Molecular and Genetic Aspects of Grain Number Determination in Rice (Oryza sativa L.)


Abstract
:1. Introduction
2. Panicle Development and GNPP Determination in Rice
2.1. Panical Development
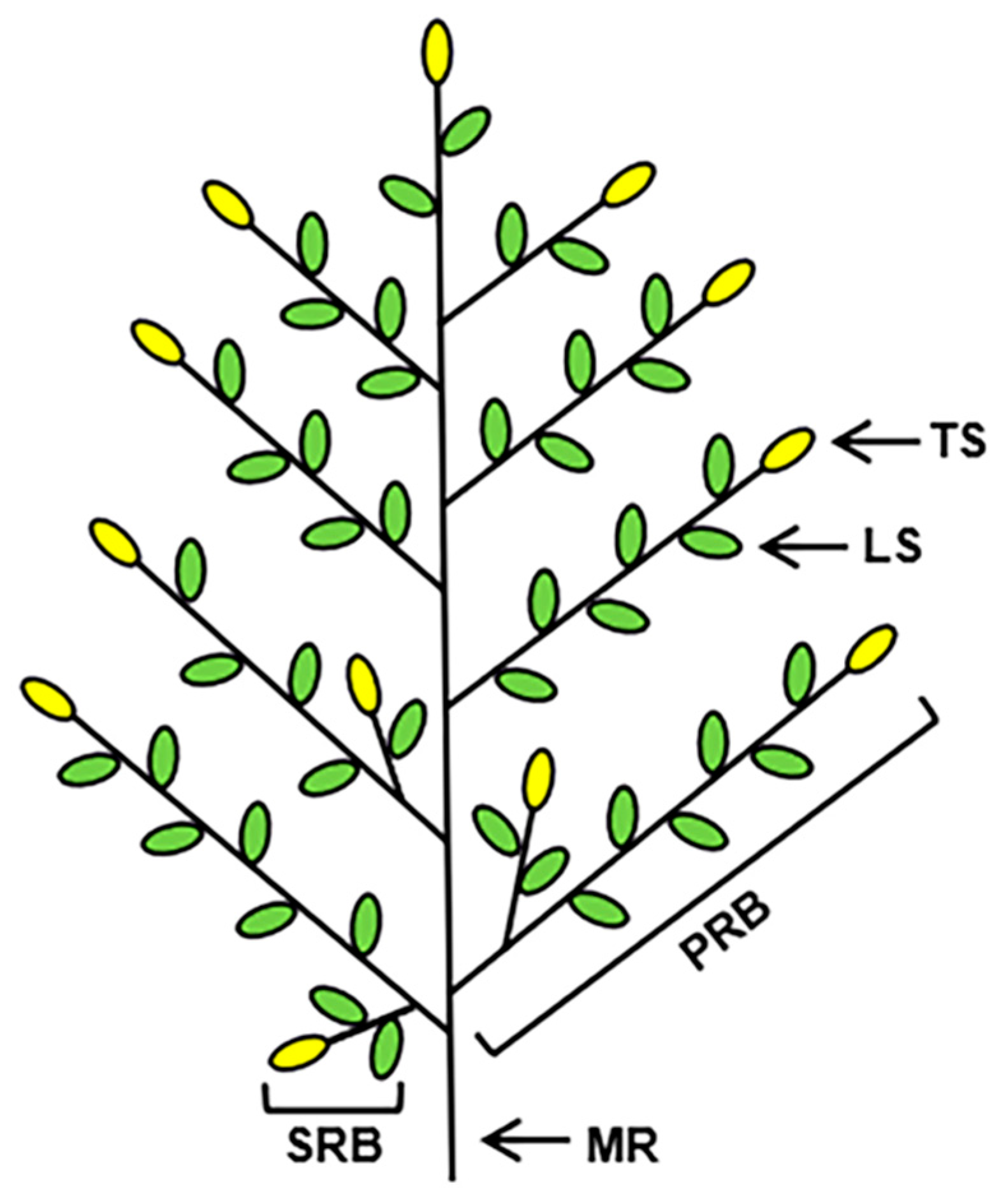
2.2. GNPP Determination in Rice
3. Functional Classification of GNPP-Related Genes and Their Regulation of GNPP
3.1. Rachis Branch Development-Related Genes and Their Regulation of GNPP
3.1.1. Positive Regulation of GNPP by Rachis Branch Development-Related Genes
3.1.2. Negative Regulation of GNPP by Rachis Branch Development-Related Genes
3.2. Phase Transition (Rachis Branch Meristem to Spikelet Meristem)-Related Genes and Their Regulation of GNPP
3.3. Spikelet-Specialisation-Related Genes and Their Regulation of GNPP
4. Other GNPP-Related Genes and Their Regulation of GNPP
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| An-1 | Awn-1 |
| APO1 | ABERRANT PANICLE ORGANIZATION 1 |
| CK | Cytokinin |
| DEP1 | DENSE AND ERECT PANICLE1 |
| DEP3 | DENSE AND ERECT PANICLE3 |
| DST | DROUGHT AND SALT TOLERANCE |
| EP3 | ERECT PANICLE3 |
| FZP | FRIZZY PANICLE |
| FZP2 | FRIZZY PANICLE 2 |
| GA | Gibberellin |
| GAD1 | GRAIN NUMBER, GRAIN LENGTH AND AWN DEVELOPMENT1 |
| Ghd7 | Grain Number, Plant Height, and Heading Date7 |
| Ghd8 | Grain Number, Plant Height, and Heading Date8 |
| GN1a | Grain Number1a |
| GN2 | Grain Number2 |
| GN4-1 | Grain Number4-1 |
| GNP1 | Grain Number per Panicle1 |
| Gnp4 | Grain Number per Panicle Gene4 |
| GNPP | Grain number per panicle |
| HAP | Heterotrimeric haem activator |
| LOG | LONELY GUY |
| IM | Inflorescence meristem |
| IPA1 | Ideal Plant Architecture 1 |
| KNOX | Knotted1-like Homeobox |
| LAX | LAX PANICLE |
| LAX2 | LAX PANICLE2 |
| LP | LARGER PANICLE |
| MOC1 | MONOCULM1 |
| NOG1 | NUMBER OF GRAINS 1 |
| OsCKX2 | Cytokinin oxidase2 |
| OSH1 | Oryza sativa Homeobox1 |
| PAP2 | PANICLE PHYTOMER 2 |
| PAY1 | PLANT ARCHITECTURE AND YIELD1 |
| PROG1 | PROSTRATE GROWTH1 |
| PRR37 | Pseudo-response REGULATOR37 |
| PTR | Peptide transporter |
| PYL | Pyrabactin Resistance-Like |
| QTL | Quantitative trait locus |
| SAM | Shoot apical meristem |
| SRBs | Secondary rachis branches |
| SP1 | Short Panicle 1 |
| SPA | Small Panicle |
| SPL14 | SOUAMOSA PROMOTER BINDING PROTEIN-LIKE 14 |
| TAW1 | TAWAWA1 |
| UTR | Untranslated region |
| WFP | WEALTHY FARMER’S PANICLE |
References
- Demont, M.; Stein, A.J. Global value of GM rice: A review of expected agronomic and consumer benefits. New Biotechnol. 2013, 30, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Shi, Z. Molecular research progress of rice panicle development. Plant Physiol. J. 2013, 49, 111–121. [Google Scholar]
- Zhou, Y.; Tao, Y.; Yuan, Y.; Zhang, Y.; Miao, J.; Zhang, R.; Yi, C.; Gong, Z.; Yang, Z.; Liang, G. Characterisation of a novel quantitative trait locus, GN4-1, for grain number and yield in rice (Oryza sativa L.). Theor. Appl. Genet. 2018, 131, 637–648. [Google Scholar] [CrossRef]
- Chen, H.; Tang, Y.; Liu, J.; Tan, L.; Jiang, J.; Wang, M.; Zhu, Z.; Sun, X.; Sun, C. Emergence of a novel chimeric gene underlying grain number in rice. Genetics 2017, 205, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Bian, J.; Shi, S.; Yu, J.; Khanzada, H.; Wassan, G.M.; Zhu, C.; Luo, X.; Tong, S.; Yang, X.; et al. Genetic analysis for the grain number heterosis of a super-hybrid rice WFYT025 combination using RNA-Seq. Rice 2018, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Tao, H.; Min, C.; Yang, Y. Research progress on rice panicle development. Mol. Plant Breed. 2015, 13, 2109–2117. [Google Scholar] [CrossRef]
- Li, S.; Zhao, B.; Yuan, D.; Duan, M.; Qian, Q.; Tang, L.; Wang, B.; Liu, X.; Zhang, J.; Wang, J.; et al. Rice zinc finger protein DST enhances grain production through controlling Gn1a/OsCKX2 expression. Proc. Natl. Acad. Sci. USA 2013, 110, 3167–3172. [Google Scholar] [CrossRef] [Green Version]
- Ikeda-Kawakatsu, K.; Yasuno, N.; Oikawa, T.; Iida, S.; Nagato, Y.; Maekawa, M.; Kyozuka, J. Expression level of ABERRANT PANICLE ORGANIZATION1 determines rice inflorescence form through control of cell proliferation in the meristem. Plant Physiol. 2009, 150, 736–747. [Google Scholar] [CrossRef] [Green Version]
- Deveshwar, P.; Prusty, A.; Sharma, S.; Tyagi, A.K. Phytohormone-Mediated molecular mechanisms involving multiple genes and QTL govern grain number in rice. Front. Genet. 2020, 11, 586462. [Google Scholar] [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.-K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. USA 2018, 115, 6058–6063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, A.; Ohmori, Y.; Kitano, H.; Taguchi-Shiobara, F.; Hirano, H.Y. Aberrant spikelet and panicle1, encoding a TOPLESS-related transcriptional co-repressor, is involved in the regulation of meristem fate in rice. Plant J. 2012, 70, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, J.; Wu, L.; Shao, Y.; Wu, Y.; Mao, C. Functional divergence of PIN1 paralogous genes in rice. Plant Cell Physiol. 2019, 60, 2720–2732. [Google Scholar] [CrossRef] [PubMed]
- Malik, N.; Ranjan, R.; Parida, S.K.; Agarwal, P.; Tyagi, A.K. Mediator subunit OsMED14_1 plays an important role in rice development. Plant J. 2020, 101, 1411–1429. [Google Scholar] [CrossRef]
- Gao, S.; Chu, C. Gibberellin metabolism and signaling: Targets for improving agronomic performance of crops. Plant Cell Physiol. 2020, 61, 1902–1911. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Xiao, G.; Zhai, M.; Pan, X.; Huang, R.; Zhang, H. CYP71D8L is a key regulator involved in growth and stress responses by mediating gibberellin homeostasis in rice. Science 2020, 71, 1160–1170. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Kurakawa, T.; Ueda, N.; Maekawa, M.; Kobayashi, K.; Kojima, M.; Nagato, Y.; Sakakibara, H.; Kyozuka, J. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature 2007, 445, 652–655. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Q.; Wu, J.; Zhang, L.; Jiao, X.; Zhang, S.; Zhang, Z.; Sun, D.; Lu, T.; Sun, Y. Two rice authentic histidine phosphotransfer proteins, OsAHP1 and OsAHP2, mediate cytokinin signaling and stress responses in rice. Plant Physiol. 2014, 165, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Hirose, N.; Makita, N.; Kojima, M.; Kamada-Nobusada, T.; Sakakibara, H. Overexpression of a type-A response regulator alters rice morphology and cytokinin metabolism. Plant Cell Physiol. 2007, 48, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Wuriyanghan, H.; Zhang, B.; Cao, W.-H.; Ma, B.; Lei, G.; Liu, Y.-F.; Wei, W.; Wu, H.-J.; Chen, L.-J.; Chen, H.W.; et al. The ethylene receptor ETR2 delays floral transition and affects starch accumulation in rice. Plant Cell 2009, 21, 1473–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sentoku, N.; Sato, Y.; Kurata, N.; Ito, Y.; Kitano, H.; Matsuoka, M. Regional expression of the Rice KN1-Type homeobox gene family during embryo, shoot, and flower development. Plant Cell 1999, 11, 1651–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, T.; Sakakibara, H.; Kojima, M.; Yamamoto, Y.; Nagasaki, H.; Inukai, Y.; Sato, Y.; Matsuoka, M. Ectopic expression of KNOTTED1-like homeobox protein induces expression of cytokinin biosynthesis genes in rice. Plant Physiol. 2006, 142, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Qian, Q.; Fu, Z.; Wang, Y.; Xiong, G.; Zeng, D.; Wang, X.; Liu, X.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qian, Q.; Fu, Z.; Zeng, D.; Meng, X.; Kyozuka, J.; Maekawa, M.; Zhu, X.; Zhang, J.; Li, J.; et al. Short panicle1 encodes a putative PTR family transporter and determines rice panicle size. Plant J. 2009, 58, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Kyozuka, J. Two-step regulation of LAX PANICLE1 protein accumulation in axillary meristem formation in rice. Plant Cell 2009, 21, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Ito, M.; Nagasawa, N.; Kyozuka, J.; Nagato, Y. Rice ABERRANT PANICLE ORGANIZATION 1, encoding an F-box protein, regulates meristem fate. Plant J. 2007, 51, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Ookawa, T.; Hobo, T.; Yano, M.; Murata, K.; Ando, T.; Miura, H.; Asano, K.; Ochiai, Y.; Ikeda, M.; Nishitani, R.; et al. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 2010, 1, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, M.; Shimamoto, K.; Kyozuka, J. Overexpression of RCN1 and RCN2, rice TERMINAL FLOWER 1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice. Plant J. 2002, 29, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Sasao, M.; Yasuno, N.; Takagi, K.; Daimon, Y.; Chen, R.; Yamazaki, R.; Tokunaga, H.; Kitaguchi, Y.; Sato, Y.; et al. TAWAWA1, a regulator of rice inflorescence architecture, functions through the suppression of meristem phase transition. Proc. Natl. Acad. Sci. USA 2013, 110, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, M.; Maekawa, M.; Shimamoto, K.; Kyozuka, J. The LAX1 and FRIZZY PANICLE 2 genes determine the inflorescence architecture of rice by controlling rachis-branch and spikelet development. Dev. Biol. 2001, 231, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, K.; Maekawa, M.; Ujiie, S.; Satake, Y.; Furutani, I.; Okamoto, H.; Shimamoto, K.; Kyozuka, J. LAX and SPA: Major regulators of shoot branching in rice. Proc. Natl. Acad. Sci. USA 2003, 100, 11765–11770. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, M.; Chujo, A.; Nagato, Y.; Shimamoto, K.; Kyozuka, J. FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development 2003, 130, 3841–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Maekawa, M.; Miyao, A.; Hirochika, H.; Kyozuka, J. PANICLE PHYTOMER2 (PAP2), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice. Plant Cell Physiol. 2010, 51, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Huang, Y.; Mao, D.; Wen, M.; Zhang, L.; Xing, Y. Regulatory role of FZP in the determination of panicle branching and spikelet formation in rice. Sci. Rep. 2016, 6, 19022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, K.; Ikeda, M.; Matsubara, A.; Song, X.-J.; Ito, M.; Asano, K.; Matsuoka, M.; Kitano, H.; Ashikari, M. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat. Genet. 2010, 42, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Shao, G.; Lu, Z.; Xiong, J.; Wang, B.; Jing, Y.; Meng, X.; Liu, G.; Ma, H.; Liang, Y.; Chen, F.; et al. Tiller bud formation regulators MOC1 and MOC3 cooperatively promote tiller bud outgrowth by activating FON1 expression in rice. Mol. Plant 2019, 12, 1090–1102. [Google Scholar] [CrossRef] [Green Version]
- Postma-Haarsma, A.D.; Verwoert, I.I.G.S.; Stronk, O.P.; Koster, J.; Lamers, G.E.M.; Hoge, J.H.C.; Meijer, A.H. Characterization of the KNOX class homeobox genes Oskn2 and Oskn3 identified in a collection of cDNA libraries covering the early stages of rice embryogenesis. Plant Mol. Biol. 1999, 39, 257–271. [Google Scholar] [CrossRef]
- Sato, Y.; Hong, S.K.; Tagiri, A.; Kitano, H.; Yamamoto, N.; Nagato, Y.; Matsuoka, M. A rice homeobox gene, OSH1, is expressed before organ differentiation in a specific region during early embryogenesis. Proc. Natl. Acad. Sci. USA 1996, 93, 8117–8122. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Ito, Y.; Sato, Y.; Kurata, N. Positive autoregulation of a KNOX gene is essential for shoot apical meristem maintenance in rice. Plant Cell 2011, 23, 4368–4381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Qian, Q.; Liu, Z.; Sun, H.; He, S.; Luo, D.; Xia, G.; Chu, C.; Li, J.; Fu, X. Natural variation at the DEP1 locus enhances grain yield in rice. Nat. Genet. 2009, 41, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, S.; Wang, Y.; Zhang, X.; Lv, B.; Luo, L.; Xi, D.; Shen, J.; Ma, H.; Ming, F. OsNAC2 encoding a NAC transcription factor that affects plant height through mediating the gibberellic acid pathway in rice. Plant J. 2015, 82, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xie, K.; Xiong, L. Conserved miR164-targeted NAC genes negatively regulate drought resistance in rice. J. Exp. Bot. 2014, 65, 2119–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, C.; Ding, W.; Wu, Y.; Yu, J.; He, X.; Shou, H.; Wu, P. Overexpression of a NAC-domain protein promotes shoot branching in rice. New Phytol. 2007, 176, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Chen, W.; Dong, J.; Li, J.; Yang, F.; Wu, Z.; Zhou, H.; Wang, W.; Zhuang, C. Overexpression of miR164b-resistant OsNAC2 improves plant architecture and grain yield in rice. J. Exp. Bot. 2018, 69, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.; Singh, A.; Jain, N.; Anand, S.; Gacche, R.; Singh, A.; Gaikwad, K.; Sharma, T.; Mohapatra, T.; Singh, N. Identification of candidate genes for grain number in rice (Oryza sativa L.). Funct. Integr. Genom. 2010, 10, 339–347. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Mi, X.-F.; Shan, J.-X.; Li, X.-M.; Xu, J.-L.; Lin, H.-X. The QTL GNP1 encodes GA20ox1, which increases grain number and yield by increasing cytokinin activity in rice panicle meristems. PLoS Genet. 2016, 12, e1006386. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Yao, G.; Zhang, H.; Dou, H.; Shi, H.; Sun, X.; Li, Z. Fine mapping and cloning of the grain number per-panicle gene (Gnp4) on chromosome 4 in rice (Oryza sativa L.). Agric. Sci. China 2011, 10, 1825–1833. [Google Scholar] [CrossRef]
- Tabuchi, H.; Zhang, Y.; Hattori, S.; Omae, M.; Shimizu-Sato, S.; Oikawa, T.; Qian, Q.; Nishimura, M.; Kitano, H.; Xie, H.; et al. LAX PANICLE2 of rice encodes a novel nuclear protein and regulates the formation of axillary meristems. Plant Cell 2011, 23, 3276–3287. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Tan, L.; Zhu, Z.; Xiao, L.; Xie, D.; Sun, C. PAY1 improves plant architecture and enhances grain yield in rice. Plant J. 2015, 83, 528–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Tang, D.; Wang, K.; Wu, X.; Lu, L.; Yu, H.; Gu, M.; Yan, C.; Cheng, Z. Mutations in the F-box gene LARGER PANICLE improve the panicle architecture and enhance the grain yield in rice. Plant Biotechnol. J. 2011, 9, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Murchie, E.H.; González-Carranza, Z.H.; Pyke, K.A.; Roberts, J.A. Decreased photosynthesis in the erect panicle 3 (ep3) mutant of rice is associated with reduced stomatal conductance and attenuated guard cell development. J. Exp. Bot. 2015, 66, 1543–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, R.; Jiang, W.; Ham, T.-H.; Choi, M.-S.; Qiao, Y.; Chu, S.-H.; Park, J.-H.; Woo, M.-O.; Jin, Z.; An, G.; et al. Map-based cloning of the ERECT PANICLE 3 gene in rice. Theor. Appl. Genet. 2009, 119, 1497–1506. [Google Scholar] [CrossRef]
- Qiao, Y.; Piao, R.; Shi, J.; Lee, S.-I.; Jiang, W.; Kim, B.-K.; Lee, J.; Han, L.; Ma, W.; Koh, H.-J. Fine mapping and candidate gene analysis of dense and erect panicle 3, DEP3, which confers high grain yield in rice (Oryza sativa L.). Theor. Appl. Genet. 2011, 122, 1439–1449. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, K.; Ai, J.; Deng, X.; Hong, Y.; Wang, X. Patatin-related phospholipase A, pPLAIIIα, modulates the longitudinal growth of vegetative tissues and seeds in rice. J. Exp. Bot. 2015, 66, 6945–6955. [Google Scholar] [CrossRef] [Green Version]
- Zahn, L.M.; Kong, H.; Leebens-Mack, J.H.; Kim, S.; Soltis, P.S.; Landherr, L.L.; Soltis, D.E.; de Pamphilis, C.W.; Ma, H. The evolution of the SEPALLATA subfamily of MADS-box genes: A preangiosperm origin with multiple duplications throughout angiosperm history. Genetics 2005, 169, 2209–2223. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Liang, W.; Yin, C.; Ji, S.; Wang, H.; Su, X.; Guo, C.; Kong, H.; Xue, H.; Zhang, D. The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol. 2010, 153, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yu, H.; Liu, J.; Wang, W.; Sun, J.; Gao, Q.; Zhang, Y.; Ma, D.; Wang, J.; Xu, Z.; et al. Loss of function of OsMADS34 leads to large sterile lemma and low grain yield in rice (Oryza sativa L.). Mol. Breed. 2016, 36, 147. [Google Scholar] [CrossRef]
- Liu, C.; Teo, Z.W.N.; Bi, Y.; Song, S.; Xi, W.; Yang, X.; Yin, Z.; Yu, H. A conserved genetic pathway determines inflorescence architecture in Arabidopsis and rice. Dev. Cell 2013, 24, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, H.; Zhou, T.; Gu, B.; Huang, X.; Shangguan, Y.; Zhu, J.; Li, Y.; Zhao, Y.; Wang, Y.; et al. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice. Plant Cell 2013, 25, 3360–3376. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Li, X.; Liu, F.; Sun, X.; Li, C.; Zhu, Z.; Fu, Y.; Cai, H.; Wang, X.; Xie, D.; et al. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364. [Google Scholar] [CrossRef]
- Jin, J.; Huang, W.; Gao, J.-P.; Yang, J.; Shi, M.; Zhu, M.-Z.; Luo, D.; Lin, H.-X. Genetic control of rice plant architecture under domestication. Nat. Genet. 2008, 40, 1365–1369. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J. Rice, rising. Nat. Genet. 2008, 40, 1273–1275. [Google Scholar] [CrossRef]
- Guo, T.; Lu, Z.-Q.; Shan, J.-X.; Ye, W.-W.; Dong, N.-Q.; Lin, H.-X. ERECTA1 acts upstream of the OsMKKK10-OsMKK4-OsMPK6 cascade to control spikelet number by regulating cytokinin metabolism in rice. Plant Cell 2020, 32, 2763–2779. [Google Scholar] [CrossRef]
- Zhu, Q.-H.; Hoque, M.S.; Dennis, E.S.; Upadhyaya, N.M. Ds tagging of BRANCHED FLORETLESS 1 (BFL1) that mediates the transition from spikelet to floret meristem in rice (Oryza sativa L). BMC Plant Biol. 2003, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Putterill, J.; Robson, F.; Lee, K.; Simon, R.; Coupland, G. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell 1995, 80, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Yano, M.; Katayose, Y.; Ashikari, M.; Yamanouchi, U.; Monna, L.; Fuse, T.; Baba, T.; Yamamoto, K.; Umehara, Y.; Nagamura, Y.; et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 2000, 12, 2473–2484. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Xu, J.; Guo, H.; Jiang, L.; Chen, S.; Yu, C.; Zhou, Z.; Hu, P.; Zhai, H.; Wan, J. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010, 153, 1747–1758. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.-H.; Wang, P.; Chen, H.-X.; Zhou, H.-J.; Li, Q.-P.; Wang, C.-R.; Ding, Z.-H.; Zhang, Y.-S.; Yu, S.-B.; Xing, Y.-Z.; et al. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol. Plant 2011, 4, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Ding, Y.; Tan, L.; Fu, Y.; Liu, F.; Zhu, Z.; Sun, X.; Sun, X.; Gu, P.; Cai, H.; et al. LHD1, an allele of DTH8/Ghd8, controls late heading date in common wild rice (Oryza rufipogon). J. Integr. Plant Biol. 2012, 54, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Thirumurugan, T.; Ito, Y.; Kubo, T.; Serizawa, A.; Kurata, N. Identification, characterization and interaction of HAP family genes in rice. Mol. Genet. Genom. 2008, 279, 279–289. [Google Scholar] [CrossRef]
- Murakami, M.; Matsushika, A.; Ashikari, M.; Yamashino, T.; Mizuno, T. Circadian-associated rice pseudo response regulators (OsPRRs): Insight into the control of flowering time. Biosci. Biotech. Biochem. 2005, 69, 410–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, B.-H.; Yoo, S.-C.; Park, J.-W.; Kwon, C.-T.; Lee, B.-D.; An, G.; Zhang, Z.; Li, J.; Li, Z.; Paek, N.-C. Natural variation in OsPRR37 regulates heading date and contributes to rice cultivation at a wide range of latitudes. Mol. Plant 2013, 6, 1877–1888. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Jin, M.; Zheng, X.-M.; Chen, J.; Yuan, D.; Xin, Y.; Wang, M.; Huang, D.; Zhang, Z.; Zhou, K.; et al. Days to heading 7, a major quantitative locus determining photoperiod sensitivity and regional adaptation in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 16337–16342. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Liu, H.; Zhou, X.; Li, Q.; Zhang, J.; Lu, L.; Liu, T.; Liu, H.; Zhang, C.; Zhang, Z.; et al. Natural variation in Ghd7.1 plays an important role in grain yield and adaptation in rice. Cell Res. 2013, 23, 969–971. [Google Scholar] [CrossRef]
- Jin, J.; Hua, L.; Zhu, Z.; Tan, L.; Zhao, X.; Zhang, W.; Liu, F.; Fu, Y.; Cai, H.; Sun, X.; et al. GAD1 encodes a secreted peptide that regulates grain number, grain length, and awn development in rice domestication. Plant Cell 2016, 28, 2453–2463. [Google Scholar] [CrossRef] [Green Version]
- Huo, X.; Wu, S.; Zhu, Z.; Liu, F.; Fu, Y.; Cai, H.; Sun, X.; Gu, P.; Xie, D.; Tan, L.; et al. NOG1 increases grain production in rice. Nat. Commun. 2017, 8, 1497. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Gene | RGAP Locus ID | Protein Products | PRB | SRB | Phase Transition | Spikelet Specialisation | Lateral Spikelet | Terminal Spikelet | GNPP | References |
|---|---|---|---|---|---|---|---|---|---|---|
| LAX1 | LOC_Os01g61480 | bHLH transcription factor | + | + | + | + | + | + | [26,32] | |
| IPA1/WFP/OsSPL14 | LOC_Os08g39890 | SOUAMOSA PROMOTER BINDING PROTEIN-LIKE protein | + | + | + | [36,37] | ||||
| MOC1 | LOC_Os06g40780 | GRAS-family nuclear protein | + | + | + | [24,38] | ||||
| OSH1 | LOC_Os03g51690 | A protein product of 364 amino acids | + | + | + | [22,39,40,41] | ||||
| SPA | Not reported | Small panicle | + | + | + | + | [32] | |||
| SP1 | LOC_Os11g12740 | A putative transporter of the peptide transporter (PTR) family | + | + | + | [25] | ||||
| DEP1 | LOC_Os09g26999 | G protein gamma subunit | + | + | + | [42] | ||||
| OsNAC2/OMTN2/Ostil1 | LOC_Os04g38720 | NAC transcription factor | + | + | + | [43,44,45,46] | ||||
| GN4-1 | Not reported | Grain number4-1 | + | + | + | [4,47] | ||||
| GNP1 | LOC_Os03g63970 | GA20-oxidase 1 | + | + | [48] | |||||
| Gnp4/LAX2 | LOC_Os04g32510 | A nuclear protein with a plant-specific conserved domain | + | + | [49,50] | |||||
| PAY1 | LOC_Os08g31470 | Peptidase | + | + | [51] | |||||
| LP/EP3 | LOC_Os02g15950 | F-box protein | − | − | − | [52,53,54] | ||||
| DEP3/OspPLAIIIδ | LOC_Os06g46350 | Patatin-related phospholipase A | − | − | − | [55,56] | ||||
| PAP2/OsMADS34 | LOC_Os03g54170 | SEP-like MADS box transcription factor | − | − | + | − | [34,57,58,59,60] | |||
| An-1 | LOC_Os04g28280 | bHLH protein | − | − | − | [61] | ||||
| PROG1 | LOC_Os07g05900 | A 161-amino-acid Cys2-His2 zinc-finger protein | − | − | − | [62,63,64] | ||||
| DST | LOC_Os03g57240 | Zinc-finger transcription factor | − | − | − | [8,65] | ||||
| GN2 | Not reported | Gain number2 | − | − | [5] | |||||
| APO1/SCM2 | LOC_Os06g45460 | F-box protein of 429 amino acids | + | + | − | + | [9,27,28] | |||
| RCN1 | LOC_Os11g05470 | ATP-binding cassette transporter | + | − | + | [29] | ||||
| RCN2 | LOC_Os02g32950 | ATP-binding cassette transporter | + | − | + | [29] | ||||
| TAW1 | LOC_Os10g33780 | Nuclear protein | + | + | − | − | + | [30] | ||
| FZP/BFL1 | LOC_Os07g47330 | ERF transcription factor | − | + | + | + | [33,35,66] | |||
| FZP2 | Not reported | Frizzy panicle 2 | − | + | + | + | [31] | |||
| GN1a/OsCKX2 | LOC_Os01g10110 | Cytokinin oxidase/dehydrogenase | − | − | − | [17] | ||||
| LOG | LOC_Os01g40630 | Cytokinin riboside 50-monophosphate phosphoribohydro-lase | + | + | + | [18] | ||||
| PYL1 PYL4 PYL6 | LOC_Os01g61210 LOC_Os03g18600 LOC_Os05g39580 | ABA receptor protein | − | − | − | [11] | ||||
| Ghd7 | LOC_Os07g15770 | CCT(CO, CO-LIKE and TIMING OF CAB1) | + | + | + | [67,68,69] | ||||
| Ghd8/DTH8/OsHAP3H/LHD1 | LOC_Os08g07740 | HAP3 subunit of the HAP (heterotrimeric haem activator) complex | + | + | + | [70,71,72,73] | ||||
| DTH7/Ghd7.1/OsPRR37 | LOC_Os07g49460 | A pseudo-response regulator protein | + | + | + | [74,75,76,77] | ||||
| GAD1 | LOC_Os08g37890 | A cysteine-rich secretory peptide | + | [78] | ||||||
| NOG1 | LOC_Os01g54860 | Enoyl-CoA hydratase/isomerase | + | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, C.; Zhu, Y.; Li, X.; Lin, Y. Molecular and Genetic Aspects of Grain Number Determination in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2021, 22, 728. https://doi.org/10.3390/ijms22020728
Yin C, Zhu Y, Li X, Lin Y. Molecular and Genetic Aspects of Grain Number Determination in Rice (Oryza sativa L.). International Journal of Molecular Sciences. 2021; 22(2):728. https://doi.org/10.3390/ijms22020728
Chicago/Turabian StyleYin, Changxi, Yanchun Zhu, Xuefei Li, and Yongjun Lin. 2021. "Molecular and Genetic Aspects of Grain Number Determination in Rice (Oryza sativa L.)" International Journal of Molecular Sciences 22, no. 2: 728. https://doi.org/10.3390/ijms22020728
APA StyleYin, C., Zhu, Y., Li, X., & Lin, Y. (2021). Molecular and Genetic Aspects of Grain Number Determination in Rice (Oryza sativa L.). International Journal of Molecular Sciences, 22(2), 728. https://doi.org/10.3390/ijms22020728