Norepinephrine Regulation of Ventromedial Hypothalamic Nucleus Astrocyte Glycogen Metabolism


{kind=link}
{kind=link}
Abstract
:1. Overview
2. Neural Regulation of Glucostasis
3. Hindbrain-to-Hypothalamic Signaling in Gluco-Regulation
4. Astrocyte Involvement in Neuro-Metabolic Stability
5. Brain Glycogen Metabolism
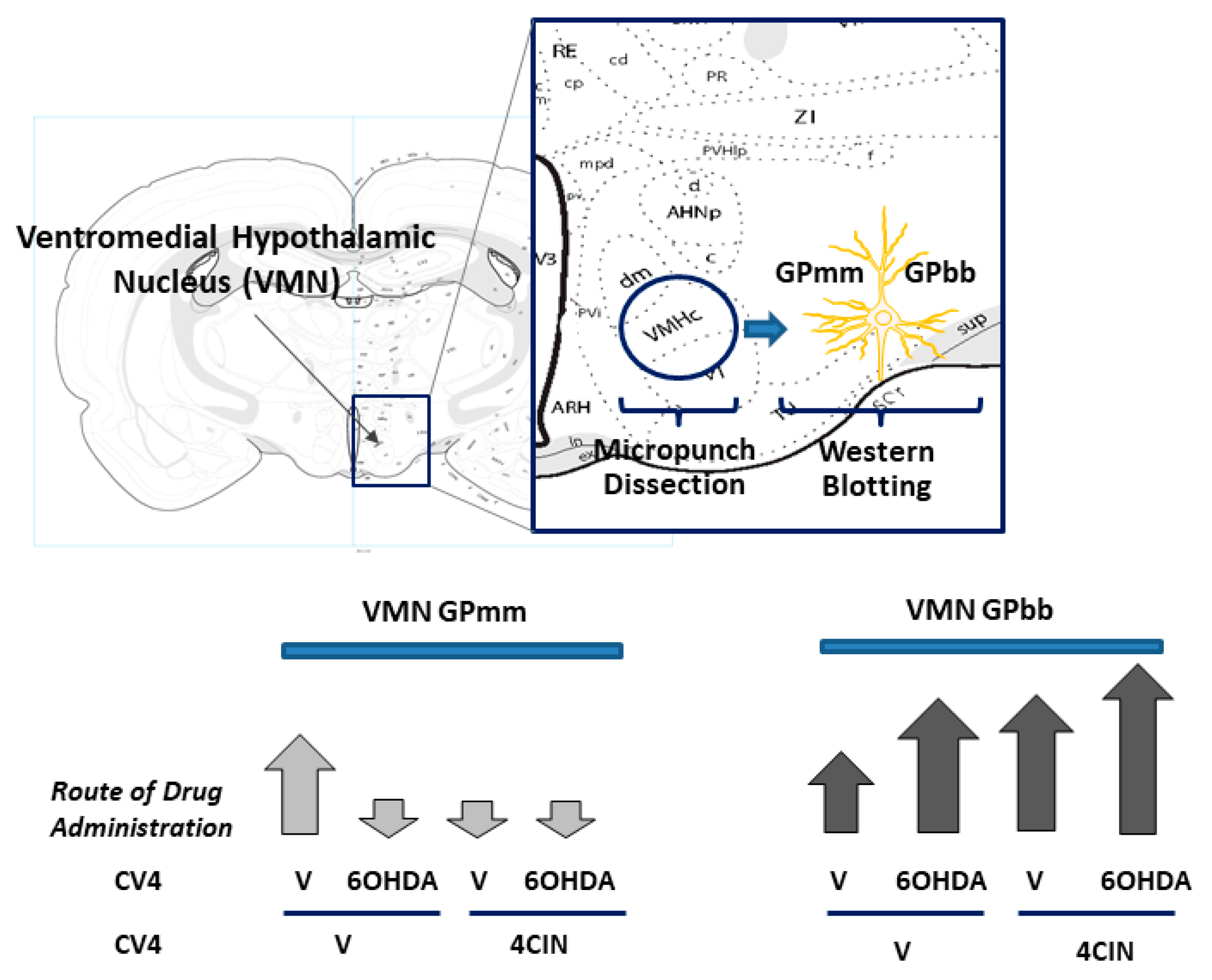
6. Effects of Pharmacological Inhibition of VMN GP Activity on Metabolic Transmitter Signaling
7. Hindbrain Noradrenergic Regulation of VMN Glycogen Metabolism
8. NE Signaling Appraises the VMN of Hindbrain Glycogen Metabolic Status
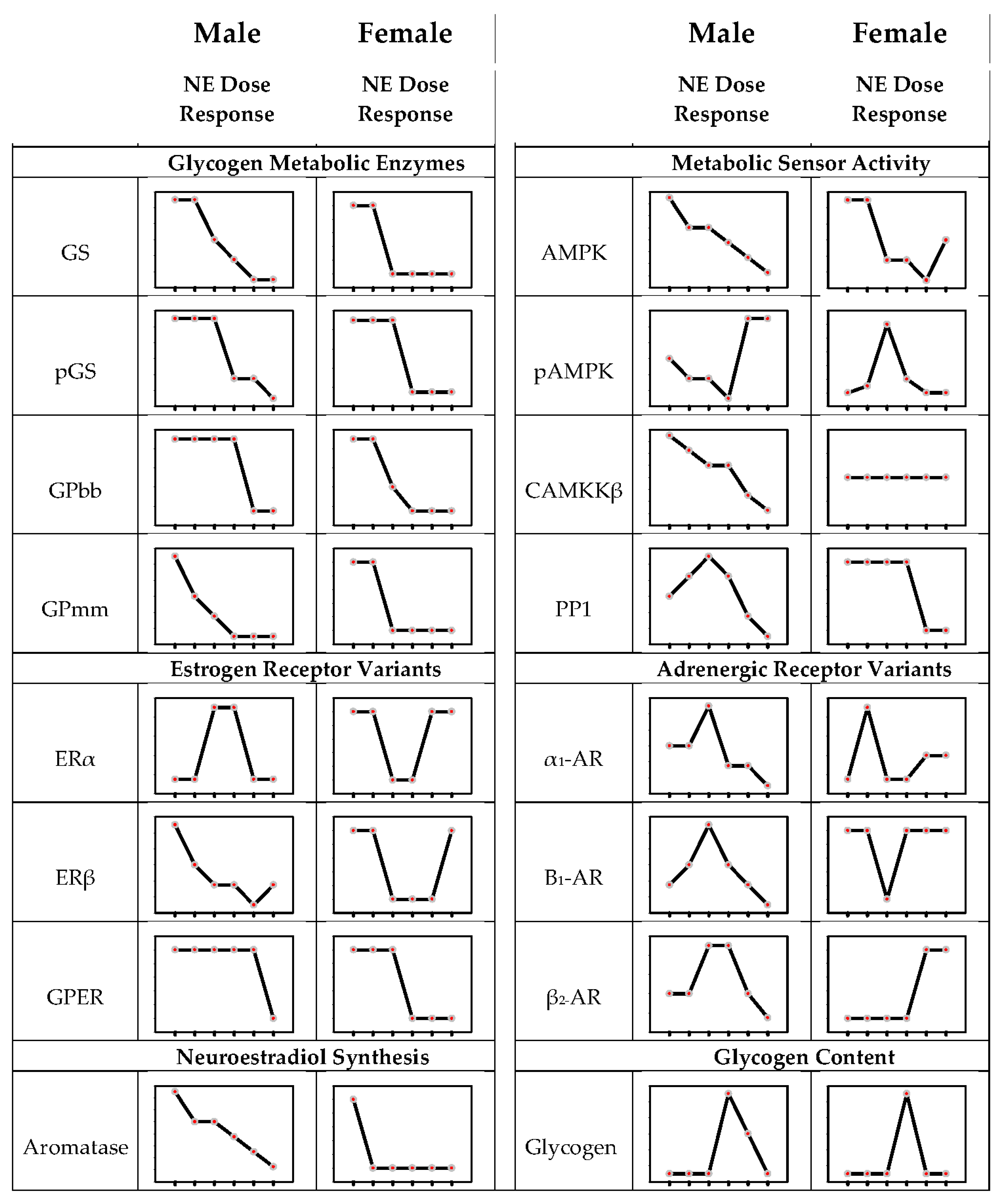
9. Concentration-Dependent Noradrenergic Regulation of Hypothalamic Astrocyte Glycogen Metabolic Enzyme and Adrenergic Receptor Expression in Male Versus Female Primary Astrocyte Cultures
10. Conclusions and Directions for Future Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| α1-AR | Alpha1-adrenergic receptor |
| α2-AR | Alpha2-adrenergic receptor |
| β1-AR | Beta1-adrenergic receptor |
| β2-AR | Beta2-adrenergic receptor |
| CaMKKβ | Calcium/calmodulin-dependent protein kinase kinase-β |
| ERα | Estrogen receptor-alpha |
| ERβ | Estrogen receptor-beta |
| GABA | γ-aminobutyric acid |
| GAD | Glutamate decarboxylase65/67 |
| GPbb | Glycogen phosphorylase brain type |
| GPER | G-protein-coupled estrogen receptor-1 |
| GPmm | Glycogen phosphorylase muscle type |
| GS | Glycogen synthase |
| NE | Norepinephrine |
| NO | Nitric oxide |
| nNOS | Neuronal nitric oxide synthase |
| PP1 | Protein phosphatase-1 |
| VMN | Ventromedial hypothalamic nucleus |
References
- Stobart, J.L.; Anderson, C.M. Multifunctional role of astrocytes as gatekeepers of neuronal energy supply. Front. Cell. Neurosci. 2013, 7, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bak, L.K.; Walls, A.B.; Schousboe, A.; Waagepetersen, H.S. Astrocytic glycogen metabolism in the healthy and diseased brain. J. Biol. Chem. 2018, 293, 7108–7116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dienel, G.A.; Carlson, G.M. Major Advances in brain glycogen research: Understanding of the roles of glycogen have evolved from emergency fuel reserve to dynamic, regulated participant in diverse brain functions. Adv. Neurobiol. 2019, 23, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cryer, P.E. Hypoglycemia in type 1 diabetes mellitus. Endocrinol. Metab. Clin. N. Am. 2010, 39, 641–654. [Google Scholar] [CrossRef] [Green Version]
- Watts, A.G.; Donovan, C.M. Sweet talk in the brain: Glucosensing, neural networks, and hypoglycemic counterregulation. Front. Neuroendocrinol. 2010, 31, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Oomura, Y.; Ono, T.; Ooyama, H.; Wayner, M.J. Glucose and osmosensitive neurones of the rat hypothalamus. Nature 1969, 222, 282–284. [Google Scholar] [CrossRef]
- Adachi, A.; Kobashi, M.; Funahashi, M. Glucose-responsive neurons in the brainstem. Obes. Res. 1995, 3, 735S–740S. [Google Scholar] [CrossRef]
- Mizuno, Y.; Oomura, Y. Glucose responding neurons in the nucleus tractus solitarius of the rat: In vitro study. Brain Res. 1984, 307, 109–116. [Google Scholar] [CrossRef]
- Ashford, M.L.J.; Boden, P.R.; Treherne, J.M. Glucose-induced excitation of hypothalamic neurones is mediated by ATP-sensitive K+ channels. Pflügers Arch. 1990, 415, 479–483. [Google Scholar] [CrossRef]
- Yettefti, K.; Orsini, J.; El Ouazzani, T.; Himmi, T.; Boyer, A.; Perrin, J. Sensitivity of nucleus tractus solitarius neurons to induced moderate hyperglycemia, with special reference to catecholaminergic regions. J. Auton. Nerv. Syst. 1995, 51, 191–197. [Google Scholar] [CrossRef]
- Silver, I.A.; Erecinska, M. Glucose-induced intracellular ion changes in sugar-sensitive hypothalamic neurons. J. Neurophysiol. 1998, 79, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Yettefti, K.; Orsini, J.-C.; Perrin, J. Characteristics of glycemia-sensitive neurons in the nucleus tractus solitarii: Possible involvement in nutritional regulation. Physiol. Behav. 1997, 61, 93–100. [Google Scholar] [CrossRef]
- Himmi, T.; Perrin, J.; Dallaporta, M.; Orsini, J.-C. Effects of lactate on glucose-sensing neurons in the solitary tract nucleus. Physiol. Behav. 2001, 74, 391–397. [Google Scholar] [CrossRef]
- Balfour, R.H.; Hansen, A.M.K.; Trapp, S. Neuronal responses to transient hypoglycaemia in the dorsal vagal complex of the rat brainstem. J. Physiol. 2005, 570, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Pénicaud, L.; Leloup, C.; Lorsignol, A.; Alquier, T.; Guillod, E. Brain glucose sensing mechanism and glucose homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 539–543. [Google Scholar] [CrossRef]
- Ritter, S.; Dinh, T.T.; Zhang, Y. Localization of hindbrain glucoreceptive sites controlling food intake and blood glucose. Brain Res. 2000, 856, 37–47. [Google Scholar] [CrossRef]
- Patil, G.D.; Briski, K.P. Lactate is a critical “sensed” variable in caudal hindbrain monitoring of CNS metabolic stasis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1777–R1786. [Google Scholar] [CrossRef] [Green Version]
- Briski, K.P.; Cherian, A.; Genabai, N.; Vavaiya, K. In situ coexpression of glucose and monocarboxylate transporter mRNAs in metabolic-sensitive caudal dorsal vagal complex catecholaminergic neurons: Transcriptional reactivity to insulin-induced hypoglycemia and caudal hindbrain glucose or lactate repletion during insulin-induced hypoglycemia. Neuroscience 2009, 164, 1152–1160. [Google Scholar] [CrossRef]
- Briski, K.P.; Marshall, E.S. Caudal brainstem Fos expression is restricted to periventricular catecholamine neuron-containing loci following intraventricular administration of 2-deoxy-D-glucose. Exp. Brain Res. 2000, 133, 547–551. [Google Scholar] [CrossRef]
- Li, A.-J.; Wang, Q.; Ritter, S. Differential responsiveness of dopamine-β-hydroxylase gene expression to glucoprivation in different catecholamine cell groups. Endocrinology 2006, 147, 3428–3434. [Google Scholar] [CrossRef] [Green Version]
- Briski, K.P.; Patil, G.D. Induction of Fos immunoreactivity labeling in rat forebrain metabolic loci by caudal fourth ventricular infusion of the monocarboxylate transporter inhibitor, alpha-cyano-4-hydroxycinnamic acid. Neuroendocrinology 2005, 82, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, P.; Tamrakar, P.; Ibrahim, B.; Briski, K.P. Hindbrain medulla catecholamine cell group involvement in lactate-sensitive hypoglycemia-associated patterns of hypothalamic norepinephrine and epinephrine activity. Neuroscience 2014, 278, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Nortley, R.; Attwell, D. Control of brain energy supply by astrocytes. Curr. Opin. Neurobiol. 2017, 47, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Laming, P.; Kimelberg, H.; Robinson, S.; Salm, A.; Hawrylak, N.; Müller, C.; Roots, B.; Ng, K. Neuronal–glial interactions and behaviour. Neurosci. Biobehav. Rev. 2000, 24, 295–340. [Google Scholar] [CrossRef]
- Bröer, S.; Rahman, B.; Pellegri, G.; Pellerin, L.; Martin, J.-L.; Verleysdonk, S.; Hamprecht, B.; Magistretti, P.J. Comparison of lactate transport in astroglial cells and monocarboxylate transporter 1 (MCT 1) expressing xenopus laevis oocytes. Expression of two different monocarboxylate transporters in astroglial cells and neurons. J. Biol. Chem. 1997, 272, 30096–30102. [Google Scholar] [CrossRef] [Green Version]
- Pellerin, L.; Magistretti, P.J. Sweet sixteen for ANLS. Br. J. Pharmacol. 2011, 32, 1152–1166. [Google Scholar] [CrossRef]
- Cambray-Deakin, M.; Pearce, B.; Morrow, C.; Murphy, S. Effects of extracellular potassium on glycogen stores of astrocytes in vitro. J. Neurochem. 1988, 51, 1846–1851. [Google Scholar] [CrossRef]
- Dringen, R.; Gebhardt, R.; Hamprecht, B. Glycogen in astrocytes: Possible function as lactate supply for neighboring cells. Brain Res. 1993, 623, 208–214. [Google Scholar] [CrossRef]
- Swanson, R.A.; Choi, D.W. Glial glycogen stores affect neuronal survival during glucose deprivation in vitro. J. Cereb. Blood Flow Metab. 1993, 13, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Magistretti, P.J. Regulation of glycogenolysis by neurotransmitters in the central nervous system. Diabete Metab. 1988, 14, 237–246. [Google Scholar]
- Magistretti, P.J. Vasoactive intestinal peptide and noradrenaline regulate energy metabolism in astrocytes: A physiological function in the control of local homeostasis within the CNS. Prog. Brain Res. 1994, 100, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J.; Sorg, O.; Yu, N.; Martin, J.-L.; Pellerin, L. Neurotransmitters regulate energy metabolism in astrocytes: Implications for the metabolic trafficking between neural cells. Dev. Neurosci. 1993, 15, 306–312. [Google Scholar] [CrossRef] [PubMed]
- DiNuzzo, M.; Giove, F.; Maraviglia, B.; Mangia, S. Monoaminergic control of cellular glucose utilization by glycogenolysis in neocortex and hippocampus. Neurochem. Res. 2015, 40, 2493–2504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walls, A.; Heimbürger, C.; Bouman, S.D.; Schousboe, A.; Waagepetersen, H.S. Robust glycogen shunt activity in astrocytes: Effects of glutamatergic and adrenergic agents. Neuroscience 2009, 158, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Sickmann, H.M.; Walls, A.B.; Bak, L.K.; Waagepetersen, H.S. Functional importance of the astrocytic glycogen-shunt and glycolysis for maintenance of an intact intra/extracellular glutamate gradient. Neurotox. Res. 2010, 18, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Obel, L.F.; Müller, M.S.; Walls, A.B.; Sickmann, H.M.; Bak, L.K.; Waagepetersen, H.S.; Schousboe, A. Brain glycogen—New perspectives on its metabolic function and regulation at the subcellular level. Front. Neuroenergetics 2012, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Gruetter, R. Glycogen: The forgotten cerebral energy store. J. Neurosci. Res. 2003, 74, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.M. Brain glycogen re-awakened. J. Neurochem. 2004, 89, 537–552. [Google Scholar] [CrossRef]
- Bélanger, M.; Allaman, I.; Magistretti, P.J. Brain energy metabolism: Focus on astrocyte-neuron metabolic cooperation. Cell Metab. 2011, 14, 724–738. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, O.W.; Fontes, J.D.; Carlson, G.M. The regulation of glycogenolysis in the brain. J. Biol. Chem. 2018, 293, 7099–7107. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, C.; Duval, R.; Cocaign, A.; Petit, E.; Bui, L.-C.; Haddad, I.; Vinh, J.; Etchebest, C.; Dupret, J.-M.; Rodrigues-Lima, F. An isozyme-specific redox switch in human brain glycogen phosphorylase modulates its allosteric activation by AMP. J. Biol. Chem. 2016, 291, 23842–23853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, C.; Dupret, J.-M.; Rodrigues-Lima, F. The structure of brain glycogen phosphorylase-from allosteric regulation mechanisms to clinical perspectives. FEBS J. 2016, 284, 546–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.S.; Pedersen, S.; Walls, A.B.; Waagepetersen, H.S.; Bak, L.K. Isoform-selective regulation of glycogen phosphorylase by energy deprivation and phosphorylation in astrocytes. Glia 2014, 63, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Alhamami, H.N.; Alshamrani, A.; Briski, K.P. Inhibition of glycogen phosphorylase stimulates ventromedial hypothalamic nucleus AMP-activated protein kinase. Physiol. Rep. 2017, 5, e13484. [Google Scholar] [CrossRef] [Green Version]
- Briski, K.P.; Alhamami, H.N.; Alshamrani, A.; Mandal, S.K.; Shakya, M.; Ibrahim, M.H.H. Sex differences and role of estradiol in hypoglycemia-associated counter-regulation. Taurine 6 2017, 1043, 359–383. [Google Scholar] [CrossRef]
- Borg, M.A.; Tamborlane, W.V.; Shulman, G.I.; Sherwin, R.S. Local lactate perfusion of the ventromedial hypothalamus suppresses hypoglycemic counterregulation. Diabetes 2003, 52, 663–666. [Google Scholar] [CrossRef] [Green Version]
- Chan, O.; Paranjape, S.A.; Horblitt, A.; Zhu, W.; Sherwin, R.S. Lactate-induced release of GABA in the ventromedial hypothalamus contributes to counterregulatory failure in recurrent hypoglycemia and diabetes. Diabetes 2013, 62, 4239–4246. [Google Scholar] [CrossRef] [Green Version]
- Alhamami, H.N.; Uddin, M.; Mahmood, A.H.; Briski, K.P. Lateral but not medial hypothalamic AMPK activation occurs at the hypoglycemic nadir in insulin-injected male rats: Impact of caudal dorsomedial hindbrain catecholamine signaling. Neuroscience 2018, 379, 103–114. [Google Scholar] [CrossRef]
- Mahmood, A.S.M.H.; Napit, P.R.; Ali, H.; Briski, K.P. Estrogen receptor involvement in noradrenergic regulation of ventromedial hypothalamic nucleus glucoregulatory neurotransmitter and stimulus-specific glycogen phosphorylase enzyme isoform expression. ASN Neuro 2020, 12, 1759091420910933. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.M.; Alhamami, H.N.; Briski, K.P. Norepinephrine regulation of ventromedial hypothalamic nucleus metabolic transmitter biomarker and astrocyte enzyme and receptor expression: Impact of 5′ AMP-activated protein kinase. Brain Res. 2019, 1711, 48–57. [Google Scholar] [CrossRef]
- Briski, K.P.; Mandal, S.K. Hindbrain lactoprivic regulation of hypothalamic neuron transactivation and gluco-regulatory neurotransmitter expression: Impact of antecedent insulin-induced hypoglycemia. Neuropeptides 2019, 77, 101962. [Google Scholar] [CrossRef] [PubMed]
- Briski, K.P.; Mandal, S.K. Hindbrain metabolic deficiency regulates ventromedial hypothalamic nucleus glycogen metabolism and glucose-regulatory signaling. Acta Neurobiol. Exp. 2020, 80, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Harik, S.; Busto, R.; Martínez, E. Norepinephrine regulation of cerebral glycogen utilization during seizures and ischemia. J. Neurosci. 1982, 2, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, T.; Soya, S.; Okamoto, M.; Ichitani, Y.; Kawanaka, K.; Soya, H. Brain glycogen decreases during prolonged exercise. J. Physiol. 2011, 589, 3383–3393. [Google Scholar] [CrossRef]
- Alshamrani, A.A.; Bheemanapally, K.; Ibrahim, M.M.; Briski, K.P. Impact of caudal hindbrain glycogen metabolism on A2 noradrenergic neuron AMPK activation and ventromedial hypothalamic nucleus norepinephrine activity and glucoregulatory neurotransmitter marker protein expression. Neuropeptides 2020, 82, 102055. [Google Scholar] [CrossRef]
- Ali, H.; Napit, P.R.; Mahmood, A.H.; Bheemanapally, K.; Alhamami, H.N.; Uddin, M.; Mandal, S.K.; Ibrahim, M.M.; Briski, K.P. Hindbrain estrogen receptor regulation of ventromedial hypothalamic glycogen metabolism and glucoregulatory transmitter expression in the hypoglycemic male rat. Neuroscience 2019, 409, 253–260. [Google Scholar] [CrossRef]
- Napit, P.R.; Ali, H.; Shakya, M.; Mandal, S.K.; Bheemanapally, K.; Mahmood, A.H.; Ibrahim, M.M.; Briski, K.P. Hindbrain estrogen receptor regulation of ventromedial hypothalamic glycogen metabolism and glucoregulatory transmitter expression in the hypoglycemic female rat. Neuroscience 2019, 411, 211–221. [Google Scholar] [CrossRef]
- Ibrahim, M.M.; Bheemanapally, K.; Sylvester, P.W.; Briski, K.P. Sex-specific estrogen regulation of hypothalamic astrocyte estrogen receptor expression and glycogen metabolism in rats. Mol. Cell. Endocrinol. 2020, 504, 110703. [Google Scholar] [CrossRef]
- Ibrahim, M.M.H.; Bheemanapally, K.; Sylvester, P.W.; Briski, K.P. Norepinephrine regulation of adrenergic receptor expression, 5′ AMP-activated protein kinase activity, and glycogen metabolism and mass in male versus female hypothalamic primary astrocyte cultures. ASN Neuro 2020, 12. [Google Scholar] [CrossRef]
- Azcoitia, I.; Yagüe, J.G.; García-Segura, L.M. Estradiol synthesis within the human brain. Neuroscience 2011, 191, 139–147. [Google Scholar] [CrossRef]
- Fester, L.; Brandt, N.; Windhorst, S.; Pröls, F.; Bläute, C.; Rune, G.M. Control of aromatase in hippocampal neurons. J. Steroid Biochem. Mol. Biol. 2016, 160, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.H.; Uddin, M.; Ibrahim, M.; Mandal, S.; Alhamami, H.; Briski, K.P. Sex differences in forebrain estrogen receptor regulation of hypoglycemic patterns of counter-regulatory hormone secretion and ventromedial hypothalamic nucleus glucoregulatory neurotransmitter and astrocyte glycogen metabolic enzyme expression. Neuropeptides 2018, 72, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Tamrakar, P.; Briski, K.P. Estradiol regulates hypothalamic astrocyte adenosine 5′-monophosphate-activated protein kinase activity: Role of hindbrain catecholamine signaling. Brain Res. Bull. 2015, 110, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Janzen, N.R.; Whitfield, J.; Hoffman, N.J. Interactive roles for AMPK and glycogen from cellular energy sensing to exercise metabolism. Int. J. Mol. Sci. 2018, 19, 3344. [Google Scholar] [CrossRef] [Green Version]
- DiNuzzo, M. Astrocyte-neuron interactions during learning may occur by lactate signaling rather than metabolism. Front. Integr. Neurosci. 2016, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugenin, E.A.; Valdebenito, S.; Gorska, A.M.; Martínez, A.D.; Bitran, M.; Sáez, J.C. Gap junctions coordinate the propagation of glycogenolysis induced by norepinephrine in the pineal gland. J. Neurochem. 2019, 151, 558–569. [Google Scholar] [CrossRef]
- Hinojosa, A.E.; García-Bueno, B.; Leza, J.C.; Madrigal, J.L.M. Regulation of CCL2/MCP-1 production in astrocytes by desipramine and atomoxetine: Involvement of α2 adrenergic receptors. Brain Res. Bull. 2011, 86, 326–333. [Google Scholar] [CrossRef]
- Morioka, N.; Abe, H.; Araki, R.; Matsumoto, N.; Zhang, F.F.; Nakamura, Y.; Nakata, Y.; Hisaoka-Nakashima, K. A β1/2 adrenergic receptor-sensitive intracellular signaling pathway modulates CCL2 production in cultured spinal astrocytes. J. Cell. Physiol. 2013, 229, 323–332. [Google Scholar] [CrossRef]
- Gutiérrez, I.L.; González-Prieto, M.; García-Bueno, B.; Caso, J.R.; Feinstein, D.L.; Madrigal, J.L.M. CCL2 induces the production of β2 adrenergic receptors and modifies astrocytic responses to noradrenaline. Mol. Neurobiol. 2018, 55, 7872–7885. [Google Scholar] [CrossRef]
- Subbarao, K.V.; Hertz, L. Effect of adrenergic agonists on glycogenolysis in primary cultures of astrocytes. Brain Res. 1990, 536, 220–226. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briski, K.P.; Ibrahim, M.M.H.; Mahmood, A.S.M.H.; Alshamrani, A.A. Norepinephrine Regulation of Ventromedial Hypothalamic Nucleus Astrocyte Glycogen Metabolism. Int. J. Mol. Sci. 2021, 22, 759. https://doi.org/10.3390/ijms22020759
Briski KP, Ibrahim MMH, Mahmood ASMH, Alshamrani AA. Norepinephrine Regulation of Ventromedial Hypothalamic Nucleus Astrocyte Glycogen Metabolism. International Journal of Molecular Sciences. 2021; 22(2):759. https://doi.org/10.3390/ijms22020759
Chicago/Turabian StyleBriski, Karen P., Mostafa M. H. Ibrahim, A. S. M. Hasan Mahmood, and Ayed A. Alshamrani. 2021. "Norepinephrine Regulation of Ventromedial Hypothalamic Nucleus Astrocyte Glycogen Metabolism" International Journal of Molecular Sciences 22, no. 2: 759. https://doi.org/10.3390/ijms22020759
APA StyleBriski, K. P., Ibrahim, M. M. H., Mahmood, A. S. M. H., & Alshamrani, A. A. (2021). Norepinephrine Regulation of Ventromedial Hypothalamic Nucleus Astrocyte Glycogen Metabolism. International Journal of Molecular Sciences, 22(2), 759. https://doi.org/10.3390/ijms22020759