Developing Tadpole Xenopus laevis as a Comparative Animal Model to Study Mycobacterium abscessus Pathogenicity

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
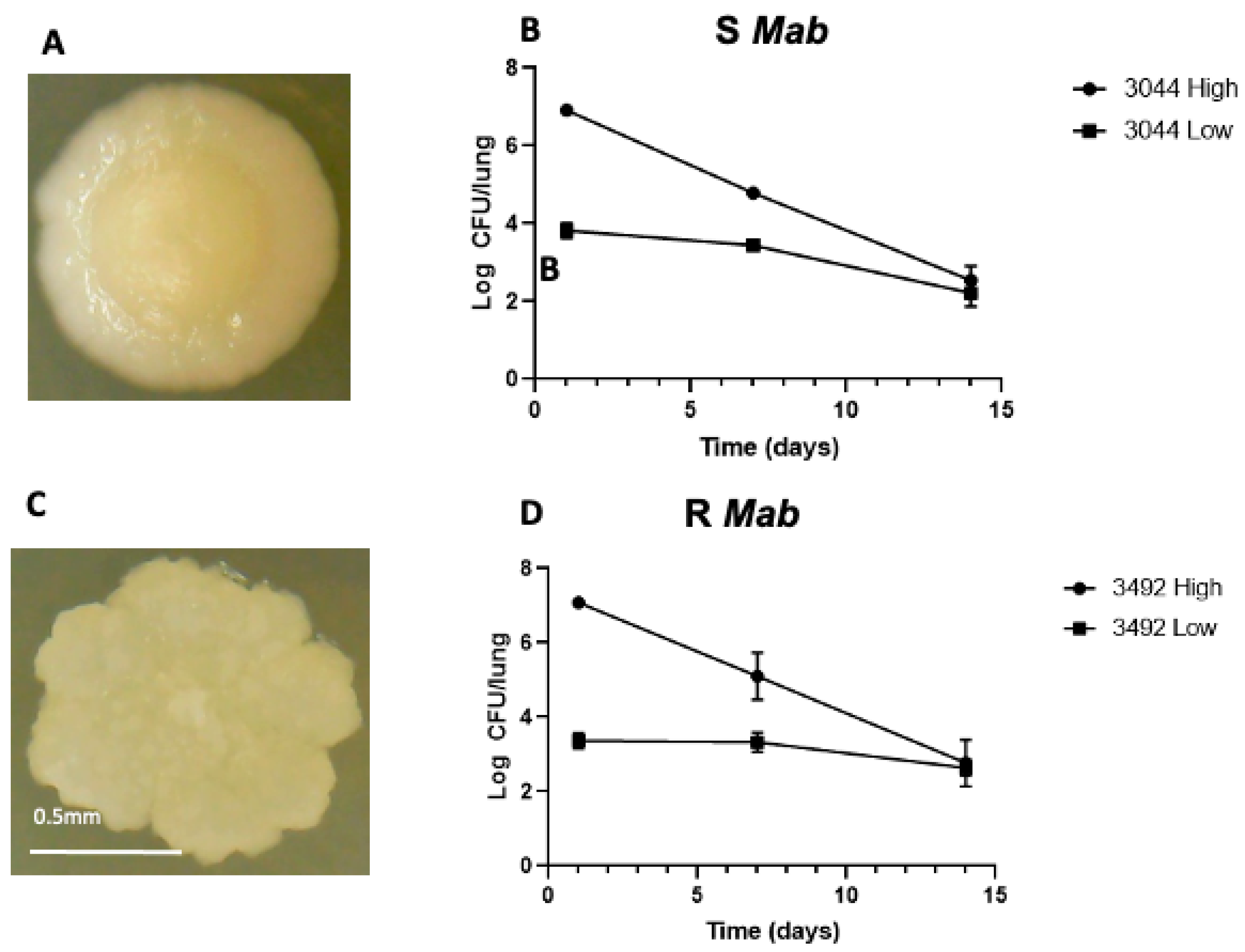
2.1. Clearance of Mab in Mice
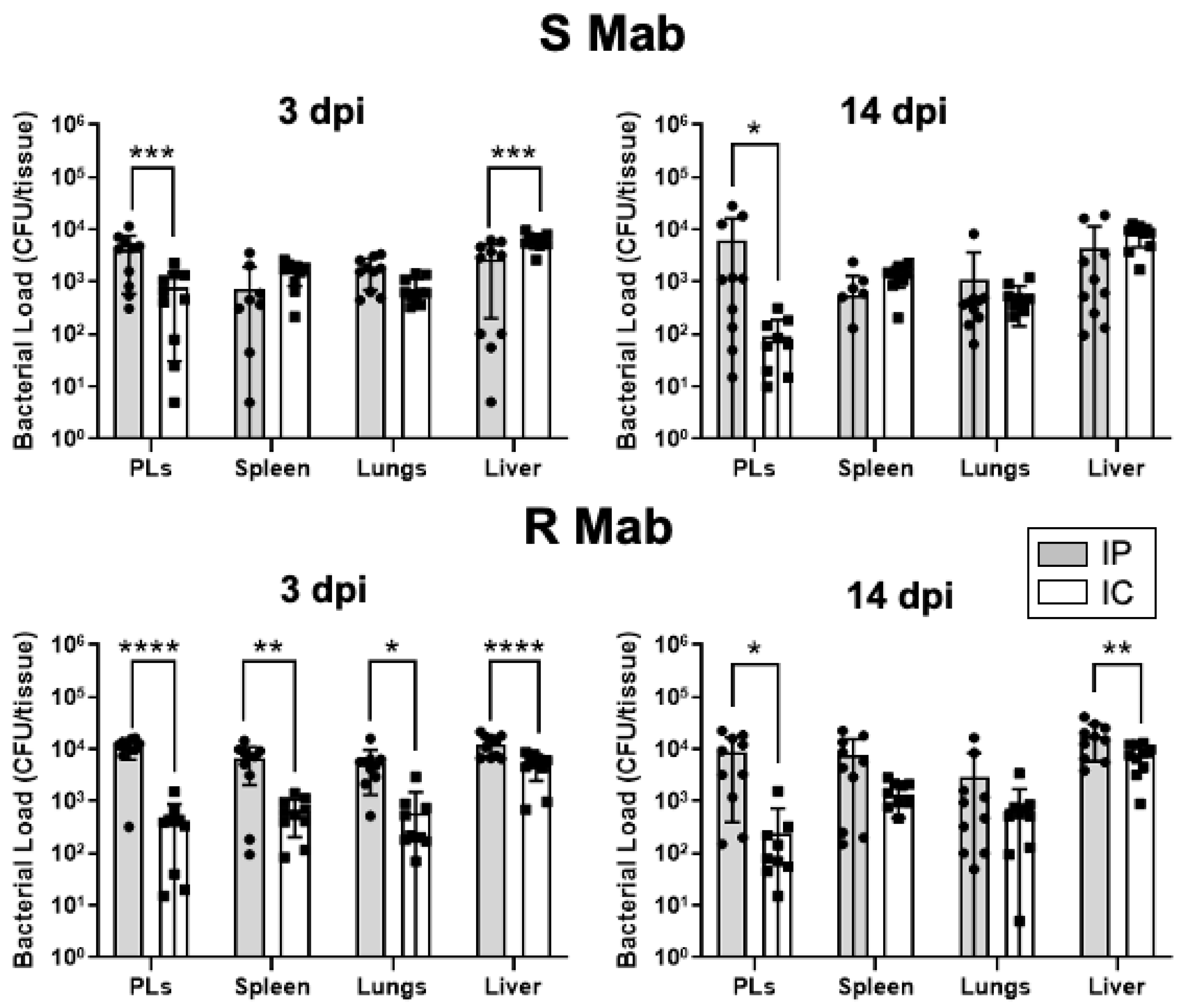
2.2. Dissemination and Persistence of S and R Mab in Intraperitoneally Infected X. laevis Tadpoles
2.3. Survival Analysis of Tadpole X. laevis Inoculated with S or R Mab
2.4. Dissemination and Persistence of S and R Mab in IC-Infected X. laevis Tadpoles
3. Discussion
4. Materials and Methods
4.1. Animal Husbandry
4.2. Mycobacterial Strains and Culture Conditions:
4.3. Mab Mouse Infection Model
4.4. Mab Inoculation in Tadpoles
4.5. Colony-Forming Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomson, R.; Tolson, C.; Carter, R.; Coulter, C.; Huygens, F.; Hargreaves, M. Isolation of Nontuberculous Mycobacteria (NTM) from Household Water and Shower Aerosols in Patients with Pulmonary Disease Caused by NTM. J. Clin. Microbiol. 2013, 51, 3006–3011. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.T.; Byrd, T.F. The rapidly growing mycobacteria: Saprophytes and parasites. Microbes Infect. 2000, 2, 1845–1853. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Wallace, R.J. Clinical and Taxonomic Status of Pathogenic Nonpigmented or Late-Pigmenting Rapidly Growing Mycobacteria. Clin. Microbiol. Rev. 2002, 15, 716–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nessar, R.; Cambau, E.; Reyrat, J.M.; Murray, A.; Gicquel, B. Mycobacterium abscessus: A new antibiotic nightmare. J. Antimicrob. Chemother. 2012, 67, 810–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catherinot, E.; Clarissou, J.; Etienne, G.; Ripoll, F.; Emile, J.F.; Daffe, M.; Perronne, C.; Soudais, C.; Gaillard, J.L.; Rottman, M. Hypervirulence of a rough variant of the Mycobacte-rium abscessus type strain. Infect. Immun. 2007, 75, 1055–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piersimoni, C.; Scarparo, C. Pulmonary infections associated with non-tuberculous mycobacteria in immunocompetent pa-tients. Lancet Infect. Dis. 2008, 8, 323–334. [Google Scholar] [CrossRef]
- Esther, C.R., Jr.; Esserman, D.A.; Gilligan, P.; Kerr, A.; Noone, P.G. Chronic Mycobacterium abscessus infection and lung function decline in cystic fibrosis. J. Cyst. Fibros. 2010, 9, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-R.; Sheng, W.-H.; Hung, C.-C.; Yu, C.-J.; Lee, L.-N.; Hsueh, P.-R. Mycobacterium abscessusComplex Infections in Humans. Emerg. Infect. Dis. 2015, 21, 1638–1646. [Google Scholar] [CrossRef]
- Bernut, A.; Herrmann, J.-L.; Ordway, D.; Kremer, L. The diverse cellular and animal models to decipher the physiopathological traits of Mycobacterium abscessus infection. Front. Cell. Infect. Microbiol. 2017, 7, 100. [Google Scholar] [CrossRef] [Green Version]
- Medjahed, H.; Gaillard, J.-L.; Reyrat, J.-M. Mycobacterium abscessus: A new player in the mycobacterial field. Trends Microbiol. 2010, 18, 117–123. [Google Scholar] [CrossRef]
- Alderwick, L.; Harrison, J.; Lloyd, G.S.; Birch, H.L. The Mycobacterial Cell Wall—Peptidoglycan and Arabinogalactan. Cold Spring Harb. Perspect. Med. 2015, 5, a021113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, A.V.; Viljoen, A.; Ghigo, E.; Herrmann, J.-L.; Kremer, L. Glycopeptidolipids, a Double-Edged Sword of the Mycobacterium abscessus Complex. Front. Microbiol. 2018, 9, 1145. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.T.; Rhoades, E.; Recht, J.; Pang, X.; Alsup, A.; Kolter, R.; Lyons, C.R.; Byrd, T.F. Spontaneous reversion of Mycobacterium abscessus from a smooth to a rough morphotype is associated with reduced expression of glycopeptidolipid and reacquisition of an invasive phenotype. Microbiology 2006, 152, 1581–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd, T.F.; Lyons, C.R. Preliminary Characterization of aMycobacterium abscessus Mutant in Human and Murine Models of Infection. Infect. Immun. 1999, 67, 4700–4707. [Google Scholar] [CrossRef] [Green Version]
- Nessar, R.; Reyrat, J.-M.; Davidson, L.B.; Byrd, T.F. Deletion of the mmpL4b gene in the Mycobacterium abscessus glycopep-tidolipid biosynthetic pathway results in loss of surface colonization capability, but enhanced ability to replicate in human macrophages and stimulate their innate immune response. Microbiology 2011, 157, 1187–1195. [Google Scholar] [CrossRef] [Green Version]
- Park, I.K.; Hsu, A.P.; Tettelin, H.; Shallom, S.J.; Drake, S.K.; Ding, L.; Wu, U.-I.; Adamo, N.; Prevots, D.R.; Olivier, K.N.; et al. Clonal Diversification and Changes in Lipid Traits and Colony Morphology in Mycobacterium abscessus Clinical Isolates. J. Clin. Microbiol. 2015, 53, 3438–3447. [Google Scholar] [CrossRef] [Green Version]
- Recht, J.; Kolter, R. Glycopeptidolipid Acetylation Affects Sliding Motility and Biofilm Formation in Mycobacterium smegmatis. J. Bacteriol. 2001, 183, 5718–5724. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, E.P.; Elloumi, H.Z.; Zelazny, A.; Ding, L.; Paulson, M.L.; Sher, A.; Bafica, A.L.; Shea, Y.R.; Holland, S.M. Mycobacterium abscessus and M. avium trigger Toll-like receptor 2 and distinct cytokine response in human cells. Am. J. Respir. Cell Mol. Biol. 2008, 39, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Malcolm, K.C.; Nichols, E.M.; Caceres, S.M.; Kret, J.E.; Martiniano, S.L.; Sagel, S.D.; Chan, E.D.; Caverly, L.; Solomon, G.M.; Reynolds, P.; et al. Mycobacterium abscessus Induces a Limited Pattern of Neutrophil Activation That Promotes Pathogen Survival. PLoS ONE 2013, 8, e57402. [Google Scholar] [CrossRef]
- Chan, E.D.; Bai, X.; Kartalija, M.; Orme, I.M.; Ordway, D. Host Immune Response to Rapidly Growing Mycobacteria, an Emerging Cause of Chronic Lung Disease. Am. J. Respir. Cell Mol. Biol. 2010, 43, 387–393. [Google Scholar] [CrossRef]
- Ryan, K.; Byrd, T.F. Mycobacterium abscessus: Shapeshifter of the Mycobacterial World. Front. Microbiol. 2018, 9, 2642. [Google Scholar] [CrossRef]
- Bernut, A.; Herrmann, J.L.; Kissa, K.; Dubremetz, J.F.; Gaillard, J.L.; Lutfalla, G.; Kremer, L. Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation. Proc. Natl. Acad. Sci. USA 2014, 111, E943–E952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obregón-Henao, A.; Arnett, K.A.; Henao-Tamayo, M.; Massoudi, L.; Creissen, E.; Andries, K.; Lenaerts, A.J.; Ordway, D. Susceptibility of Mycobacterium abscessus to Antimycobacterial Drugs in Preclinical Models. Antimicrob. Agents Chemother. 2015, 59, 6904–6912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, C.S.; James, B. Plasticity of lung development in the amphibian, Xenopus laevis. Biol. Open 2013, 2, 1324–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, J.; Ohta, Y. Comparative and developmental study of the immune system inXenopus. Dev. Dyn. 2009, 238, 1249–1270. [Google Scholar] [CrossRef] [Green Version]
- Rhoo, K.H.; Edholm, E.S.; Forzán, M.J.; Khan, A.; Waddle, A.W.; Pavelka, M.S.; Robert, J. Distinct Host–Mycobacterial Pathogen Interac-tions between Resistant Adult and Tolerant Tadpole Life Stages of Xenopus laevis. J. Immunol. 2019, 203, 2679–2688. [Google Scholar]
- Manabe, Y.C.; Bishai, W.R. Latent Mycobacterium tuberculosis–persistence, patience and winning by waiting. Nat. Med. 2000, 6, 1327–1329. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Schwager, J.; Flajnik, M.F. The Immune System of Xenopus. Annu. Rev. Immunol. 1989, 7, 251–275. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; Blair, P.J.; Davis, A.T. Thymus Ontogeny in Frogs: T-Cell Renewal at Metamorphosis. Dev. Immunol. 1992, 2, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Nieuwkoop, P.; Faber, J. Normal Table of Xenopus Laevis (Daudin); Garland Publishing: New York, NY, USA, 1994; p. 252. [Google Scholar]
- Gregoire, S.A.; Byam, J.; Pavelka, M.S., Jr. Galk-based suicide vector mediated allelic exchange in Mycobacterium abscessus. Microbiology 2017, 163, 1399. [Google Scholar] [CrossRef]
- Zahn, N.; Levin, M.; Adams, D.S. The Zahn drawings: New illustrations of Xenopus embryo and tadpole stages for studies of craniofacial development. Development 2017, 144, 2708–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, A.; Shoen, C.; Cynamon, M.; Dimitrakopoulou, D.; Paiola, M.; Pavelka, M.S., Jr.; Robert, J. Developing Tadpole Xenopus laevis as a Comparative Animal Model to Study Mycobacterium abscessus Pathogenicity. Int. J. Mol. Sci. 2021, 22, 806. https://doi.org/10.3390/ijms22020806
Lopez A, Shoen C, Cynamon M, Dimitrakopoulou D, Paiola M, Pavelka MS Jr., Robert J. Developing Tadpole Xenopus laevis as a Comparative Animal Model to Study Mycobacterium abscessus Pathogenicity. International Journal of Molecular Sciences. 2021; 22(2):806. https://doi.org/10.3390/ijms22020806
Chicago/Turabian StyleLopez, Arianna, Carolyn Shoen, Michael Cynamon, Dionysia Dimitrakopoulou, Matthieu Paiola, Martin S. Pavelka, Jr., and Jacques Robert. 2021. "Developing Tadpole Xenopus laevis as a Comparative Animal Model to Study Mycobacterium abscessus Pathogenicity" International Journal of Molecular Sciences 22, no. 2: 806. https://doi.org/10.3390/ijms22020806
APA StyleLopez, A., Shoen, C., Cynamon, M., Dimitrakopoulou, D., Paiola, M., Pavelka, M. S., Jr., & Robert, J. (2021). Developing Tadpole Xenopus laevis as a Comparative Animal Model to Study Mycobacterium abscessus Pathogenicity. International Journal of Molecular Sciences, 22(2), 806. https://doi.org/10.3390/ijms22020806