
Promising Antiviral Activities of Natural Flavonoids against SARS-CoV-2 Targets: Systematic Review

 ,
,  ,
,  ,
,  and
and
Abstract
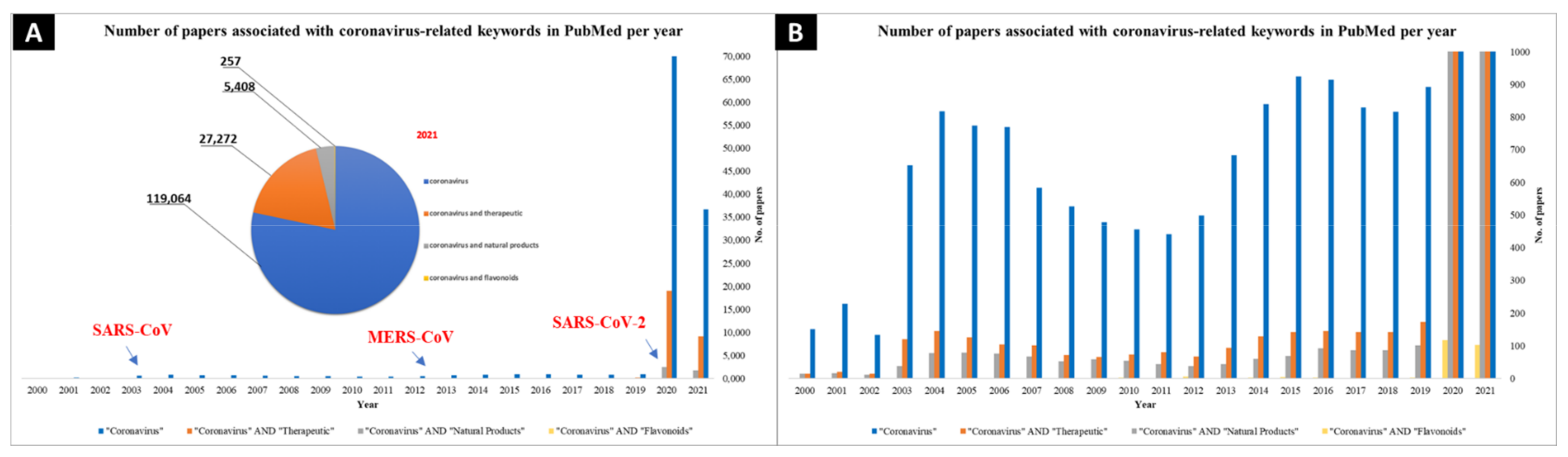
:1. Introduction
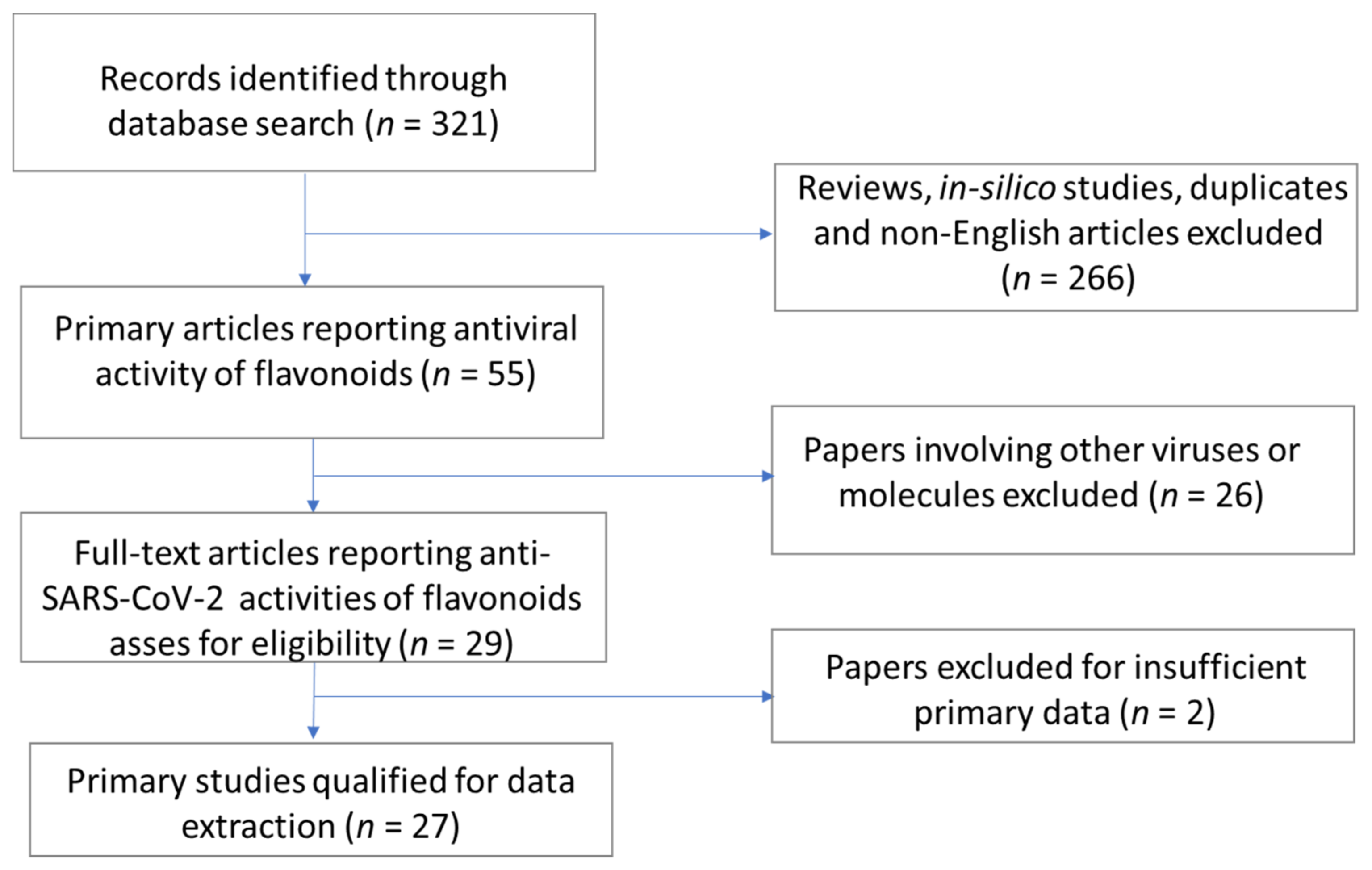
2. Methods
3. Results and Discussion
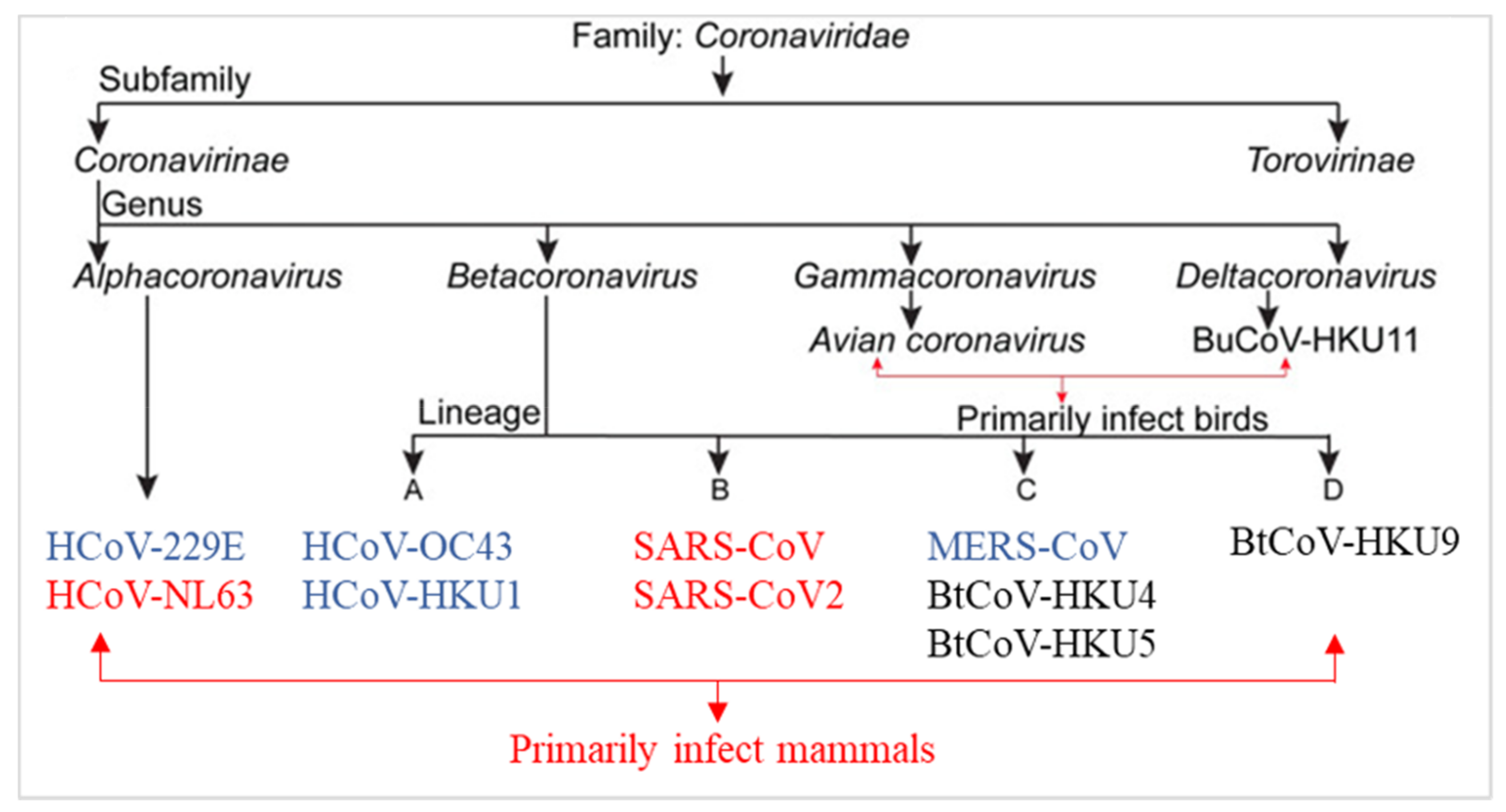
3.1. Coronaviruses Biology and Therapeutic Strategies for the Treatment of COVID-19 Infection
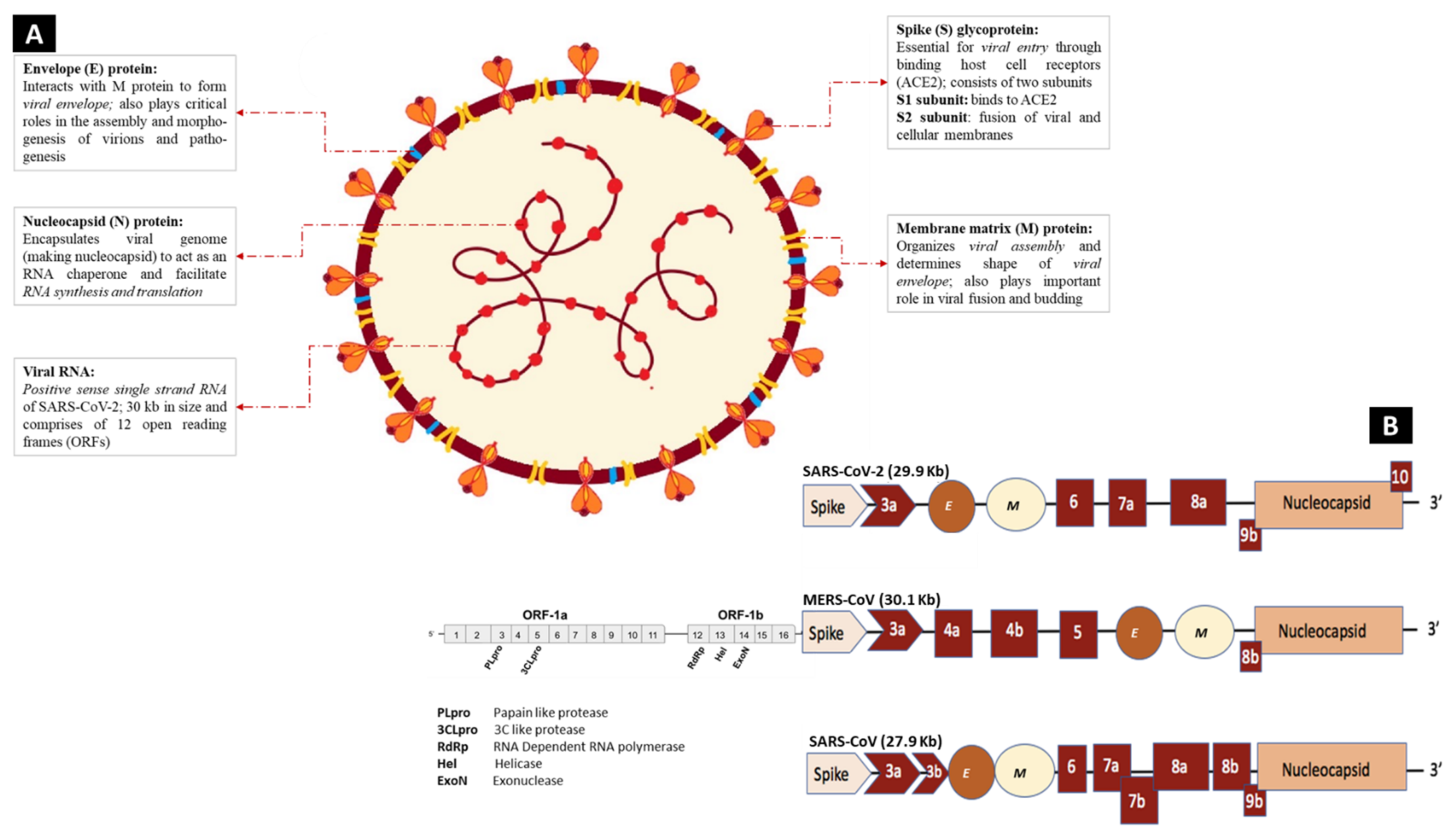
3.1.1. Genomic Characterization and Structure of SARS-CoV-2
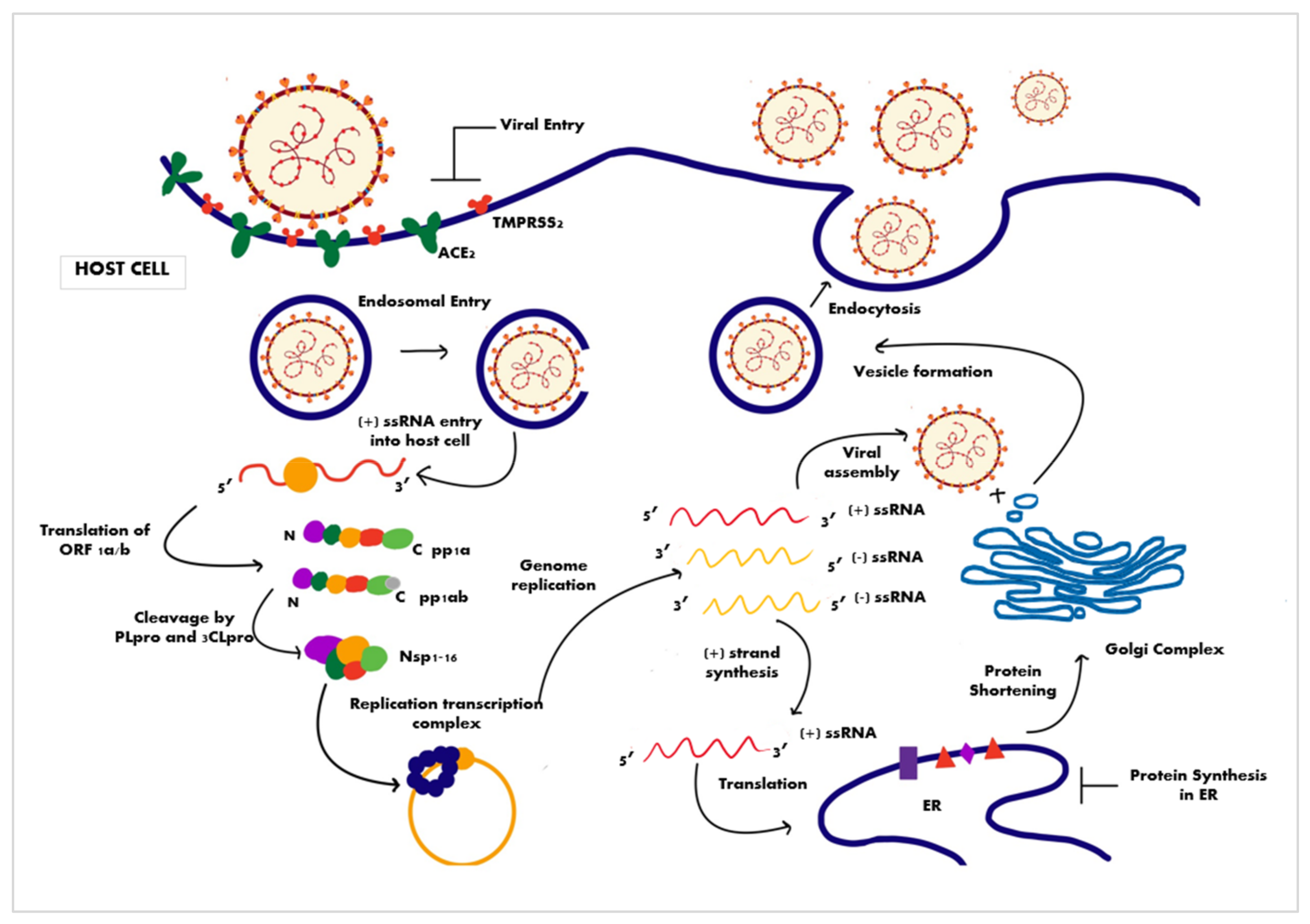
3.1.2. Mechanism of Cell Entry and Life Cycle of the Virus
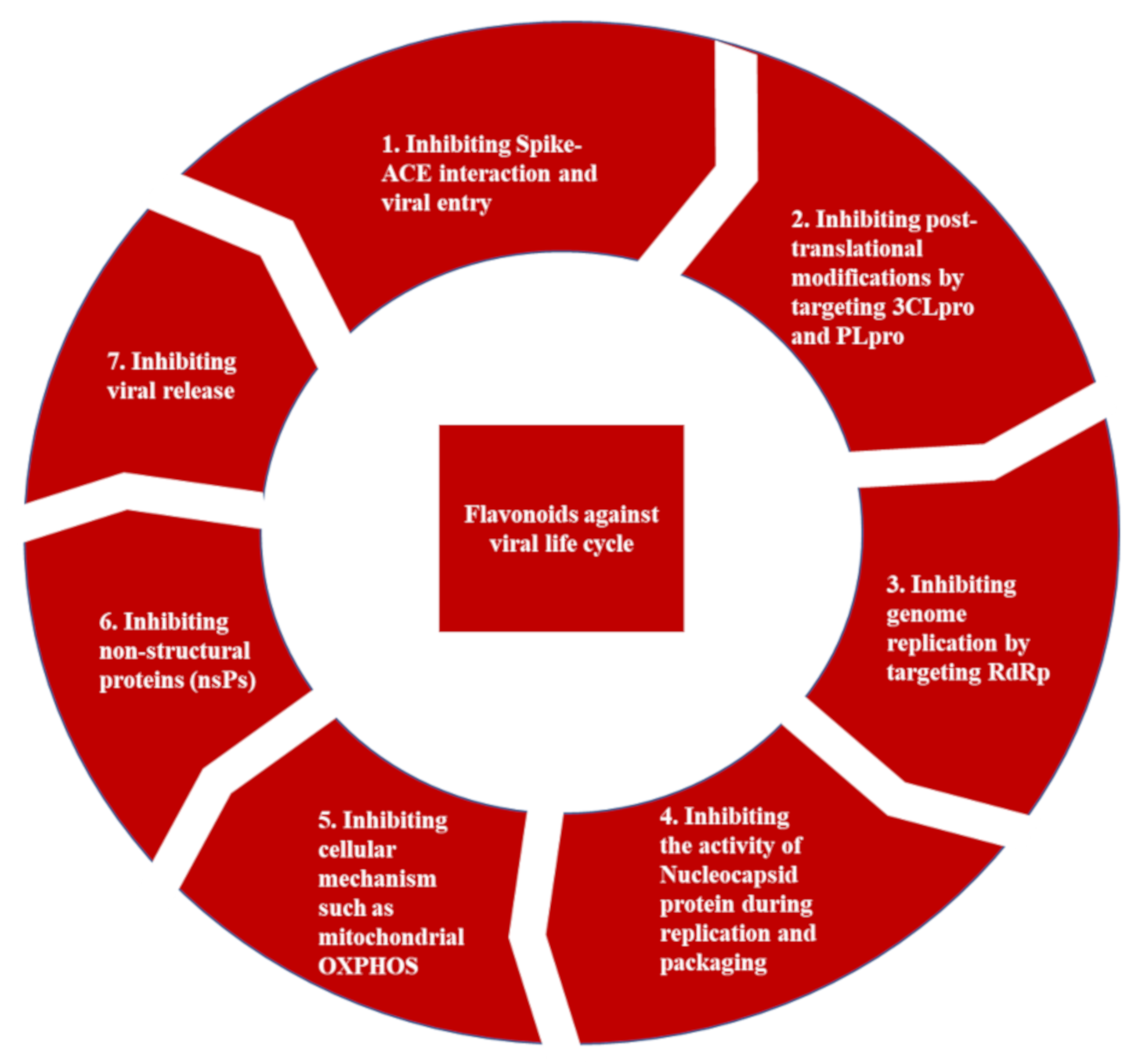
3.1.3. Promising Therapeutic Strategies for the Treatment of COVID-19 Infection
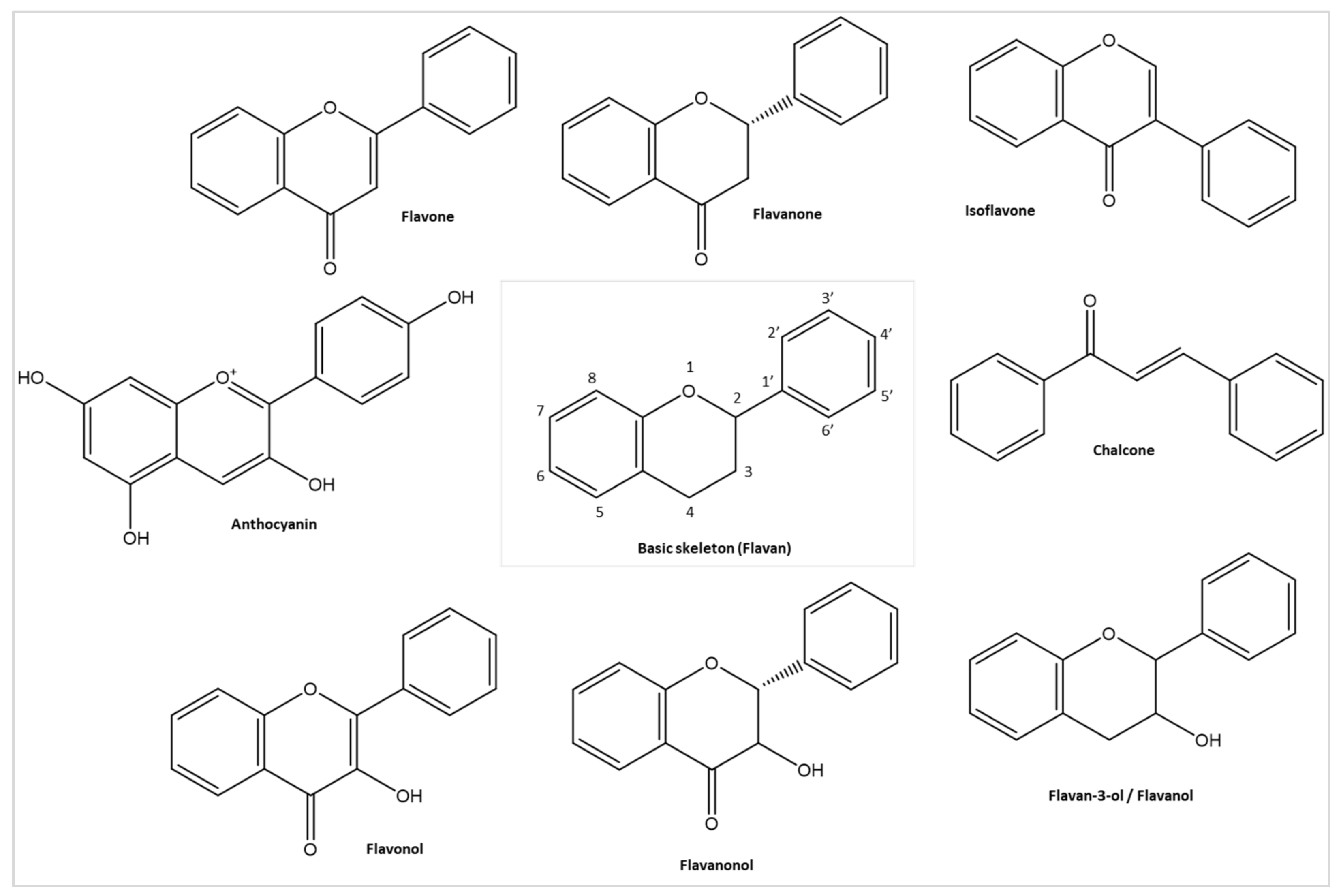
3.2. Flavonoids and Their Antiviral Properties
3.2.1. Antiviral Activities of Flavonoid against Non-Respiratory Viruses
3.2.2. Antiviral Activities of Flavonoid against Respiratory Viruses
3.3. Antiviral Properties against Coronaviruses, Including SARS-CoV and MERS-CoV
3.3.1. SARS-CoV
3.3.2. MERS-CoV
3.4. Antiviral Activity of Flavonoids against SARS-CoV-2
3.4.1. Antiviral Activity of Flavonoids against SARS-CoV-2 Proteases (3CLpro and PLpro)
Flavonols and Flavanonols
Flavones, Flavanones, and Isoflavones
Flavan-3-ols/Flavanols
Others
3.4.2. Flavonoids against SARS-CoV-2 Spike RBD and hACE2 Interaction
Flavonols
Flavones and Flavanones
Flavan-3-ols/Flavanols and Others
3.4.3. Antiviral Activities of Flavonoids against Other SARS-CoV-2 Targets
Flavones against SARS-CoV-2 RNA-Dependent RNA Polymerase (RdRp)
Flavan-3-ols/Flavanols against SARS-CoV-2 Nucleocapsid Protein
Flavonoids against Other SARS-CoV-2 Targets
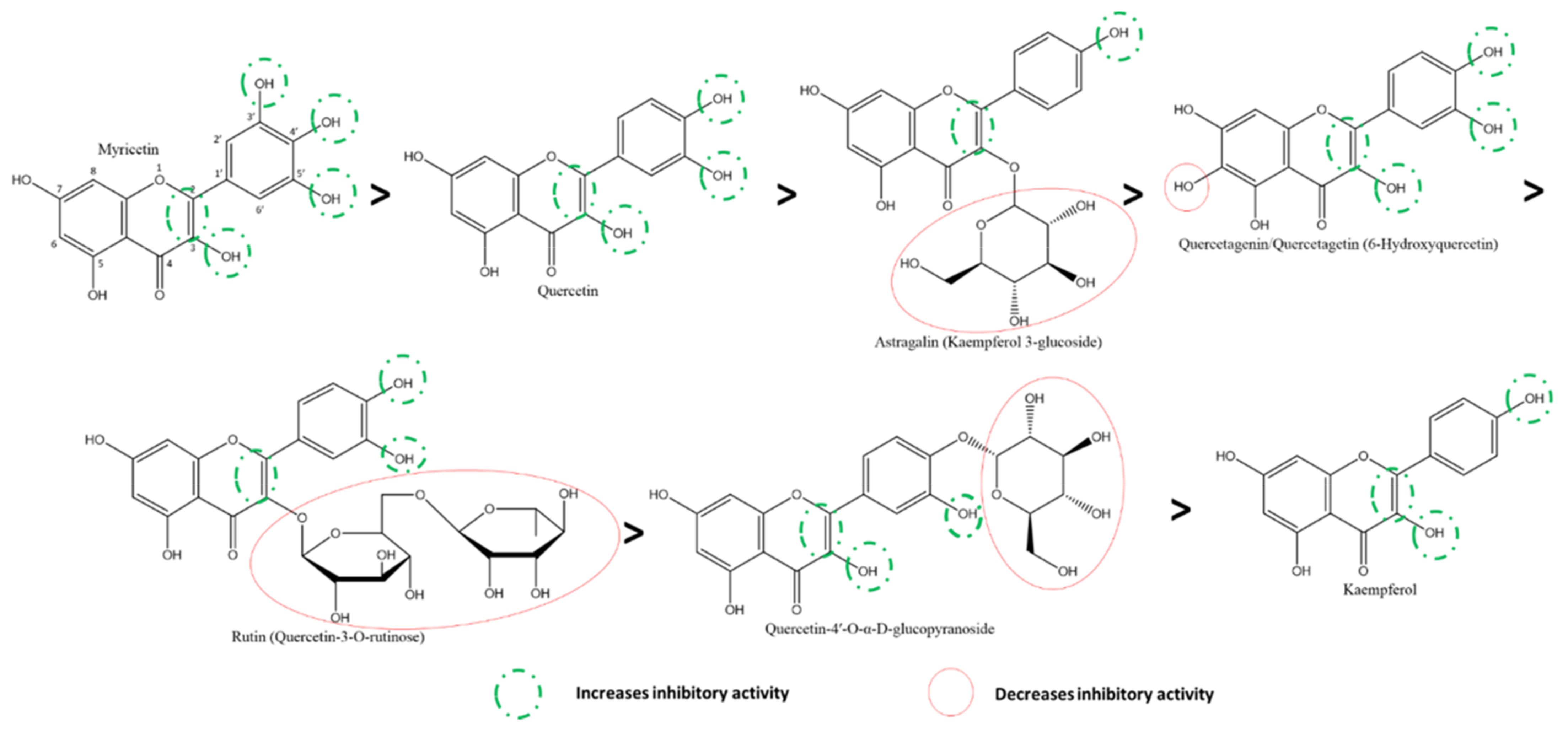
3.5. Structure-Activity Relationships (SARs) of Flavonoids
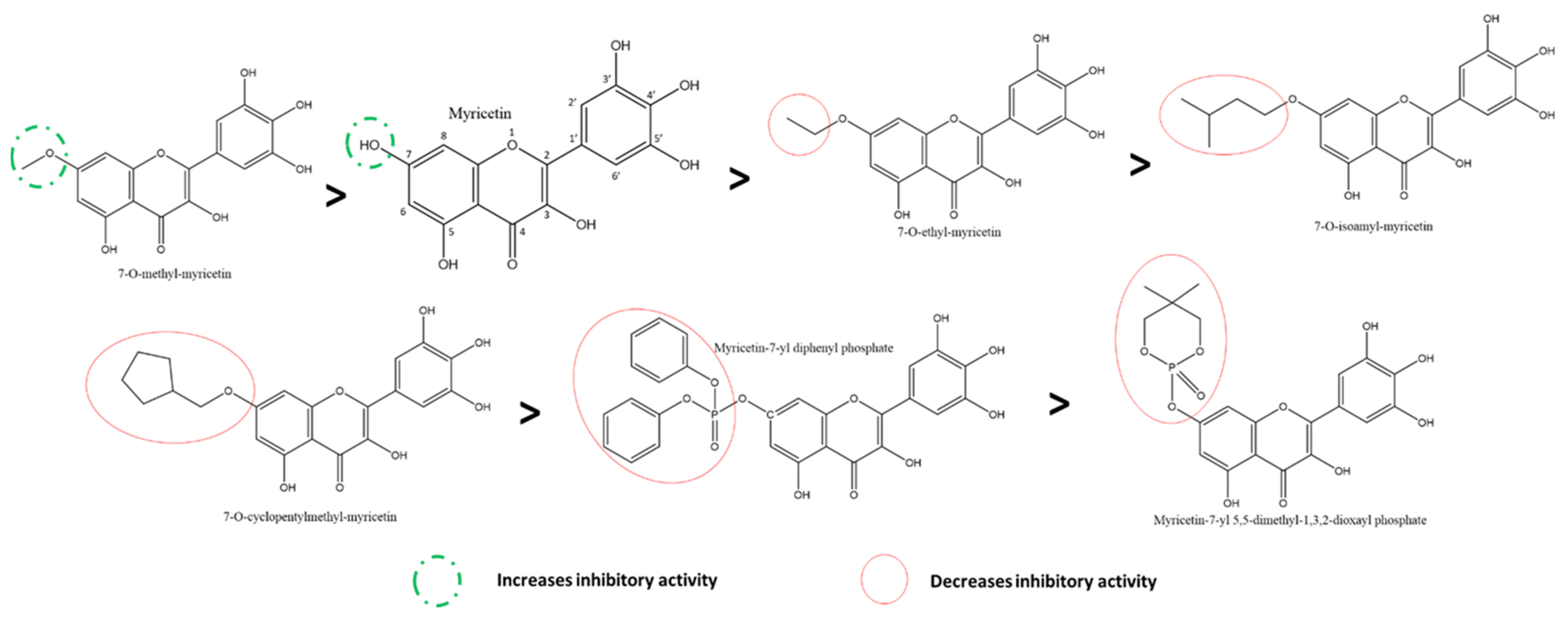
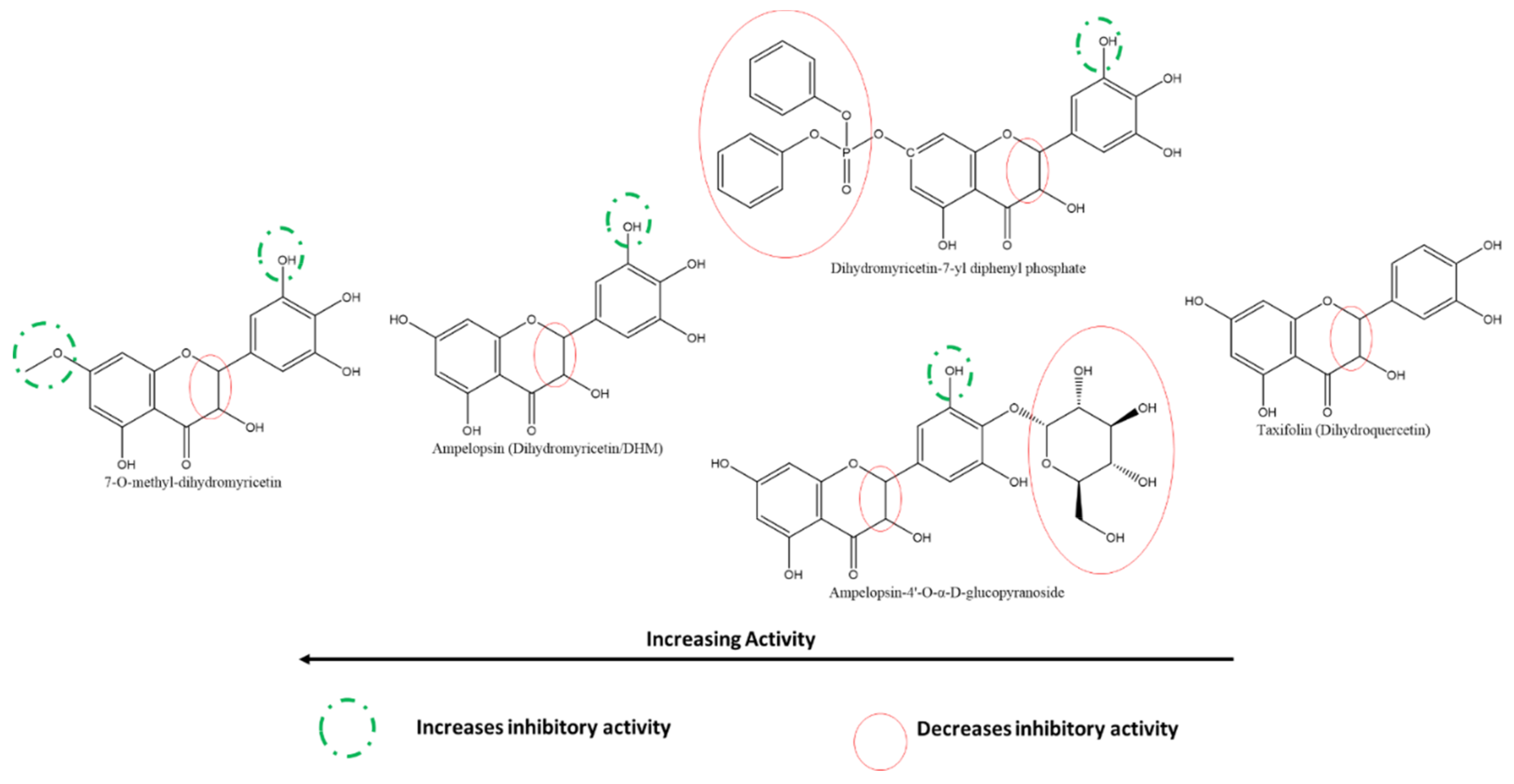
3.5.1. Effect of Flavonols and Flavanonols on SARS-CoV-2 3CLpro
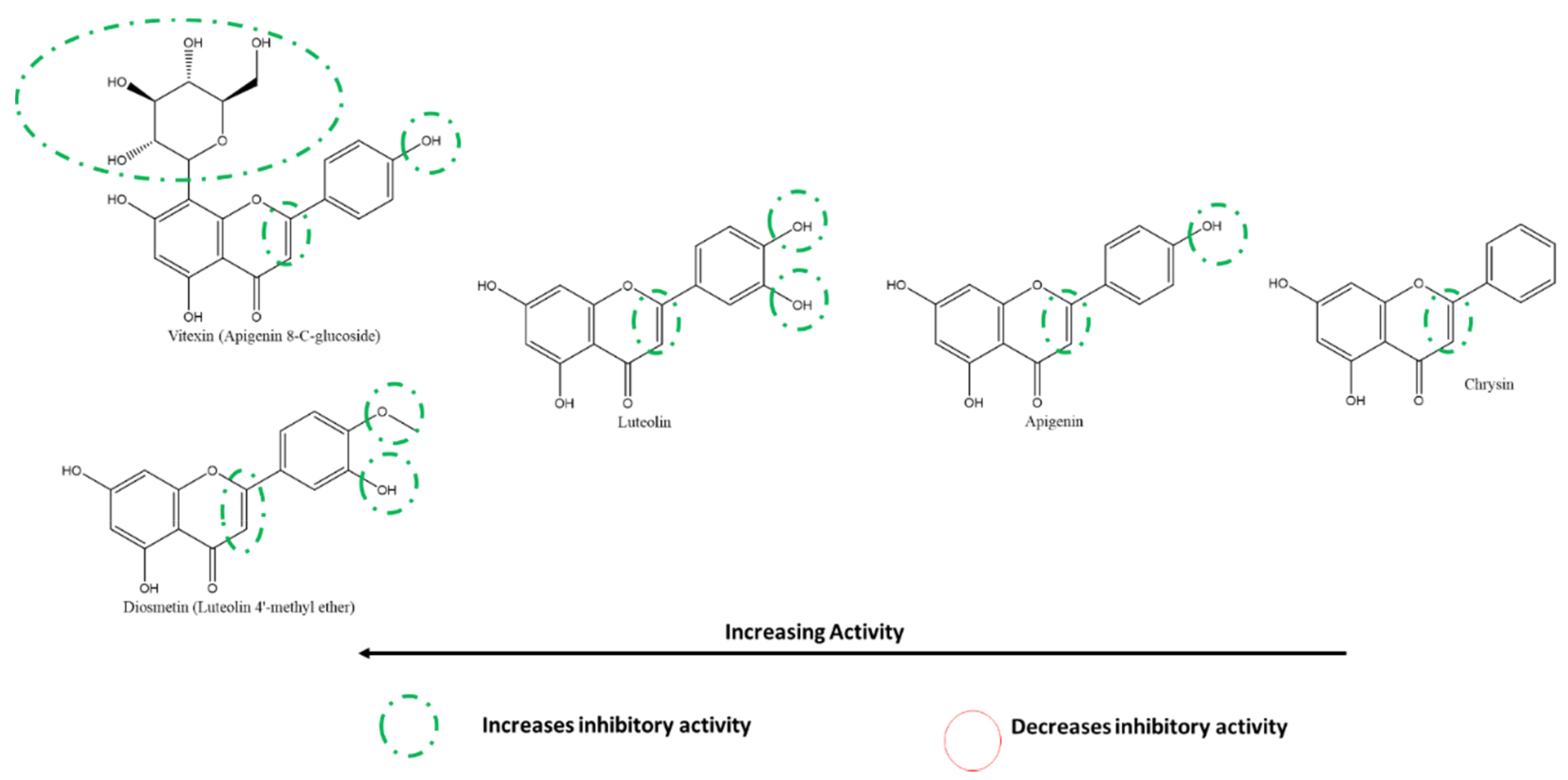
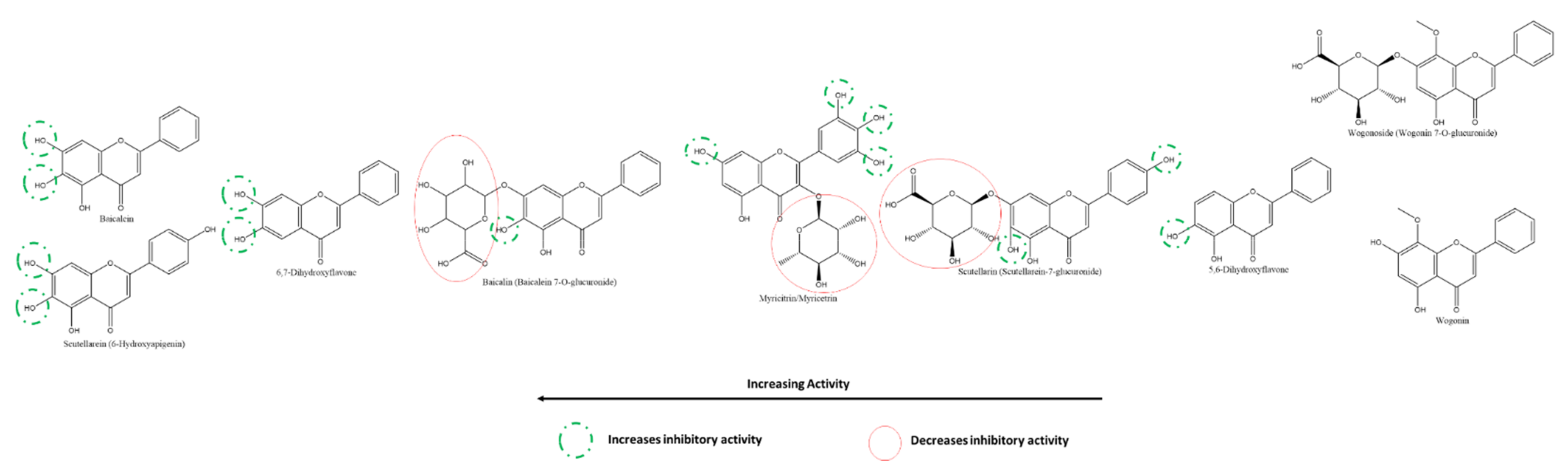
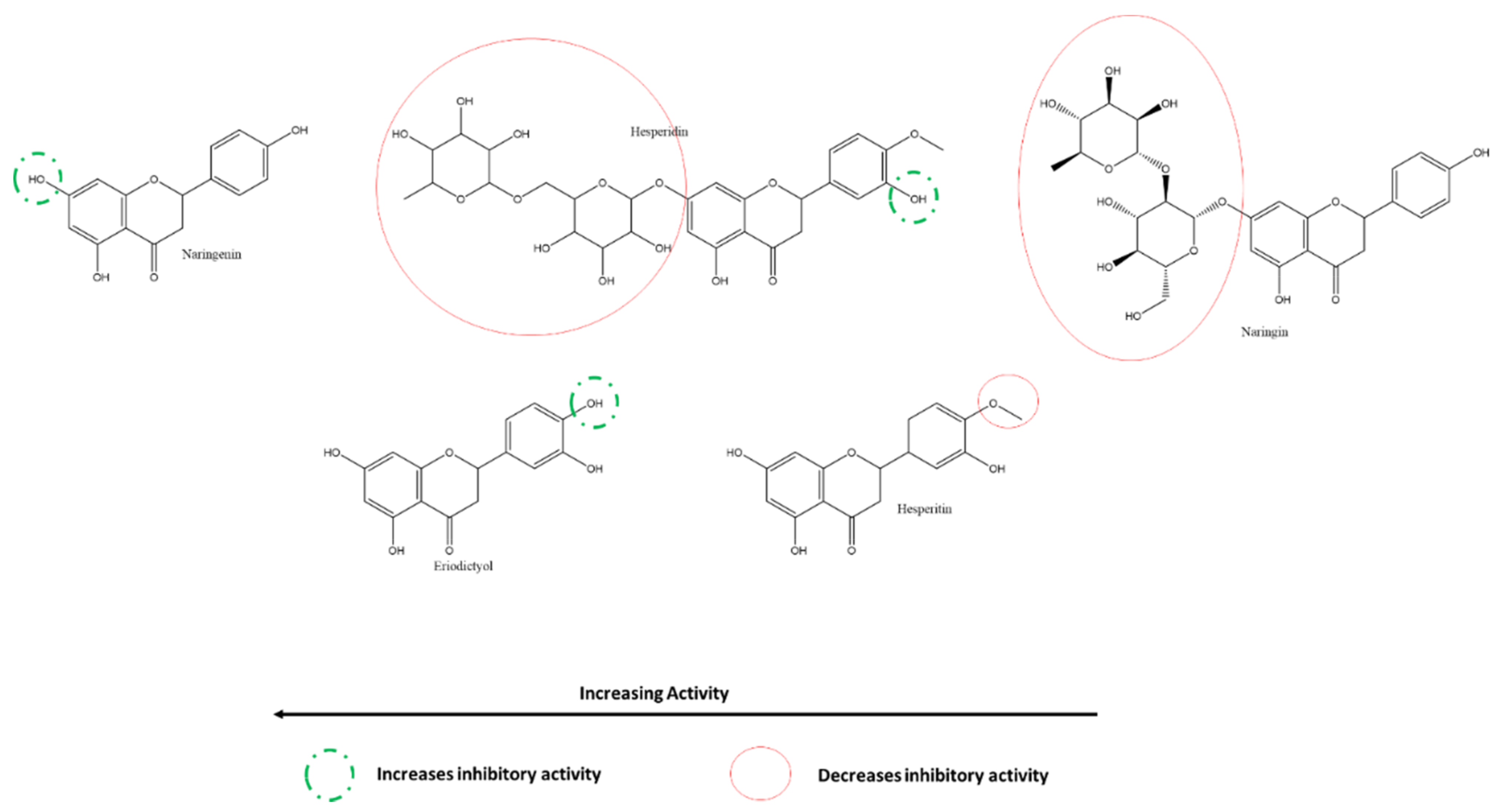
3.5.2. Effect of Flavones and Flavanones on SARS-CoV-2 3CLpro
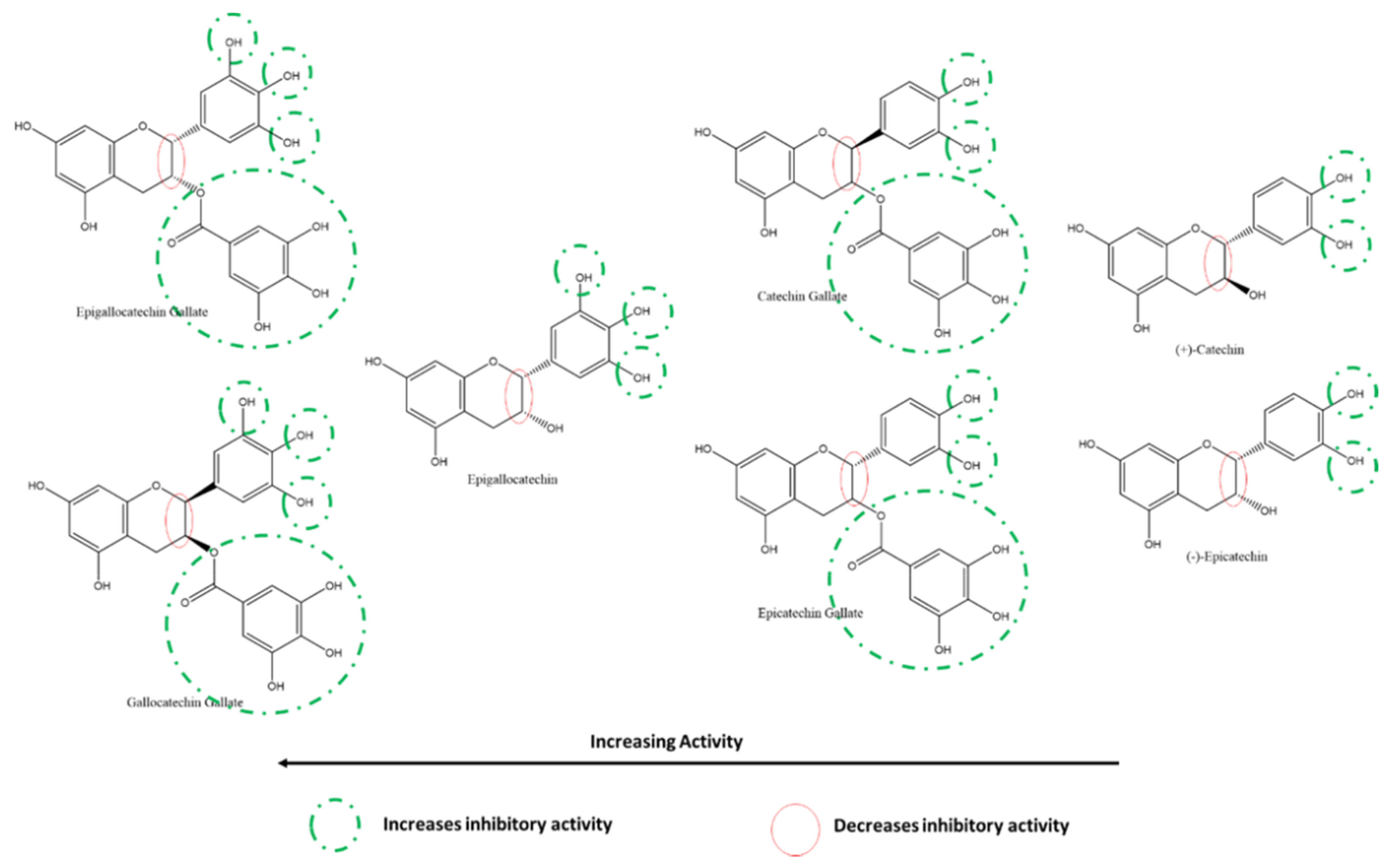
3.5.3. Effect of Flavan-3-ols/Flavanols on SARS-CoV-2 3CLpro
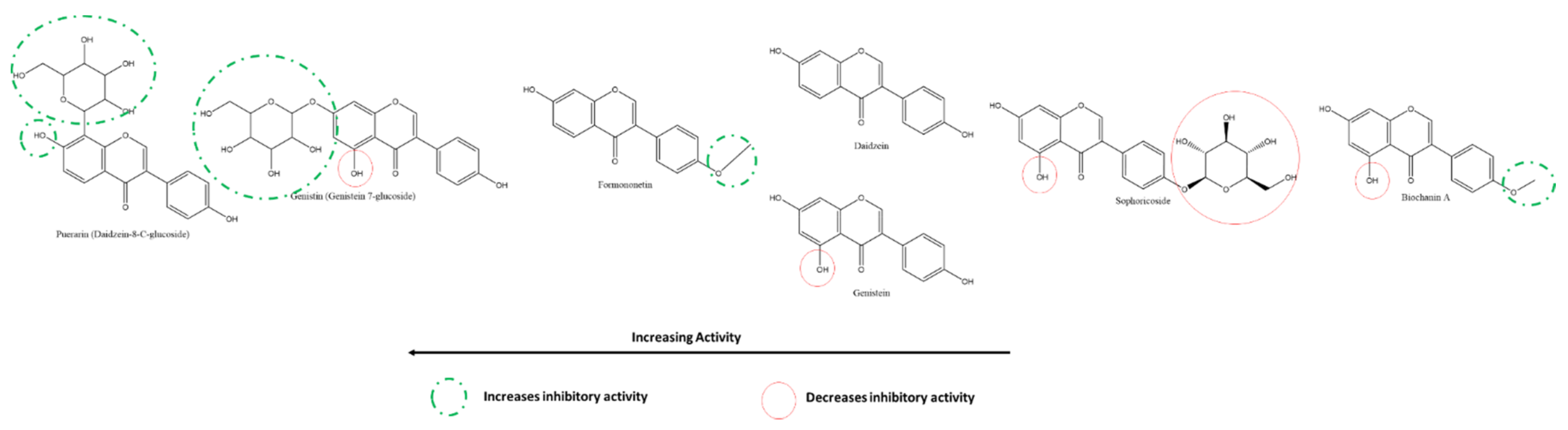
3.5.4. Effect of Isoflavones on SARS-CoV-2 3CLpro
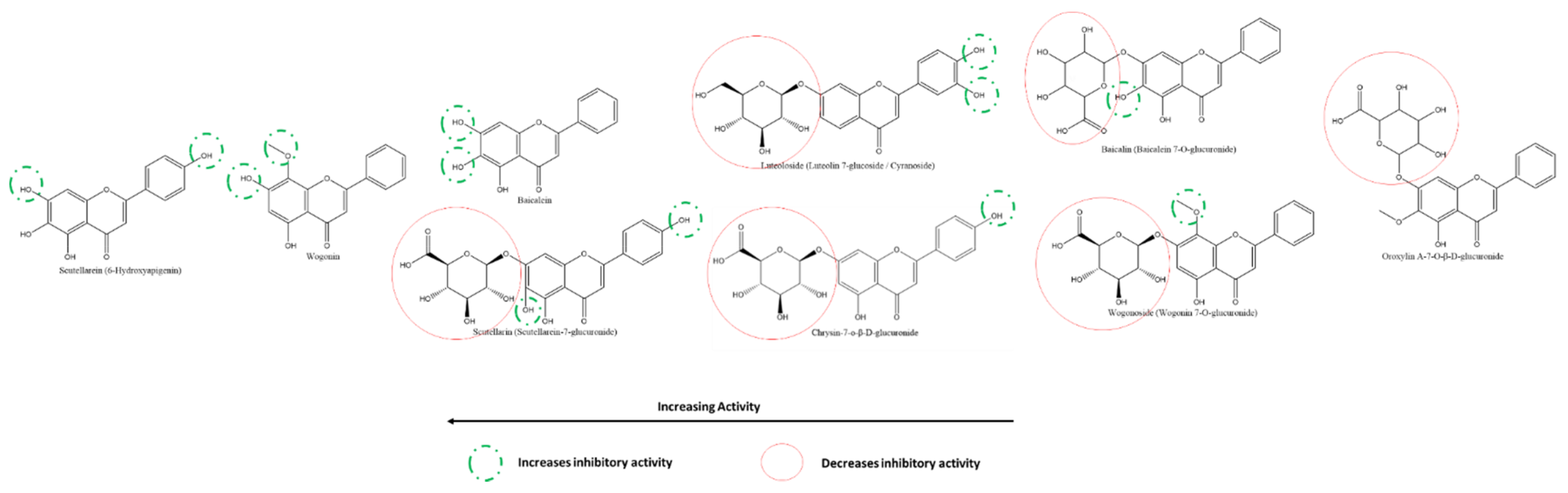
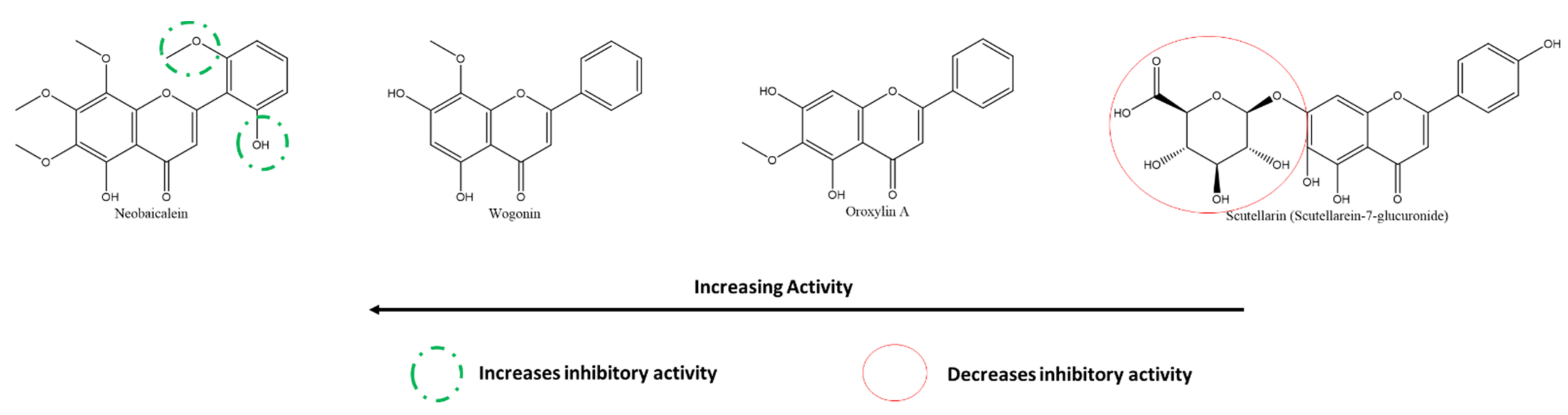
3.5.5. Effect of Flavones on SARS-CoV-2 PLpro
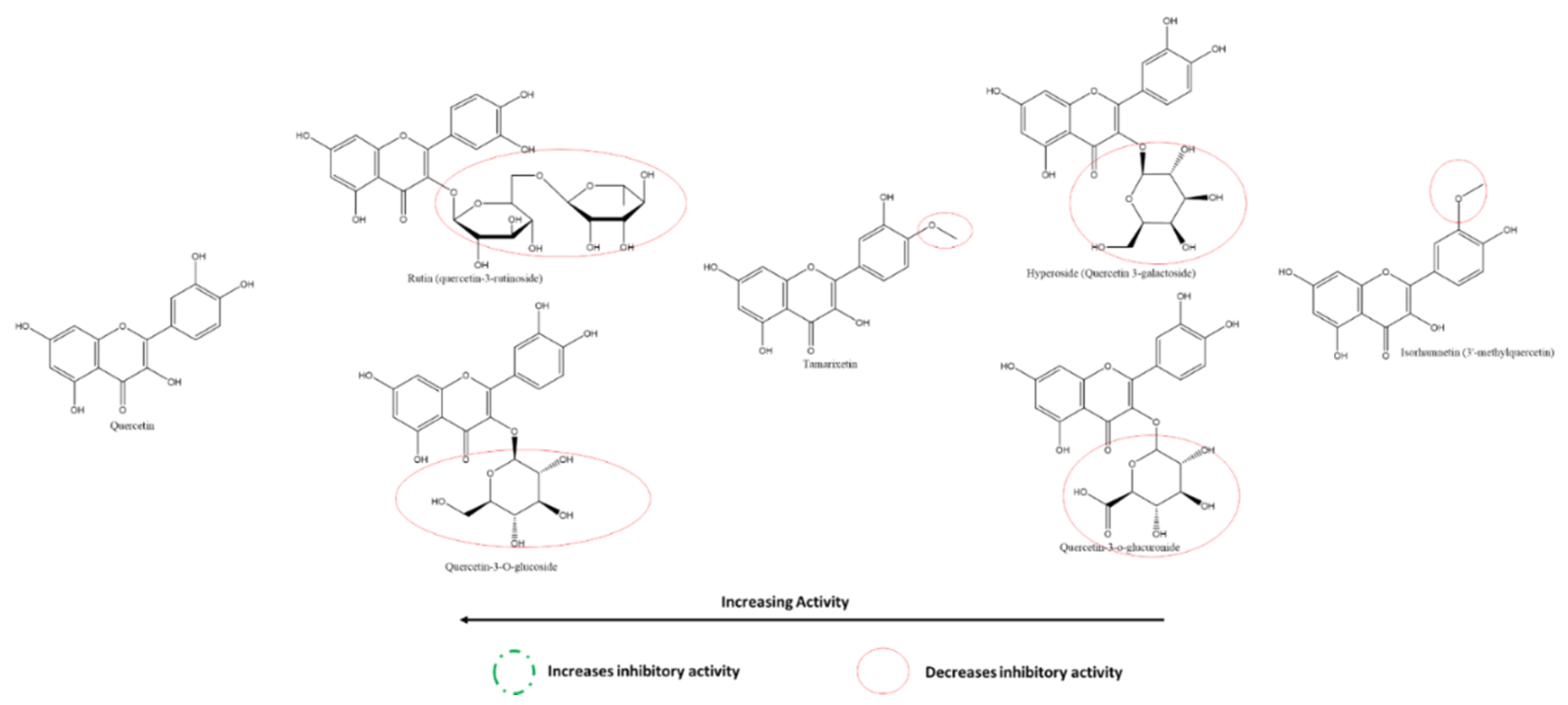
3.5.6. Effect of Flavonols on SARS-CoV-2 Spike Protein and hACE2 Receptor Interaction
3.5.7. Effect of Flavones on SARS-CoV-2 Spike Protein and hACE2 Receptor Interaction
3.6. Flavonoids as Potential Inhibitors of SARS-CoV-2 Proteins: In Silico Studies
3.6.1. Flavonoids against SARS-CoV-2 MPro
3.6.2. Flavonoids against SARS-CoV-2 Spike Glycoprotein
3.6.3. Flavonoids against SARS-CoV-2 PLPro
3.6.4. Flavonoids against SARS-CoV-2 RdRp
3.6.5. Flavonoids against SARS-CoV-2 Helicase
3.6.6. Flavonoids against SARS-CoV-2 ACE2
3.7. Clinical Trials and Future Prospects
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, M.; Yu, Y.; Sun, L.-M.; Xing, J.-Q.; Li, T.; Zhu, Y.; Wang, M.; Yu, Y.; Xue, W.; Xia, T.; et al. GCG inhibits SARS-CoV-2 replication by disrupting the liquid phase condensation of its nucleocapsid protein. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 17 August 2021).
- “About Variants of the Virus that Causes COVID-19 | CDC”. Available online: https://stacks.cdc.gov/view/cdc/104698 (accessed on 15 August 2021).
- Loganathan, S.K.; Schleicher, K.; Malik, A.; Quevedo, R.; Langille, E.; Teng, K.; Oh, R.H.; Rathod, B.; Tsai, R.; Samavarchi-Tehrani, P.; et al. Rare driver mutations in head and neck squamous cell carcinomas converge on NOTCH signaling. Science 2020, 367, 1264–1269. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. In Coronaviruses—Methods and Protocols; Humana Press: New York, NY, USA, 2015; Volume 1282, pp. 1–23. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.F.-W.; Lau, S.K.P.; To, K.; Cheng, V.C.C.; Woo, P.C.Y.; Yuen, K.-Y. Middle East Respiratory Syndrome Coronavirus: Another Zoonotic Betacoronavirus Causing SARS-Like Disease. Clin. Microbiol. Rev. 2015, 28, 465–522. [Google Scholar] [CrossRef] [Green Version]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Peng, W.; Zhu, Y.; Lu, S.; Zhou, M.; Lin, W.; Wu, W.; Huang, S.; Jiang, L.; Luo, X.; et al. Recent progress in understanding 2019 novel coronavirus (SARS-CoV-2) associated with human respiratory disease: Detection, mechanisms and treatment. Int. J. Antimicrob. Agents 2020, 55, 105950. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.; Manjunath, K.; Ranjan, R.K.; Kaushik, S.; Kumar, S.; Verma, V. COVID-19 pandemic: Insights into structure, function, and hACE2 receptor recognition by SARS-CoV-2. PLoS Pathog. 2020, 16, e1008762. [Google Scholar] [CrossRef]
- Mishra, C.B.; Pandey, P.; Sharma, R.D.; Malik, Z.; Mongre, R.K.; Lynn, A.M.; Prasad, R.; Jeon, R.; Prakash, A. Identifying the natural polyphenol catechin as a multi-targeted agent against SARS-CoV-2 for the plausible therapy of COVID-19: An integrated computational approach. Brief. Bioinform. 2020, 22, 1346–1360. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.S.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-Spectrum In Vitro Activity and In Vivo Efficacy of the Antiviral Protein Griffithsin against Emerging Viruses of the Family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- Ertekin, S.S.; Morgado-Carrasco, D.; Forns, X.; Mascaró, J.M. Complete Remission of Hypertrophic Discoid Cutaneous Lupus Erythematosus after Treatment of Chronic Hepatitis C with Direct-Acting Antivirals. JAMA Dermatol. 2020, 156, 471. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.A.; Chung, R.T. Shortening treatment with direct-acting antivirals in HCV-positive organ transplantation. Lancet Gastroenterol. Hepatol. 2020, 5, 626–627. [Google Scholar] [CrossRef]
- Choi, H.-J.; Kim, J.-H.; Lee, C.-H.; Ahn, Y.-J.; Song, J.-H.; Baek, S.-H.; Kwon, D.-H. Antiviral activity of quercetin 7-rhamnoside against porcine epidemic diarrhea virus. Antivir. Res. 2009, 81, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Nunes, V.S.; Paschoal, D.F.S.; Costa, L.A.S.; Dos Santos, H.F. Antivirals virtual screening to SARS-CoV-2 non-structural proteins. J. Biomol. Struct. Dyn. 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, Y.; Singh, H.; Patel, C.N. In silico prediction of potential inhibitors for the main protease of SARS-CoV-2 using molecular docking and dynamics simulation based drug-repurposing. J. Infect. Public Health 2020, 13, 1210–1223. [Google Scholar] [CrossRef]
- Gurung, A.B.; Ali, M.A.; Lee, J.; Farah, M.A.; Al-Anazi, K.M. Unravelling lead antiviral phytochemicals for the inhibition of SARS-CoV-2 Mpro enzyme through in silico approach. Life Sci. 2020, 255, 117831. [Google Scholar] [CrossRef] [PubMed]
- Koulgi, S.; Jani, V.; Uppuladinne, V.N.M.; Sonavane, U.; Joshi, R.N. Natural plant products as potential inhibitors of RNA dependent RNA polymerase of Severe Acute Respiratory Syndrome Coronavirus-2. PLoS ONE 2021, 16, e0251801. [Google Scholar] [CrossRef]
- Shah, B.; Modi, P.; Sagar, S.R. In silico studies on therapeutic agents for COVID-19: Drug repurposing approach. Life Sci. 2020, 252, 117652. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of flavonoids against coronavirus infection. Chem. Interact. 2020, 328, 109211. [Google Scholar] [CrossRef]
- Reynolds, D.; Huesemann, M.; Edmundson, S.; Sims, A.; Hurst, B.; Cady, S.; Beirne, N.; Freeman, J.; Berger, A.; Gao, S. Viral inhibitors derived from macroalgae, microalgae, and cyanobacteria: A review of antiviral potential throughout pathogenesis. Algal Res. 2021, 57, 102331. [Google Scholar] [CrossRef] [PubMed]
- WHO Global Report on Traditional and Complementary Medicine. 2019. Available online: https://apps.who.int/iris/handle/10665/312342 (accessed on 15 August 2021).
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Zhang, H.; Luo, H.; Zhu, L.; Jiang, P.; et al. Small Molecules Blocking the Entry of Severe Acute Respiratory Syndrome Coronavirus into Host Cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef] [Green Version]
- Chaari, A. Inhibition of human islet amyloid polypeptide aggregation and cellular toxicity by oleuropein and derivatives from olive oil. Int. J. Biol. Macromol. 2020, 162, 284–300. [Google Scholar] [CrossRef]
- Thevarajan, I.; Nguyen, T.H.O.; Koutsakos, M.; Druce, J.; Caly, L.; van de Sandt, C.E.; Jia, X.; Nicholson, S.; Catton, M.; Cowie, B.; et al. Breadth of concomitant immune responses prior to patient recovery: A case report of non-severe COVID-19. Nat. Med. 2020, 26, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising natural compounds against viral infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Fuzimoto, A.D.; Isidoro, C. The antiviral and coronavirus-host protein pathways inhibiting properties of herbs and natural compounds-Additional weapons in the fight against the COVID-19 pandemic? J. Tradit. Complement. Med. 2020, 10, 405–419. [Google Scholar] [CrossRef]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic Potential of Plant Phenolic Acids in the Treatment of Cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koklesova, L.; Liskova, A.; Samec, M.; Zhai, K.; Al-Ishaq, R.K.; Bugos, O.; Šudomová, M.; Biringer, K.; Pec, M.; Adamkov, M.; et al. Protective Effects of Flavonoids Against Mitochondriopathies and Associated Pathologies: Focus on the Predictive Approach and Personalized Prevention. Int. J. Mol. Sci. 2021, 22, 8649. [Google Scholar] [CrossRef] [PubMed]
- Al-Ishaq, R.; Liskova, A.; Kubatka, P.; Büsselberg, D. Enzymatic Metabolism of Flavonoids by Gut Microbiota and Its Impact on Gastrointestinal Cancer. Cancers 2021, 13, 3934. [Google Scholar] [CrossRef]
- Liskova, A.; Samec, M.; Koklesova, L.; Brockmueller, A.; Zhai, K.; Abdellatif, B.; Siddiqui, M.; Biringer, K.; Kudela, E.; Pec, M.; et al. Flavonoids as an effective sensitizer for anti-cancer therapy: Insights into multi-faceted mechanisms and applicability towards individualized patient profiles. EPMA J. 2021, 12, 155–176. [Google Scholar] [CrossRef]
- Chaari, A.; Abdellatif, B.; Nabi, F.; Khan, R.H. Date palm (Phoenix dactylifera L.) fruit’s polyphenols as potential inhibitors for human amylin fibril formation and toxicity in type 2 diabetes. Int. J. Biol. Macromol. 2020, 164, 1794–1808. [Google Scholar] [CrossRef] [PubMed]
- Chaari, A.; Bendriss, G.; Zakaria, D.; McVeigh, C. Importance of Dietary Changes during the Coronavirus Pandemic: How to Upgrade Your Immune Response. Front. Public Health 2020, 8, 476. [Google Scholar] [CrossRef]
- Shereen, M.A.; Khan, S.; Kazmi, A.; Bashir, N.; Siddique, R. COVID-19 infection: Emergence, transmission, and characteristics of human coronaviruses. J. Adv. Res. 2020, 24, 91–98. [Google Scholar] [CrossRef]
- Piccolella, S.; Crescente, G.; Faramarzi, S.; Formato, M.; Pecoraro, M.T.; Pacifico, S. Polyphenols vs. Coronaviruses: How Far Has Research Moved Forward? Molecules 2020, 25, 4103. [Google Scholar] [CrossRef]
- Satarker, S.; Nampoothiri, M. Structural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2. Arch. Med. Res. 2020, 51, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.K. Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Springer Nature: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Güler, G.; Özdemir, H.; Omar, D.; Akdoğan, G. Coronavirus disease 2019 (COVID-19): Biophysical and biochemical aspects of SARS-CoV-2 and general characteristics. Prog. Biophys. Mol. Biol. 2021, 164, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Genet. 2018, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Al Adem, K.; Shanti, A.; Stefanini, C.; Lee, S. Inhibition of SARS-CoV-2 Entry into Host Cells Using Small Molecules. Pharmaceuticals 2020, 13, 447. [Google Scholar] [CrossRef]
- Snijder, E.; Decroly, E.; Ziebuhr, J. The Nonstructural Proteins Directing Coronavirus RNA Synthesis and Processing. Adv. Virus Res. 2016; 96, 59–126. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, Y.; Pan, Y.; Zhao, Z.J. Structure analysis of the receptor binding of 2019-nCoV. Biochem. Biophys. Res. Commun. 2020, 525, 135–140. [Google Scholar] [CrossRef]
- Arndt, A.L.; Larson, B.J.; Hogue, B.G. A Conserved Domain in the Coronavirus Membrane Protein Tail Is Important for Virus Assembly. J. Virol. 2010, 84, 11418–11428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Levy, E.; Delvin, E.; Marcil, V.; Spahis, S. Can phytotherapy with polyphenols serve as a powerful approach for the prevention and therapy tool of novel coronavirus disease 2019 (COVID-19)? Am. J. Physiol. Metab. 2020, 319, E689–E708. [Google Scholar] [CrossRef] [PubMed]
- Belouzard, S.; Chu, V.C.; Whittaker, G.R. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc. Natl. Acad. Sci. USA 2009, 106, 5871–5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamid, S.; Mir, M.Y.; Rohela, G.K. Novel coronavirus disease (COVID-19): A pandemic (epidemiology, pathogenesis and potential therapeutics). New Microbes New Infect. 2020, 35, 100679. [Google Scholar] [CrossRef]
- Guo, Y.-R.; Cao, Q.-D.; Hong, Z.-S.; Tan, Y.-Y.; Chen, S.-D.; Jin, H.-J.; Tan, K.-S.; Wang, D.-Y.; Yan, Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak—An update on the status. Mil. Med. Res. 2020, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Grunewald, M.; Perlman, S. Coronaviruses: An Updated Overview of Their Replication and Pathogenesis. Coronaviruses 2020, 2203, 1–29. [Google Scholar] [CrossRef]
- Jeong, G.U.; Song, H.; Yoon, G.Y.; Kim, D.; Kwon, Y.-C. Therapeutic Strategies against COVID-19 and Structural Characterization of SARS-CoV-2: A Review. Front. Microbiol. 2020, 11, 1723. [Google Scholar] [CrossRef]
- Das, G.; Ghosh, S.; Garg, S.; Ghosh, S.; Jana, A.; Samat, R.; Mukherjee, N.; Roy, R. An overview of key potential therapeutic strategies for combat in the COVID-19 battle. RSC Adv. 2020, 10, 28243–28266. [Google Scholar] [CrossRef]
- Alzaabi, M.M.; Hamdy, R.; Ashmawy, N.S.; Hamoda, A.M.; Alkhayat, F.; Khademi, N.N.; Al Joud, S.M.A.; El-Keblawy, A.A.; Soliman, S.S.M. Flavonoids are promising safe therapy against COVID-19. Phytochem. Rev. 2021, 1–22. [Google Scholar] [CrossRef]
- Wang, T.-Y.; Li, Q.; Bi, K.-S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2017, 13, 12–23. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutting, W.C.; Dreisbach, R.H.; Azima, M.; Neff, B.J.; Brown, B.J.; Wray, J. Antiviral chemotherapy. V. Further report on flavonoids. Stanf. Med. Bull. 1951, 9, 236–242. [Google Scholar]
- Gábor, M.; Eperjessy, E. Antibacterial Effect of Fisetin and Fisetinidin. Nature 1966, 212, 1273. [Google Scholar] [CrossRef]
- Pusztai, R.; Béládi, I.; Bakai, M.; Mucsi, I.; Kukán, E. Study on the effect of flavonoids and related substances. I. The effect of quercetin on different viruses. Acta Microbiol. Acad. Sci. Hung. 1966, 13, 113–118. [Google Scholar]
- Kincl, F.A.; Romo, J.; Rosenkranz, G.; Sondheimer, F. 804. The constituents of Casimiroa edulis llave et lex. Part I. The seed. J. Chem. Soc. 1956, 4163–4169. [Google Scholar] [CrossRef]
- Piccolella, S.; Crescente, G.; Candela, L.; Pacifico, S. Nutraceutical polyphenols: New analytical challenges and opportunities. J. Pharm. Biomed. Anal. 2019, 175, 112774. [Google Scholar] [CrossRef] [PubMed]
- Piccolella, S.; Pacifico, S. Plant-Derived Polyphenols: A Chemopreventive and Chemoprotectant Worth-Exploring Resource in Toxicology. Adv. Mol. Toxicol. 2015, 9, 161–214. [Google Scholar] [CrossRef]
- Martin, K.W.; Ernst, E. Antiviral agents from plants and herbs: A systematic review. Antivir. Ther. 2003, 8, 77–90. [Google Scholar] [CrossRef]
- Helenius, A. Virus Entry: Looking Back and Moving Forward. J. Mol. Biol. 2018, 430, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Denaro, M.; Smeriglio, A.; Barreca, D.; De Francesco, C.; Occhiuto, C.; Milano, G.; Trombetta, D. Antiviral activity of plants and their isolated bioactive compounds: An update. Phytotherapy Res. 2019, 34, 742–768. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Kaleem, M.; Ahmed, Z.; Shafiq, H. Therapeutic potential of flavonoids and their mechanism of action against microbial and viral infections—A review. Food Res. Int. 2015, 77, 221–235. [Google Scholar] [CrossRef]
- Béládi, I.; Pusztai, R.; Mucsi, I.; Bakay, M.; Gabor, M. Activity of some flavonoids against viruses. Ann. N. Y. Acad. Sci. 1977, 284, 358–364. [Google Scholar] [CrossRef]
- Kaul, T.N.; Middleton, E.; Ogra, P.L. Antiviral effect of flavonoids on human viruses. J. Med. Virol. 1985, 15, 71–79. [Google Scholar] [CrossRef]
- Vlietinck, A.J.; Berghe, D.A.V. Can ethnopharmacology contribute to the development of antiviral drugs? J. Ethnopharmacol. 1991, 32, 141–153. [Google Scholar] [CrossRef]
- Debiaggi, M.; Tateo, F.; Pagani, L.; Luini, M.; Romero, E. Effects of propolis flavonoids on virus infectivity and replication. Microbiologica 1990, 13, 207–213. [Google Scholar]
- Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, Z.; Du, J.; Hu, Y.; Liu, L.; Yang, F.; Jin, Q. Anti- Japanese-Encephalitis-Viral Effects of Kaempferol and Daidzin and Their RNA-Binding Characteristics. PLoS ONE 2012, 7, e30259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lani, R.; Hassandarvish, P.; Shu, M.-H.; Phoon, W.H.; Chu, J.J.H.; Higgs, S.; Vanlandingham, D.; Abu Bakar, S.; Zandi, K. Antiviral activity of selected flavonoids against Chikungunya virus. Antivir. Res. 2016, 133, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-C.; Chen, M.-C.; Li, S.; Lin, C.-C.; Wang, T.T. Antiviral activity of nobiletin against chikungunya virus in vitro. Antivir. Ther. 2017, 22, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Lani, R.; Hassandarvish, P.; Chiam, C.W.; Moghaddam, E.; Chu, J.J.H.; Rausalu, K.; Merits, A.; Higgs, S.; VanLandingham, D.L.; Abu Bakar, S.; et al. Antiviral activity of silymarin against chikungunya virus. Sci. Rep. 2015, 5, 11421. [Google Scholar] [CrossRef] [Green Version]
- Zandi, K.; Teoh, B.-T.; Sam, S.-S.; Wong, P.-F.; Mustafa, M.R.; AbuBakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 2011, 8, 560. [Google Scholar] [CrossRef] [Green Version]
- Chiow, K.; Phoon, M.; Putti, T.; Tan, B.K.; Chow, V.T. Evaluation of antiviral activities of Houttuynia cordata Thunb. extract, quercetin, quercetrin and cinanserin on murine coronavirus and dengue virus infection. Asian Pac. J. Trop. Med. 2015, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zandi, K.; Teoh, B.-T.; Sam, S.-S.; Wong, P.-F.; Mustafa, M.R.; AbuBakar, S. Novel antiviral activity of baicalein against dengue virus. BMC Complement. Altern. Med. 2012, 12, 214. [Google Scholar] [CrossRef] [Green Version]
- Moghaddam, E.J.; Teoh, B.-T.; Sam, S.-S.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a metabolite of baicalein with antiviral activity against dengue virus. Sci. Rep. 2014, 4, 5452. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Qiao, H.; Lv, Y.; Wang, J.; Chen, X.; Hou, Y.; Tan, R.; Li, E. Apigenin Inhibits Enterovirus-71 Infection by Disrupting Viral RNA Association with trans-Acting Factors. PLoS ONE 2014, 9, e110429. [Google Scholar] [CrossRef]
- Dai, W.; Bi, J.; Li, F.; Wang, S.; Huang, X.; Meng, X.; Sun, B.; Wang, D.; Kong, W.; Jiang, C.; et al. Antiviral Efficacy of Flavonoids against Enterovirus 71 Infection in Vitro and in Newborn Mice. Viruses 2019, 11, 625. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, Y.; Wu, T.; Jin, Y.; Cheng, J.; Wan, C.; Qian, W.; Xing, F.; Shi, W. The Antiviral Effect of Baicalin on Enterovirus 71 In Vitro. Viruses 2015, 7, 4756–4771. [Google Scholar] [CrossRef] [Green Version]
- Tsai, F.-J.; Lin, C.-W.; Lai, C.-C.; Lan, Y.-C.; Lai, C.-H.; Hung, C.-H.; Hsueh, K.-C.; Lin, T.-H.; Chang, H.C.; Wan, L.; et al. Kaempferol inhibits enterovirus 71 replication and internal ribosome entry site (IRES) activity through FUBP and HNRP proteins. Food Chem. 2011, 128, 312–322. [Google Scholar] [CrossRef]
- Nahmias, Y.; Goldwasser, J.; Casali, M.; van Poll, D.; Wakita, T.; Chung, R.T.; Yarmush, M.L. Apolipoprotein B-dependent hepatitis C virus secretion is inhibited by the grapefruit flavonoid naringenin. Hepatology 2008, 47, 1437–1445. [Google Scholar] [CrossRef] [Green Version]
- Shibata, C.; Ohno, M.; Otsuka, M.; Kishikawa, T.; Goto, K.; Muroyama, R.; Kato, N.; Yoshikawa, T.; Takata, A.; Koike, K. The flavonoid apigenin inhibits hepatitis C virus replication by decreasing mature microRNA122 levels. Virology 2014, 462–463, 42–48. [Google Scholar] [CrossRef]
- Ferenci, P.; Scherzer, T.; Kerschner, H.; Rutter, K.; Beinhardt, S.; Hofer, H.; Schöniger–Hekele, M.; Holzmann, H.; Steindl–Munda, P. Silibinin Is a Potent Antiviral Agent in Patients With Chronic Hepatitis C Not Responding to Pegylated Interferon/Ribavirin Therapy. Gastroenterology 2008, 135, 1561–1567. [Google Scholar] [CrossRef]
- Bachmetov, L.; Gal-Tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman-Galam, T.; Sathiyamoorthy, P.; Golan-Goldhirsh, A.; Benhar, I.; Tur-Kaspa, R.; Zemel, R. Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. J. Viral Hepat. 2011, 19, e81–e88. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, J.F.; Lima, C.S.; Pereira, C.M.; Bittar, C.; Batista, M.; Nazaré, A.C.; Polaquini, C.R.; Zothner, C.; Harris, M.; Rahal, P.; et al. Flavonoids from Pterogyne nitens Inhibit Hepatitis C Virus Entry. Sci. Rep. 2017, 7, 16127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauter, D.; Schwarz, S.; Wang, K.; Zhang, R.; Sun, B.; Schwarz, W. Genistein as Antiviral Drug against HIV Ion Channel. Planta Med. 2014, 80, 682–687. [Google Scholar] [CrossRef]
- Tao, J.; Hu, Q.; Yang, J.; Li, R.; Li, X.; Lu, C.; Chen, C.; Wang, L.; Shattock, R.; Ben, K. In vitro anti-HIV and -HSV activity and safety of sodium rutin sulfate as a microbicide candidate. Antivir. Res. 2007, 75, 227–233. [Google Scholar] [CrossRef]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral activities of flavonoids. Biomed. Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef]
- Ge, M.; Xiao, Y.; Chen, H.; Luo, F.; Du, G.; Zeng, F. Multiple antiviral approaches of (–)-epigallocatechin-3-gallate (EGCG) against porcine reproductive and respiratory syndrome virus infection in vitro. Antivir. Res. 2018, 158, 52–62. [Google Scholar] [CrossRef]
- Mehany, T.; Khalifa, I.; Barakat, H.; Althwab, S.A.; Alharbi, Y.M.; El-Sohaimy, S. Polyphenols as promising biologically active substances for preventing SARS-CoV-2: A review with research evidence and underlying mechanisms. Food Biosci. 2021, 40, 100891. [Google Scholar] [CrossRef]
- Imanishi, N.; Tuji, Y.; Katada, Y.; Maruhashi, M.; Konosu, S.; Mantani, N.; Terasawa, K.; Ochiai, H. Additional Inhibitory Effect of Tea Extract on the Growth of Influenza A and B Viruses in MDCK Cells. Microbiol. Immunol. 2002, 46, 491–494. [Google Scholar] [CrossRef]
- Roschek, B.; Fink, R.; McMichael, M.D.; Li, D.; Alberte, R.S. Elderberry flavonoids bind to and prevent H1N1 infection in vitro. Phytochemistry 2009, 70, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Choi, H. Silymarin efficacy against influenza A virus replication. Phytomedicine 2011, 18, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Omrani, M.; Keshavarz, M.; Ebrahimi, S.N.; Mehrabi, M.; McGaw, L.J.; Abdalla, M.A.; Mehrbod, P. Potential Natural Products Against Respiratory Viruses: A Perspective to Develop Anti-COVID-19 Medicines. Front. Pharmacol. 2021, 11, 586993. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Faris, A.N.; Comstock, A.T.; Wang, Q.; Nanua, S.; Hershenson, M.B.; Sajjan, U.S. Quercetin inhibits rhinovirus replication in vitro and in vivo. Antivir. Res. 2012, 94, 258–271. [Google Scholar] [CrossRef]
- Desideri, N.; Conti, C.; Sestili, I.; Tomao, P.; Stein, M.L.; Orsi, N. In vitro Evaluation of the Anti-Picornavirus Activities of New Synthetic Flavonoids. Antivir. Chem. Chemother. 1995, 6, 298–306. [Google Scholar] [CrossRef]
- Clark, K.; Grant, P.; Sarr, A.; Belakere, J.; Swaggerty, C.; Phillips, T.; Woode, G. An in vitro study of theaflavins extracted from black tea to neutralize bovine rotavirus and bovine coronavirus infections. Vet.-Microbiol. 1998, 63, 147–157. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.-Y.; Kim, D.; Naguyen, T.T.H.; Park, S.-J.; Chang, J.S.; Park, K.H. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorganic Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.-S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzym. Inhib. Med. Chem. 2019, 35, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.H.; Woo, H.-J.; Kang, H.-K.; Nguyen, V.D.; Kim, Y.-M.; Kim, D.-W.; Ahn, S.-A.; Xia, Y.; Kim, D. Flavonoid-mediated inhibition of SARS coronavirus 3C-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.K.; Curtis-Long, M.J.; Lee, K.H.; Kim, D.W.; Ryu, H.W.; Yuk, H.J.; Park, K.H. Geranylated flavonoids displaying SARS-CoV papain-like protease inhibition from the fruits of Paulownia tomentosa. Bioorganic Med. Chem. 2013, 21, 3051–3057. [Google Scholar] [CrossRef]
- Schwarz, S.; Sauter, D.; Wang, K.; Zhang, R.; Sun, B.; Karioti, A.; Bilia, A.R.; Efferth, T.; Schwarz, W. Kaempferol Derivatives as Antiviral Drugs against the 3a Channel Protein of Coronavirus. Planta Med. 2014, 80, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.-S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.-W.; Jee, J.-G.; Keum, Y.-S.; Jeong, Y.-J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorganic Med. Chem. Lett. 2012, 22, 4049–4054. [Google Scholar] [CrossRef]
- Ho, T.-Y.; Wu, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef]
- Chen, S.; Chan, K.; Jiang, Y.; Kao, R.Y.; Lu, H.; Fan, K.; Cheng, V.; Tsui, W.; Hung, I.F.N.; Lee, T. In vitro susceptibility of 10 clinical isolates of SARS coronavirus to selected antiviral compounds. J. Clin. Virol. 2004, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, H.; Kim, S.; Shin, D.H.; Kim, M. Characteristics of flavonoids as potent MERS-CoV 3C-like protease inhibitors. Chem. Biol. Drug Des. 2019, 94, 2023–2030. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, M.; Jiang, H.; Suzuki, Y.; Li, X.; Xiao, P.; Tanaka, T.; Ling, H.; Yang, B.; Saitoh, H.; Zhang, L.; et al. Procyanidins and butanol extract of Cinnamomi Cortex inhibit SARS-CoV infection. Antivir. Res. 2009, 82, 73–81. [Google Scholar] [CrossRef]
- Roh, C. A facile inhibitor screening of SARS coronavirus N protein using nanoparticle-based RNA oligonucleotide. Int. J. Nanomed. 2012, 7, 2173–2179. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Lee, J.M.; Lee, N.-R.; Kim, D.-E.; Jeong, Y.-J.; Chong, Y. Investigation of the pharmacophore space of Severe Acute Respiratory Syndrome coronavirus (SARS-CoV) NTPase/helicase by dihydroxychromone derivatives. Bioorg. Med. Chem. Lett. 2009, 19, 4538–4541. [Google Scholar] [CrossRef]
- Solnier, J.; Fladerer, J.-P. Flavonoids: A complementary approach to conventional therapy of COVID-19? Phytochem. Rev. 2020, 20, 773–795. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Yao, S.; Zhao, W.; Zhang, Y.; Liu, J.; Shao, Q.; Wang, Q.; Li, M.; Xie, H.; Shang, W.; et al. Identification of pyrogallol as a warhead in design of covalent inhibitors for the SARS-CoV-2 3CL protease. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Jung, J.-H.; Kim, M.-K.; Lim, S.; Choi, J.-M.; Chung, B.; Kim, D.-W.; Kim, D. The Inhibitory Effects of Plant Derivate Polyphenols on the Main Protease of SARS Coronavirus 2 and Their Structure–Activity Relationship. Molecules 2021, 26, 1924. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ye, F.; Sun, Q.; Liang, H.; Li, C.; Li, S.; Lu, R.; Huang, B.; Tan, W.; Lai, L. Scutellaria baicalensis extract and baicalein inhibit replication of SARS-CoV-2 and its 3C-like protease in vitro. J. Enzym. Inhib. Med. Chem. 2021, 36, 497–503. [Google Scholar] [CrossRef]
- Gao, J.; Ding, Y.; Wang, Y.; Liang, P.; Zhang, L.; Liu, R. Oroxylin A is a severe acute respiratory syndrome coronavirus 2-spiked pseudotyped virus blocker obtained from Radix Scutellariae using angiotensin-converting enzyme II/cell membrane chromatography. Phytotherapy Res. 2021, 35, 3194–3204. [Google Scholar] [CrossRef] [PubMed]
- Du, A.; Zheng, R.; Disoma, C.; Li, S.; Chen, Z.; Li, S.; Liu, P.; Zhou, Y.; Shen, Y.; Liu, S.; et al. Epigallocatechin-3-gallate, an active ingredient of Traditional Chinese Medicines, inhibits the 3CLpro activity of SARS-CoV-2. Int. J. Biol. Macromol. 2021, 176, 1–12. [Google Scholar] [CrossRef]
- Xiao, T.; Cui, M.; Zheng, C.; Wang, M.; Sun, R.; Gao, D.; Bao, J.; Ren, S.; Yang, B.; Lin, J.; et al. Myricetin Inhibits SARS-CoV-2 Viral Replication by Targeting Mpro and Ameliorates Pulmonary Inflammation. Front. Pharmacol. 2021, 12, 669642. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhu, G.-H.; Zhang, Y.-N.; Hu, Q.; Wang, H.-N.; Yu, H.-N.; Qin, X.-Y.; Guan, X.-Q.; Xiang, Y.-W.; Tang, H.; et al. Flavonoids in Ampelopsis grossedentata as covalent inhibitors of SARS-CoV-2 3CLpro: Inhibition potentials, covalent binding sites and inhibitory mechanisms. Int. J. Biol. Macromol. 2021, 187, 976–987. [Google Scholar] [CrossRef]
- Abian, O.; Ortega-Alarcon, D.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Vega, S.; Reyburn, H.T.; Rizzuti, B.; Velazquez-Campoy, A. Structural stability of SARS-CoV-2 3CLpro and identification of quercetin as an inhibitor by experimental screening. Int. J. Biol. Macromol. 2020, 164, 1693–1703. [Google Scholar] [CrossRef]
- Rizzuti, B.; Grande, F.; Conforti, F.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Ortega-Alarcon, D.; Vega, S.; Reyburn, H.; Abian, O.; Velazquez-Campoy, A. Rutin Is a Low Micromolar Inhibitor of SARS-CoV-2 Main Protease 3CLpro: Implications for Drug Design of Quercetin Analogs. Biomedicines 2021, 9, 375. [Google Scholar] [CrossRef]
- Su, H.-X.; Yao, S.; Zhao, W.-F.; Li, M.-J.; Liu, J.; Shang, W.-J.; Xie, H.; Ke, C.-Q.; Hu, H.-C.; Gao, M.-N.; et al. Anti-SARS-CoV-2 activities in vitro of Shuanghuanglian preparations and bioactive ingredients. Acta Pharmacol. Sin. 2020, 41, 1167–1177. [Google Scholar] [CrossRef]
- Owis, A.I.; El-Hawary, M.S.; El Amir, D.; Refaat, H.; Alaaeldin, E.; Aly, O.M.; Elrehany, M.A.; Kamel, M.S. Flavonoids of Salvadora persica L. (meswak) and its liposomal formulation as a potential inhibitor of SARS-CoV-2. RSC Adv. 2021, 11, 13537–13544. [Google Scholar] [CrossRef]
- Pitsillou, E.; Liang, J.; Ververis, K.; Hung, A.; Karagiannis, T.C. Interaction of small molecules with the SARS-CoV-2 papain-like protease: In silico studies and in vitro validation of protease activity inhibition using an enzymatic inhibition assay. J. Mol. Graph. Model. 2021, 104, 107851. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Raghuvanshi, R.; Ceylan, F.D.; Bolling, B.W. Quercetin and Its Metabolites Inhibit Recombinant Human Angiotensin-Converting Enzyme 2 (ACE2) Activity. J. Agric. Food Chem. 2020, 68, 13982–13989. [Google Scholar] [CrossRef]
- Güler, H.I.; Şal, F.A.; Can, Z.; Kara, Y.; Yildiz, O.; Beldüz, A.O.; Çanakçi, S.; Kolayli, S. Targeting CoV-2 spike RBD and ACE-2 interaction with flavonoids of Anatolian propolis by in silico and in vitro studies in terms of possible COVID-19 therapeutics. Turk. J. Boil. 2021, 45, 530–548. [Google Scholar] [CrossRef]
- Henss, L.; Auste, A.; Schürmann, C.; Schmidt, C.; von Rhein, C.; Mühlebach, M.D.; Schnierle, B.S. The green tea catechin epigallocatechin gallate inhibits SARS-CoV-2 infection. J. Gen. Virol. 2021, 102, 001574. [Google Scholar] [CrossRef] [PubMed]
- Biagioli, M.; Marchianò, S.; Roselli, R.; Di Giorgio, C.; Bellini, R.; Bordoni, M.; Gidari, A.; Sabbatini, S.; Francisci, D.; Fiorillo, B.; et al. Discovery of a AHR pelargonidin agonist that counter-regulates Ace2 expression and attenuates ACE2-SARS-CoV-2 interaction. Biochem. Pharmacol. 2021, 188, 114564. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Ta, W.; Tang, W.; Hua, R.; Wang, J.; Wang, C.; Lu, W. Potential antiviral activity of isorhamnetin against SARS-CoV -2 spike pseudotyped virus in vitro. Drug Dev. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Musall, K.; Oo, A.; Cao, D.; Liang, B.; Hassandarvish, P.; Lan, S.; Slack, R.; Kirby, K.; Bassit, L.; et al. Baicalein and Baicalin Inhibit SARS-CoV-2 RNA-Dependent-RNA Polymerase. Microorganisms 2021, 9, 893. [Google Scholar] [CrossRef]
- Huang, S.; Liu, Y.; Zhang, Y.; Zhang, R.; Zhu, C.; Fan, L.; Pei, G.; Zhang, B.; Shi, Y. Baicalein inhibits SARS-CoV-2/VSV replication with interfering mitochondrial oxidative phosphorylation in a mPTP dependent manner. Signal Transduct. Target. Ther. 2020, 5, 1–3. [Google Scholar] [CrossRef]
- Hong, S.; Seo, S.H.; Woo, S.-J.; Kwon, Y.; Song, M.; Ha, N.-C. Epigallocatechin Gallate Inhibits the Uridylate-Specific Endoribonuclease Nsp15 and Efficiently Neutralizes the SARS-CoV-2 Strain. J. Agric. Food Chem. 2021, 69, 5948–5954. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Park, R.; Park, Y.-I.; Cha, Y.-E.; Yamamoto, A.; Lee, J.I.; Park, J. EGCG, a green tea polyphenol, inhibits human coronavirus replication in vitro. Biochem. Biophys. Res. Commun. 2021, 547, 23–28. [Google Scholar] [CrossRef]
- Clementi, N.; Scagnolari, C.; D’Amore, A.; Palombi, F.; Criscuolo, E.; Frasca, F.; Pierangeli, A.; Mancini, N.; Antonelli, G.; Clementi, M.; et al. Naringenin is a powerful inhibitor of SARS-CoV-2 infection in vitro. Pharmacol. Res. 2020, 163, 105255. [Google Scholar] [CrossRef] [PubMed]
- Pitsillou, E.; Liang, J.; Ververis, K.; Lim, K.W.; Hung, A.; Karagiannis, T.C. Identification of Small Molecule Inhibitors of the Deubiquitinating Activity of the SARS-CoV-2 Papain-Like Protease: In silico Molecular Docking Studies and in vitro Enzymatic Activity Assay. Front. Chem. 2020, 8, 623971. [Google Scholar] [CrossRef]
- Leal, C.M.; Leitão, S.G.; Sausset, R.; Mendonça, S.C.; Nascimento, P.H.A.; Cheohen, C.F.d.A.R.; Esteves, M.E.A.; da Silva, M.L.; Gondim, T.S.; Monteiro, M.E.S.; et al. Flavonoids from Siparuna cristata as Potential Inhibitors of SARS-CoV-2 Replication. Rev. Bras. Farm. 2021, 1–9. [Google Scholar] [CrossRef]
- Song, J.; Zhang, L.; Xu, Y.; Yang, D.; Yang, S.; Zhang, W.; Wang, J.; Tian, S.; Yang, S.; Yuan, T.; et al. The comprehensive study on the therapeutic effects of baicalein for the treatment of COVID-19 in vivo and in vitro. Biochem. Pharmacol. 2020, 183, 114302. [Google Scholar] [CrossRef] [PubMed]
- Cherrak, S.A.; Merzouk, H.; Mokhtari-Soulimane, N. Potential bioactive glycosylated flavonoids as SARS-CoV-2 main protease inhibitors: A molecular docking and simulation studies. PLoS ONE 2020, 15, e0240653. [Google Scholar] [CrossRef]
- Teli, D.M.; Shah, M.B.; Chhabria, M.T. In silico Screening of Natural Compounds as Potential Inhibitors of SARS-CoV-2 Main Protease and Spike RBD: Targets for COVID-19. Front. Mol. Biosci. 2021, 7, 599079. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Dubey, A.; Yadava, U.; Mishra, S.K.; Kang, S.G.; Dwivedi, V.D. Exploration of natural compounds with anti-SARS-CoV-2 activity via inhibition of SARS-CoV-2 Mpro. Brief. Bioinform. 2021, 22, 1361–1377. [Google Scholar] [CrossRef] [PubMed]
- Rakshit, G.; Dagur, P.; Satpathy, S.; Patra, A.; Jain, A.; Ghosh, M. Flavonoids as potential therapeutics against novel coronavirus disease-2019 (nCOVID-19). J. Biomol. Struct. Dyn. 2021, 1–13. [Google Scholar] [CrossRef]
- Yu, R.; Chen, L.; Lan, R.; Shen, R.; Li, P. Computational screening of antagonists against the SARS-CoV-2 (COVID-19) coronavirus by molecular docking. Int. J. Antimicrob. Agents 2020, 56, 106012. [Google Scholar] [CrossRef] [PubMed]
- Akhter, S.; Batool, A.I.; Selamoglu, Z.; Sevindik, M.; Eman, R.; Mustaqeem, M.; Aslam, M. Effectiveness of Natural Antioxidants against SARS-CoV-2? Insights from the In-Silico World. Antibiotics 2021, 10, 1011. [Google Scholar] [CrossRef]
- Batool, F.; Mughal, E.U.; Zia, K.; Sadiq, A.; Naeem, N.; Javid, A.; Ul-Haq, Z.; Saeed, M. Synthetic flavonoids as potential antiviral agents against SARS-CoV-2 main protease. J. Biomol. Struct. Dyn. 2020, 1–12. [Google Scholar] [CrossRef]
- Ziebuhr, J.; Gorbalenya, A.; Snijder, E. Virus-encoded proteinases and proteolytic processing in the Nidovirales. J. Gen. Virol. 2000, 81, 853–879. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.-S.; Wang, S.-Q.; Zhu, Y.; Wei, D.-Q.; Guo, H.; Sirois, S.; Chou, K.-C. Polyprotein cleavage mechanism of SARS-CoV Mpro and chemical modification of the octapeptide. Peptides 2004, 25, 1857–1864. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Hilgenfeld, R. Conservation of substrate specificities among coronavirus main proteases. J. Gen. Virol. 2002, 83, 595–599. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilgenfeld, R. From SARS to MERS: Crystallographic studies on coronaviral proteases enable antiviral drug design. FEBS J. 2014, 281, 4085–4096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Lin, D.; Kusov, Y.; Nian, Y.; Ma, Q.; Wang, J.; Von Brunn, A.; Leyssen, P.; Lanko, K.; Neyts, J.; et al. α-Ketoamides as Broad-Spectrum Inhibitors of Coronavirus and Enterovirus Replication: Structure-Based Design, Synthesis, and Activity Assessment. J. Med. Chem. 2020, 63, 4562–4578. [Google Scholar] [CrossRef] [PubMed]
- Qamar, M.T.U.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.-L. Structural basis of SARS-CoV-2 3CLpro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 10, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.S.; Sushma, P.; Dharmashekar, C.; Beelagi, M.S.; Prasad, S.K.; Shivamallu, C.; Prasad, A.; Syed, A.; Marraiki, N.; Prasad, K.S. In silico evaluation of flavonoids as effective antiviral agents on the spike glycoprotein of SARS-CoV-2. Saudi J. Biol. Sci. 2020, 28, 1040–1051. [Google Scholar] [CrossRef]
- Lung, J.; Lin, Y.; Yang, Y.; Chou, Y.; Shu, L.; Cheng, Y.; Liu, H.T.; Wu, C. The potential chemical structure of anti- SARS-CoV -2 RNA-dependent RNA polymerase. J. Med. Virol. 2020, 92, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Shawan, M.M.A.K.; Halder, S.K.; Hasan, A. Luteolin and abyssinone II as potential inhibitors of SARS-CoV-2: An in silico molecular modeling approach in battling the COVID-19 outbreak. Bull. Natl. Res. Cent. 2021, 45, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.S.; Jagdale, S.S.; Bansode, S.B.; Shankar, S.S.; Tellis, M.B.; Pandya, V.K.; Chugh, A.; Giri, A.P.; Kulkarni, M.J. Discovery of potential multi-target-directed ligands by targeting host-specific SARS-CoV-2 structurally conserved main protease. J. Biomol. Struct. Dyn. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Du, Q. Potential Natural Compounds for Preventing SARS-CoV-2 (2019-nCoV) Infection. Available online: https://www.preprints.org/manuscript/202001.0358/v3 (accessed on 3 June 2021).
- Utomo, R.Y.; Ikawati, M.; Meiyanto, E. Revealing the Potency of Citrus and Galangal Con-stituents to Halt SARS-CoV-2 In-fection. Preprints 2020, 2020030214. [Google Scholar] [CrossRef] [Green Version]
- Topcagic, A.; Zeljkovic, S.C.; Karalija, E.; Galijasevic, S.; Sofic, E. Evaluation of phenolic profile, enzyme inhibitory and antimicrobial activities of Nigella sativa L. seed extracts. Bosn. J. Basic Med. Sci. 2017, 17, 286–294. [Google Scholar] [CrossRef] [Green Version]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Class Flavonoid | Natural Source (N.S.)/Extract | Efficacy of N.S. Extract | Mode of Action | Methods Used | IC50 (µM) | EC50 (µM) | % Inhibition | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Flavonol | 7-O-methyl-myricetin | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 0.30 ± 0.00 | 12.59 ± 4.41 | [117] | |||
| 7-O-ethyl-myricetin | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 0.74 ± 0.06 | 51.01 ± 12.79 | [117] | ||||
| 7-O-isoamyl-myricetin | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 1.92 ± 0.16 | 31.54 ± 0.74 | [117] | ||||
| 7-O-cyclopentylmethyl-myricetin | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 2.45 ± 0.26 | 7.56 ± 2.34 | [117] | ||||
| Astragalin (kaempferol 3-glucoside) | Black garlic extract | IC50: 137 ± 10 µg/mL, 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 143 ± 9 | 61 at 200 µM | [118] | ||
| Herbacetin (8-hydroxykaempferol) | Flaxseed hulls, Rhodiola | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 59.1 ± 1.9 at 50 µM | [119] | ||||
| Hyperoside (quercetin 3-galactoside) | Nelumbo nucifera | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 5.2 at 10 µM | [117] | ||||
| Icaritin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 31 at 200 µM | [118] | |||
| Isorhamnetin (3-methylquercetin) | Pears, olive oil, wine | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | −2.6 at 10 µM | [117] | ||||
| Kaempferol | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 16 at 200 µM | [118] | |||
| TCM | Binds to 3CLpro active site | Vero E6 Cells | 34.46 | [120] | |||||
| Kaempferide | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 8.1 at 10 µM | [117] | |||||
| TCM | Binds to the 3CLpro substrate binding site. | FRET assay | >100 | [121] | |||||
| Myricetin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 43 ± 1 | 80 at 200 µM | [118] | ||
| Polygoni avicularis | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 2.86 ± 0.23 | [119] | |||||
| Positions itself in the 3CLpro binding pocket | FRET assay, BEAS-2B cells | 3.684 ± 0.076 | 97.79 at 50 µM | [122] | |||||
| Binds at the catalytic site within the extended substrate-binding pocket | FRET assay, Vero E6 Cells, qRT-PCR | 0.63 ± 0.01 | 8.00 ± 2.05 | 97.6 at 10 µM | [117] | ||||
| Ampelopsis grossedentata extract | 99.74% inhibition at 100 μg/mL IC50 = 3.44 μg/mL | Modify key residue in domain III of 3CLpro | FRET assay | 1.21 (60 min pre-incubation) 21.44 (0.5 min pre-incubation) | [123] | ||||
| Myricetin-7-yl 5,5-dimethyl-1,3,2-dioxayl phosphate | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 6.62 ± 0.42 | 33.45 ± 11.96 | [117] | ||||
| Myricetin-7-yl diphenyl phosphate | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 3.13 ± 0.37 | 3.15 ± 0.84 | [117] | ||||
| Quercetagenin/quercetagenin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 145 ± 6 | 58 at 200 µM | [118] | ||
| Eriocaulon buergerianum | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 1.24 ± 0.14 | [119] | |||||
| Quercetin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 93 ± 5 | 74 at 200 µM | [118] | ||
| TCM | Binds to the 3CLpro substrate binding site. | FRET assay | 97.460 ± 2.263 | [121] | |||||
| Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 41.3 at 10 µM | [117] | ||||||
| Binds to SARS-CoV-2 3CLpro active site | FRET assay | Kiapp = 21 μM Ki = 7.4 μM Kd = 2.7 μM (no NaCl) Kd = 150 mM (150 mM NaCl) | [124] | ||||||
| Quercetin-4′-O-α-D-glucopyranoside | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 26 at 200 µM | [118] | |||
| Rutin (quercetin-3-O-rutinose) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 45 at 200 µM | [118] | |||
| Binds to the 3CLpro catalytic site. | FRET assay | 32 | 43 at 30 µM 65 at 60 µM 80 at 120 µM | [125] | |||||
| Flavanonol | Ampelopsin (dihydromyricetin/DHM) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 128 ± 5 | 64 at 200 µM | [118] | |
| Ampelopsis japonica | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 1.20 ± 0.09 | [119] | |||||
| Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 1.14 ± 0.03 | 13.56 ± 2.50 | 93.8 at 10 µM | [117] | ||||
| Ampelopsis grossedentata extract | 99.74% inhibition at 100 μg/mL IC50 = 3.44 μg/mL | Modify key residue in domain III of 3CLpro | FRET assay | 4.91 (60 min pre-incubation) 34.61 (0.5 min pre-incubation) | [123] | ||||
| Isodihydromyricetin | Ampelopsis grossedentata extract | 99.74% inhibition at 100 μg/mL IC50 = 3.44 μg/mL | Modify key residue in domain III of 3CLpro | FRET assay | 3.73 (60 min pre-incubation) 29.04 (0.5 min pre-incubation) | [123] | |||
| Dihydromyricetin-7-yl diphenyl phosphate | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 1.84 ± 0.22 | 9.03 ± 1.36 | [117] | ||||
| 7-O-methyl-dihydromyricetin | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 0.26 ± 0.02 | 11.50 ± 4.57 | [117] | ||||
| Ampelopsin-4′-O-α-D-glucopyranoside | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 195 ± 5 | 50 at 200 µM | [118] | ||
| Taxifolin | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 28.0 at 10 µM | [117] | |||||
| Ampelopsis grossedentata extract | 99.74% inhibition at 100 μg/mL IC50 = 3.44 μg/mL | Inhibits 3CLpro activity | FRET assay | 72.72 (60 min pre-incubation) | [123] | ||||
| Flavanone | (±)-Eriodyctiol | Interacts with 3CLpro catalytic site | FRET assay, Vero E6 Cells, qRT-PCR | 34.5 at 10 µM | [117] | ||||
| Hesperidin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 22 at 200 µM | [118] | |||
| Hesperetin | Interacts with 3CLpro catalytic site | FRET assay | 13.8 at 10 µM | [117] | |||||
| Naringenin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 150 ± 10 | 57 at 200 µM | [118] | ||
| TCM | Binds to the 3CLpro substrate binding site. | FRET assay | >1000 | [121] | |||||
| Naringin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 18 at 200 µM | [118] | |||
| Flavones | 5,6-dihydroxyflavone | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 26.6 ± 0.4 at 50 µM | [119] | ||||
| 6,7-dihydroxyflavone | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 56.7 ± 2.0 at 50 µM | [119] | |||||
| Apigenin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 25 at 200 µM | [118] | |||
| Interacts with 3CLpro catalytic site | FRET assay | −1.0 at 10 µM | [117] | ||||||
| Baicalein | Scutellaria baicalensis | IC50: 8.52 ± 0.54 µg/mL EC50: 0.74 ± 0.36 µg/mL CC50: > 500 µg/mL | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 0.39 ± 0.12 | 2.92 ± 0.06 | [119] | ||
| Binds to the core of the substrate-binding pocket, preventing substrate access to the active site | Vero E6 cells/CCK8 assays/qRT-PCR | 0.94 ± 0.20 | 2.49 ± 1.19 | 99.4 at 100 µM 87 at 10 µM | [126] | ||||
| Baicalin | Scutellaria baicalensis | IC50: 8.52 ± 0.54 µg/mL EC50: 0.74 ± 0.36 µg/mL CC50: > 500 µg/mL | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 83.4 ± 0.9 | 41.5 ± 0.6 at 50 µM | [119] | ||
| Binds to 3CLpro active site | Vero E6 cells/CCK8 assays/qRT-PCR | 6.41 ± 0.95 | 27.87 ± 12.5 | 97.6 at 100 µM 68.9 at 10 µM | [126] | ||||
| Chrysin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 9 at 200 µM | [118] | |||
| Binds to the 3CLpro substrate binding site. | Vero cells/RT PCR | 2.6 ± 1.1 at 50 µM | [119] | ||||||
| Chrysin-7-O-β-D-glucoronide | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 50.6 at 100 μM 24.2 at 10 μM | [126] | ||||
| Diosmetin | Interacts with 3CLpro catalytic site | FRET assay | 11.3 at 10 µM | [117] | |||||
| Luteolin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 45 at 200 µM | [118] | |||
| TCM | Binds to the 3CLpro substrate binding site. | FRET assay | 89.670 ± 4.712 | [121] | |||||
| Interacts with 3CLpro catalytic site | FRET assay | −4.1 at 10 µM | [117] | ||||||
| Luteoloside (cyranoside) | L. japonica | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 65.4 at 100 μM 14.8 at 10 μM | [126] | ||||
| Myricitrin | Polygoni avicularis | Binds to the 3CLpro substrate binding site | Vero cells/RT PCR | 30.8 ± 4.6 at 50 µM | [119] | ||||
| Ampelopsis grossedentata extract | 99.74% inhibition at 100 μg/mL IC50 = 3.44 μg/mL | Modify key residue in domain III of 3CLpro | FRET assay | 14.22 (60 min pre-incubation) | [123] | ||||
| Oroxylin A-7-O-β-D-glucuronide | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 33.0 at 100 μM | [126] | ||||
| Scutellarein (6-hydroxyapigenin) | Scutellaria, Erigerontis herba | Binds to the 3CLpro substrate binding site. | Vero cells/RT PCR | 5.80 ± 0.22 | [119] | ||||
| Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 3.02 ± 0.11 | 101.6 at 100 µM 90.7 at 10 µM | [126] | ||||
| Scutellarin (scutellarein-7-glucuronide) | Scutellaria, Erigerontis herba | Binds to the 3CLpro substrate binding site. | Vero cells/RT PCR | 28.9 ± 1.6 at 50 µM | [119] | ||||
| Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 76.8 at 100 µM 18.9 at 10 µM | [126] | |||||
| Vitexin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 180 ± 6 | 52 at 200 µM | [118] | ||
| Wogonin | Scutellaria baicalensis | IC50: 8.52 ± 0.54 µg/mL EC50: 0.74 ± 0.36 µg/mL | Binds to the 3CLpro substrate binding site. | Vero cells/RT PCR | 6.1 ± 0.8 at 50 µM | [119] | |||
| Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 3.6 at 100 µM | [126] | ||||||
| Wogonoside | Scutellaria baicalensis | IC50: 8.52 ± 0.54 µg/mL EC50: 0.74 ± 0.36 µg/mL | Binds to the 3CLpro substrate binding site. | Vero cells/RT PCR | 8.5 ± 3.3 at 50 µM | [119] | |||
| Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 20.4 at 100 µM | [126] | ||||||
| Isoflavones | Biochanin A | Interacts with 3CLpro catalytic site | FRET assay | 5 at 10 µM | [117] | ||||
| Daidzein | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 56 | 100 at 200 µM | [118] | ||
| Interacts with 3CLpro catalytic site | FRET assay | 13.9 at 10 µM | [117] | ||||||
| Formononetin | Interacts with 3CLpro catalytic site | FRET assay | 16.0 at 10 µM | [117] | |||||
| Genistein | Interacts with 3CLpro catalytic site | FRET assay | 15.0 at 10 µM | [117] | |||||
| Genistin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 48 at 200 µM | [118] | |||
| Interacts with 3CLpro catalytic site | FRET assay | 25.5 at 10 µM | [117] | ||||||
| Puerarin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 42 ± 2 | 100 at 200 µM | [118] | ||
| Sophoricoside | Interacts with 3CLpro catalytic site | FRET assay | 10.3 at 10 µM | [117] | |||||
| Flavan-3-ols/Flavanols | Catechin | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 9 at 200 µM | [118] | ||
| Interacts with 3CLpro catalytic site | FRET assay | 14.0 at 10 µM | [117] | ||||||
| Catechin gallate (CG) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 21 at 200 µM | [118] | |||
| Epicatechin (EC) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 8 at 200 µM | [118] | |||
| Epicatechin gallate (ECG) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 21 at 200 µM | [118] | |||
| Epigallocatechin (EGC) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 23 at 200 µM | [118] | |||
| Epigallocatechin gallate (EGCG) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 171 ± 5 | 53 at 200 µM | [118] | ||
| TCM | Binds to the 3CLpro substrate binding site. | FRET assay | 0.847 ± 0.005 | [121] | |||||
| Gallocatechin gallate (GCG) | Black garlic extract | IC50: 137 ± 10 µg/mL 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 50 at 200 µM | [118] | |||
| Tannoid | Tannic acid | Black garlic extract | IC50: 137 ± 10 µg/mL, 100% inhibition at 0.5 mg/mL | Inhibits 3CLpro activity | FRET assay | 9 | 100 at 200 µM | [118] | |
| Others | Mixture of 11 flavonols | Salvadora persica L. | Inhibits 3CLpro activity | 3CL protease assay, A549 cells | 8.59 ± 0.3 μg mL−1 | 85.56 ± 1.12% | [127] |
| Class | Flavonoid | Natural Source (N.S.)/Extract | Mode of Action | Methods Used | % Inhibition | Reference |
|---|---|---|---|---|---|---|
| Flavonol | Myricetin | Interacts with 3CLpro catalytic site | FRET assay | 50 at 159.10 ± 38.33 µM | [117] | |
| Rutin | Binds to naphthalene inhibitor binding pocket | PLpro enzymatic inhibition assay | 38 at 100 µM | [128] | ||
| Flavones | Baicalein | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 45.1 at 50 µM, 12.4 at 12.5 µM | [126] |
| Baicalin | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 15.9 at 50 µM | [126] | |
| Chrysin-7-O-β-D-glucuronide | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 16.3 at 50 µM | [126] | |
| Luteoloside (cyranoside) | L. japonica | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 21.5 at 50 μM | [126] | |
| Oroxylin A-7-O-β-D-glucuronide | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 7.4 at 50 μM | [126] | |
| Scutellarein | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 65.7 at 50 µM, 14.4 at 12.5 µM | [126] | |
| Scutellarin | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 41.1 at 50 µM, 12.7 at 12.5 µM | [126] | |
| Wogonin | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 52.0 at 50 µM, 35.9 at 12.5 µM | [126] | |
| Wogonoside | Scutellaria baicalensis | Binds to 3CLpro active site | Vero E6 cells/CCK8 assays | 14.4 at 50 µM | [126] | |
| Flavan-3-ols/Flavanols | Epicatechin gallate (ECG) | Binds to naphthalene inhibitor binding pocket | PLpro enzymatic inhibition assay | 20 at 100 µM | [128] | |
| Epigallocatechin gallate (EGCG) | Binds to naphthalene inhibitor binding pocket | PLpro enzymatic inhibition assay | 13 at 100 µM | [128] | ||
| Anthocyanin | Cyanidin-3-O-glucoside (chrysanthemin) | Binds to naphthalene inhibitor binding pocket | PLpro enzymatic inhibition assay | 20 at 100 µM | [128] | |
| Others | Hypericin | Binds to naphthalene inhibitor binding pocket | PLpro enzymatic inhibition assay | 97 at 100 µ M87 at 50 µM | [128] |
| Class | Flavonoid | Natural Source (N.S.)/Extract | Efficacy of N.S. Extract | Mode of Action | Methods Used | IC50 (µM) | % Inhibition | Reference |
|---|---|---|---|---|---|---|---|---|
| Flavonol | Isorhamnetin | Sea buckthorn berry | Binds to three residues involved in spike RBD–ACE2 interaction | HEK293 cells/SPR assay | Kd = 2.51 ± 0.68 μM | [125] | ||
| Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 14.7 ± 1.4 at 10 µM | [129] | |||||
| Quercetin | Hippophae rhamnoides L. | Binds to three residues involved in spike RBD–ACE2 interaction | HEK293 cells/SPR assay | Kd = 5.92 ± 0.92 μM | [125] | |||
| Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | At 2.5 min: 4.48 At 10.5 min: 29.5 | 66.2 ± 2.2 at 10 µM | [129] | ||||
| Quercetin-3-O-galactoside (hyperoside) | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 34.2 ± 3.7 at 10 µM | [129] | ||||
| Quercetin-3-O-glucuronide | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 33.1 ± 4.9 at 10 µM | [129] | ||||
| Quercetin-3-O-glucoside (isoquercetin) | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 47.7 ± 3.7 at 10 µM | [129] | ||||
| Rutin (quercetin-3-)-rutinose) | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 48.3 ± 4.7 at 10 µM | [129] | ||||
| Tamarixetin | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 41.5 ± 5.0 at 10 µM | [129] | ||||
| Flavanone | (±)-Eriodictyol | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 24.4 ± 1.4 at 10 µM | [129] | |||
| Hesperetin | Anatolian Propolis | IC50: 1.14 µL | S1 colorimetric assay | 16,880 | [130] | |||
| Pinocembrin | Anatolian Propolis | IC50: 1.14 µL | S1 colorimetric assay | 29,530 | [130] | |||
| Flavones | Luteolin | Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 37.1 ± 0.6 at 10 µM | [129] | |||
| Neobaicalein | Radix Scutellariae | Binds to ACE2 receptor | CMC, HEK293T cells, CK8 assay, SPR assay | 83.8 | [120] | |||
| Oroxylin A | Radix Scutellariae | Binds to ACE2 receptor | CMC, HEK293T cells, CK8 assay, SPR assay | 164.6 | [120] | |||
| Scutellarin | Radix Scutellariae | Binds to ACE2 receptor | CMC, HEK293T cells, CK8 assay, SPR assay | 170.9 | [120] | |||
| Wogonin | Radix Scutellariae | Binds to ACE2 receptor | CMC, HEK293T cells, CK8 assay, SPR assay | 137.6 | [120] | |||
| Flavan-3-ols/Flavanols | Epicatechin (EC) | Green tea | Interferes with SARS-CoV-2 spike RBD–ACE2 interaction | HEK293T-ACE2 cells/Huh7 cells/Vero cells | >20 µg/mL | [131] | ||
| Inhibits rhACE2 activity | MCA Fluorescence, rhACE2 cells | 27.4 ± 5.7 at 10 µM | [129] | |||||
| Epigallocatechin gallate (EGCG) | Green tea | Interferes with SARS-CoV-2 spike RBD–ACE2 interaction | HEK293T-ACE2 cells/Huh7 cells/Vero cells/Plaque reduction assay | 2.47 µg/mL | [131] | |||
| Anthocyanin | Pelargonidin | Binds to fatty acid binding pocket on spike RBD and attenuates spike–ACE2 interaction Reduces SARS-CoV-2 replication | ACE2-SARS-CoV-2 spike inhibitor screening assay Vero cells/Plaque assay | Screening Assay: >5 (10 µM), >15 (20 µM), >40 (50 µM) | [132] |
| Target | Class | Flavonoid | Natural Source (N.S.)/Extract | Mode of Action | Methods Used | IC50 (µM) | EC50 (µM) | % Inhibition | Reference |
|---|---|---|---|---|---|---|---|---|---|
| RdRp | Flavone | Baicalein | Scutellaria baicalensis | Binding to NiRAN domain and the palm subdomain | Vero CCL-81/Calu-3 cells/MTS assay/qRT-PCR assay/293T cells/Huh7.5 cells | Vero: 4.5 ± 0.2 Calu-3: 1.2 + 0.03 | 99.8 at 20 µM | [134] | |
| Flavone | Baicalin | Scutellaria baicalensis | Inhibits RdRp | Vero CCL-81/Calu-3 cells/MTS assay/qRT-PCR assay/293T cells/Huh7.5 cells | Vero: 9.0 ± 0.08 Calu-3: 8.0 ± 0.11 | 98 at 20 µM | [134] | ||
| N | Flavanol/flavan-3-ol | Gallocatechin Gallate (GCG) | Green Tea | Disrupts the LLPS of N by interfering with N-RNA binding | H1299 cells RT-qPCR | 44.4 | [1] | ||
| Mitochondrial OXPHOS | Flavone | Baicalein | Scutellaria baicalensis | Oxygen consumption inhibitor | Vero E6 cells | 10 | [135] | ||
| nsP15 | Flavanol/flavan-3-ol | epigallocatechin gallate (EGCG) | Green Tea extract | Binds to nsp15 active site | Endoribonuclease assay, plaque assay, Vero cells | 1.62 ± 0.36 | PRNT50: 0.2 µM | [136] | |
| Flavone | Baicalin | Scutellaria baicalensis, Scutellaria lateriflora | Endoribonuclease assay, plaque assay, Vero cells | 7.98 ± 1.46 | PRNT50: 83.3 µM | [136] | |||
| Flavone | Baicalein | Scutellaria baicalensis, Scutellaria lateriflora | Endoribonuclease assay, plaque assay, Vero cells | 8.61 | [136] | ||||
| Flavonol | Quercetin | Onion peels, red grapes, green leafy vegetables | Endoribonuclease assay, plaque assay, Vero cells | 13.79 | [136] |
| Study Title | Study Type | Number of Subjects Enrolled | Status |
|---|---|---|---|
| Nigella Sativa in COVID-19 (NCT04401202) | Prospective, Randomized, Open-label | 183 COVID-19 positive participants * | Completed |
| Efficacy of Psidii Guava’s Extract For COVID-19 (NCT04810728) | Experimental, randomized, double-blind clinical trial | 90 COVID-19 positive participants b/w 13-59 yrs. | Phase 3 |
| The Effectiveness of Phytotherapy in SARS-CoV 2 (COVID-19) (NCT04851821) | Randomized, double—masked, interventional clinical Trial with Parallel Assignment | 80 COVID-19 positive participants * | Phase 1 |
| Masitinib Combined with Isoquercetin and Best Supportive Care in Hospitalized Patients With Moderate and Severe COVID-19 (NCT04622865) | Randomized, double-blinded, triple-masked interventional clinical trial with Parallel Assignment | 200 COVID-19 positive participants * | Phase 2 |
| Quercetin In The Treatment of SARS-CoV 2 (NCT04853199) | Randomized, double-blinded, triple-masked interventional clinical trial with Parallel Assignment | 200 COVID-19 positive participants * | Early Phase 1 |
| Randomized Proof-of-Concept Trial to Evaluate the Safety and Explore the Effectiveness of Resveratrol, a Plant Polyphenol, for COVID-19 (NCT04400890) | Randomized placebo-controlled, double-blinded, quadruple-masked, interventional clinical trial | 100 COVID-19 positive participants ≥ 45 yrs. | Phase 2 |
| Tannin Specific Natural Extract for COVID-19 Infection (NCT04403646) | Double-blind, randomized, triple-masked | 124 COVID-19 positive participants * | n/a |
| P2Et Extract in the Symptomatic Treatment of Subjects With COVID-19 (NCT04410510) | Double-blind, randomized, triple-masked, interventional clinical trial with parallel assignment | 100 COVID-19 positive participants * | Phase 2/3 |
| COVID-19, Hospitalized, Patients, Nasafytol (NCT04844658) | Standard-of-care comparative, open-labelled, parallel two-arms and randomized trial | 50 COVID-19 positive participants * | Recruiting |
| Study to Investigate the Clinical Benefits of Dietary Supplement Quercetin for Managing Early COVID-19 Symptoms at Home (NCT04861298) | Open-labelled, randomized, parallel-assignment, interventional trial | 142 COVID-19 positive participants * | Recruiting |
| Complementary Intervention for COVID-19 (NCT04487964) | Open-labelled, non-randomized | 70 COVID-19 positive participants * | Recruiting |
| The Study of Quadruple Therapy Zinc, Quercetin, Bromelain and Vitamin C on the Clinical Outcomes of Patients Infected With COVID-19 (NCT04468139) | Open-labelled, single-assignment, interventional trial | 60 COVID-19 positive participants * | Phase 4 |
| Evaluation of the Effect of Anatolian Propolis on COVID-19 in Healthcare Professionals (NCT04680819) | Observational, prospective cohort study | 50 HCWs at risk for developing COVID-19 | Not yet recruiting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaul, R.; Paul, P.; Kumar, S.; Büsselberg, D.; Dwivedi, V.D.; Chaari, A. Promising Antiviral Activities of Natural Flavonoids against SARS-CoV-2 Targets: Systematic Review. Int. J. Mol. Sci. 2021, 22, 11069. https://doi.org/10.3390/ijms222011069
Kaul R, Paul P, Kumar S, Büsselberg D, Dwivedi VD, Chaari A. Promising Antiviral Activities of Natural Flavonoids against SARS-CoV-2 Targets: Systematic Review. International Journal of Molecular Sciences. 2021; 22(20):11069. https://doi.org/10.3390/ijms222011069
Chicago/Turabian StyleKaul, Ridhima, Pradipta Paul, Sanjay Kumar, Dietrich Büsselberg, Vivek Dhar Dwivedi, and Ali Chaari. 2021. "Promising Antiviral Activities of Natural Flavonoids against SARS-CoV-2 Targets: Systematic Review" International Journal of Molecular Sciences 22, no. 20: 11069. https://doi.org/10.3390/ijms222011069
APA StyleKaul, R., Paul, P., Kumar, S., Büsselberg, D., Dwivedi, V. D., & Chaari, A. (2021). Promising Antiviral Activities of Natural Flavonoids against SARS-CoV-2 Targets: Systematic Review. International Journal of Molecular Sciences, 22(20), 11069. https://doi.org/10.3390/ijms222011069