Genetic Restrictive Cardiomyopathy: Causes and Consequences—An Integrative Approach



Abstract
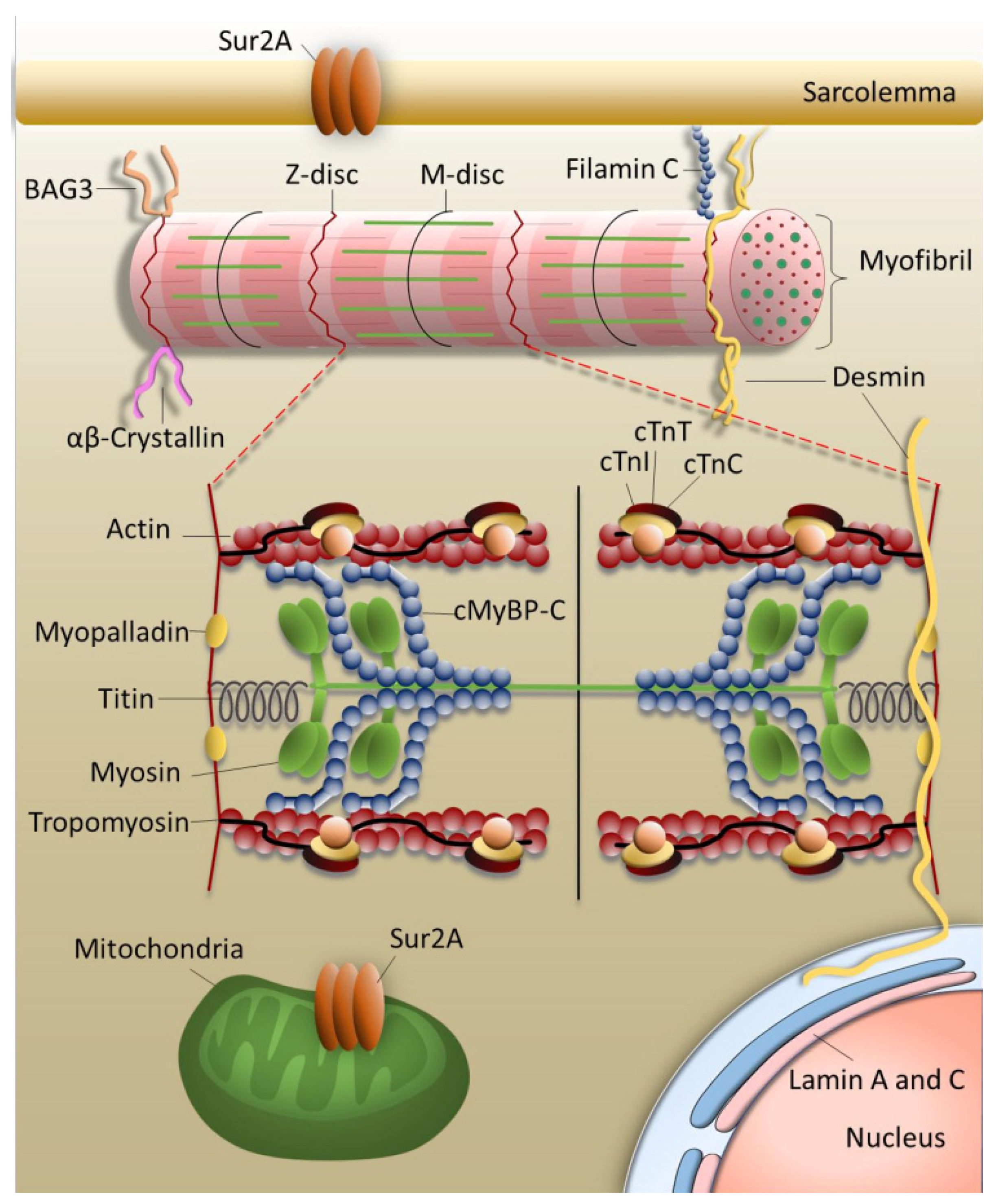
:1. The Sarcomere
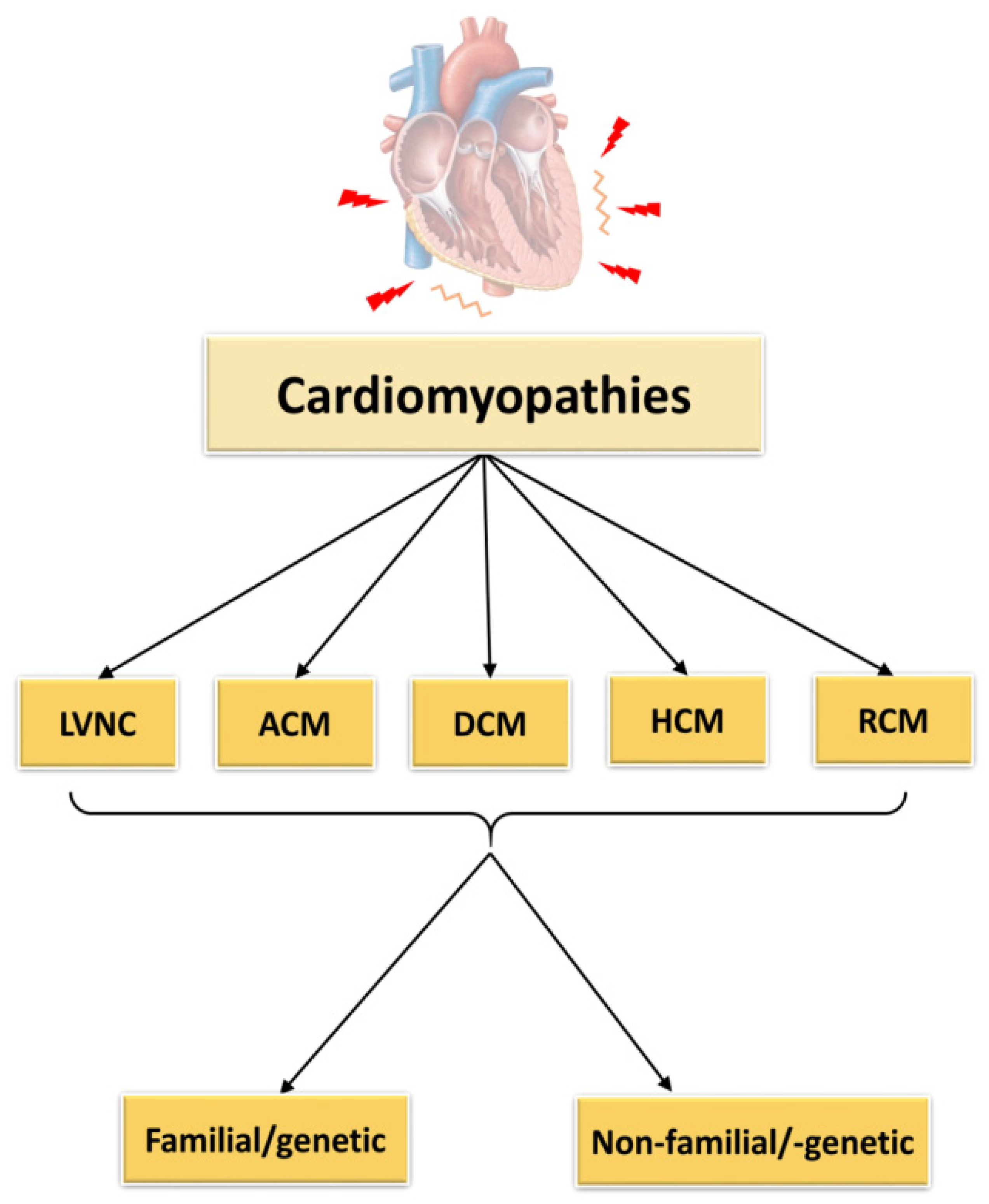
2. Cardiomyopathies
2.1. Left Ventricular Non-Compaction Cardiomyopathy
2.2. Arrhythmogenic Cardiomyopathy
2.3. Dilated Cardiomyopathy
2.4. Hypertrophic Cardiomyopathy
2.5. Restrictive Cardiomyopathy
Pediatric RCM
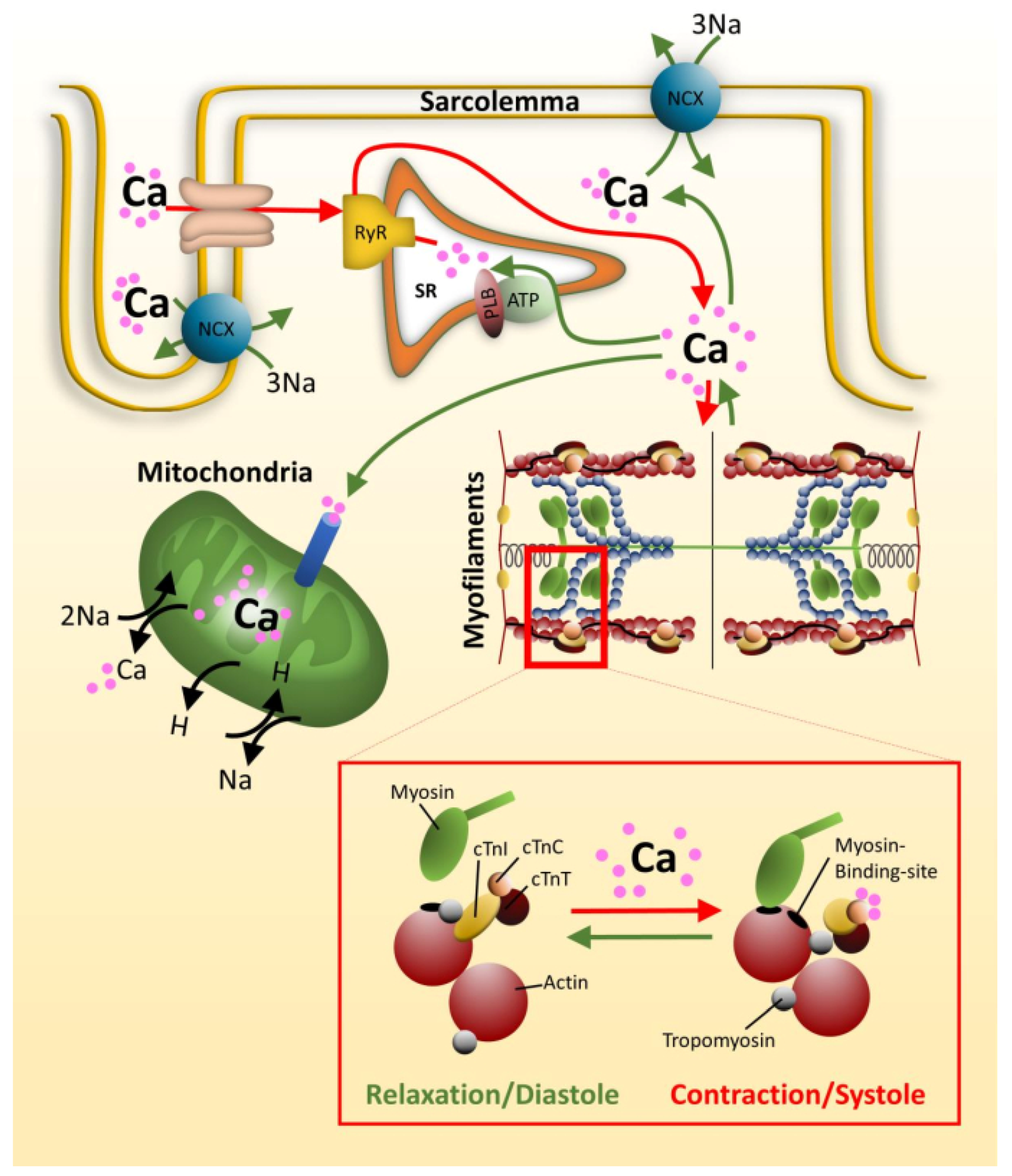
3. Molecular Mechanisms in RCM
3.1. Contractile Dysfunction
3.1.1. Calcium Signaling
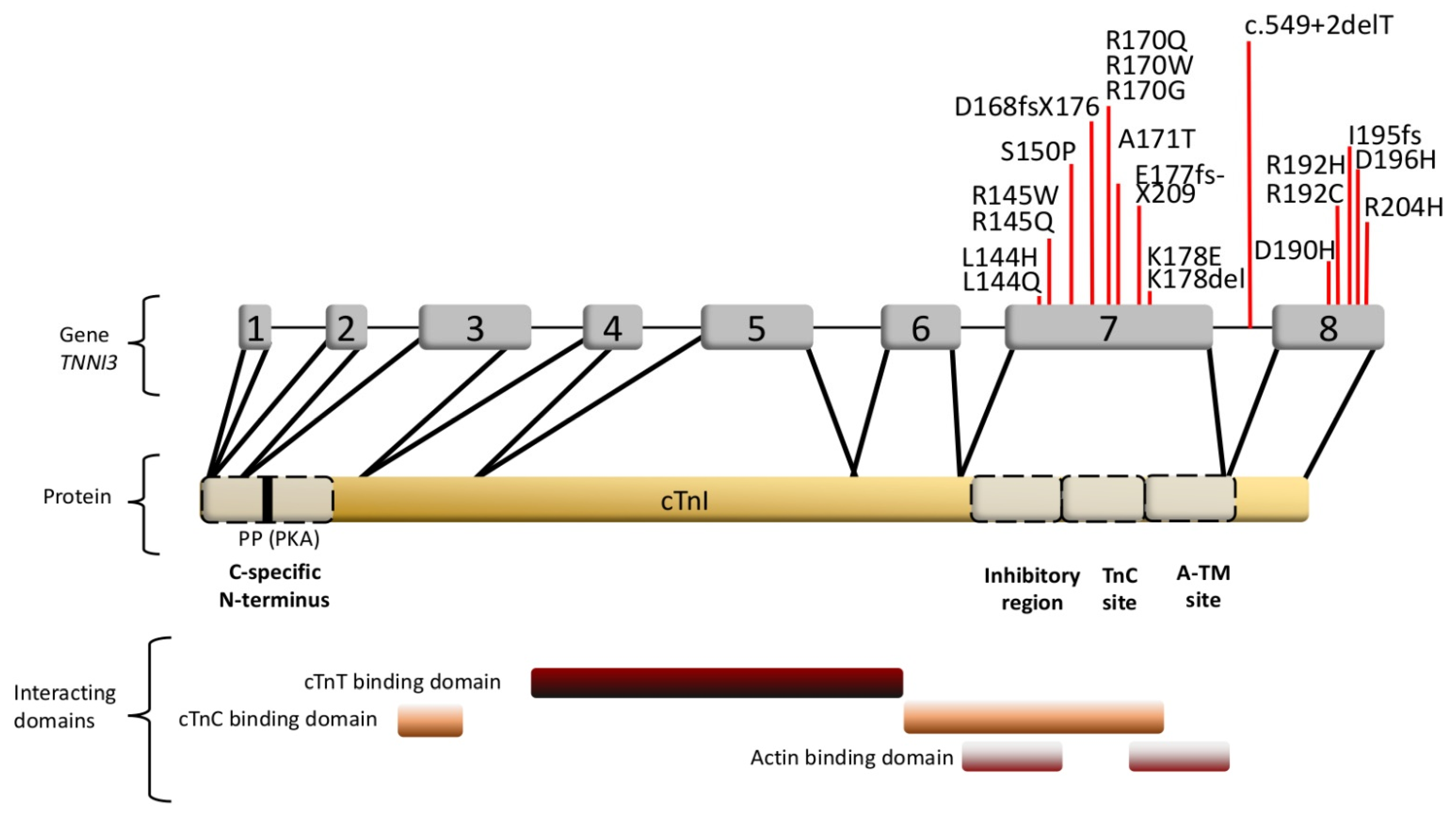
3.1.2. Cardiac Troponin I and the Interplay of Sarcomeric Proteins in RCM
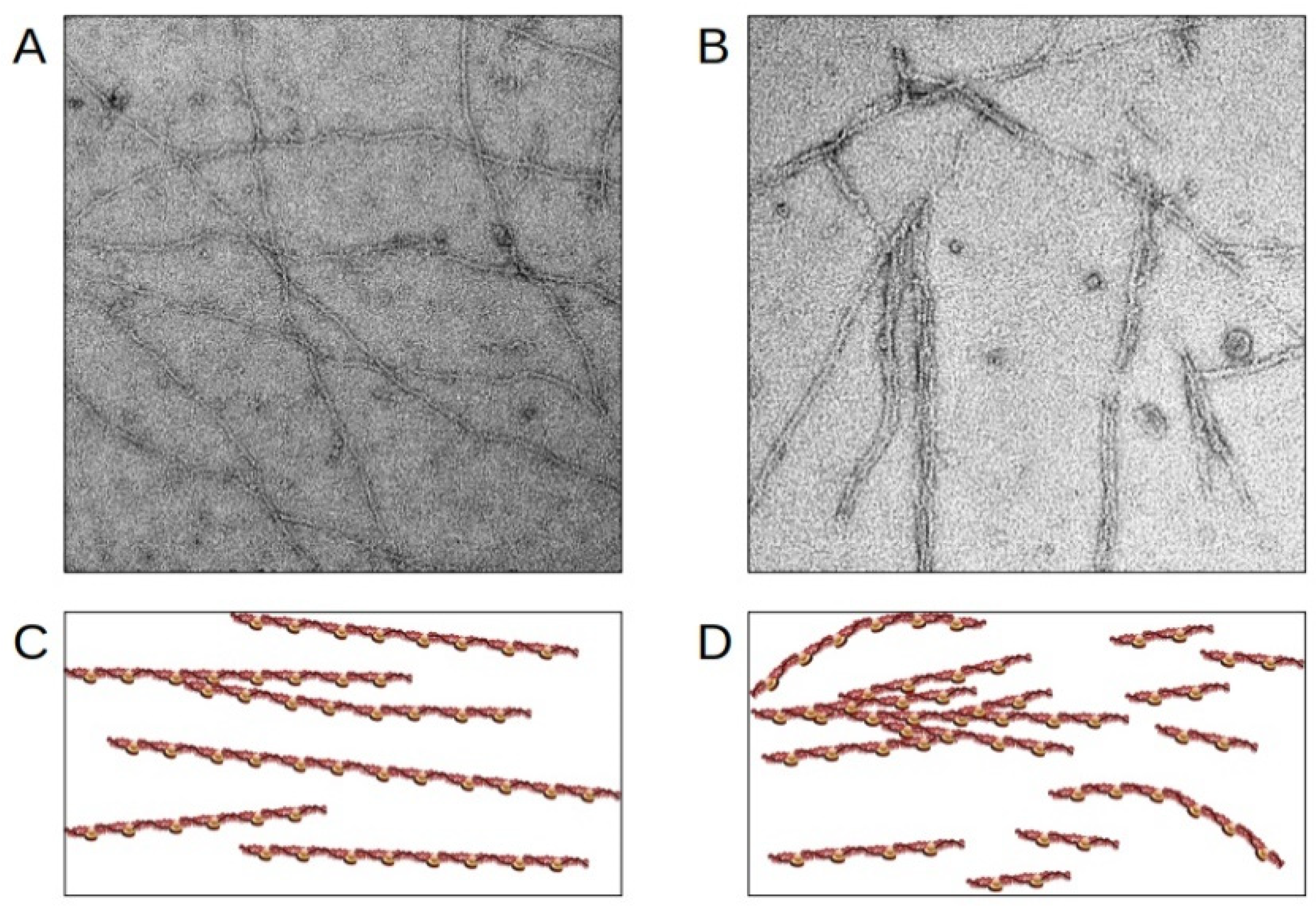
3.2. Protein Aggregation
3.3. Gene Expression and Mosaicism
4. Problems and Prospects in RCM
4.1. Diversity and Diagnostic Disparity of Cardiomyopathies
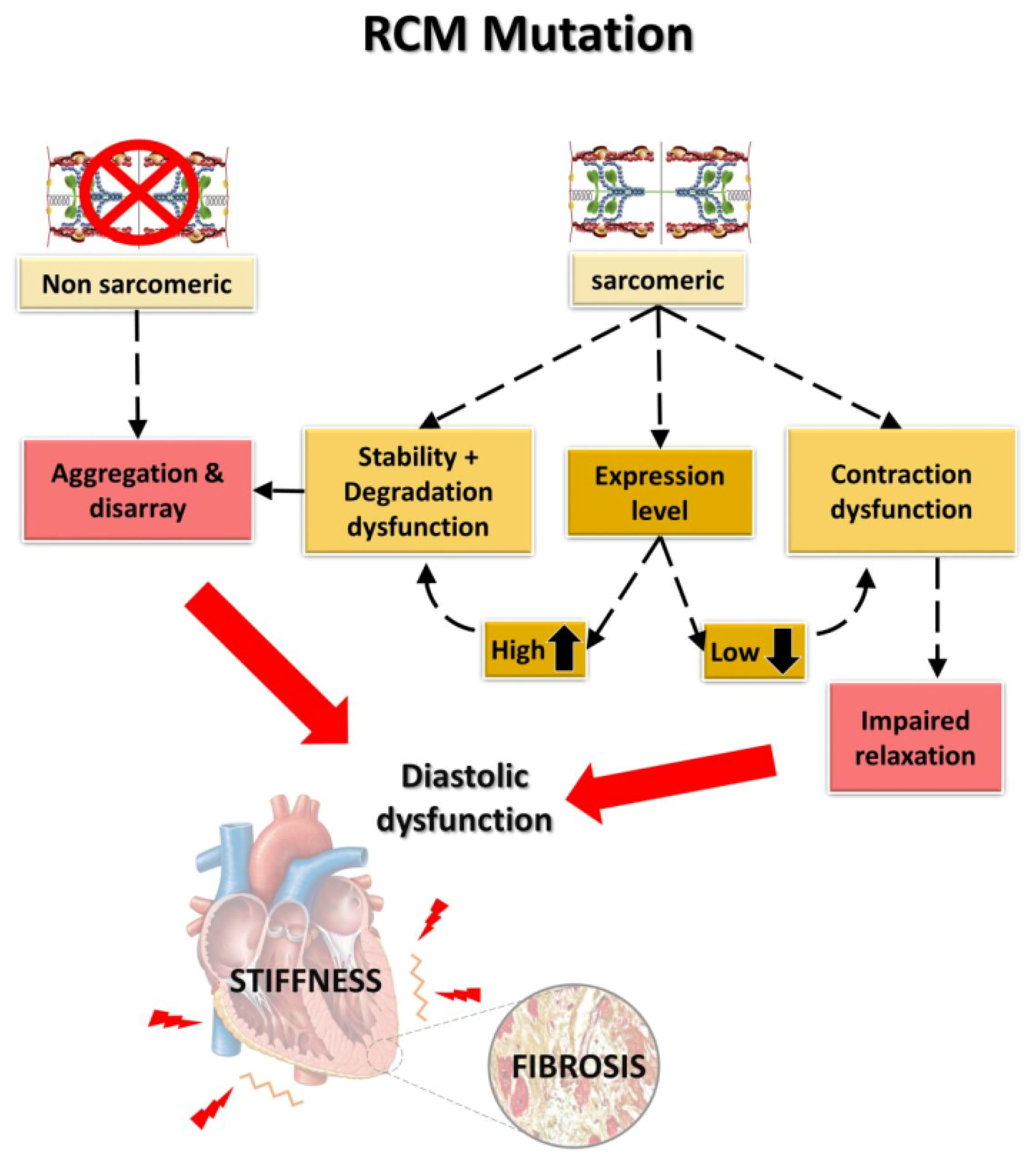
4.2. Development of the RCM Phenotype: An Integrated Approach
4.3. Therapeutic Options
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanson, J.; Huxley, H.E. Structural Basis of the Cross-Striations in Muscle. Nature 1953, 172, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Huxley, A.F.; Niedergerke, R. Structural Changes in Muscle during Contraction. Nature 1954, 173, 971–973. [Google Scholar] [CrossRef] [PubMed]
- Elliott, P.; Andersson, B.; Arbustini, E.; Bilinska, Z.; Cecchi, F.; Charron, P.; Dubourg, O.; Kühl, U.; Maisch, B.; McKenna, W.J.; et al. Classification of the Cardiomyopathies: A Position Statement from the European Society of Cardiology Working Group on Myocardial and Pericardial Diseases. Eur. Heart J. 2008, 29, 270–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nugent, A.W.; Daubeney, P.E.F.; Chondros, P.; Carlin, J.B.; Colan, S.D.; Cheung, M.; Davis, A.M.; Chow, C.W.; Weintraub, R.G. Clinical Features and Outcomes of Childhood Hypertrophic Cardiomyopathy: Results from a National Population-Based Study. Circulation 2005, 112, 1332–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferies, J.L.; Wilkinson, J.D.; Sleeper, L.A.; Colan, S.D.; Lu, M.; Pahl, E.; Kantor, P.F.; Everitt, M.D.; Webber, S.A.; Kaufman, B.D.; et al. Cardiomyopathy Phenotypes and Outcomes for Children With Left Ventricular Myocardial Noncompaction: Results From the Pediatric Cardiomyopathy Registry. J. Card. Fail. 2015, 21, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Moric-Janiszewska, E.; Markiewicz-Łoskot, G. Genetic Heterogeneity of Left Ventricular Noncompaction Cardiomyopathy. Clin. Cardiol. 2008, 31, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Miszalski-Jamka, K.; Jefferies, J.L.; Mazur, W.; Głowacki, J.; Hu, J.; Lazar, M.; Gibbs, R.A.; Liczko, J.; Kłyś, J.; Venner, E.; et al. Novel Genetic Triggers and Genotype-Phenotype Correlations in Patients with Left Ventricular Noncompaction. Circ. Cardiovasc. Genet. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- England, J.; Pang, K.L.; Parnall, M.; Haig, M.I.; Loughna, S. Cardiac Troponin T Is Necessary for Normal Development in the Embryonic Chick Heart. J. Anat. 2016, 229, 436–449. [Google Scholar] [CrossRef] [Green Version]
- Groeneweg, J.A.; Bhonsale, A.; James, C.A.; Te Riele, A.S.; Dooijes, D.; Tichnell, C.; Murray, B.; Wiesfeld, A.C.P.; Sawant, A.C.; Kassamali, B.; et al. Clinical Presentation, Long-Term Follow-Up, and Outcomes of 1001 Arrhythmogenic Right Ventricular Dysplasia/ Cardiomyopathy Patients and Family Members. Circ. Cardiovasc. Genet. 2015, 8, 437–446. [Google Scholar] [CrossRef]
- Beffagna, G.; Zorzi, A.; Pilichou, K.; Perazzolo Marra, M.; Rigato, I.; Corrado, D.; Migliore, F.; Rampazzo, A.; Bauce, B.; Basso, C.; et al. Arrhythmogenic Cardiomyopathy. Eur. Heart J. 2020, 22, 1147–1148. [Google Scholar] [CrossRef]
- Corrado, D.; Basso, C.; Judge, D.P. Arrhythmogenic Cardiomyopathy. Circ. Res. 2017, 121, 785–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merner, N.D.; Hodgkinson, K.A.; Haywood, A.F.M.; Connors, S.; French, V.M.; Drenckhahn, J.D.; Kupprion, C.; Ramadanova, K.; Thierfelder, L.; McKenna, W.; et al. Arrhythmogenic Right Ventricular Cardiomyopathy Type 5 Is a Fully Penetrant, Lethal Arrhythmic Disorder Caused by a Missense Mutation in the TMEM43 Gene. Am. J. Hum. Genet. 2008, 82, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Quarta, G.; Syrris, P.; Ashworth, M.; Jenkins, S.; Zuborne Alapi, K.; Morgan, J.; Muir, A.; Pantazis, A.; McKenna, W.J.; Elliott, P.M. Mutations in the Lamin A/C Gene Mimic Arrhythmogenic Right Ventricular Cardiomyopathy. Eur. Heart J. 2012, 33, 1128–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rijsingen, I.A.W.; Van Der Zwaag, P.A.; Groeneweg, J.A.; Nannenberg, E.A.; Jongbloed, J.D.H.; Zwinderman, A.H.; Pinto, Y.M.; Lekanne Dit Deprez, R.H.; Post, J.G.; Tan, H.L.; et al. Outcome in Phospholamban R14del Carriers Results of a Large Multicentre Cohort Study. Circ. Cardiovasc. Genet. 2014, 7, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Veerman, C.C.; Wilde, A.A.M.; Lodder, E.M. The Cardiac Sodium Channel Gene SCN5A and Its Gene Product NaV1.5: Role in Physiology and Pathophysiology. Gene 2015, 573, 177–187. [Google Scholar] [CrossRef]
- Taylor, M.; Graw, S.; Sinagra, G.; Barnes, C.; Slavov, D.; Brun, F.; Pinamonti, B.; Salcedo, E.E.; Sauer, W.; Pyxaras, S.; et al. Genetic Variation in Titin in Arrhythmogenic Right Ventricular Cardiomyopathy-Overlap Syndromes. Circulation 2011, 124, 876–885. [Google Scholar] [CrossRef] [Green Version]
- Brun, F.; Barnes, C.V.; Sinagra, G.; Slavov, D.; Barbati, G.; Zhu, X.; Graw, S.L.; Spezzacatene, A.; Pinamonti, B.; Merlo, M.; et al. Titin and Desmosomal Genes in the Natural History of Arrhythmogenic Right Ventricular Cardiomyopathy. J. Med. Genet. 2014, 51, 669–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oz, S.; Yonath, H.; Visochyk, L.; Ofek, E.; Landa, N.; Reznik-Wolf, H.; Ortiz-Genga, M.; Monserrat, L.; Ben-Gal, T.; Goitein, O.; et al. Reduction in Filamin C Transcript Is Associated with Arrhythmogenic Cardiomyopathy in Ashkenazi Jews. Int. J. Cardiol. 2020, 317, 133–138. [Google Scholar] [CrossRef]
- Hall, C.L.; Akhtar, M.M.; Sabater-Molina, M.; Futema, M.; Asimaki, A.; Protonotarios, A.; Dalageorgou, C.; Pittman, A.M.; Suarez, M.P.; Aguilera, B.; et al. Filamin C Variants Are Associated with a Distinctive Clinical and Immunohistochemical Arrhythmogenic Cardiomyopathy Phenotype. Int. J. Cardiol. 2020, 307, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asimaki, A.; Syrris, P.; Wichter, T.; Matthias, P.; Saffitz, J.E.; McKenna, W.J. A Novel Dominant Mutation in Plakoglobin Causes Arrhythmogenic Right Ventricular Cardiomyopathy. Am. J. Hum. Genet. 2007, 81, 964–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heide, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, R.M.; Fidler, L. Right Ventricular Cardiomyopathy in the Young: An Emerging Challenge. Hear. Rhythm 2009, 6, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Kriebel, T.; Korte, T.; Kandolf, R.; Jux, C.; Windhagen-Mahnert, B.; Bökenkamp, R.; Bertram, H.; Paul, T. Arrhythmogene Rechtsventrikuläre Dysplasie/Kardiomyopathie -Diagnostik Im Kindesalter. Z. Kardiol. 2003, 92, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Dungan, W.T.; Garson, A.; Gillette, P.C. Arrhythmogenic Right Ventricular Dysplasia: A Cause of Ventricular Tachycardia in Children with Apparently Normal Hearts. Am. Heart J. 1981, 102, 745–750. [Google Scholar] [CrossRef]
- Pinto, Y.M.; Elliott, P.M.; Arbustini, E.; Adler, Y.; Anastasakis, A.; Böhm, M.; Duboc, D.; Gimeno, J.; De Groote, P.; Imazio, M.; et al. Proposal for a Revised Definition of Dilated Cardiomyopathy, Hypokinetic Non-Dilated Cardiomyopathy, and Its Implications for Clinical Practice: A Position Statement of the ESC Working Group on Myocardial and Pericardial Diseases. Eur. Heart J. 2016, 37, 1850–1858. [Google Scholar] [CrossRef] [Green Version]
- Fadl, S.; Wåhlander, H.; Fall, K.; Cao, Y.; Sunnegårdh, J. The Highest Mortality Rates in Childhood Dilated Cardiomyopathy Occur during the First Year after Diagnosis. Acta Paediatr. Int. J. Paediatr. 2018, 107, 672–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipshultz, S.E.; Cochran, T.R.; Briston, D.A.; Brown, S.R.; Sambatakos, P.J.; Miller, T.L.; Carrillo, A.A.; Corcia, L.; Sanchez, J.E.; Diamond, M.B.; et al. Pediatric Cardiomyopathies: Causes, Epidemiology, Clinical Course, Preventive Strategies and Therapies. Future Cardiol. 2013, 9, 817–848. [Google Scholar] [CrossRef] [Green Version]
- Burkett, E.L.; Hershberger, R.E. Clinical and Genetic Issues in Familial Dilated Cardiomyopathy. J. Am. Coll. Cardiol. 2005, 45, 969–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershberger, R.E.; Hedges, D.J.; Morales, A. Dilated Cardiomyopathy: The Complexity of a Diverse Genetic Architecture. Nat. Rev. Cardiol. 2013, 10, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Serra, A.; Toro, R.; Sarquella-Brugada, G.; de Gonzalo-Calvo, D.; Cesar, S.; Carro, E.; Llorente-Cortes, V.; Iglesias, A.; Brugada, J.; Brugada, R.; et al. Genetic Basis of Dilated Cardiomyopathy. Int. J. Cardiol. 2016, 224, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-J.; Chen, C.-S.; Yiang, G.-T.; Tsai, A.P.-Y.; Liao, W.-T.; Wu, M.-Y. Advanced Evolution of Pathogenesis Concepts in Cardiomyopathies. J. Clin. Med. 2019, 8, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, J.; Frese, K.S.; Peil, B.; Kloos, W.; Keller, A.; Nietsch, R.; Feng, Z.; Müller, S.; Kayvanpour, E.; Vogel, B.; et al. Atlas of the Clinical Genetics of Human Dilated Cardiomyopathy. Eur. Heart J. 2015, 36, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Hinson, J.T.; Chopra, A.; Nafissi, N.; Polacheck, W.J.; Benson, C.C.; Swist, S.; Gorham, J.; Yang, L.; Schafer, S.; Sheng, C.C.; et al. Titin Mutations in IPS Cells Define Sarcomere Insufficiency as a Cause of Dilated Cardiomyopathy. Science 2015, 349, 982–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franaszczyk, M.; Chmielewski, P.; Truszkowska, G.; Stawinski, P.; Michalak, E.; Rydzanicz, M.; Sobieszczanska-Malek, M.; Pollak, A.; Szczygieł, J.; Kosinska, J.; et al. Titin Truncating Variants in Dilated Cardiomyopathy -Prevalence and Genotype-Phenotype Correlations. PLoS ONE 2017, 12, e0169007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigli, M.; Begay, R.L.; Morea, G.; Graw, S.L.; Sinagra, G.; Taylor, M.R.G.; Granzier, H.; Mestroni, L. A Review of the Giant Protein Titin in Clinical Molecular Diagnostics of Cardiomyopathies. Front. Cardiovasc. Med. 2016, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansweijer, J.A.; Nieuwhof, K.; Russo, F.; Hoorntje, E.T.; Jongbloed, J.D.H.; Lekanne Deprez, R.H.; Postma, A.V.; Bronk, M.; van Rijsingen, I.A.W.; de Haij, S.; et al. Truncating Titin Mutations Are Associated with a Mild and Treatable Form of Dilated Cardiomyopathy. Eur. J. Heart Fail. 2017, 19, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Herrero Galán, E.; Dominguez, F.; Martinez-Martin, I.; Sanchez-Gonzalez, C.; Vicente, N.; Lalaguna, L.; Bonzon-Kulichenko, E.; Calvo, E.; Gonzalez-Lopez, E.; Cobo-Marcos, M.; et al. Conserved Cysteines in Titin Sustain the Mechanical Function of Cardiomyocytes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Maron, B.J.; Rowin, E.J.; Casey, S.A.; Haas, T.S.; Chan, R.H.M.; Udelson, J.E.; Garberich, R.F.; Lesser, J.R.; Appelbaum, E.; Manning, W.J.; et al. Risk Stratification and Outcome of Patients with Hypertrophic Cardiomyopathy ≥60 Years of Age. Circulation 2013, 127, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Maron, B.J.; Rowin, E.J.; Casey, S.A.; Link, M.S.; Lesser, J.R.; Chan, R.H.M.; Garberich, R.F.; Udelson, J.E.; Maron, M.S. Hypertrophic Cardiomyopathy in Adulthood Associated with Low Cardiovascular Mortality with Contemporary Management Strategies. J. Am. Coll. Cardiol. 2015, 65, 1915–1928. [Google Scholar] [CrossRef] [Green Version]
- Aro, A.L.; Nair, S.G.; Reinier, K.; Jayaraman, R.; Stecker, E.C.; Uy-Evanado, A.; Rusinaru, C.; Jui, J.; Chugh, S.S. Population Burden of Sudden Death Associated with Hypertrophic Cardiomyopathy. Circulation 2017, 136, 1665–1667. [Google Scholar] [CrossRef]
- Teekakirikul, P.; Zhu, W.; Huang, H.C.; Fung, E. Hypertrophic Cardiomyopathy: An Overview of Genetics and Management. Biomolecules 2019, 9, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marian, A.J.; Wu, Y.; Lim, D.S.; McCluggage, M.; Youker, K.; Yu, Q.T.; Brugada, R.; DeMayo, F.; Quinones, M.; Roberts, R. A Transgenic Rabbit Model for Human Hypertrophic Cardiomyopathy. J. Clin. Investig. 1999, 104, 1683–1692. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, S.J.; Dooijes, D.; Dos Remedios, C.; Michels, M.; Lamers, J.M.J.; Winegrad, S.; Schlossarek, S.; Carrier, L.; Cate, F.J.T.; Stienen, G.J.M.; et al. Cardiac Myosin-Binding Protein C Mutations and Hypertrophic Cardiomyopathy. Circulation 2009, 119, 1473–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidman, C.E.; Seidman, J.G. Identifying Sarcomere Gene Mutations in Hypertrophic Cardiomyopathy: A Personal History. Circ. Res. 2011, 108, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schober, T.; Huke, S.; Venkataraman, R.; Gryshchenko, O.; Kryshtal, D.; Hwang, H.S.; Baudenbacher, F.J.; Knollmann, B.C. Myofilament Ca Sensitization Increases Cytosolic Ca Binding Affinity, Alters Intracellular Ca Homeostasis, and Causes Pause-Dependent Ca-Triggered Arrhythmia. Circ. Res. 2012, 111, 170–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knollmann, B.C.; Kirchhof, P.; Sirenko, S.G.; Degen, H.; Greene, A.E.; Schober, T.; Mackow, J.C.; Fabritz, L.; Potter, J.D.; Morad, M. Familial Hypertrophic Cardiomyopathy-Linked Mutant Troponin T Causes Stress-Induced Ventricular Tachycardia and Ca2+-Dependent Action Potential Remodeling. Circ. Res. 2003, 92, 428–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sequeira, V.; Najafi, A.; Wijnker, P.J.M.; Dos Remedios, C.G.; Michels, M.; Kuster, D.W.D.; Van Der Velden, J. ADP-Stimulated Contraction: A Predictor of Thin-Filament Activation in Cardiac Disease. Proc. Natl. Acad. Sci. USA 2015, 112, E7003–E7012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sequeira, V.; Bertero, E.; Maack, C. Energetic Drain Driving Hypertrophic Cardiomyopathy. FEBS Lett. 2019, 593, 1616–1626. [Google Scholar] [CrossRef] [Green Version]
- Tariq, M. Importance of Genetic Evaluation and Testing in Pediatric Cardiomyopathy. World J. Cardiol. 2014, 6, 1156. [Google Scholar] [CrossRef]
- Nihoyannopoulos, P.; Dawson, D. Restrictive Cardiomyopathies. Eur. J. Echocardiogr. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Yokochi, T. Transthyretin Cardiac Amyloidosis: An Update on Diagnosis and Treatment. ESC Hear. Fail. 2019, 6, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, J.; Kubo, T.; Duque, M.; Uribe, W.; Shaw, A.; Murphy, R.; Gimeno, J.R.; Elliott, P.; McKenna, W.J. Idiopathic Restrictive Cardiomyopathy Is Part of the Clinical Expression of Cardiac Troponin I Mutations. J. Clin. Investig. 2003, 111, 209–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego-Delgado, M.; Delgado, J.F.; Brossa-Loidi, V.; Palomo, J.; Marzoa-Rivas, R.; Perez-Villa, F.; Salazar-Mendiguchía, J.; Ruiz-Cano, M.J.; Gonzalez-Lopez, E.; Padron-Barthe, L.; et al. Idiopathic Restrictive Cardiomyopathy Is Primarily a Genetic Disease. J. Am. Coll. Cardiol. 2016, 67, 3021–3023. [Google Scholar] [CrossRef] [PubMed]
- Kaski, J.P.; Syrris, P.; Burch, M.; Tomé Esteban, M.T.; Fenton, M.; Christiansen, M.; Andersen, P.S.; Sebire, N.; Ashworth, M.; Deanfield, J.E.; et al. Idiopathic Restrictive Cardiomyopathy in Children Is Caused by Mutations in Cardiac Sarcomere Protein Genes. Heart 2008, 94, 1478–1484. [Google Scholar] [CrossRef]
- Kostareva, A.; Kiselev, A.; Gudkova, A.; Frishman, G.; Ruepp, A.; Frishman, D.; Smolina, N.; Tarnovskaya, S.; Nilsson, D.; Zlotina, A.; et al. Genetic Spectrum of Idiopathic Restrictive Cardiomyopathy Uncovered by Next-Generation Sequencing. PLoS ONE 2016, 11, e0163362. [Google Scholar] [CrossRef] [Green Version]
- Pruszczyk, P.; Kostera-Pruszczyk, A.; Shatunov, A.; Goudeau, B.; Dramiñska, A.; Takeda, K.; Sambuughin, N.; Vicart, P.; Strelkov, S.V.; Goldfarb, L.G.; et al. Restrictive Cardiomyopathy with Atrioventricular Conduction Block Resulting from a Desmin Mutation. Int. J. Cardiol. 2007, 117, 244–253. [Google Scholar] [CrossRef]
- Brodehl, A.; Hakimi, S.A.P.; Stanasiuk, C.; Ratnavadivel, S.; Hendig, D.; Gaertner, A.; Gerull, B.; Gummert, J.; Paluszkiewicz, L.; Milting, H. Restrictive Cardiomyopathy Is Caused by a Novel Homozygous Desmin (DES) Mutation p.Y122H Leading to a Severe Filament Assembly Defect. Genes 2019, 10, 918. [Google Scholar] [CrossRef] [Green Version]
- Ojrzyńska, N.; Bilińska, Z.T.; Franaszczyk, M.; Płoski, R.; Grzybowski, J. Restrictive Cardiomyopathy Due to Novel Desmin Gene Mutation. Kardiol. Pol. 2017, 75, 723. [Google Scholar] [CrossRef] [Green Version]
- Kiselev, A.; Vaz, R.; Knyazeva, A.; Khudiakov, A.; Tarnovskaya, S.; Liu, J.; Sergushichev, A.; Kazakov, S.; Frishman, D.; Smolina, N.; et al. De Novo Mutations in FLNC Leading to Early-Onset Restrictive Cardiomyopathy and Congenital Myopathy. Hum. Mutat. 2018, 39, 1161–1172. [Google Scholar] [CrossRef]
- Brodehl, A.; Gaertner-Rommel, A.; Klauke, B.; Grewe, S.A.; Schirmer, I.; Peterschröder, A.; Faber, L.; Vorgerd, M.; Gummert, J.; Anselmetti, D.; et al. The Novel AB-Crystallin (CRYAB) Mutation p.D109G Causes Restrictive Cardiomyopathy. Hum. Mutat. 2017, 38, 947–952. [Google Scholar] [CrossRef]
- Gomes, A.V.; Liang, J.; Potter, J.D. Mutations in Human Cardiac Troponin I That Are Associated with Restrictive Cardiomyopathy Affect Basal ATPase Activity and the Calcium Sensitivity of Force Development. J. Biol. Chem. 2005, 280, 30909–30915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.; Wen, H.; Edwards, T.; Metzger, J.M. Allele and Species Dependent Contractile Defects by Restrictive and Hypertrophic Cardiomyopathy-Linked Troponin I Mutants. J. Mol. Cell. Cardiol. 2008, 44, 891–904. [Google Scholar] [CrossRef] [Green Version]
- Mouton, J.; Pellizzon, A.; Goosen, A.; Kinnear, C.; Herbst, P.; Brink, P.; Moolman-Smook, J. Diagnostic Disparity and Identification of Two TNNI3 Gene Mutations, One Novel and One Arising de Novo, in South African Patients with Restrictive Cardiomyopathy and Focal Ventricular Hypertrophy. Cardiovasc. J. Afr. 2015, 26, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Wijngaard, A.; Volders, P.; van Tintelen, J.P.; Jongbloed, J.D.H.; van den Berg, M.P.; Lekanne Deprez, R.H.; Mannens, M.M.A.M.; Hofmann, N.; Slegtenhorst, M.; Dooijes, D.; et al. Recurrent and Founder Mutations in the Netherlands: Cardiac Troponin I (TNNI3) Gene Mutations as a Cause of Severe Forms of Hypertrophic and Restrictive Cardiomyopathy. Netherlands Hear. J. 2011, 19, 344–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.P.; Lu, C.X.; Zhao, X.Y.; Liang, R.J.; Lian, H.; Routledge, M.; Wu, W.; Zhang, X.; Fan, Z.J. Restrictive Cardiomyopathy Resulting from a Troponin I Type 3 Mutation in a Chinese Family. Chin. Med. Sci. J. 2016, 31, 1–7. [Google Scholar] [CrossRef]
- Kostareva, A.; Gudkova, A.; Sjöberg, G.; Mörner, S.; Semernin, E.; Krutikov, A.; Shlyakhto, E.; Sejersen, T. Deletion in TNNI3 Gene Is Associated with Restrictive Cardiomyopathy. Int. J. Cardiol. 2009, 131, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Cimiotti, D.; Fujita-Becker, S.; Möhner, D.; Smolina, N.; Budde, H.; Wies, A.; Morgenstern, L.; Gudkova, A.; Sejersen, T.; Sjöberg, G.; et al. Infantile Restrictive Cardiomyopathy: cTnIR170G/W Impair the Interplay of Sarcomeric Proteins and the Integrity of Thin Filaments. PLoS ONE 2020, 15, e0229227. [Google Scholar] [CrossRef]
- Kaski, J.P.; Syrris, P.; Esteban, M.T.T.; Jenkins, S.; Pantazis, A.; Deanfield, J.E.; McKenna, W.J.; Elliott, P.M. Prevalence of Sarcomere Protein Gene Mutations in Preadolescent Children with Hypertrophic Cardiomyopathy. Circ. Cardiovasc. Genet. 2009, 2, 436–441. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Yogasundaram, H.; Basu, R.; Wang, F.; Paterson, D.I.; Alastalo, T.P.; Oudit, G.Y. Novel Dominant-Negative Mutation in Cardiac Troponin i Causes Severe Restrictive Cardiomyopathy. Circ. Hear. Fail. 2017, 10. [Google Scholar] [CrossRef]
- Pantou, M.P.; Gourzi, P.; Gkouziouta, A.; Armenis, I.; Kaklamanis, L.; Zygouri, C.; Constantoulakis, P.; Adamopoulos, S.; Degiannis, D. A Case Report of Recessive Restrictive Cardiomyopathy Caused by a Novel Mutation in Cardiac Troponin I (TNNI3). BMC Med. Genet. 2019, 20, 1–6. [Google Scholar] [CrossRef]
- Gambarin, F.I.; Tagliani, M.; Arbustini, E. Pure Restrictive Cardiomyopathy Associated with Cardiac Troponin I Gene Mutation: Mismatch between the Lack of Hypertrophy and the Presence of Disarray. Heart 2008, 94, 1257. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.W.; Hitz, M.P.; Andelfinger, G. Ventricular Septal Defect and Restrictive Cardiomyopathy in a Paediatric TNNI3 Mutation Carrier. Cardiol. Young 2010, 20, 574–576. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.C.; Michels, V.V.; Pellikka, P.A.; Ballew, J.D.; Karst, M.L.; Herron, K.J.; Nelson, S.M.; Rodeheffer, R.J.; Olson, T.M. Cardiac Troponin T Mutation in Familial Cardiomyopathy with Variable Remodeling and Restrictive Physiology. Clin. Genet. 2008, 74, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, T.; Szczesna, D.; Housmans, P.R.; Zhao, J.; De Freitas, F.; Gomes, A.V.; Culbreath, L.; McCue, J.; Wang, Y.; Xu, Y.; et al. Abnormal Contractile Function in Transgenic Mice Expressing a Familial Hypertrophic Cardiomyopathy-Linked Troponin T (I79N) Mutation. J. Biol. Chem. 2001, 276, 3743–3755. [Google Scholar] [CrossRef] [Green Version]
- Peddy, S.B.; Vricella, L.A.; Crosson, J.E.; Oswald, G.L.; Cohn, R.D.; Cameron, D.E.; Valle, D.; Loeys, B.L. Infantile Restrictive Cardiomyopathy Resulting from a Mutation in the Cardiac Troponin T Gene. Pediatrics 2006, 117, 1830–1833. [Google Scholar] [CrossRef]
- Pinto, J.R.; Parvatiyar, M.S.; Jones, M.A.; Liang, J.; Potter, J.D. A Troponin T Mutation That Causes Infantile Restrictive Cardiomyopathy Increases Ca2+ Sensitivity of Force Development and Impairs the Inhibitory Properties of Troponin. J. Biol. Chem. 2008, 283, 2156–2166. [Google Scholar] [CrossRef] [Green Version]
- Ploski, R.; Rydzanicz, M.; Ksiazczyk, T.M.; Franaszczyk, M.; Pollak, A.; Kosinska, J.; Michalak, E.; Stawinski, P.; Ziolkowska, L.; Bilinska, Z.T.; et al. Evidence for Troponin C (TNNC1) as a Gene for Autosomal Recessive Restrictive Cardiomyopathy with Fatal Outcome in Infancy. Am. J. Med. Genet. Part A 2016, 170, 3241–3248. [Google Scholar] [CrossRef]
- Pinto, J.R.; Parvatiyar, M.S.; Jones, M.A.; Liang, J.; Ackerman, M.J.; Potter, J.D. A Functional and Structural Study of Tropon in C Mutations Related to Hypertrophic Cardiomyopathy. J. Biol. Chem. 2009, 284, 19090–19100. [Google Scholar] [CrossRef] [Green Version]
- Pinto, J.R.; Reynaldo, D.P.; Parvatiyar, M.S.; Dweck, D.; Liang, J.; Jones, M.A.; Sorenson, M.M.; Potter, J.D. Strong Cross-Bridges Potentiate the Ca2+ Affinity Changes Produced by Hypertrophic Cardiomyopathy Cardiac Troponin C Mutants in Myofilaments: A Fast Kinetic Approach. J. Biol. Chem. 2011, 286, 1005–1013. [Google Scholar] [CrossRef] [Green Version]
- Dorsch, L.M.; Kuster, D.W.D.; Jongbloed, J.D.H.; Boven, L.G.; van Spaendonck-Zwarts, K.Y.; Suurmeijer, A.J.H.; Vink, A.; du Marchie Sarvaas, G.J.; van den Berg, M.P.; van der Velden, J.; et al. The Effect of Tropomyosin Variants on Cardiomyocyte Function and Structure That Underlie Different Clinical Cardiomyopathy Phenotypes. Int. J. Cardiol. 2021, 323, 251–258. [Google Scholar] [CrossRef]
- Peled, Y.; Gramlich, M.; Yoskovitz, G.; Feinberg, M.S.; Afek, A.; Polak-Charcon, S.; Pras, E.; Sela, B.A.; Konen, E.; Weissbrod, O.; et al. Titin Mutation in Familial Restrictive Cardiomyopathy. Int. J. Cardiol. 2014, 171, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Greenway, S.C.; Wilson, G.J.; Wilson, J.; George, K.; Kantor, P.F. Sudden Death in an Infant with Angina, Restrictive Cardiomyopathy, and Coronary Artery Bridging. An Unusual Phenotype for a β-Myosin Heavy Chain (MYH7) Sarcomeric Protein Mutation. Circ. Hear. Fail. 2012, 5, e92–e93. [Google Scholar]
- Neagoe, O.; Ciobanu, A.; Diaconu, R.; Mirea, O.; Donoiu, I.; Militaru, C. A Rare Case of Familial Restrictive Cardiomyopathy with Mutations in MYH7 and ABCC9 Genes. Discoveries 2019, 7, e99. [Google Scholar] [CrossRef]
- Fan, L.L.; Guo, S.; Jin, J.Y.; He, Z.J.; Zhao, S.P.; Xiang, R.; Zhao, W. Whole Exome Sequencing Identified a 13 Base Pair MYH7 Deletion-Mutation in a Patient with Restrictive Cardiomyopathy and Left Ventricle Hypertrophy. Ann. Clin. Lab. Sci. 2019, 49, 838–840. [Google Scholar]
- Ware, S.M.; Quinn, M.E.; Ballard, E.T.; Miller, E.; Uzark, K.; Spicer, R.L. Pediatric Restrictive Cardiomyopathy Associated with a Mutation in β-Myosin Heavy Chain. Clin. Genet. 2008, 73, 165–170. [Google Scholar] [CrossRef]
- Karam, S.; Raboisson, M.J.; Ducreux, C.; Chalabreysse, L.; Millat, G.; Bozio, A.; Bouvagnet, P. A de Novo Mutation of the Beta Cardiac Myosin Heavy Chain Gene in an Infantile Restrictive Cardiomyopathy. Congenit. Heart Dis. 2008, 3, 138–143. [Google Scholar] [CrossRef]
- Wu, W.; Lu, C.X.; Wang, Y.N.; Liu, F.; Chen, W.; Liu, Y.T.; Han, Y.C.; Cao, J.; Zhang, S.Y.; Zhang, X. Novel Phenotype-Genotype Correlations of Restrictive Cardiomyopathy with Myosin-Binding Protein C (MYBPC3) Gene Mutations Tested by next-Generation Sequencing. J. Am. Heart Assoc. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Bahrudin, U.; Morisaki, H.; Morisaki, T.; Ninomiya, H.; Higaki, K.; Nanba, E.; Igawa, O.; Takashima, S.; Mizuta, E.; Miake, J.; et al. Ubiquitin-Proteasome System Impairment Caused by a Missense Cardiac Myosin-Binding Protein C Mutation and Associated with Cardiac Dysfunction in Hypertrophic Cardiomyopathy. J. Mol. Biol. 2008, 384, 896–907. [Google Scholar] [CrossRef] [Green Version]
- Brodehl, A.; Ferrier, R.A.; Hamilton, S.J.; Greenway, S.C.; Brundler, M.A.; Yu, W.; Gibson, W.T.; Mckinnon, M.L.; Mcgillivray, B.; Alvarez, N.; et al. Mutations in FLNC Are Associated with Familial Restrictive Cardiomyopathy. Hum. Mutat. 2016, 37, 269–279. [Google Scholar] [CrossRef]
- Roldán-Sevilla, A.; Palomino-Doza, J.; de Juan, J.; Sánchez, V.; Domínguez-González, C.; Salguero-Bodes, R.; Arribas-Ynsaurriaga, F. Missense Mutations in the FLNC Gene Causing Familial Restrictive Cardiomyopathy. Circ. Genom. Precis. Med. 2019, 12, e002388. [Google Scholar] [CrossRef] [PubMed]
- Tucker, N.R.; McLellan, M.A.; Hu, D.; Ye, J.; Parsons, V.A.; Mills, R.W.; Clauss, S.; Dolmatova, E.; Shea, M.A.; Milan, D.J.; et al. Novel Mutation in FLNC (Filamin C) Causes Familial Restrictive Cardiomyopathy. Circ. Cardiovasc. Genet. 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, J.; Tariq, M.; Geddes, G.; Kindel, S.; Miller, E.M.; Ware, S.M. Novel Pathogenic Variants in Filamin C Identified in Pediatric Restrictive Cardiomyopathy. Hum. Mutat. 2018, 39, 2083–2096. [Google Scholar] [CrossRef] [PubMed]
- Purevjav, E.; Arimura, T.; Augustin, S.; Huby, A.C.; Takagi, K.; Nunoda, S.; Kearney, D.L.; Taylor, M.D.; Terasaki, F.; Bos, J.M.; et al. Molecular Basis for Clinical Heterogeneity in Inherited Cardiomyopathies Due to Myopalladin Mutations. Hum. Mol. Genet. 2012, 21, 2039–2053. [Google Scholar] [CrossRef] [Green Version]
- Huby, A.C.; Mendsaikhan, U.; Takagi, K.; Martherus, R.; Wansapura, J.; Gong, N.; Osinska, H.; James, J.F.; Kramer, K.; Saito, K.; et al. Disturbance in Z-Disk Mechanosensitive Proteins Induced by a Persistent Mutant Myopalladin Causes Familial Restrictive Cardiomyopathy. J. Am. Coll. Cardiol. 2014, 64, 2765–2776. [Google Scholar] [CrossRef] [Green Version]
- Paller, M.S.; Martin, C.M.; Pierpont, M.E. Restrictive Cardiomyopathy: An Unusual Phenotype of a Lamin A Variant. ESC Hear. Fail. 2018, 5, 724–726. [Google Scholar] [CrossRef]
- Schänzer, A.; Rupp, S.; Gräf, S.; Zengeler, D.; Jux, C.; Akintürk, H.; Gulatz, L.; Mazhari, N.; Acker, T.; Van Coster, R.; et al. Dysregulated Autophagy in Restrictive Cardiomyopathy Due to Pro209Leu Mutation in BAG3. Mol. Genet. Metab. 2018, 123, 388–399. [Google Scholar] [CrossRef]
- Ranthe, M.F.; Carstensen, L.; Øyen, N.; Jensen, M.K.; Axelsson, A.; Wohlfahrt, J.; Melbye, M.; Bundgaard, H.; Boyd, H.A. Risk of Cardiomyopathy in Younger Persons with a Family History of Death from Cardiomyopathy: A Nationwide Family Study in a Cohort of 3.9 Million Persons. Circulation 2015, 132, 1013–1019. [Google Scholar] [CrossRef] [Green Version]
- Wittekind, S.G.; Ryan, T.D.; Gao, Z.; Zafar, F.; Czosek, R.J.; Chin, C.W.; Jefferies, J.L. Contemporary Outcomes of Pediatric Restrictive Cardiomyopathy: A Single-Center Experience. Pediatr. Cardiol. 2019, 40, 694–704. [Google Scholar] [CrossRef]
- Webber, S.A.; Lipshultz, S.E.; Sleeper, L.A.; Lu, M.; Wilkinson, J.D.; Addonizio, L.J.; Canter, C.E.; Colan, S.D.; Everitt, M.D.; Jefferies, J.L.; et al. Outcomes of Restrictive Cardiomyopathy in Childhood and the Influence of Phenotype: A Report from the Pediatric Cardiomyopathy Registry. Circulation 2012, 126, 1237–1244. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, J.; Arbustini, E. Restrictive Cardiomyopathy. Curr. Opin. Cardiol. 2009, 24, 214–220. [Google Scholar] [CrossRef]
- Mogensen, J.; Hey, T.; Lambrecht, S. A Systematic Review of Phenotypic Features Associated With Cardiac Troponin I Mutations in Hereditary Cardiomyopathies. Can. J. Cardiol. 2015, 31, 1377–1385. [Google Scholar] [CrossRef]
- Ding, W.H.; Han, L.; Xiao, Y.Y.; Mo, Y.; Yang, J.; Wang, X.F.; Jin, M. Role of Whole-Exome Sequencing in Phenotype Classification and Clinical Treatment of Pediatric Restrictive Cardiomyopathy. Chin. Med. J. 2017, 130, 2823–2828. [Google Scholar] [CrossRef] [PubMed]
- Veltman, J.A.; Brunner, H.G. De Novo Mutations in Human Genetic Disease. Nat. Rev. Genet. 2012, 13, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, N.; Kooij, V.; Van Dijk, S.; Merkus, D.; Paulus, W.J.; Dos Remedios, C.; Duncker, D.J.; Stienen, G.J.M.; Van Der Velden, J. Sarcomeric Dysfunction in Heart Failure. Cardiovasc. Res. 2008, 77, 649–658. [Google Scholar] [CrossRef]
- Yumoto, F.; Lu, Q.W.; Morimoto, S.; Tanaka, H.; Kono, N.; Nagata, K.; Ojima, T.; Takahashi-Yanaga, F.; Miwa, Y.; Sasaguri, T.; et al. Drastic Ca2+ Sensitization of Myofilament Associated with a Small Structural Change in Troponin I in Inherited Restrictive Cardiomyopathy. Biochem. Biophys. Res. Commun. 2005, 338, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Solaro, R.J. Increased Ca2+ affinity of Cardiac Thin Filaments Reconstituted with Cardiomyopathy-Related Mutant Cardiac Troponin I. J. Biol. Chem. 2006, 281, 13471–13477. [Google Scholar] [CrossRef] [Green Version]
- Sparrow, A.J.; Sievert, K.; Patel, S.; Chang, Y.F.; Broyles, C.N.; Brook, F.A.; Watkins, H.; Geeves, M.A.; Redwood, C.S.; Robinson, P.; et al. Measurement of Myofilament-Localized Calcium Dynamics in Adult Cardiomyocytes and the Effect of Hypertrophic Cardiomyopathy Mutations. Circ. Res. 2019, 124, 1228–1239. [Google Scholar] [CrossRef]
- Frazier, A.H.; Ramirez-Correa, G.A.; Murphy, A.M. Molecular Mechanisms of Sarcomere Dysfunction in Dilated and Hypertrophic Cardiomyopathy. Prog. Pediatr. Cardiol. 2011, 31, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Priori, S.G.; Blomstrom-Lundqvist, C.; Mazzanti, A.; Bloma, N.; Borggrefe, M.; Camm, J.; Elliott, P.M.; Fitzsimons, D.; Hatala, R.; Hindricks, G.; et al. 2015 ESC Guidelines for the Management of Patients with Ventricular Arrhythmias and the Prevention of Sudden Cardiac Death the Task Force for the Management of Patients with Ventricular Arrhythmias and the Prevention of Sudden Cardiac Death of the Europea. Eur. Heart J. 2015, 36, 2793–2867l. [Google Scholar] [CrossRef] [Green Version]
- Rivenes, S.M.; Kearney, D.L.; Smith, E.O.; Towbin, J.A.; Denfield, S.W. Sudden Death and Cardiovascular Collapse in Children with Restrictive Cardiomyopathy. Circulation 2000, 102, 876–882. [Google Scholar] [CrossRef]
- Denfield, S.W.; Rosenthal, G.; Gajarski, R.J.; Bricker, J.T.; Schowengerdt, K.O.; Price, J.K.; Towbin, J.A. Restrictive Cardiomyopathies in Childhood: Etiologies and Natural History. Texas Hear. Inst. J. 1997, 24, 38–44. [Google Scholar]
- Linke, W.A. Sense and Stretchability: The Role of Titin and Titin-Associated Proteins in Myocardial Stress-Sensing and Mechanical Dysfunction. Cardiovasc. Res. 2008, 77, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Kooij, V.; Saes, M.; Jaquet, K.; Zaremba, R.; Foster, D.B.; Murphy, A.M.; dos Remedios, C.G.; van der Velden, J.; Stienen, G.J.M. Effect of Troponin I Ser23/24 Phosphorylation on Ca2+ -Sensitivity in Human Myocardium Depends on the Phosphorylation Background. J. Mol. Cell. Cardiol. 2010, 48, 954–963. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Schmidtmann, A.; Redlich, A.; Westerdorf, B.; Jaquet, K.; Thieleczek, R. Effects of Phosphorylation and Mutation R145G on Human Cardiac Troponin I Function. Biochemistry 2001, 40, 14593–14602. [Google Scholar] [CrossRef]
- Dvornikov, A.V.; Smolin, N.; Zhang, M.; Martin, J.L.; Robia, S.L.; De Tombe, P.P. Restrictive Cardiomyopathy Troponin I R145W Mutation Does Not Perturb Myofilament Length-Dependent Activation in Human Cardiac Sarcomeres. J. Biol. Chem. 2016, 291, 21817–21828. [Google Scholar] [CrossRef] [Green Version]
- Westfall, M.V. Contribution of Post-Translational Phosphorylation to Sarcomere-Linked Cardiomyopathy Phenotypes. Front. Physiol. 2016, 7, 407. [Google Scholar] [CrossRef] [Green Version]
- Tadros, H.J.; Life, C.S.; Garcia, G.; Pirozzi, E.; Jones, E.G.; Datta, S.; Parvatiyar, M.S.; Chase, P.B.; Allen, H.D.; Kim, J.J.; et al. Meta-Analysis of Cardiomyopathy-Associated Variants in Troponin Genes Identifies Loci and Intragenic Hot Spots That Are Associated with Worse Clinical Outcomes. J. Mol. Cell. Cardiol. 2020, 142, 118–125. [Google Scholar] [CrossRef]
- Takeda, S.; Yamashita, A.; Maeda, K.; Maéda, Y. Structure of the Core Domain of Human Cardiac Troponin in the Ca(2+)-Saturated Form. Nature 2003, 424, 35–41. [Google Scholar] [CrossRef]
- Metzger, J.M.; Westfall, M.V. Covalent and Noncovalent Modification of Thin Filament Action: The Essential Role of Troponin in Cardiac Muscle Regulation. Circ. Res. 2004, 94, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Meriin, A.B.; Narayanan, A.; Meng, L.; Alexandrov, I.; Varelas, X.; Cissé, I.I.; Sherman, M.Y. Hsp70–Bag3 Complex Is a Hub for Proteotoxicity-Induced Signaling That Controls Protein Aggregation. Proc. Natl. Acad. Sci. USA 2018, 115, E7043–E7052. [Google Scholar] [CrossRef] [Green Version]
- Rauch, J.N.; Tse, E.; Freilich, R.; Mok, S.A.; Makley, L.N.; Southworth, D.R.; Gestwicki, J.E. BAG3 Is a Modular, Scaffolding Protein That Physically Links Heat Shock Protein 70 (Hsp70) to the Small Heat Shock Proteins. J. Mol. Biol. 2017, 429, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Dimauro, I.; Antonioni, A.; Mercatelli, N.; Caporossi, D. The Role of AB-Crystallin in Skeletal and Cardiac Muscle Tissues. Cell Stress Chaperones 2018, 23, 491–505. [Google Scholar] [CrossRef]
- Islam, M.; Diwan, A.; Mani, K. Come Together: Protein Assemblies, Aggregates and the Sarcostat at the Heart of Cardiac Myocyte Homeostasis. Front. Physiol. 2020, 11, 586. [Google Scholar] [CrossRef]
- Schlossarek, S.; Englmann, D.R.; Sultan, K.R.; Sauer, M.; Eschenhagen, T.; Carrier, L. Defective Proteolytic Systems in MYBPC3-Targeted Mice with Cardiac Hypertrophy. Basic Res. Cardiol. 2012, 107, 1–13. [Google Scholar] [CrossRef]
- Salman, O.F.; El-Rayess, H.M.; Abi Khalil, C.; Nemer, G.; Refaat, M.M. Inherited Cardiomyopathies and the Role of Mutations in Non-Coding Regions of the Genome. Front. Cardiovasc. Med. 2018, 5, 77. [Google Scholar] [CrossRef]
- Frisso, G.; Detta, N.; Coppola, P.; Mazzaccara, C.; Pricolo, M.R.; D’Onofrio, A.; Limongelli, G.; Calabrò, R.; Salvatore, F. Functional Studies and in Silico Analyses to Evaluate Non-Coding Variants in Inherited Cardiomyopathies. Int. J. Mol. Sci. 2016, 17, 1883. [Google Scholar] [CrossRef] [Green Version]
- Lander, J.; Ware, S.M. Copy Number Variation in Congenital Heart Defects. Curr. Genet. Med. Rep. 2014, 2, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Singer, E.S.; Ross, S.B.; Skinner, J.R.; Weintraub, R.G.; Ingles, J.; Semsarian, C.; Bagnall, R.D. Characterization of Clinically Relevant Copy-Number Variants from Exomes of Patients with Inherited Heart Disease and Unexplained Sudden Cardiac Death. Genet. Med. 2020, 1–8. [Google Scholar] [CrossRef]
- Franaszczyk, M.; Truszkowska, G.; Chmielewski, P.; Rydzanicz, M.; Kosinska, J.; Rywik, T.; Biernacka, A.; Spiewak, M.; Kostrzewa, G.; Stepien-Wojno, M.; et al. Analysis of De Novo Mutations in Sporadic Cardiomyopathies Emphasizes Their Clinical Relevance and Points to Novel Candidate Genes. J. Clin. Med. 2020, 9, 370. [Google Scholar] [CrossRef] [Green Version]
- Ceyhan-Birsoy, O.; Pugh, T.J.; Bowser, M.J.; Hynes, E.; Frisella, A.L.; Mahanta, L.M.; Lebo, M.S.; Amr, S.S.; Funke, B.H. Next Generation Sequencing-Based Copy Number Analysis Reveals Low Prevalence of Deletions and Duplications in 46 Genes Associated with Genetic Cardiomyopathies. Mol. Genet. Genom. Med. 2016, 4, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Helms, A.S.; Davis, F.M.; Coleman, D.; Bartolone, S.N.; Glazier, A.A.; Pagani, F.; Yob, J.M.; Sadayappan, S.; Pedersen, E.; Lyons, R.; et al. Sarcomere Mutation-Specific Expression Patterns in Human Hypertrophic Cardiomyopathy. Circ. Cardiovasc. Genet. 2014, 7, 434–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, S.; Schultz, I.; Becker, E.; Montag, J.; Borchert, B.; Francino, A.; Navarro-Lopez, F.; Perrot, A.; Özcelik, C.; Osterziel, K.J.; et al. Unequal Allelic Expression of Wild-Type and Mutated β-Myosin in Familial Hypertrophic Cardiomyopathy. Basic Res. Cardiol. 2011, 106, 1041–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montag, J.; Kowalski, K.; Makul, M.; Ernstberger, P.; Radocaj, A.; Beck, J.; Becker, E.; Tripathi, S.; Keyser, B.; Mühlfeld, C.; et al. Burst-like Transcription of Mutant and Wildtype MYH7-Alleles as Possible Origin of Cell-to-Cell Contractile Imbalance in Hypertrophic Cardiomyopathy. Front. Physiol. 2018, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Parbhudayal, R.Y.; Garra, A.R.; Götte, M.J.W.; Michels, M.; Pei, J.; Harakalova, M.; Asselbergs, F.W.; van Rossum, A.C.; van der Velden, J.; Kuster, D.W.D. Variable Cardiac Myosin Binding Protein-C Expression in the Myofilaments Due to MYBPC3 Mutations in Hypertrophic Cardiomyopathy. J. Mol. Cell. Cardiol. 2018, 123, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, L.; Jean-Charles, P.-Y.; Nan, C.; Chen, G.; Tian, J.; Jin, J.-P.; Gelb, I.J.; Huang, X. Dose-Dependent Diastolic Dysfunction and Early Death in a Mouse Model with Cardiac Troponin Mutations. J. Mol. Cell. Cardiol. 2013, 62, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Kraft, T.; Montag, J.; Radocaj, A.; Brenner, B. Hypertrophic Cardiomyopathy: Cell-to-Cell Imbalance in Gene Expression and Contraction Force as Trigger for Disease Phenotype Development. Circ. Res. 2016, 119, 992–995. [Google Scholar] [CrossRef] [Green Version]
- Kraft, T.; Montag, J. Altered Force Generation and Cell-to-Cell Contractile Imbalance in Hypertrophic Cardiomyopathy. Pflugers Arch. Eur. J. Physiol. 2019, 471, 719–733. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and Guidelines for the Interpretation of Sequence Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Burke, M.A.; Cook, S.A.; Seidman, J.G.; Seidman, C.E. Clinical and Mechanistic Insights into the Genetics of Cardiomyopathy. J. Am. Coll. Cardiol. 2016, 68, 2871–2886. [Google Scholar] [CrossRef]
- Maron, B.J.; Towbin, J.A.; Thiene, G.; Antzelevitch, C.; Corrado, D.; Arnett, D.; Moss, A.J.; Seidman, C.E.; Young, J.B. Contemporary Definitions and Classification of the Cardiomyopathies. Circulation 2006, 113, 1807–1816. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.V.; Potter, J.D. Cellular and Molecular Aspects of Familial Hypertrophic Cardiomyopathy Caused by Mutations in the Cardiac Troponin I Gene. Mol. Cell. Biochem. 2004, 263, 99–114. [Google Scholar] [CrossRef]
- Kimura, A.; Harada, H.; Park, J.E.; Nishi, H.; Satoh, M.; Takahashi, M.; Hiroi, S.; Sasaoka, T.; Ohbuchi, N.; Nakamura, T.; et al. Mutations in the Cardiac Troponin I Gene Associated with Hypertrophic Cardiomyopathy. Nat. Genet. 1997, 17, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.W.; Jang, M.A.; Jang, S.Y.; Seo, S.H.; Seong, M.W.; Park, S.S.; Ki, C.S.; Kim, D.K. Diverse Phenotypic Expression of Cardiomyopathies in a Family with TNNI3 p.Arg145Trp Mutation. Korean Circ. J. 2017, 47, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, P.A.; Leinwand, L.A. Cellular Mechanisms of Cardiomyopathy. J. Cell Biol. 2011, 194, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semsarian, C.; Healey, M.J.; Fatkin, D.; Giewat, M.; Duffy, C.; Seidman, C.E.; Seidman, J.G. A Polymorphic Modifier Gene Alters the Hypertrophic Response in a Murine Model of Familial Hypertrophic Cardiomyopathy. J. Mol. Cell. Cardiol. 2001, 33, 2055–2060. [Google Scholar] [CrossRef]
- Mosqueira, D.; Mannhardt, I.; Bhagwan, J.R.; Lis-Slimak, K.; Katili, P.; Scott, E.; Hassan, M.; Prondzynski, M.; Harmer, S.C.; Tinker, A.; et al. CRISPR/Cas9 Editing in Human Pluripotent Stemcell-Cardiomyocytes Highlights Arrhythmias, Hypocontractility, and Energy Depletion as Potential Therapeutic Targets for Hypertrophic Cardiomyopathy. Eur. Heart J. 2018, 39, 3879–3892. [Google Scholar] [CrossRef]
- Song, L.; Su, M.; Wang, S.; Zou, Y.; Wang, X.; Wang, Y.; Cui, H.; Zhao, P.; Hui, R.; Wang, J. MiR-451 Is Decreased in Hypertrophic Cardiomyopathy and Regulates Autophagy by Targeting TSC1. J. Cell. Mol. Med. 2014, 18, 2266–2274. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, D.; Li, Y. LncRNAs in Cardiac Hypertrophy: From Basic Science to Clinical Application. J. Cell. Mol. Med. 2020, 24, 11638–11645. [Google Scholar] [CrossRef]
- Zhang, C.J.; Huang, Y.; De Lu, J.; Lin, J.; Ge, Z.R.; Huang, H. Upregulated MicroRNA-132 Rescues Cardiac Fibrosis and Restores Cardiocyte Proliferation in Dilated Cardiomyopathy through the Phosphatase and Tensin Homolog–Mediated PI3K/Akt Signal Transduction Pathway. J. Cell. Biochem. 2019, 120, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Nan, C.; Chen, Y.; Tian, J.; Jean-Charles, P.-Y.; Getfield, C.; Wang, X.; Huang, X. Calcium Desensitizer Catechin Reverses Diastolic Dysfunction in Mice with Restrictive Cardiomyopathy. Arch. Biochem. Biophys. 2015, 573, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.S.; Schwartz, J.H.; Gundapaneni, B.; Elliott, P.M.; Merlini, G.; Waddington-Cruz, M.; Kristen, A.V.; Grogan, M.; Witteles, R.; Damy, T.; et al. Tafamidis Treatment for Patients with Transthyretin Amyloid Cardiomyopathy. N. Engl. J. Med. 2018, 379, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Popkova, T.; Hajek, R.; Jelinek, T. Monoclonal Antibodies in the Treatment of AL Amyloidosis: Co-Targetting the Plasma Cell Clone and Amyloid Deposits. Br. J. Haematol. 2020, 189, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Kovacs, J.J.; McLaurin, A.; Vance, J.M.; Ito, A.; Yao, T.P. The Deacetylase HDAC6 Regulates Aggresome Formation and Cell Viability in Response to Misfolded Protein Stress. Cell 2003, 115, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Koga, H.; Kawaguchi, Y.; Tang, W.; Wong, E.; Gao, Y.S.; Pandey, U.B.; Kaushik, S.; Tresse, E.; Lu, J.; et al. HDAC6 Controls Autophagosome Maturation Essential for Ubiquitin-Selective Quality-Control Autophagy. EMBO J. 2010, 29, 969–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Protein | Variant | Allele Origin | No. of Affected Families (No. Mutation Carriers) | Age at Presentation/Onset (Youngest Patient) | Molecular Effects/Remarks | Ref. # |
|---|---|---|---|---|---|---|
| TNNI3/cTnI | p.L144Q | unknown | - | 17 years | Myofibrillar disarray, Ca2+-sensitization + incr. basal force, red. maximal force (in reconstituted filaments), low ATPase inhibition + maximal activity, decreased incorporation in thin filaments | [52,61,62] |
| p.L144H | familial | 1(3) | 20–30 years | mild hypertrophy | [63] | |
| p.R145W | familial | 2(2) | 19 years | Low ATPase inhibition, Ca2+-sensitization (reconstituted filaments), decreased incorporation in thin filaments | [61,62,64] | |
| p.R145Q | unknown | - | 9 years | Occurred in combination with R192C | [64] | |
| p.S150P | familial | 1(3) | unknown | [65] | ||
| c.549+2delT | De novo | - | <1 year | truncation | [64] | |
| p.D168fsX176 | unknown | 1(1) | 23 years | unknown | [66] | |
| p.R170G | De novo | - | 3 years | Ca2+-sensitization (skinned fibers), thin filament instability, impaired interaction with cMyBP-C and Tpm | [55,67] | |
| p.R170W | De novo | - | 8 months | Ca2+-sensitization (skinned fibers), thin filament instability, impaired cMyBP-C interaction, decreased incorporation in thin filaments | [55,67] | |
| p.R170Q | De novo | - | 15 years | Unknown; also associated with HCM in another study | [63,68] | |
| p.A171T | unknown | - | 63 years | Mild Ca2+-sensitization | [52,61] | |
| p.E177fsX209 | De novo | - | 6 years | disarray | [54] | |
| p.K178E | De novo | - | 3 years | Low ATPase inhibition, Ca2+-sensitization (skinned fibers), increased max. ATPase activity | [52,61] | |
| p.K178del | De novo | - | <11 years | unknown | [64] | |
| p.D190H | familial | 1(13) | 11 years | Ca2+-sensitization (in-vitro ATPase activity), red. cooperativity | [52] | |
| p.R192C | unknown | - | 9 years | Occurred in combination with R145Q | [64] | |
| p.R192H | De novo | - | 16 years | Ca2+-sensitization, red. cooperativity (in-vitro ATPase activity), impaired relaxation, disarray, fibrosis (mouse model), increased incorporation in thin filaments | [52,61] | |
| p.I195fs | De novo | - | 24 years | Mild fibrosis and hypertrophy, no aggregation | [69] | |
| p.D196H | familial | 1(10) | 41 years | 3 members homozygous, heterozygous relatives asymptomatic; fibrosis, mild hypertrophy, no deposits | [70] | |
| p.R204H | De novo | - | 14 years; 3 years | unknown | [71,72] | |
| TNNT2/cTnT | p.I79N | familial | 1(9) | 53 years | No disarray (biopsy); fibrosis; Ca2+-sensitization (mice, skinned fibers); also associated with HCM in other studies | [73,74] |
| p.E96del | De novo | - | 1 year | Disarray, fibrosis (biopsy); Ca2+-sensitization (skinned fibers, in-vitro ATPase activity); impaired inhibition (ATPase), impaired relaxation (skinned fibers), with fetal TnI the effects are less severe | [75,76] | |
| p.E136K | familial | 1(3) | 3.5 years | Myocyte vacuolation, no disarray | [54] | |
| TNNC1/cTnC | p.A8V and p.D145E | familial | 1(4) | 8 months | Compound heterozygosity; HCM to RCM transition; Ca2+-sensitization (fibers), slow Ca2+_off rate, impaired relaxation | [77,78,79] |
| TPM1/Tpm | p.E62Q and p.M281T | familial | 1(11) | 9 years | Compound heterozygosity; disruption of sarcomeres (biopsy), reduced Ca2+ transient amplitudes (HL-1 cells) | [80] |
| TTN/Titin | p.Y7621C | familial | 1(5) | 12 years | Fibrosis, myofilament degradation, Z-disk distortion | [81] |
| MYH7/MHC | p.Y386C | De novo | - | 9 months | Mild fibrosis, no disarray (biopsy) | [82] |
| p.R721K | familial | 1(1) | 43 years | Together with p.Sur2A-R1186Q; arrhythmia | [83] | |
| 13bp del | De novo | - | 49 years | unknown | [84] | |
| p.G768R | familial | 1(2) | 15 months | No tissue abnormalities (biopsy), no disarray or fibrosis | [85] | |
| p.838L | De novo | - | 5 months | Mild disarray, no infiltration (biopsy), arrhythmia | [86] | |
| MYBPC3/cMyBP-C | p.Q463X | familial | 1(3) | 34 years | unknown | [87] |
| p.E334K | De novo | - | 45 years | Increased polyubiquitinylation and degradation (cell model) | [87,88] | |
| DES/desmin | p.Y122H | familial | 1(1) | 19 years | Homozygous; impaired intermediate filament assembly, desmin aggregates (iPSC) | [57] |
| 735+1G>T | familial | 1(1) | 46 years | Myopathy, alternate splicing | [58] | |
| p.E413K | familial | 1(3) | 30 years | Desmin aggregation, granulofilamentous deposits (biopsy); disruption of intermediate filaments, aggregation (cell model) | [56] | |
| FLNC/filamin C | p.A1183L | De novo | - | 6 months | perinuclear aggregates (zebrafish skeletal muscle) | [59] |
| p.A1186V | De novo | - | 1.4 years | Absence of filamin C and desmin in intercalated discs, Z-line streaming (biopsy); perinuclear aggregates (zebrafish skeletal muscle) | [59] | |
| p.S1624L | familial | 1(4) | 3 years | Aggregates, Z-disk disorganization, impaired desmin localization (biopsy, cell model) | [89] | |
| p.G2151S | familial | 1 (2) | 15 years | Sarcomere disorganization, Filamin C deposits (biopsy) | [90] | |
| p.I2160F | familial | 1 (6) | 15 years | disrupted Z-disks, impaired desmin localization (biopsy, cell model) | [89] | |
| p.V2297M | familial | 1 (5) | 44 years | Impaired Filamin C association with sarcomeres (biopsy), reduced contractility (ESC model) | [91] | |
| p.P2298L | familial | 1 (8) | 3 years | No deposits, no sarcomeric disarray (biopsy); perinuclear Filamin C and actin aggregation (cell model) | [92] | |
| p.P2301L | De novo | - | 41 years | Significant fibrosis, no deposits (biopsy) | [90] | |
| p.Y2563C | De novo | - | 1 year | Regular sarcomeric structure, no deposits (biopsy); randomly distributed Filamin C and actin aggregates (cell model) | [92] | |
| MYPN/myopalladin | p.Q529X | familial | 1 (3) | 7 years | Truncated variant without NEBL and a-actinin binding domains; nuclear deformations, myofibrillar degeneration, impaired a-actinin localization, CARP aggregation, Z-disk disruption (biopsy, cell model); nuclear enrichment of truncated Mypn, impaired MAPK signaling, down-regulation of CARP (mice) | [93,94] |
| LMNA/Lamin A | p.E279RfsX201 | familial | 1(2) | 53 years | No infiltration (biopsy) | [95] |
| CRYAB/crystallin αB | p.D109G | familial | 1(2) | 19 years | Protein aggregation, Z-disk disruption (biopsy, cell model) | [60] |
| BAG3/Bag3 | p.P209L | unknown | - | 8 years | Aggregation of BAG3 and desmin, Z-disk alterations, myofibrillar disarray, undegraded autophagosomes, increased autophagy regulators (biopsy) | [96] |
| ABCC9/Sur2A | p.R1186Q | familial | 1(1) | 43 years | Together with p.MHC-R721K; arrhythmia | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cimiotti, D.; Budde, H.; Hassoun, R.; Jaquet, K. Genetic Restrictive Cardiomyopathy: Causes and Consequences—An Integrative Approach. Int. J. Mol. Sci. 2021, 22, 558. https://doi.org/10.3390/ijms22020558
Cimiotti D, Budde H, Hassoun R, Jaquet K. Genetic Restrictive Cardiomyopathy: Causes and Consequences—An Integrative Approach. International Journal of Molecular Sciences. 2021; 22(2):558. https://doi.org/10.3390/ijms22020558
Chicago/Turabian StyleCimiotti, Diana, Heidi Budde, Roua Hassoun, and Kornelia Jaquet. 2021. "Genetic Restrictive Cardiomyopathy: Causes and Consequences—An Integrative Approach" International Journal of Molecular Sciences 22, no. 2: 558. https://doi.org/10.3390/ijms22020558
APA StyleCimiotti, D., Budde, H., Hassoun, R., & Jaquet, K. (2021). Genetic Restrictive Cardiomyopathy: Causes and Consequences—An Integrative Approach. International Journal of Molecular Sciences, 22(2), 558. https://doi.org/10.3390/ijms22020558