
PIF4 and PIF4-Interacting Proteins: At the Nexus of Plant Light, Temperature and Hormone Signal Integrations


Abstract
:1. Introduction
2. Brief History of PIF4
3. PIF4-Interacting Photoreceptors
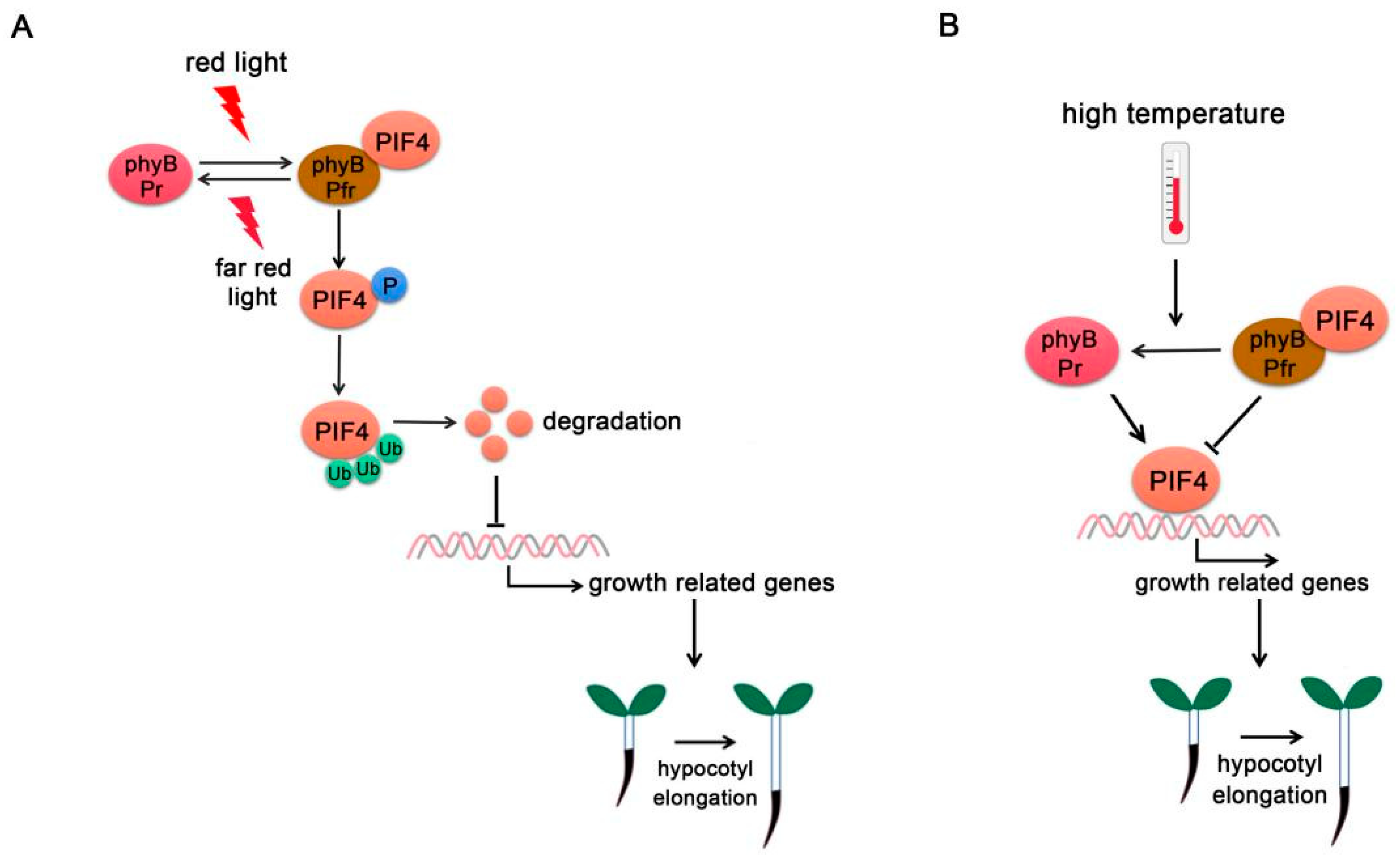
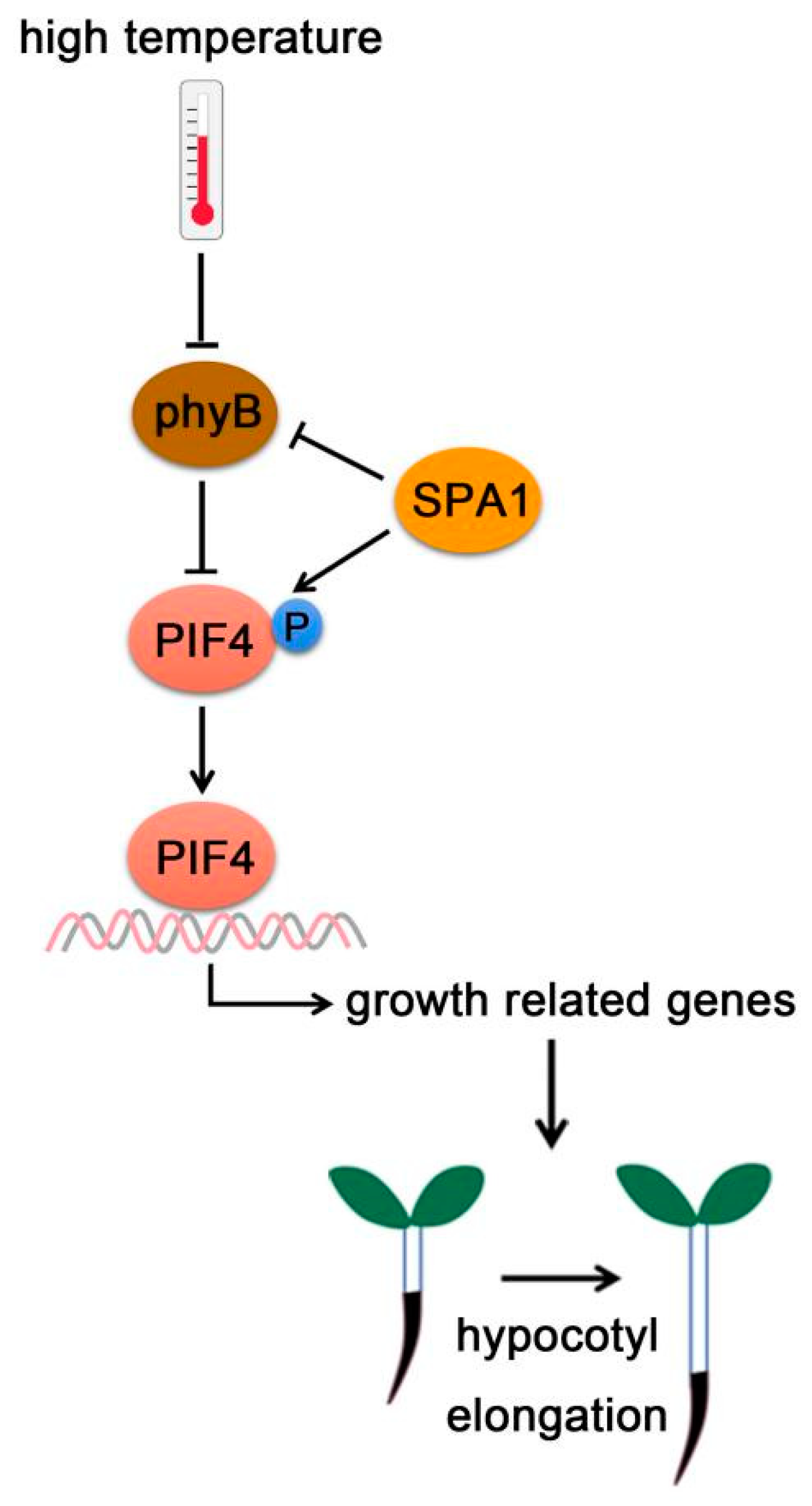
3.1. Phytochromes
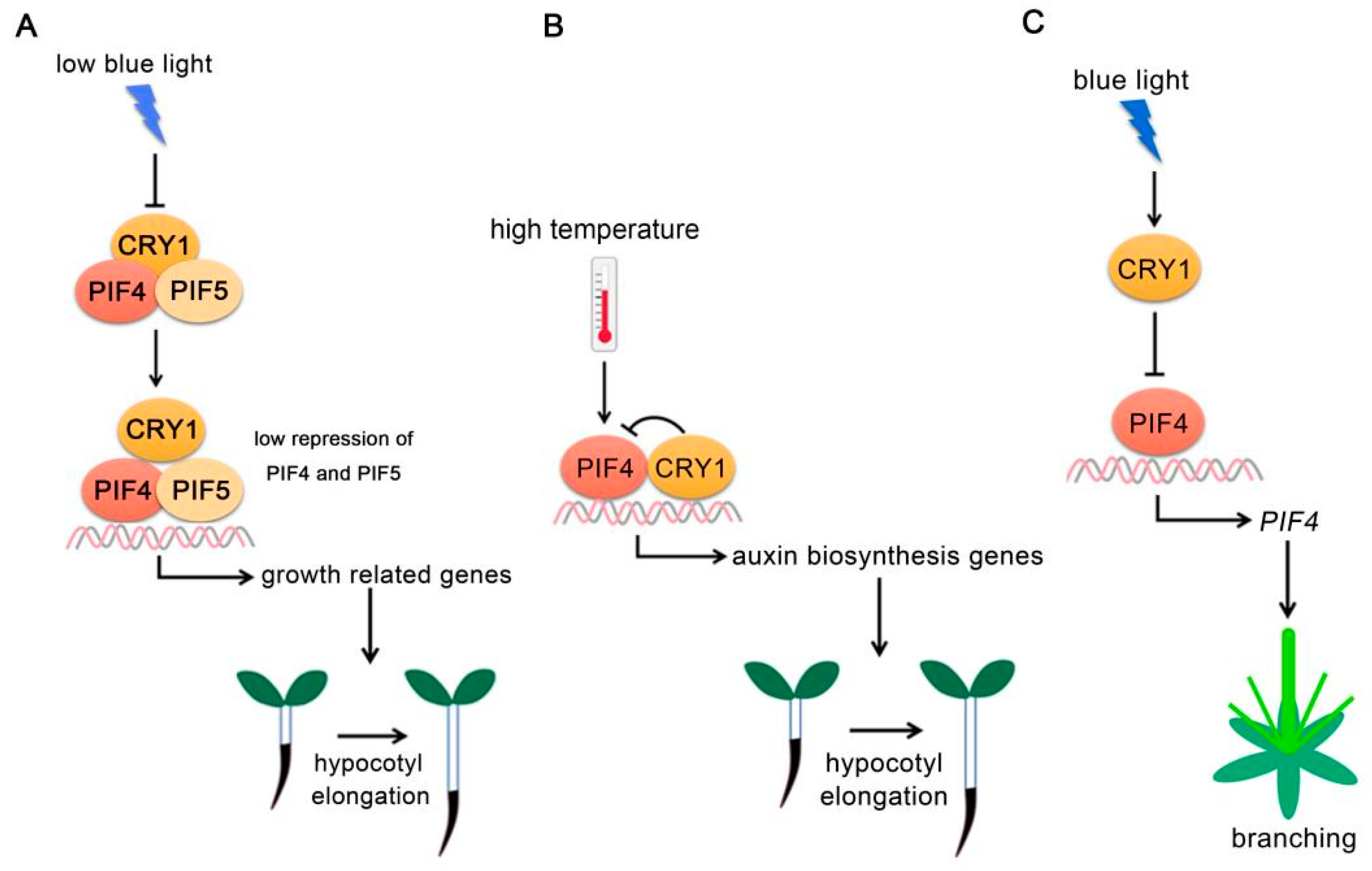
3.2. Cryptochromes
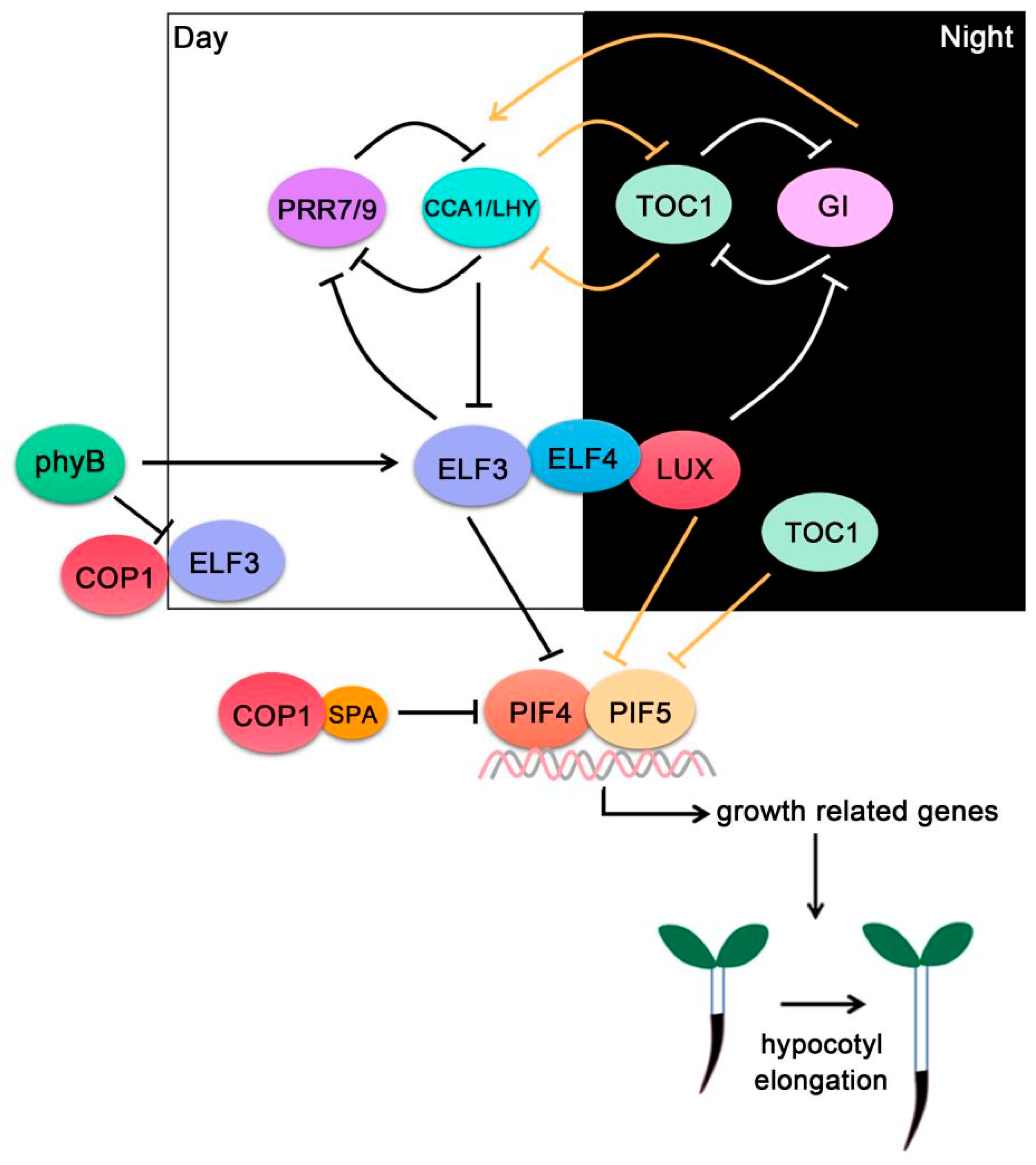
4. PIF4-Interacting Circadian Clock Components
4.1. ELF3
4.2. PRRs
5. PIF4-Interacting Plant Hormone Signaling Components
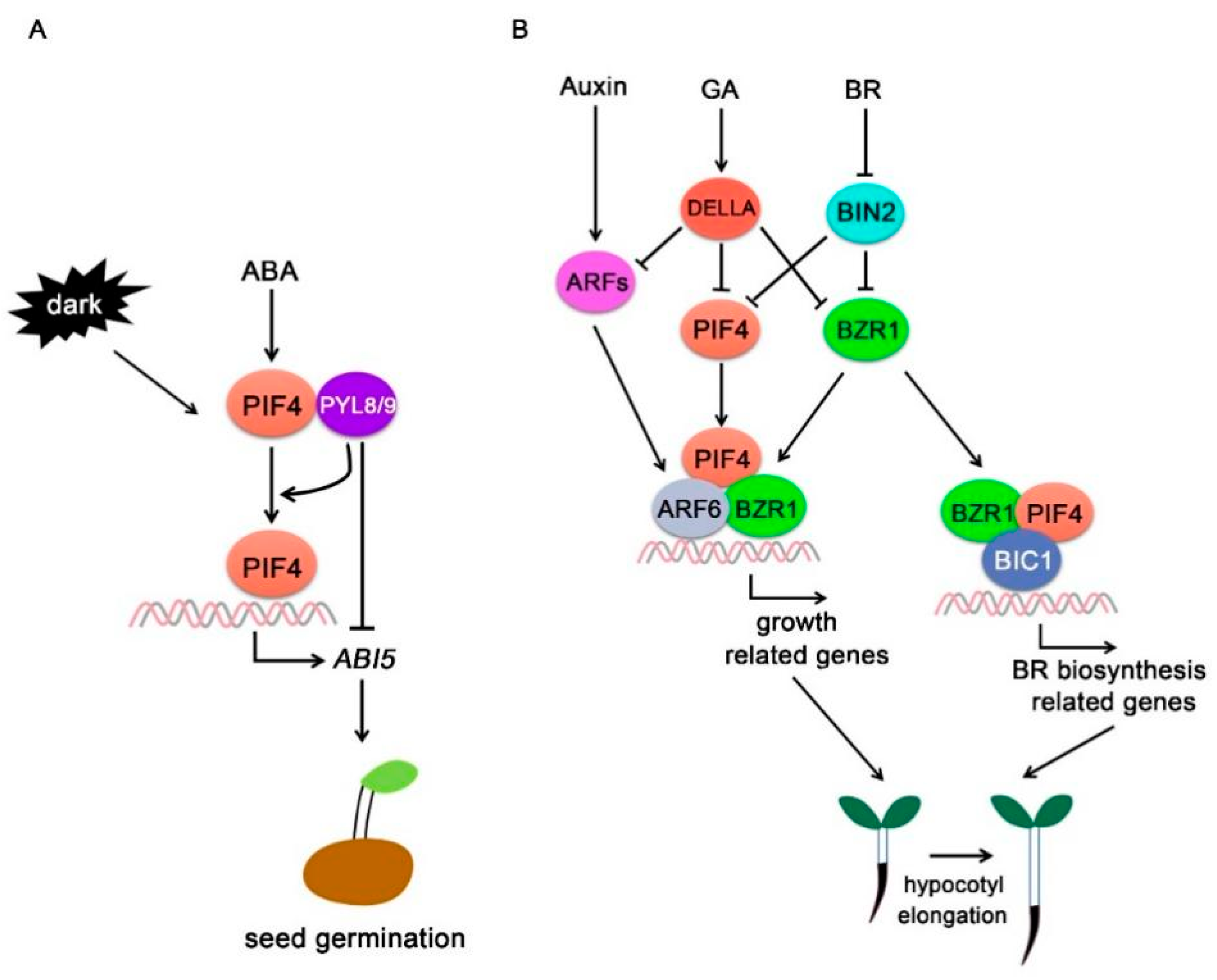
5.1. Abscisic Acid (ABA)
5.2. Gibberellins (GAs)
5.3. Brassinosteroid (BR)
5.4. Auxin
6. PIF4-Interacting Transcriptional Regulators
6.1. Positive Regulators
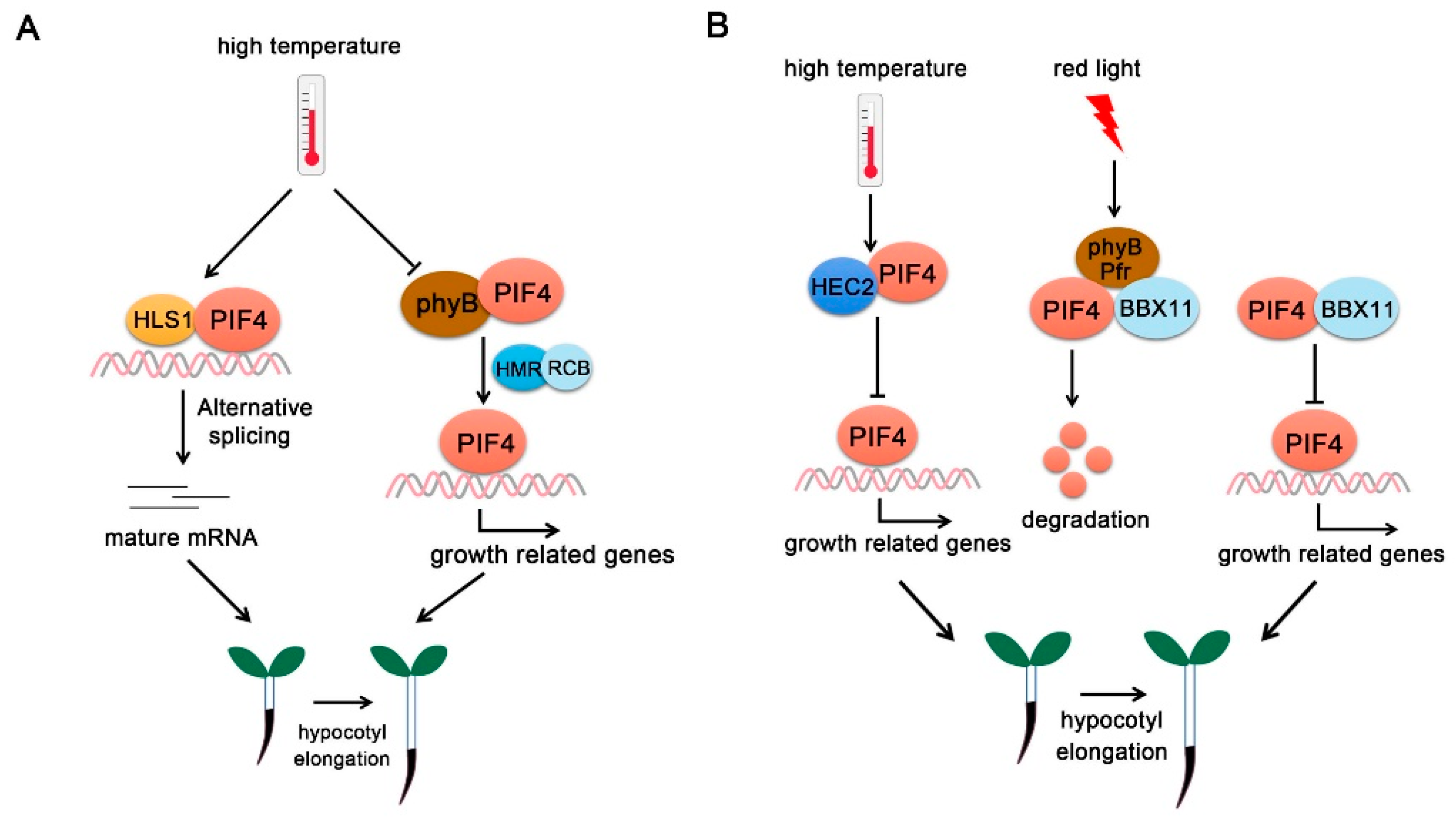
6.1.1. HLS1
6.1.2. HMR
6.2. Negative Regulators
6.2.1. BBX11
6.2.2. HEC2
7. PIF4-Interacting Kinase
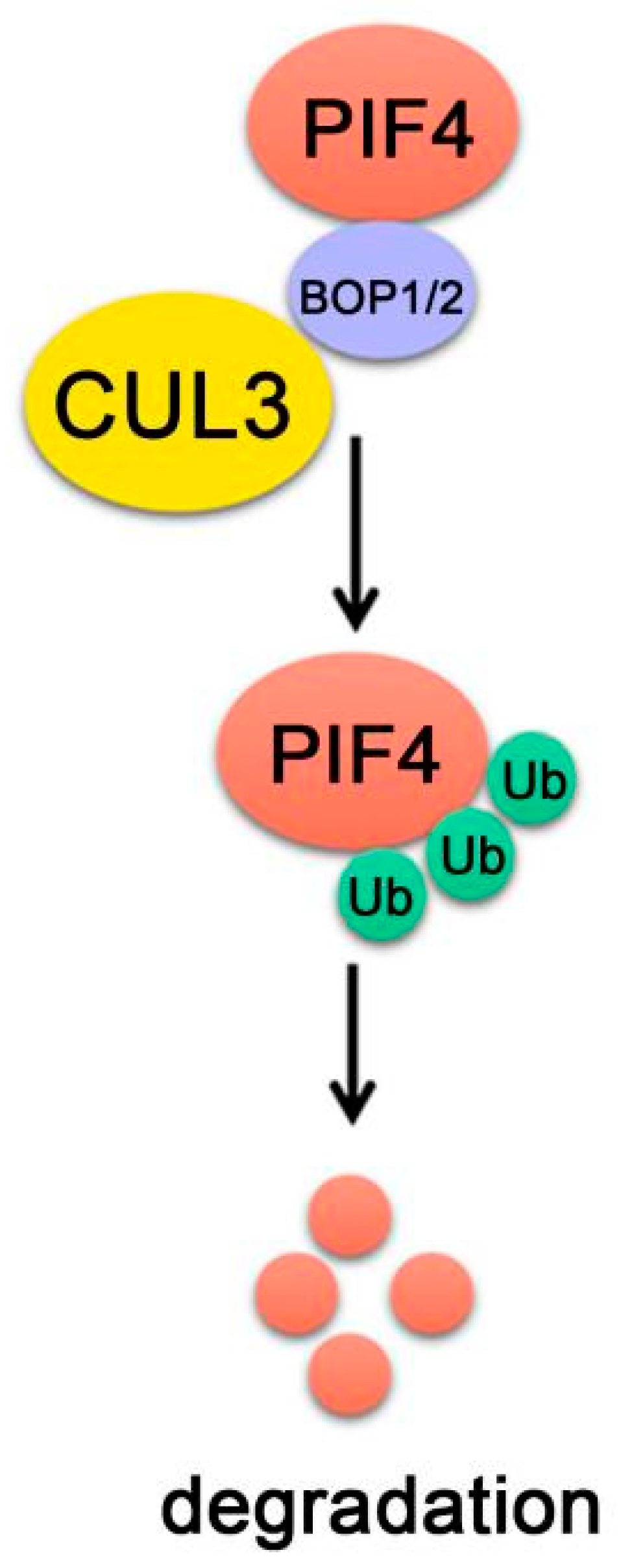
8. PIF4-Interacting E3 Ubiquitin Ligase
9. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, H.; Lin, R.; Deng, X.W. Photobiology: Light signal transduction and photomorphogenesis. J. Integr. Plant. Biol. 2020, 62, 1267–1269. [Google Scholar] [CrossRef]
- von Arnim, A.; Deng, X.-W. Light control of seedling development. Annu. Rev. Plant. Biol. 1996, 47, 215–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casal, J.J.; Balasubramanian, S. Thermomorphogenesis. Annu. Rev. Plant Biol. 2019, 70, 321–346. [Google Scholar] [CrossRef]
- Jeong, J.; Choi, G. Phytochrome-interacting factors have both shared and distinct biological roles. Mol. Cells 2013, 35, 371–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, V.N.; Xu, X.; Huq, E. Molecular bases for the constitutive photomorphogenic phenotypes in Arabidopsis. Development 2018, 145, dev.169870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huq, E.; Quail, P.H. PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis. EMBO J. 2002, 21, 2441–2450. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Cashmore, A.R. HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature 1993, 366, 162–166. [Google Scholar] [CrossRef]
- Qi, L.; Liu, S.; Li, C.; Fu, J.; Jing, Y.; Cheng, J.; Li, H.; Zhang, D.; Wang, X.; Dong, X.; et al. PHYTOCHROME-INTERACTING FACTORS Interact with the ABA Receptors PYL8 and PYL9 to Orchestrate ABA Signaling in Darkness. Mol. Plant. 2020, 13, 414–430. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.-Y.; Wang, Z.-Y. Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nature 2012, 14, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Yan, B.; Dong, H.; He, G.; Zhou, Y.; Sun, J. BIC1 acts as a transcriptional coactivator to promote brassinosteroid signaling and plant growth. EMBO J. 2021, 40, e104615. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.-Y.; Bai, M.-Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.-Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. eLife 2014, 3, e03031. [Google Scholar] [CrossRef] [PubMed]
- De Lucas, M.; Davière, J.-M.; Rodríguez-Falcón, M.; Pontin, M.; Pedraz, J.M.I.; Lorrain, S.; Fankhauser, C.; Blazquez, M.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Paik, I.; Huq, E. SPAs promote thermomorphogenesis by regulating the phyB-PIF4 module in Arabidopsis. Development 2020, 147, dev189233. [Google Scholar] [CrossRef]
- Song, Z.; Heng, Y.; Bian, Y.; Xiao, Y.; Liu, J.; Zhao, X.; Xu, D. BBX11 promotes red light-mediated photomorphogenic development by modulating phyB-PIF4 signaling. aBIOTECH 2021, 2, 117–130. [Google Scholar] [CrossRef]
- Qiu, Y.; Li, M.; Kim, R.J.-A.; Moore, C.M.; Chen, M. Daytime temperature is sensed by phytochrome B in Arabidopsis through a transcriptional activator HEMERA. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nieto, C.; Salmerón, V.L.; Davière, J.-M.; Prat, S. ELF3-PIF4 Interaction Regulates Plant Growth Independently of the Evening Complex. Curr. Biol. 2015, 25, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-Y.; Oh, E.; Wang, T.; Wang, Z.-Y. TOC1–PIF4 interaction mediates the circadian gating of thermoresponsive growth in Arabidopsis. Nat. Commun. 2016, 7, 13692. [Google Scholar] [CrossRef]
- Lee, S.; Zhu, L.; Huq, E. An autoregulatory negative feedback loop controls thermomorphogenesis in Arabidopsis. PLoS Genet. 2021, 17, e1009595. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Lin, J.; Zhu, Z. PIF4 and HOOKLESS1 Impinge on Common Transcriptome and Isoform Regulation in Thermomorphogenesis. Plant. Commun. 2020, 1, 100034. [Google Scholar] [CrossRef]
- Zhang, B.; Holmlund, M.; Lorrain, S.; Norberg, M.; Bakó, L.; Fankhauser, C.; Nilsson, O. BLADE-ON-PETIOLE proteins act in an E3 ubiquitin ligase complex to regulate PHYTOCHROME INTERACTING FACTOR 4 abundance. eLife 2017, 6, e26759. [Google Scholar] [CrossRef]
- Yan, Y.; Li, C.; Dong, X.; Li, H.; Zhang, D.; Zhou, Y.; Li, J. MYB30 Is a Key Negative Regulator of Arabidopsis Photomorphogenic Development That Promotes PIF4 and PIF5 Protein Accumulation in the Light. Plant. Cell 2020, 32, 2196–2215. [Google Scholar] [CrossRef] [PubMed]
- Huai, J.; Zhang, X.; Li, J.; Ma, T.; Zha, P.; Jing, Y.; Lin, R. SEUSS and PIF4 Coordinately Regulate Light and Temperature Signaling Pathways to Control Plant Growth. Mol. Plant. 2018, 11, 928–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Yu, H.; Yuan, R.; Yang, Y.; An, F.; Qin, G. Arabidopsis Transcription Factor TCP5 Controls Plant Thermomorphogenesis by Positively Regulating PIF4 Activity. iScience 2019, 15, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, M.; Tepperman, J.M.; Quail, P.H. PIF3, a Phytochrome-Interacting Factor Necessary for Normal Photoinduced Signal Transduction, Is a Novel Basic Helix-Loop-Helix Protein. Cell 1998, 95, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Al-Sady, B.; Ni, W.; Kircher, S.; Schäfer, E.; Quail, P.H. Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol. Cell 2006, 23, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.; Huq, E.; Kikis, E.A.; Al-Sady, B.; Lanzatella, C.; Quail, P.H. A novel molecular recognition motif necessary for targeting photoactivated phytochrome signaling to specific basic helix-loop-helix transcription factors. Plant. Cell 2004, 16, 3033–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.A.; Lee, S.H.; Patel, D.; Kumar, S.V.; Spartz, A.K.; Gu, C.; Ye, S.; Yu, P.; Breen, G.; Cohen, J.; et al. PHYTOCHROME-INTERACTING FACTOR 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. USA 2011, 108, 20231–20235. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Qi, L.; Li, Y.; Chu, J.; Li, C. PIF4–Mediated Activation of YUCCA8 Expression Integrates Temperature into the Auxin Pathway in Regulating Arabidopsis Hypocotyl Growth. PLoS Genet. 2012, 8, e1002594. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Li, X.; Guo, Y.; Chu, J.; Fang, S.; Yan, C.; Liu, H. Cryptochrome 1 interacts with PIF4 to regulate high temperature-mediated hypocotyl elongation in response to blue light. Proc. Natl. Acad. Sci. USA. 2016, 113, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Xin, R.; Bu, Q.; Shen, H.; Dang, J.; Huq, E. A Negative Feedback Loop between PHYTOCHROME INTER-ACTING FACTORs and HECATE Proteins Fine-Tunes Photomorphogenesis in Arabidopsis. Plant. Cell 2016, 28, 855–874. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 2007, 8, 217–230. [Google Scholar] [CrossRef]
- Leivar, P.; Quail, P.H. PIFs: Pivotal components in a cellular signaling hub. Trends Plant. Sci. 2011, 16, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Paik, I.; Zhu, L.; Huq, E. Illuminating Progress in Phytochrome-Mediated Light Signaling Pathways. Trends Plant. Sci. 2015, 20, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Ni, W.; Xu, S.L.; Tepperman, J.M.; Stanley, D.J.; Maltby, D.A.; Gross, J.D.; Quail, P.H. A mutually assured de-struction mechanism attenuates light signaling in Arabidopsis. Science 2014, 344, 1160–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christians, M.J.; Gingerich, D.; Hua, Z.; Lauer, T.D.; Vierstra, R.D. The Light-Response BTB1 and BTB2 Proteins Assemble Nuclear Ubiquitin Ligases That Modify Phytochrome B and D Signaling in Arabidopsis. Plant Physiol. 2012, 160, 118–134. [Google Scholar] [CrossRef] [Green Version]
- Sharrock, R.A.; Quail, P.H. Novel phytochrome sequences in Arabidopsis thaliana: Structure, evolution, and dif-ferential expression of a plant regulatory photoreceptor family. Genes Dev. 1989, 3, 1745–1757. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.-H.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.K.; Box, M.S.; Charoensawan, V.; Cortijo, S.; et al. Phytochromes function as thermosensors inArabidopsis. Science 2016, 354, 886–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legris, M.; Klose, C.; Burgie, E.S.; Rojas, C.C.R.; Neme, M.; Hiltbrunner, A.; Wigge, P.A.; Schäfer, E.; Vierstra, R.D.; Casal, J.J. Phytochrome B integrates light and temperature signals in Arabidopsis. Science 2016, 354, 897–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, P.D.; Batschauer, A.; Hays, J.B. PHH1, a novel gene from Arabidopsis thaliana that encodes a protein similar to plant blue-light photoreceptors and microbial photolyases. Mol. Genet. Genom. 1996, 253, 259–265. [Google Scholar] [CrossRef]
- Cashmore, A.R.; Jarillo, J.A.; Wu, Y.; Liu, D. Cryptochromes: Blue Light Receptors for Plants and Animals. Science 1999, 284, 760–765. [Google Scholar] [CrossRef]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of Flowering Time by Arabidopsis Photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Ballaré, C.L. Keeping up with the neighbours: Phytochrome sensing and other signalling mechanisms. Trends Plant Sci. 1999, 4, 97–102. [Google Scholar] [CrossRef]
- Keller, M.M.; Jaillais, Y.; Pedmale, U.V.; Moreno, J.; Chory, J.; Ballaré, C.L. Cryptochrome 1 and phytochrome B control shade-avoidance responses in Arabidopsis via partially independent hormonal cascades. Plant. J. 2011, 67, 195–207. [Google Scholar] [CrossRef]
- Keuskamp, D.H.; Sasidharan, R.; Pierik, R. Physiological regulation and functional significance of shade avoidance responses to neighbors. Plant. Signal. Behav. 2010, 5, 655–662. [Google Scholar] [CrossRef] [Green Version]
- de Wit, M.; Keuskamp, D.H.; Bongers, F.J.; Hornitschek, P.; Gommers, C.; Reinen, E.; Martínez-Cerón, C.; Fankhauser, C.; Pierik, R. Integration of Phytochrome and Cryptochrome Signals Determines Plant Growth during Competition for Light. Curr. Biol. 2016, 26, 3320–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedmale, U.V.; Huang, S.-S.C.; Zander, M.; Cole, B.J.; Hetzel, J.; Ljung, K.; Reis, P.A.; Sridevi, P.; Nito, K.; Nery, J.R.; et al. Cryptochromes Interact Directly with PIFs to Control Plant Growth in Limiting Blue Light. Cell 2015, 164, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Zhai, H.; Xiong, L.; Li, H.; Lyu, X.; Yang, G.; Zhao, T.; Liu, J.; Liu, B. Cryptochrome 1 Inhibits Shoot Branching by Repressing the Self-Activated Transciption Loop of PIF4 in Arabidopsis. Plant. Commun. 2020, 1, 100042. [Google Scholar] [CrossRef]
- Nozue, K.; Covington, M.F.; Duek, P.D.; Lorrain, S.; Fankhauser, C.; Harmer, S.; Maloof, J.N. Rhythmic growth explained by coincidence between internal and external cues. Nature 2007, 448, 358–361. [Google Scholar] [CrossRef]
- Michael, T.P.; Breton, G.; Hazen, S.P.; Priest, H.; Mockler, T.C.; Kay, S.A.; Chory, J. A morning-specific phyto-hormone gene expression program underlying rhythmic plant growth. PLoS Biol. 2008, 6, e225. [Google Scholar] [CrossRef] [Green Version]
- Niwa, Y.; Yamashino, T.; Mizuno, T. The Circadian Clock Regulates the Photoperiodic Response of Hypocotyl Elongation through a Coincidence Mechanism in Arabidopsis thaliana. Plant. Cell Physiol. 2009, 50, 838–854. [Google Scholar] [CrossRef] [Green Version]
- de Montaigu, A.; Tóth, R.; Coupland, G. Plant development goes like clockwork. Trends Genet. 2010, 26, 296–306. [Google Scholar] [CrossRef]
- Pokhilko, A.; Fernández, A.P.; Edwards, K.; Southern, M.M.; Halliday, K.; Millar, A.J. The clock gene circuit in Arabidopsis includes a repressilator with additional feedback loops. Mol. Syst. Biol. 2012, 8, 574. [Google Scholar] [CrossRef]
- Farre, E.; Harmer, S.; Harmon, F.G.; Yanovsky, M.J.; Kay, S.A. Overlapping and Distinct Roles of PRR7 and PRR9 in the Arabidopsis Circadian Clock. Curr. Biol. 2005, 15, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, T.; Nakamichi, N. Pseudo-Response Regulators (PRRs) or True Oscillator Components (TOCs). Plant. Cell Physiol. 2005, 46, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Nusinow, D.; Helfer, A.; Hamilton, E.E.; King, J.J.; Imaizumi, T.; Schultz, T.; Farre, E.; Kay, S.A. The ELF4–ELF3–LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 2011, 475, 398–402. [Google Scholar] [CrossRef]
- Matsushika, A.; Makino, S.; Kojima, M.; Mizuno, T. Circadian Waves of Expression of the APRR1/TOC1 Family of Pseudo-Response Regulators in Arabidopsis thaliana: Insight into the Plant Circadian Clock. Plant. Cell Physiol. 2000, 41, 1002–1012. [Google Scholar] [CrossRef] [Green Version]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.-H.; Sakakibara, H. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 Are Transcriptional Repressors in the Arabidopsis Circadian Clock. Plant. Cell 2010, 22, 594–605. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, Y.; He, Y.; Wang, Y.; Wang, L. Pseudo Response Regulators Regulate Photoperiodic Hypocotyl Growth by Repressing PIF4/5 Transcription. Plant Physiol. 2020, 183, 686–699. [Google Scholar] [CrossRef] [Green Version]
- Murata, Y.; Mori, I.C.; Munemasa, S. Diverse Stomatal Signaling and the Signal Integration Mechanism. Annu. Rev. Plant. Biol. 2015, 66, 369–392. [Google Scholar] [CrossRef]
- Qi, J.; Song, C.P.; Wang, B.; Zhou, J.; Kangasjärvi, J.; Zhu, J.K.; Gong, Z. Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack. J. Integr. Plant. Biol. 2018, 60, 805–826. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, K.; Nishimura, N.; Hitomi, K.; Getzoff, E.D.; Schroeder, J.I. Early abscisic acid signal transduction mechanisms: Newly discovered components and newly emerging questions. Genes Dev. 2010, 24, 1695–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.-P. Gibberellin Metabolism, Perception and Signaling Pathways in Arabidopsis. Arab. Book 2008, 6, e0103. [Google Scholar] [CrossRef]
- Tyler, L.; Thomas, S.G.; Hu, J.; Dill, A.; Alonso, J.M.; Ecker, J.; Sun, T.-P. DELLA Proteins and Gibberellin-Regulated Seed Germination and Floral Development in Arabidopsis. Plant. Physiol. 2004, 135, 1008–1019. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Chory, J.; Fankhauser, C. Light Signal Transduction in Higher Plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabadí, D.; Gil, J.; Blázquez, M.A.; García-Martínez, J.L. Gibberellins Repress Photomorphogenesis in Darkness. Plant Physiol. 2004, 134, 1050–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.-P.; Gubler, F. MOLECULAR MECHANISM OF GIBBERELLIN SIGNALING IN PLANTS. Annu. Rev. Plant. Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Matsuoka, M.; Steber, C.M. A role for the ubiquitin–26S-proteasome pathway in gibberellin signaling. Trends Plant. Sci. 2003, 8, 492–497. [Google Scholar] [CrossRef]
- Achard, P.; Liao, L.; Jiang, C.; Desnos, T.; Bartlett, J.; Fu, X.; Harberd, N.P. DELLAs contribute to plant photomorphogenesis. Plant Physiol. 2007, 143, 1163–1172. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Martinez, C.; Gusmaroli, G.; Wang, Y.; Zhou, J.; Wang, F.; Chen, L.; Yu, L.; Pedraz, J.M.I.; Kircher, S.; et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature 2008, 451, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Yu, R.; Fan, L.-M.; Wei, N.; Chen, H.; Deng, X.W. DELLA-mediated PIF degradation contributes to coordination of light and gibberellin signalling in Arabidopsis. Nat. Commun. 2016, 7, 11868. [Google Scholar] [CrossRef] [Green Version]
- Clouse, S.D.; Sasse, J.M. BRASSINOSTEROIDS: Essential Regulators of Plant Growth and Development. Annu. Rev. Plant Biol. 1998, 49, 427–451. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.-Y.; Shang, J.-X.; Oh, E.; Fan, M.; Bai, Y.; Zentella, R.; Sun, T.-P.; Wang, Z.-Y. Brassinosteroid, gibberellin and phytochrome impinge on a common transcription module in Arabidopsis. Nature 2012, 14, 810–817. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, B.; Xu, Y.; Li, H.; Li, S.; Zhang, D.; Mao, Z.; Guo, S.; Yang, C.; Weng, Y.; et al. The Cyclophilin CYP20-2 Modulates the Conformation of BRASSINAZOLE-RESISTANT1, Which Binds the Promoter of FLOWERING LOCUS D to Regulate Flowering in Arabidopsis. Plant. Cell 2013, 25, 2504–2521. [Google Scholar] [CrossRef] [Green Version]
- Chaiwanon, J.; Wang, W.; Zhu, J.-Y.; Oh, E.; Wang, Z.-Y. Information Integration and Communication in Plant Growth Regulation. Cell 2016, 164, 1257–1268. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Oh, E. PIF4 Integrates Multiple Environmental and Hormonal Signals for Plant Growth Regulation in Arabidopsis. Mol. Cells 2016, 39, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Chory, J.; Nagpal, P.; Peto, C.A. Phenotypic and Genetic Analysis of det2, a New Mutant That Affects Light-Regulated Seedling Development in Arabidopsis. Plant Cell 1991, 3, 445–459. [Google Scholar] [CrossRef]
- Clouse, S.D.; Langford, M.; McMorris, T.C.; du Jardin, P.; Rojas-Beltran, J.; Gebhardt, C.; Brasseur, R. A Brassinosteroid-Insensitive Mutant in Arabidopsis thaliana Exhibits Multiple Defects in Growth and Development. Plant Physiol. 1996, 111, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Nagpal, P.; Vitart, V.; McMorris, T.C.; Chory, J. A Role for Brassinosteroids in Light-Dependent Development of Arabidopsis. Science 1996, 272, 398–401. [Google Scholar] [CrossRef]
- Szekeres, M.; Németh, K.; Koncz-Kálmán, Z.; Mathur, J.; Kauschmann, A.; Altmann, T.; Koncz, C. Brassinosteroids rescue the deficiency of CYP90, a cytochrome P450, controlling cell elongation and de-etiolation in Arabidopsis. Cell 1996, 85, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-W.; Guan, S.; Sun, Y.; Deng, Z.; Tang, W.; Shang, J.-X.; Sun, Y.; Burlingame, A.L.; Wang, Z.-Y. Brassinosteroid signal transduction from cell-surface receptor kinases to nuclear transcription factors. Nature 2009, 11, 1254–1260. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Bai, M.-Y.; Oh, E.; Zhu, J.-Y. Brassinosteroid Signaling Network and Regulation of Photomorphogenesis. Annu. Rev. Genet. 2012, 46, 701–724. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, Z.Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Martínez, C.; Espinosa-Ruíz, A.; De Lucas, M.; Bernardo-García, S.; Franco-Zorrilla, J.M.; Prat, S. PIF 4-induced BR synthesis is critical to diurnal and thermomorphogenic growth. EMBO J. 2018, 37, e99552. [Google Scholar] [CrossRef]
- Stewart, J.L.; Nemhauser, J.L. Do Trees Grow on Money? Auxin as the Currency of the Cellular Economy. Cold Spring Harb. Perspect. Biol. 2009, 2, a001420. [Google Scholar] [CrossRef] [Green Version]
- Del Bianco, M.; Kepinski, S. Context, Specificity, and Self-Organization in Auxin Response. Cold Spring Harb. Perspect. Biol. 2010, 3, a001578. [Google Scholar] [CrossRef] [Green Version]
- Chapman, E.J.; Estelle, M. Mechanism of Auxin-Regulated Gene Expression in Plants. Annu. Rev. Genet. 2009, 43, 265–285. [Google Scholar] [CrossRef] [Green Version]
- Vernoux, T.; Brunoud, G.; Farcot, E.; Morin, V.; Daele, H.V.D.; Legrand, J.; Oliva, M.; Das, P.; Larrieu, A.; Wells, D.; et al. The auxin signalling network translates dynamic input into robust patterning at the shoot apex. Mol. Syst. Biol. 2011, 7, 508. [Google Scholar] [CrossRef]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A Role for Flavin Monooxygenase-Like Enzymes in Auxin Biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, P.; Ecker, J.R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant. Cell 1990, 2, 513–523. [Google Scholar]
- Lehman, A.; Black, R.; Ecker, J. HOOKLESS1, an Ethylene Response Gene, Is Required for Differential Cell Elongation in the Arabidopsis Hypocotyl. Cell 1996, 85, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.-J.; Lai, Z.; Lee, S.; Yun, D.J.; Mengiste, T. Arabidopsis HOOKLESS1 regulates responses to pathogens and abscisic acid through interaction with MED18 and acetylation of WRKY33 and ABI5 chromatin. Plant. Cell 2016, 28, 1662–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Zhu, Z. HOOKLESS1 is a positive regulator in Arabidopsis thermomorphogenesis. Sci. China Life Sci. 2018, 62, 423–425. [Google Scholar] [CrossRef]
- Chen, M.; Galvão, R.M.; Li, M.; Burger, B.; Bugea, J.; Bolado, J.; Chory, J. Arabidopsis HEMERA/pTAC12 Initiates Photomorphogenesis by Phytochromes. Cell 2010, 141, 1230–1240. [Google Scholar] [CrossRef] [Green Version]
- Nevarez, P.A.; Qiu, Y.; Inoue, H.; Yoo, C.Y.; Benfey, P.N.; Schnell, D.J.; Chen, M. Mechanism of Dual Targeting of the Phytochrome Signaling Component HEMERA/pTAC12 to Plastids and the Nucleus. Plant. Physiol. 2017, 173, 1953–1966. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Li, M.; Pasoreck, E.K.; Long, L.; Shi, Y.; Galvão, R.M.; Chou, C.L.; Wang, H.; Sun, A.Y.; Zhang, Y.C.; et al. HEMERA Couples the Proteolysis and Transcriptional Activity of PHYTOCHROME INTERACTING FACTORs in Arabidopsis Photomorphogenesis. Plant. Cell 2015, 27, 1409–1427. [Google Scholar] [CrossRef] [Green Version]
- Pfalz, J.; Liere, K.; Kandlbinder, A.; Dietz, K.J.; Oelmüller, R. pTAC2, -6, and -12 are components of the transcriptionally active plastid chromosome that are required for plastid gene expression. Plant. Cell 2006, 18, 176–197. [Google Scholar] [CrossRef] [Green Version]
- Galvão, R.M.; Li, M.; Kothadia, S.M.; Haskel, J.D.; Decker, P.V.; Van Buskirk, E.K.; Chen, M. Photoactivated phytochromes interact with HEMERA and promote its accumulation to establish photomorphogenesis in Arabidopsis. Genes Dev. 2012, 26, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Pasoreck, E.K.; Yoo, C.Y.; He, J.; Wang, H.; Bajracharya, A.; Chen, M. RCB initiates Arabidopsis thermo-morphogenesis by stabilizing the thermoregulator PIF4 in the daytime. Nat. Commun. 2021, 12, 2042. [Google Scholar] [CrossRef]
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–470. [Google Scholar] [CrossRef]
- Vaishak, K.; Yadukrishnan, P.; Bakshi, S.; Kushwaha, A.K.; Ramachandran, H.; Job, N.; Babu, D.; Datta, S. The B-box bridge between light and hormones in plants. J. Photochem. Photobiol. B Biol. 2018, 191, 164–174. [Google Scholar] [CrossRef]
- Song, Z.; Yan, T.; Liu, J.; Bian, Y.; Heng, Y.; Lin, F.; Xu, D. BBX28/BBX29, HY5 and BBX30/31 form a feedback loop to fine-tune photomorphogenic development. Plant J. 2020, 104, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Xu, D. COP1 and BBXs-HY5-mediated light signal transduction in plants. New Phytol. 2020, 228, 1748–1753. [Google Scholar] [CrossRef]
- Khanna, R.; Kronmiller, B.; Maszle, D.R.; Coupland, G.; Holm, M.; Mizuno, T.; Wu, S.-H. The Arabidopsis B-Box Zinc Finger Family. Plant. Cell 2009, 21, 3416–3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Hettiarachchi, C.; Johansson, H.; Holm, M. SALT TOLERANCE HOMOLOG2, a B-box protein in Arabidopsis that activates transcription and positively regulates light-mediated development. Plant Cell 2007, 19, 3242–3255. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Hettiarachchi, G.; Deng, X.-W.; Holm, M. Arabidopsis CONSTANS-LIKE3 Is a Positive Regulator of Red Light Signaling and Root Growth. Plant Cell 2005, 18, 70–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Johansson, H.; Hettiarachchi, C.; Irigoyen, M.L.; Desai, M.; Rubio, V.; Holm, M. LZF1/SALT TOLERANCE HOMOLOG3, an Arabidopsis B-box protein involved in light-dependent development and gene expression, undergoes COP1-mediated ubiquitination. Plant Cell 2008, 20, 2324–2338. [Google Scholar]
- Holtan, H.E.; Bandong, S.; Marion, C.M.; Adam, L.; Tiwari, S.; Shen, Y.; Maloof, J.; Maszle, D.R.; Ohto, M.-A.; Preuss, S.; et al. BBX32, an Arabidopsis B-Box Protein, Functions in Light Signaling by Suppressing HY5-Regulated Gene Expression and Interacting with STH2/BBX21. Plant. Physiol. 2011, 156, 2109–2123. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.-Y.; Sun, Y.; Cao, D.-M.; Bai, M.-Y.; Luo, X.-M.; Yang, H.-J.; Wei, C.-Q.; Zhu, S.-W.; Sun, Y.; Chong, K.; et al. BZS1, a B-box Protein, Promotes Photomorphogenesis Downstream of Both Brassinosteroid and Light Signaling Pathways. Mol. Plant 2012, 5, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangappa, S.; Crocco, C.D.; Johansson, H.; Datta, S.; Hettiarachchi, C.; Holm, M.; Botto, J.F. The Arabidopsis B-BOX Protein BBX25 Interacts with HY5, Negatively Regulating BBX22 Expression to Suppress Seedling Photomorphogenesis. Plant Cell 2013, 25, 1243–1257. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Q.; Sarmast, M.K.; Jiang, J.; Dehesh, K. The Transcriptional Regulator BBX19 Promotes Hypocotyl Growth by Facilitating COP1-Mediated EARLY FLOWERING3 Degradation in Arabidopsis. Plant. Cell 2015, 27, 1128–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Jiang, Y.; Li, J.; Lin, F.; Holm, M.; Deng, X.W. BBX21, an Arabidopsis B-box protein, directly activates HY5 and is targeted by COP1 for 26S proteasome-mediated degradation. Proc. Natl. Acad. Sci. USA 2016, 113, 7655–7660. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Jiang, Y.; Li, J.; Holm, M.; Deng, X.W. The B-Box Domain Protein BBX21 Promotes Photomorphogenesis. Plant. Physiol. 2017, 176, 2365–2375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huai, J.; Shang, F.; Xu, G.; Tang, W.; Jing, Y.; Lin, R. A PIF1/PIF3-HY5-BBX23 Transcription Factor Cascade Affects Photomorphogenesis. Plant. Physiol. 2017, 174, 2487–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Job, N.; Yadukrishnan, P.; Bursch, K.; Datta, S.; Johansson, H. Two B-Box Proteins Regulate Photomorphogenesis by Oppositely Modulating HY5 through their Diverse C-Terminal Domains. Plant. Physiol. 2018, 176, 2963–2976. [Google Scholar] [CrossRef]
- Lin, F.; Jiang, Y.; Li, J.; Yan, T.; Fan, L.; Liang, J.; Chen, Z.J.; Xu, D.; Deng, X.W. B-BOX DOMAIN PROTEIN28 Negatively Regulates Photomorphogenesis by Repressing the Activity of Transcription Factor HY5 and Undergoes COP1-Mediated Degradation. Plant. Cell 2018, 30, 2006–2019. [Google Scholar] [CrossRef] [Green Version]
- Heng, Y.; Jiang, Y.; Zhao, X.; Zhou, H.; Wang, X.; Deng, X.W.; Xu, D. BBX4, a phyB-interacting and modulated regulator, directly interacts with PIF3 to fine tune red light-mediated photomorphogenesis. Proc. Natl. Acad. Sci. USA 2019, 116, 26049–26056. [Google Scholar] [CrossRef]
- Heng, Y.; Lin, F.; Jiang, Y.; Ding, M.; Yan, T.; Lan, H.; Zhou, H.; Zhao, X.; Xu, D.; Deng, X.W. B-Box Containing Proteins BBX30 and BBX31, Acting Downstream of HY5, Negatively Regulate Photomorphogenesis in Arabidopsis. Plant. Physiol. 2019, 180, 497–508. [Google Scholar] [CrossRef]
- Yadav, A.; Bakshi, S.; Yadukrishnan, P.; Lingwan, M.; Dolde, U.; Wenkel, S.; Datta, S. The B-Box-Containing MicroProtein miP1a/BBX31 Regulates Photomorphogenesis and UV-B Protection. Plant. Physiol. 2019, 179, 1876–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bursch, K.; Toledo-Ortiz, G.; Pireyre, M.; Lohr, M.; Braatz, C.; Johansson, H. Identification of BBX proteins as rate-limiting cofactors of HY5. Nat. Plants 2020, 6, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Bian, Y.; Liu, J.; Sun, Y.; Xu, D. B-box proteins: Pivotal players in light-mediated development in plants. J. Integr. Plant. Biol. 2020, 62, 1293–1309. [Google Scholar] [CrossRef]
- Wu, Q.; Kuang, K.; Lyu, M.; Zhao, Y.; Li, Y.; Li, J.; Pan, Y.; Shi, H.; Zhong, S. Allosteric deactivation of PIFs and EIN3 by microproteins in light control of plant development. Proc. Natl. Acad. Sci. USA 2020, 117, 18858–18868. [Google Scholar] [CrossRef]
- Zhao, X.; Heng, Y.; Wang, X.; Deng, X.W.; Xu, D. A Positive Feedback Loop of BBX11-BBX21-HY5 Promotes Photomorphogenic Development in Arabidopsis. Plant. Commun. 2020, 1, 100045. [Google Scholar] [CrossRef]
- Gremski, K.; Ditta, G.; Yanofsky, M.F. The HECATE genes regulate female reproductive tract development in Arabidopsis thaliana. Development 2007, 134, 3593–3601. [Google Scholar] [CrossRef] [Green Version]
- Kathare, P.K.; Xu, X.; Nguyen, A.; Huq, E. A COP1-PIF-HEC regulatory module fine-tunes photomorphogenesis in Arabidopsis. Plant. J. 2020, 104, 113–123. [Google Scholar] [CrossRef]
- Hoecker, U. The activities of the E3 ubiquitin ligase COP1/SPA, a key repressor in light signaling. Curr. Opin. Plant. Biol. 2017, 37, 63–69. [Google Scholar] [CrossRef]
- Hoecker, U.; Tepperman, J.M.; Quail, P.H. SPA1, a WD-Repeat Protein Specific to Phytochrome A Signal Transduction. Science 1999, 284, 496–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laubinger, S.; Fittinghoff, K.; Hoecker, U. The SPA Quartet: A Family of WD-Repeat Proteins with a Central Role in Suppression of Photomorphogenesis in Arabidopsis. Plant. Cell 2004, 16, 2293–2306. [Google Scholar] [CrossRef] [Green Version]
- Pham, V.N.; Kathare, P.K.; Huq, E. Phytochromes and Phytochrome Interacting Factors. Plant. Physiol. 2017, 176, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Paik, I.; Chen, F.; Ngoc Pham, V.; Zhu, L.; Kim, J.I.; Huq, E. A phyB-PIF1-SPA1 kinase regulatory complex promotes photomorphogenesis in Arabidopsis. Nat. Commun. 2019, 10, 4216. [Google Scholar] [CrossRef] [Green Version]
- Ha, C.M.; Kim, G.-T.; Kim, B.C.; Jun, J.H.; Soh, M.-S.; Ueno, Y.; Machida, Y.; Tsukaya, H.; Gil Nam, H. The BLADE-ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis. Development 2003, 130, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Hepworth, S.; Zhang, Y.; McKim, S.; Li, X.; Haughn, G.W. BLADE-ON-PETIOLE–Dependent Signaling Controls Leaf and Floral Patterning in Arabidopsis. Plant. Cell 2005, 17, 1434–1448. [Google Scholar] [CrossRef]
- Norberg, M.; Holmlund, M.; Nilsson, O. The BLADE ON PETIOLE genes act redundantly to control the growth and development of lateral organs. Development 2005, 132, 2203–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.A.; Quail, P.H. Phytochrome functions in Arabidopsis development. J. Exp. Bot. 2009, 61, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Hwang, G.; Kim, S.; Thi, T.N.; Kim, H.; Jeong, J.; Oh, E. The epidermis coordinates thermoresponsive growth through the phyB-PIF4-auxin pathway. Nat. Commun. 2020, 11, 1053. [Google Scholar] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Number | Annotation | Category | Reference |
|---|---|---|---|
| AT2G18790 | PHYB | Red light photoreceptor | [6] |
| AT4G08920 | CRY1 | Blue light photoreceptor | [7] |
| AT5G53160 | PYL8 | ABA receptor | [8] |
| AT1G01360 | PYL9 | ABA receptor | [8] |
| AT1G75080 | BZR1 | Transcription factor in BR signaling | [9] |
| AT3G44450 | BIC1 | Regulator of CRY activity | [10] |
| AT1G30330 | ARF6 | Transcription factor in auxin signaling | [11] |
| AT2G01570 | RGA | GA signaling repressor | [12] |
| AT2G46340 | SPA1 | PhyA signaling repressor which has Ser/Thr kinase activity | [13] |
| AT2G47890 | BBX11 | B-box family transcription factor | [14] |
| AT2G34640 | HMR | Transcription activator | [15] |
| AT2G25930 | ELF3 | Transcriptional regulator repressing clock- and growth-associated transcription factors to regulate the circadian rhythm and hypocotyl elongation | [16] |
| AT5G61380 | TOC1 | Transcription factor | [17] |
| AT5G24470 | PRR5 | Transcription factor | [17] |
| AT5G02810 | PRR7 | Transcription factor | [17] |
| AT2G46790 | PRR9 | Transcription factor | [17] |
| AT3G50330 | HEC2 | Transcription factor | [18] |
| AT4G37580 | HLS1 | Transcriptional regulator | [19] |
| AT2G40360 | BOP1 | E3 ubiquitin ligase | [20] |
| AT2G41370 | BOP2 | E3 ubiquitin ligase | [20] |
| AT3G28910 | MYB30 | Transcription factor | [21] |
| AT1G43850 | SEU | Transcriptional regulator | [22] |
| AT5G60970 | TCP5 | Transcription factor | [23] |
| AT3G02150 | TCP13 | Transcription factor | [23] |
| AT5G08070 | TCP17 | Transcription factor | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Zhu, Z. PIF4 and PIF4-Interacting Proteins: At the Nexus of Plant Light, Temperature and Hormone Signal Integrations. Int. J. Mol. Sci. 2021, 22, 10304. https://doi.org/10.3390/ijms221910304
Xu Y, Zhu Z. PIF4 and PIF4-Interacting Proteins: At the Nexus of Plant Light, Temperature and Hormone Signal Integrations. International Journal of Molecular Sciences. 2021; 22(19):10304. https://doi.org/10.3390/ijms221910304
Chicago/Turabian StyleXu, Yang, and Ziqiang Zhu. 2021. "PIF4 and PIF4-Interacting Proteins: At the Nexus of Plant Light, Temperature and Hormone Signal Integrations" International Journal of Molecular Sciences 22, no. 19: 10304. https://doi.org/10.3390/ijms221910304
APA StyleXu, Y., & Zhu, Z. (2021). PIF4 and PIF4-Interacting Proteins: At the Nexus of Plant Light, Temperature and Hormone Signal Integrations. International Journal of Molecular Sciences, 22(19), 10304. https://doi.org/10.3390/ijms221910304