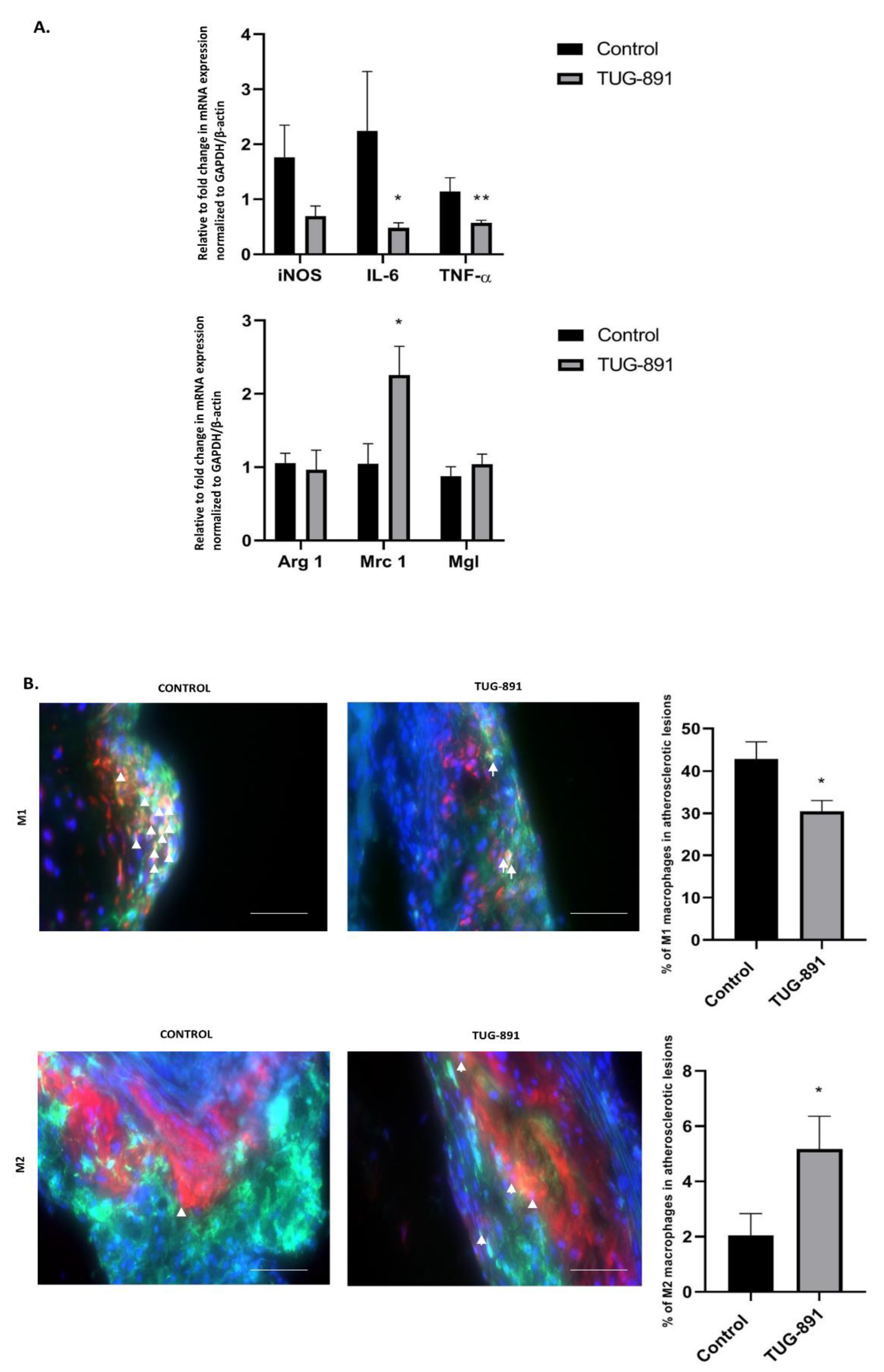
2.3. Effects of TUG-891 on In-Plaque and Peritoneal Macrophage Phenotype
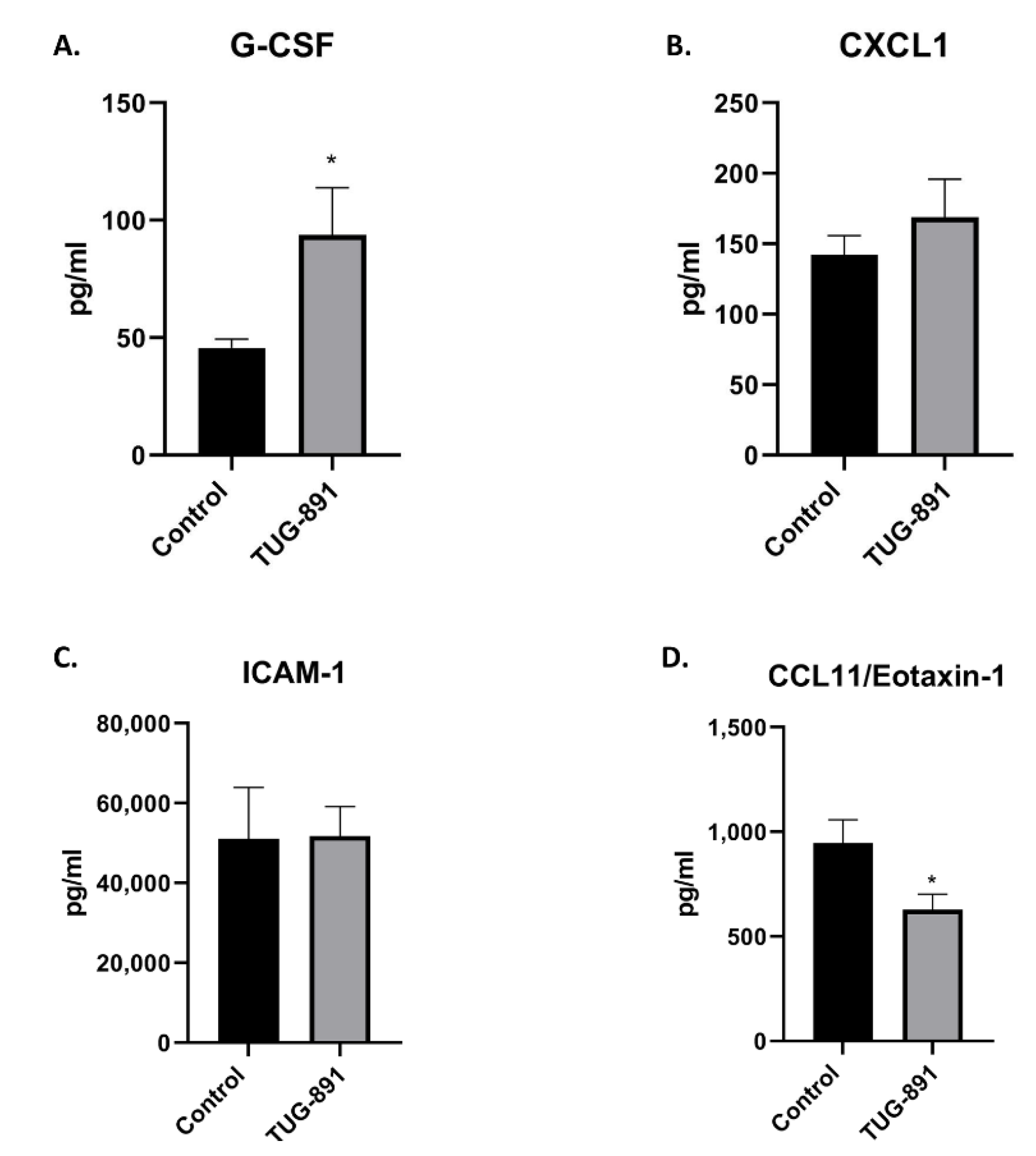
2.4. Effects of TUG-891 on Plasma Levels of Inflammatory Markers
3. Discussion
Limitations and Future Directions
4. Materials and Methods
4.1. Animal Experiments
4.2. Analysis of Atherosclerotic Plaque
4.3. Real Time (RT)-PCR
4.4. Biochemical Methods
4.5. Flow Cytometry
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AH-7614 | AH-7614-4-Methyl-N-9H-xanthen-9-yl-benzenesulfonamide |
| DAPI | 4′:6-diamidino-2-phenylindole |
| DMSO | dimethyl sulfoxide |
| FBS | fetal bovine serum |
| FFAR4/GPR120 | Free Fatty Acid Receptor 4 |
| GADP | glyceraldehyde 3-phosphate dehydrogenase |
| HDL | high-density lipoproteins |
| HE | hematoxylin/eosin |
| iCAM-1 | intercellular adhesion molecule 1 |
| IL-1β | interleukin 1 beta |
| iNOS | inducible nitric oxide synthase |
| M1 | classically activated macrophages |
| M2 | alternatively activated macrophages |
| RT-PCR | reverse transcription polymerase chain reaction |
| SMA | smooth muscle actin |
| TC | total cholesterol |
| TG | triglyceride |
| TNF-α | tumor necrosis factor α |
| TUG-891 | 4-[(4-Fluoro-4′-methyl[1,1′-biphenyl]-2-yl)methoxy]-benzenepropanoic acid |
References
- Bisgaard, L.S.; Mogensen, C.K.; Rosendahl, A.; Cucak, H.; Nielsen, L.B.; Rasmussen, S.E.; Pedersen, T.X. Bone marrow-derived and peritoneal macrophages have different inflammatory response to oxLDL and M1/M2 marker expression—Implications for atherosclerosis research. Sci. Rep. 2016, 6, 35234. [Google Scholar] [CrossRef]
- Ross, R. Atherosclerosis--an inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Jiang, T.; Jiang, D.; You, D.; Zhang, L.; Liu, L.; Zhao, Q. Agonism of GPR120 prevents ox-LDL-induced attachment of monocytes to endothelial cells. Chem.-Biol. Interact. 2019, 316, 108916. [Google Scholar] [CrossRef]
- Bobryshev, Y.V.; Ivanova, E.A.; Chistiakov, D.A.; Nikiforov, N.G.; Orekhov, A.N. Macrophages and Their Role in Atherosclerosis: Pathophysiology and Transcriptome Analysis. BioMed Res. Int. 2016, 2016, 9582430. [Google Scholar] [CrossRef] [Green Version]
- Barrett, T.J. Macrophages in Atherosclerosis Regression. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 20–33. [Google Scholar] [CrossRef]
- Milligan, G.; Alvarez-Curto, E.; Hudson, B.D.; Prihandoko, R.; Tobin, A.B. FFA4/GPR120: Pharmacology and therapeutic opportunities. Trends Pharmacol. Sci. 2017, 38, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Shimpukade, B.; Hudson, B.D.; Hovgaard, C.K.; Milligan, G.; Ulven, T. Discovery of a potent and selective GPR120 agonist. J. Med. Chem. 2012, 55, 4511–4515. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Obesity and Free Fatty Acids (FFA). Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams-Bey, Y.; Boularan, C.; Vural, A.; Huang, N.N.; Hwang, I.Y.; Shan-Shi, C.; Kehrl, J.H. Omega-3 free fatty acids suppress macrophage inflammasome activation by inhibiting NF-κB activation and enhancing autophagy. PLoS ONE 2014, 9, e97957. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ballantyne, L.L.; Che, X.; Mewburn, J.D.; Kang, J.X.; Barkley, R.M.; Murphy, R.C.; Yu, Y.; Funk, C.D. Endogenously Generated Omega-3 Fatty Acids Attenuate Vascular Inflammation and Neointimal Hyperplasia by Interaction With Free Fatty Acid Receptor 4 in Mice. J. Am. Heart Assoc. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Kamata, R.; Bumdelger, B.; Kokubo, H.; Fujii, M.; Yoshimura, K.; Ishida, T.; Ishida, M.; Yoshizumi, M. EPA Prevents the Development of Abdominal Aortic Aneurysms through Gpr-120/Ffar-4. PLoS ONE 2016, 11, e0165132. [Google Scholar] [CrossRef]
- Grundmann, M.; Bender, E.; Schamberger, J.; Eitner, F. Pharmacology of Free Fatty Acid Receptors and Their Allosteric Modulators. Int. J. Mol. Sci. 2021, 22, 1763. [Google Scholar] [CrossRef]
- Suski, M.; Kiepura, A.; Wiśniewska, A.; Kuś, K.; Skałkowska, A.; Stachyra, K.; Stachowicz, A.; Gajda, M.; Korbut, R.; Olszanecki, R. Anti-atherosclerotic action of GW9508—Free fatty acid receptors activator—In apoE-knockout mice. Pharmacol. Rep. 2019, 71, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ren, X.; Tian, X.-F.; Cheng, X.-L.; Zhao, Y.-Y.; Li, Q.-Y.; Duan, Z.-Y.; Tian, L.-F.; Chen, Z.; Lu, J.-M.; et al. Protective effects of GPR120 agonist-programmed macrophages on renal interstitial fibrosis in unilateral ureteral obstruction (UUO) rats. Biomed. Pharmacother. 2019, 117, 109172. [Google Scholar] [CrossRef]
- Lin, P.; Ji, H.-H.; Li, Y.-J.; Guo, S.-D. Macrophage Plasticity and Atherosclerosis Therapy. Front. Mol. Biosci. 2021, 8, 679797. [Google Scholar] [CrossRef]
- Hara, T.; Hirasawa, A.; Sun, Q.; Sadakane, K.; Itsubo, C.; Iga, T.; Adachi, T.; Koshimizu, T.-A.; Hashimoto, T.; Asakawa, Y.; et al. Novel selective ligands for free fatty acid receptors GPR120 and GPR40. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 380, 247–255. [Google Scholar] [CrossRef]
- Su, X.-L.; Liu, Y.-G.; Shi, M.; Zhao, Y.-Y.; Liang, X.-Y.; Zhang, L.-J.; Wei, L.-L.; Zhao, Y.-F. The GPR120 Agonist TUG-891 Inhibits the Motility and Phagocytosis of Mouse Alveolar Macrophages. BioMed Res. Int. 2020, 2020, 1706168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wang, L.; Qiu, J.; Zha, D.; Sun, Q.; Chen, C. Linoleic Acid Stimulates [Ca2+]i Increase in Rat Pancreatic Beta-Cells through Both Membrane Receptor- and Intracellular Metabolite-Mediated Pathways. PLoS ONE 2013, 8, e60255. [Google Scholar] [CrossRef] [Green Version]
- Nagarkar, D.R.; Bowman, E.R.; Schneider, D.; Wang, Q.; Shim, J.; Zhao, Y.; Linn, M.; McHenry, C.L.; Gosangi, B.; Bentley, J.K.; et al. Rhinovirus Infection of Allergen-Sensitized and -Challenged Mice Induces Eotaxin Release from Functionally Polarized Macrophages. J. Immunol. 2010, 185, 2525–2535. [Google Scholar] [CrossRef] [PubMed]
- Kiepura, A.; Stachyra, K.; Olszanecki, R. Anti-Atherosclerotic Potential of Free Fatty Acid Receptor 4 (FFAR4). Biomedicines 2021, 9, 467. [Google Scholar] [CrossRef] [PubMed]
- Gozal, D.; Qiao, Z.; Almendros, I.; Zheng, J.; Khalyfa, A.; Shimpukade, B.; Ulven, T. Treatment with TUG891, a free fatty acid receptor 4 agonist, restores adipose tissue metabolic dysfunction following chronic sleep fragmentation in mice. Int. J. Obes. 2016, 40, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Schilperoort, M.; Van Dam, A.D.; Hoeke, G.; Shabalina, I.G.; Okolo, A.; Hanyaloglu, A.C.; Dib, L.H.; Mol, I.M.; Caengprasath, N.; Chan, Y.; et al. The GPR 120 agonist TUG-891 promotes metabolic health by stimulating mitochondrial respiration in brown fat. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewska, A.; Olszanecki, R.; Totoń-Żurańska, J.; Kuś, K.; Stachowicz, A.; Suski, M.; Gębska, A.; Gajda, M.; Jawień, J.; Korbut, R. Anti-Atherosclerotic Action of Agmatine in ApoE-Knockout Mice. Int. J. Mol. Sci. 2017, 18, 1706. [Google Scholar] [CrossRef]
- Stachowicz, A.; Wiśniewska, A.; Kuś, K.; Kiepura, A.; Gębska, A.; Gajda, M.; Białas, M.; Totoń-Żurańska, J.; Stachyra, K.; Suski, M.; et al. The Influence of Trehalose on Atherosclerosis and Hepatic Steatosis in Apolipoprotein E Knockout Mice. Int. J. Mol. Sci. 2019, 20, 1552. [Google Scholar] [CrossRef] [Green Version]
- Surmiak, M.; Hubalewska-Mazgaj, M.; Wawrzycka-Adamczyk, K.; Musiał, J.; Sanak, M. Neutrophil MiRNA-128-3p is Decreased During Active Phase of Granulomatosis with Polyangiitis. Curr. Genom. 2015, 16, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Total Cholesterol [mmol/L] | HDL [mmol/L] | LDL [mmol/L] | TG [mmol/L] |
|---|---|---|---|---|
| apoE−/− | 16.06 ± 2.55 | 1.28 ± 0.19 | 13.96 ± 2.03 | 1.02 ± 0.08 |
| apoE−/− + TUG-891 | 16.29 ± 2.87 | 1.29 ± 0.18 | 14. 37 ± 2.56 | 1.005 ± 0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiepura, A.; Stachyra, K.; Wiśniewska, A.; Kuś, K.; Czepiel, K.; Suski, M.; Ulatowska-Białas, M.; Surmiak, M.; Olszanecki, R. The Anti-Atherosclerotic Action of FFAR4 Agonist TUG-891 in ApoE–Knockout Mice Is Associated with Increased Macrophage Polarization towards M2 Phenotype. Int. J. Mol. Sci. 2021, 22, 9772. https://doi.org/10.3390/ijms22189772
Kiepura A, Stachyra K, Wiśniewska A, Kuś K, Czepiel K, Suski M, Ulatowska-Białas M, Surmiak M, Olszanecki R. The Anti-Atherosclerotic Action of FFAR4 Agonist TUG-891 in ApoE–Knockout Mice Is Associated with Increased Macrophage Polarization towards M2 Phenotype. International Journal of Molecular Sciences. 2021; 22(18):9772. https://doi.org/10.3390/ijms22189772
Chicago/Turabian StyleKiepura, Anna, Kamila Stachyra, Anna Wiśniewska, Katarzyna Kuś, Klaudia Czepiel, Maciej Suski, Magdalena Ulatowska-Białas, Marcin Surmiak, and Rafał Olszanecki. 2021. "The Anti-Atherosclerotic Action of FFAR4 Agonist TUG-891 in ApoE–Knockout Mice Is Associated with Increased Macrophage Polarization towards M2 Phenotype" International Journal of Molecular Sciences 22, no. 18: 9772. https://doi.org/10.3390/ijms22189772
APA StyleKiepura, A., Stachyra, K., Wiśniewska, A., Kuś, K., Czepiel, K., Suski, M., Ulatowska-Białas, M., Surmiak, M., & Olszanecki, R. (2021). The Anti-Atherosclerotic Action of FFAR4 Agonist TUG-891 in ApoE–Knockout Mice Is Associated with Increased Macrophage Polarization towards M2 Phenotype. International Journal of Molecular Sciences, 22(18), 9772. https://doi.org/10.3390/ijms22189772