
Quantitative Analysis of the Cardiac Phosphoproteome in Response to Acute β-Adrenergic Receptor Stimulation In Vivo

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Regulation of Protein Expression in Response to Acute β-AR Stimulation
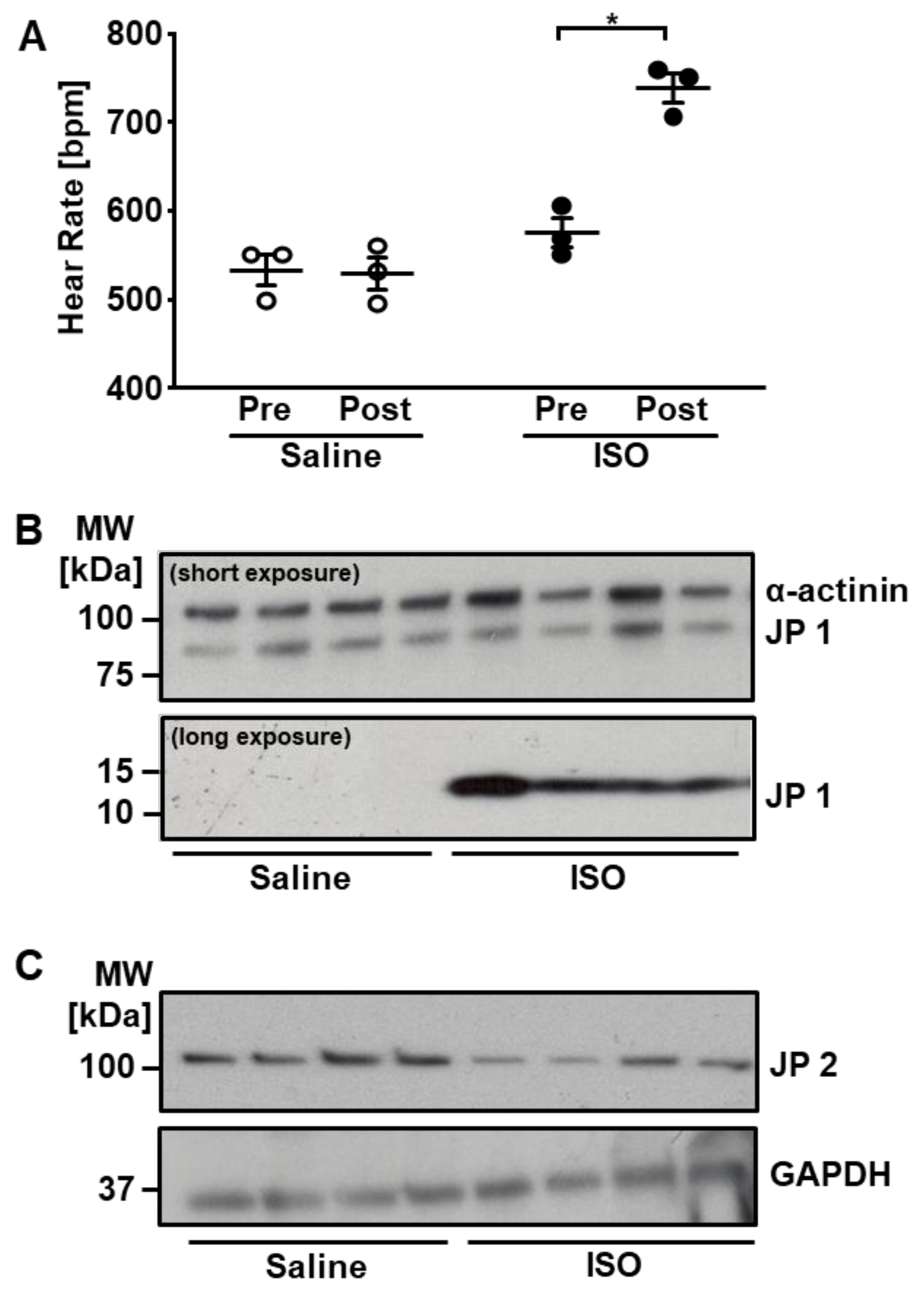
2.2. Cardiac Expression and ISO-Induced Fragmentation of Junctophilin-1
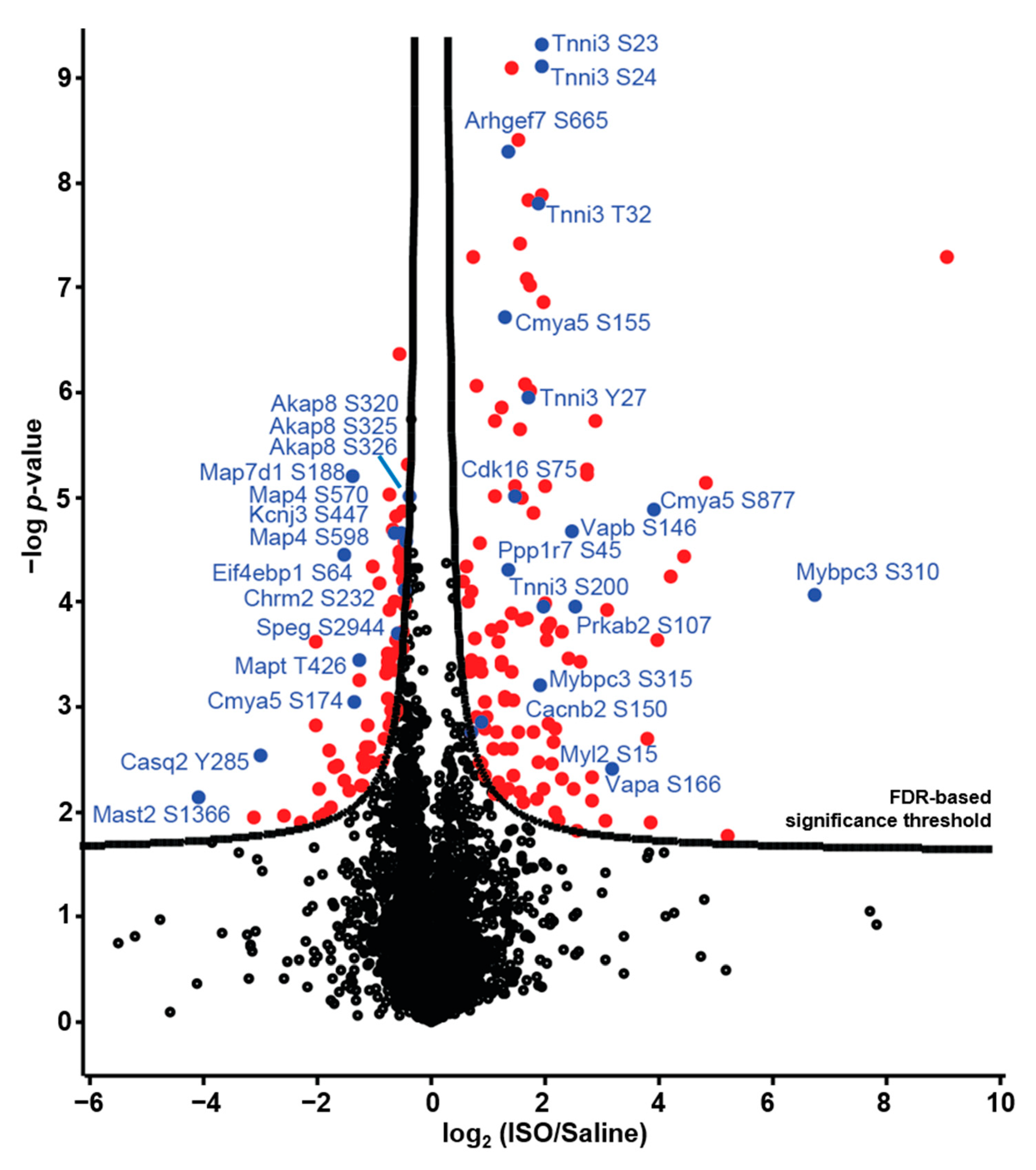
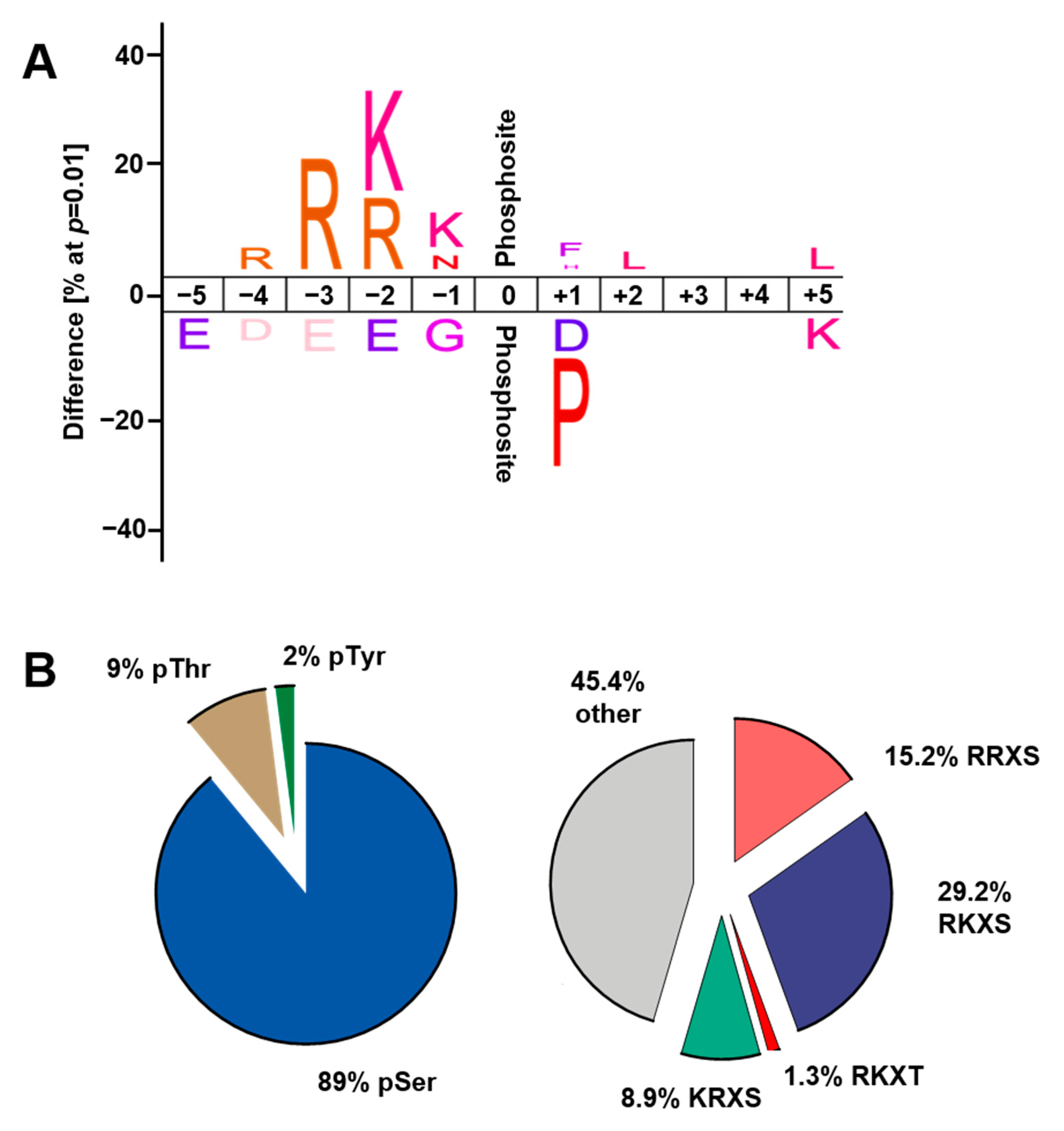
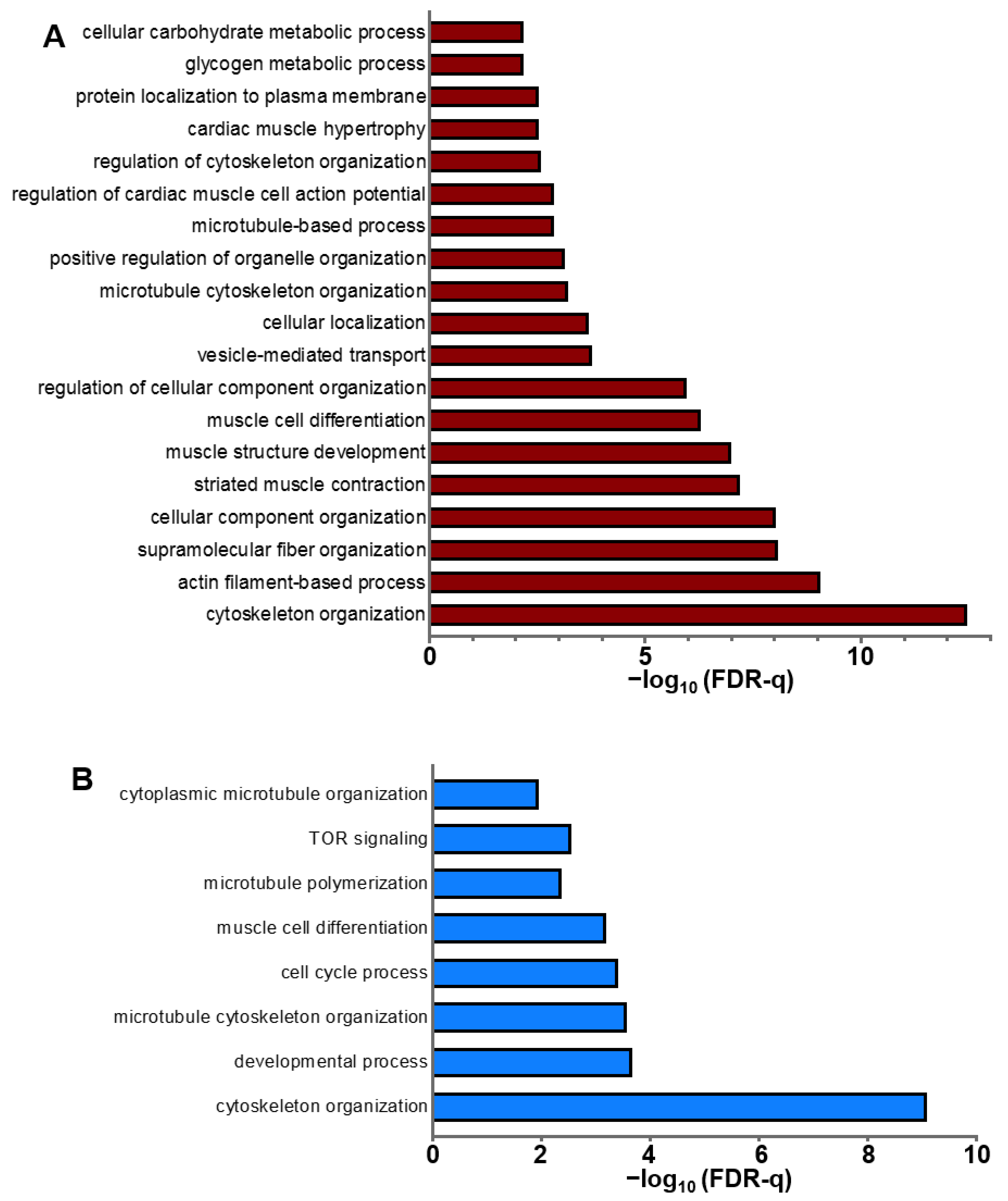
2.3. Regulation of Cardiac Protein Phosphorylation in Response to Acute β-AR Stimulation
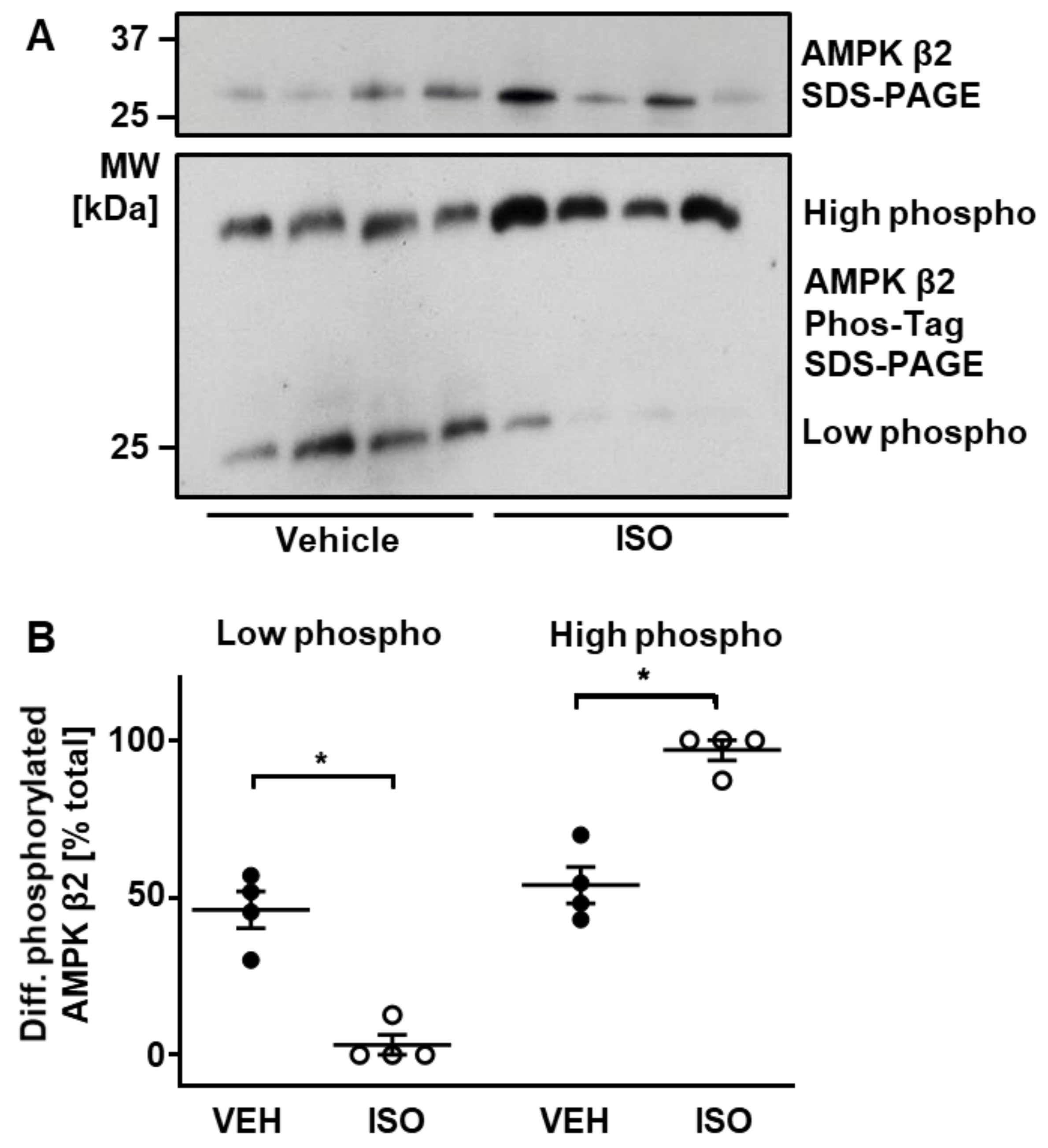
2.4. ISO-Induced Phosphorylation of AMPK β2 Subunit
3. Discussion
3.1. Cardiac Expression and Fragmentation of JP1
3.2. Cardiac Protein Phosphorylation
3.3. Kinases and Phosphatases Displaying Altered Phosphorylation in Response to ISO
3.4. ISO-Induced Protein Dephosphorylation
3.5. Study Limitations
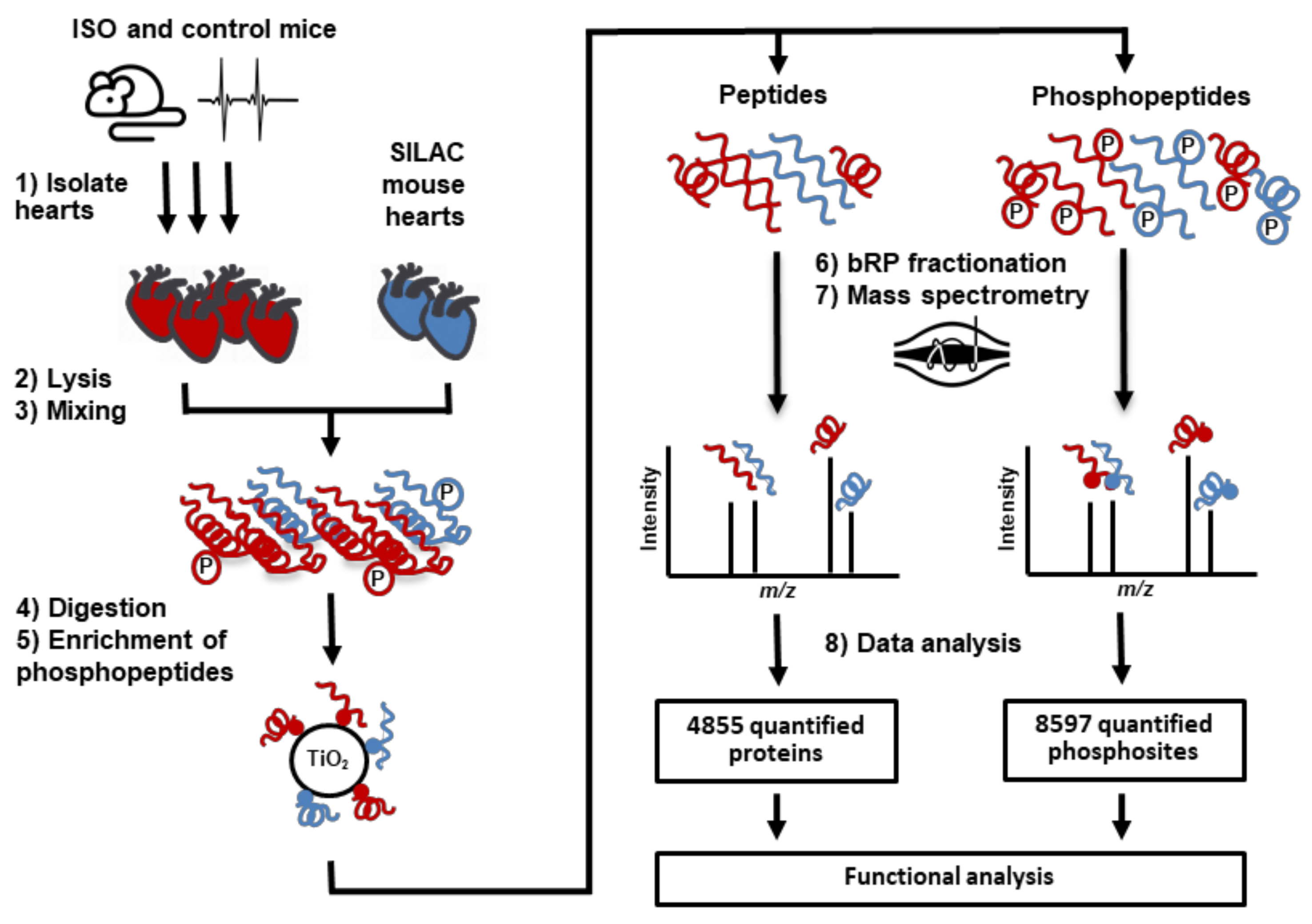
4. Materials and Methods
4.1. Preparation of Mouse Tissue Samples
4.2. Protein Extraction and Digestion
4.3. Phosphopeptide Enrichment
4.4. High-pH Reverse Phase Fractionation
4.5. Mass Spectrometric Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rockman, H.A.; Koch, W.J.; Lefkowitz, R.J. Seven-transmembrane-spanning receptors and heart function. Nature 2002, 415, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Dao, K.K.D.; Teigen, K.; Kopperud, R.; Hodneland, E.; Schwede, F.; Christensen, A.E.; Martinez, A.; Døskeland, S.O. Epac1 and cAMP-dependent protein kinase holoenzyme have similar cAMP affinity, but their cAMP domains have distinct structural features and cyclic nucleotide recognition. J. Biol. Chem. 2006, 281, 21500–21511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, M.; Brown, J.H. β-Adrenergic receptor signaling in the heart: Role of CaMKII. J. Mol. Cell. Cardiol. 2010, 48, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, S.; Meyer-Roxlau, S.; El-Armouche, A. Role of protein phosphatase inhibitor-1 in cardiac beta adrenergic pathway. J. Mol. Cell. Cardiol. 2016, 101, 116–126. [Google Scholar] [CrossRef]
- Ranieri, A.; Kemp, E.; Burgoyne, J.R.; Avkiran, M. β-Adrenergic regulation of cardiac type 2A protein phosphatase through phosphorylation of regulatory subunit B56δ at S573. J. Mol. Cell. Cardiol. 2018, 115, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Cuello, F.; Mayr, U.; Hao, Z.; Hornshaw, M.; Ehler, E.; Avkiran, M.; Mayr, M. Proteomics analysis of the cardiac myofilament subproteome reveals dynamic alterations in phosphatase subunit distribution. Mol. Cell. Proteom. 2010, 9, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Hulme, J.T.; Westenbroek, R.E.; Scheuer, T.; Catterall, W.A. Phosphorylation of serine 1928 in the distal C-terminal domain of cardiac Ca v 1.2 channels during β1-adrenergic regulation. Proc. Natl. Acad. Sci. USA 2006, 103, 16574–16579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, B.; Jiang, M.T.; Zhao, M.; Yang, D.; Sutherland, C.; Lai, F.A.; Walsh, M.P.; Warltier, D.C.; Cheng, H.; Chen, S.R.W. Characterization of a novel PKA phosphorylation site, serine-2030, reveals no PKA hyperphosphorylation of the cardiac ryanodine receptor in canine heart failure. Circ. Res. 2005, 96, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Armouche, A.; Pamminger, T.; Ditz, D.; Zolk, O.; Eschenhagen, T. Decreased protein and phosphorylation level of the protein phosphatase inhibitor-1 in failing human hearts. Cardiovasc. Res. 2004, 61, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Rotman, E.I.; De Jongh, K.S.; Florio, V.; Lai, Y.; Catterall, W.A. Specific phosphorylation of a site in the full-length form of the alpha-1 subunit of the cardiac L-type calcium channel by cAMP-dependent protein kinase. Biochem. 1996, 35, 10392–10402. [Google Scholar]
- Bossuyt, J.; Ai, X.; Moorman, J.R.; Pogwizd, S.M.; Bers, D.M. Expression and phosphorylation of the Na-pump regulatory subunit phospholemman in heart failure. Circ. Res. 2005, 97, 558–565. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.E.; Blagoev, B.; Kratchmarova, I.; Kristensen, D.B.; Steen, H.; Pandey, A.; Mann, M. Stable isotope labeling by amino acids in cell culture, SILAC, as a simple and accurate approach to expression proteomics. Mol. Cell. Proteom. 2002, 1, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Rauniyar, N.; McClatchy, D.B.; Yates, J.R. Stable isotope labeling of mammals (SILAM) for in vivo quantitative proteomic analysis. Methods 2013, 61, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Krüger, M.; Moser, M.; Ussar, S.; Thievessen, I.; Luber, C.A.; Forner, F.; Schmidt, S.; Zanivan, S.; Fässler, R.; Mann, M. SILAC Mouse for Quantitative Proteomics Uncovers Kindlin-3 as an Essential Factor for Red Blood Cell Function. Cell 2008, 134, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monetti, M.; Nagaraj, N.; Sharma, K.; Mann, M. Large-scale phosphosite quantification in tissues by a spike-in SILAC method. Nat. Methods 2011, 8, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Geiger, T.; Wisniewski, J.R.; Cox, J.; Zanivan, S.; Kruger, M.; Ishihama, Y.; Mann, M. Use of stable isotope labeling by amino acids in cell culture as a spike-in standard in quantitative proteomics. Nat. Protoc. 2011, 6, 147–157. [Google Scholar] [CrossRef]
- Murphy, R.M.; Dutka, T.L.; Horvath, D.; Bell, J.R.; Delbridge, L.M.; Lamb, G.D. Ca2+-dependent proteolysis of junctophilin-1 and junctophilin-2 in skeletal and cardiac muscle. J. Physiol. 2013, 591, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Wang, Y.; Chen, B.; Wang, Y.; Yuan, J.; Zhang, L.; Hall, D.; Wu, J.; Shi, Y.; Zhu, Q.; et al. E-C coupling structural protein junctophilin-2 encodes a stress-adaptive transcription regulator. Science 2018, 362, eaan3303. [Google Scholar] [CrossRef] [PubMed]
- Takeshima, H.; Komazaki, S.; Nishi, M.; Iino, M.; Kangawa, K. Junctophilins: A novel family of junctional membrane complex proteins. Mol. Cell 2000, 6, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Golini, L.; Chouabe, C.; Berthier, C.; Cusimano, V.; Fornaro, M.; Bonvallet, R.; Formoso, L.; Giacomello, E.; Jacquemond, V.; Sorrentino, V. Junctophilin 1 and 2 proteins interact with the L-type Ca 2+ channel dihydropyridine receptors (DHPRs) in skeletal muscle. J. Biol. Chem. 2011, 286, 43717–43725. [Google Scholar] [CrossRef] [Green Version]
- Guo, A.; Hall, D.; Zhang, C.; Peng, T.; Miller, J.D.; Kutschke, W.; Grueter, C.E.; Johnson, F.L.; Lin, R.Z.; Song, L.S. Molecular determinants of calpain-dependent cleavage of junctophilin-2 protein in cardiomyocytes. J. Biol. Chem. 2015, 290, 17946–17955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layland, J.; Grieve, D.J.; Cave, A.C.; Sparks, E.; Solaro, R.J.; Shah, A.M. Essential role of troponin I in the positive inotropic response to isoprenaline in mouse hearts contracting auxotonically. J. Physiol. 2004, 556, 835–847. [Google Scholar] [CrossRef]
- Sadayappan, S.; Gulick, J.; Osinska, H.; Barefield, D.; Cuello, F.; Avkiran, M.; Lasko, V.M.; Lorenz, J.N.; Maillet, M.; Martin, J.L.; et al. A critical function for Ser-282 in cardiac myosin binding protein-C phosphorylation and cardiac function. Circ. Res. 2011, 109, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, B.E.; Pearson, R.B. Protein kinase recognition sequence motifs. Trends Biochem. Sci. 1990, 15, 342–346. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Warden, S.M.; Richardson, C.; O’Donnell, J.J.; Stapleton, D.; Kemp, B.E.; Witters, L.A. Post-translational modifications of the β-1 subunit of AMP-activated protein kinase affect enzyme activity and cellular localization. Biochem. J. 2001, 354, 275–283. [Google Scholar] [CrossRef]
- Mitchelhill, K.I.; Michell, B.J.; House, C.M.; Stapleton, D.; Dyck, J.; Gamble, J.; Ullrich, C.; Witters, L.A.; Kemp, B.E. Posttranslational modifications of the 5’-AMP-activated protein kinase β1 subunit. J. Biol. Chem. 1997, 272, 24475–24479. [Google Scholar] [CrossRef] [Green Version]
- Gavi, S.; Shumay, E.; Wang, H.Y.; Malbon, C.C. G-protein-coupled receptors and tyrosine kinases: Crossroads in cell signaling and regulation. Trends Endocrinol. Metab. 2006, 17, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Najafi, A.; Sequeira, V.; Kuster, D.W.D.; van der Velden, J. β-adrenergic receptor signalling and its functional consequences in the diseased heart. Eur. J. Clin. Investig. 2016, 46, 362–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijnker, P.J.M.; Foster, D.B.; Tsao, A.L.; Frazier, A.H.; dos Remedios, C.G.; Murphy, A.M.; Stienen, G.J.M.; van der Velden, J. Impact of site-specific phosphorylation of protein kinase a sites ser23 and ser24 of cardiac troponin in human cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, 260–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layland, J.; Solaro, R.J.; Shah, A.M. Regulation of cardiac contractile function by troponin I phosphorylation. Cardiovasc. Res. 2005, 66, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Kirk, J.A.; Ji, W.; Dos Remedios, C.G.; Kass, D.A.; Van Eyk, J.E.; Murphy, A.M. Multiple reaction monitoring to identify site-specific troponin i phosphorylated residues in the failing human heart. Circulation 2012, 126, 1828–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salhi, H.E.; Walton, S.D.; Hassel, N.C.; Brundage, E.A.; de Tombe, P.P.; Janssen, P.M.L.; Davis, J.P.; Biesiadecki, B.J. Cardiac troponin I tyrosine 26 phosphorylation decreases myofilament Ca2+ sensitivity and accelerates deactivation. J. Mol. Cell. Cardiol. 2014, 76, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Wijnker, P.J.M.; Li, Y.; Zhang, P.; Foster, D.B.; dos Remedios, C.; Van Eyk, J.E.; Stienen, G.J.M.; Murphy, A.M.; van der Velden, J. A novel phosphorylation site, Serine 199, in the C-terminus of cardiac troponin I regulates calcium sensitivity and susceptibility to calpain-induced proteolysis. J. Mol. Cell. Cardiol. 2015, 82, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graeser, R.; Gannon, J.; Poon, R.Y.C.; Dubois, T.; Aitken, A.; Hunt, T. Regulation of the CDK-related protein kinase PCTAIRE-1 and its possible role in neurite outgrowth in Neuro-2A cells. J. Cell Sci. 2002, 115, 3479–3490. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, K.; Gong, K.; Fu, A.K.Y.; Ip, N.Y. Pctaire1 phosphorylates N-ethylmaleimide-sensitive fusion protein: Implications in the regulation of its hexamerization and exocytosis. J. Biol. Chem. 2006, 281, 9852–9858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirwany, N.A.; Zou, M.H. AMPK in cardiovascular health and disease. Acta Pharmacol. Sinica 2010, 31, 1075–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornton, C.; Snowden, M.A.; Carling, D. Identification of a novel AMP-activated protein kinase β subunit isoform that is highly expressed in skeletal muscle. J. Biol. Chem. 1998, 273, 12443–12450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.P.; Heierhorst, J.; Mann, R.J.; Mitchelhill, K.I.; Michell, B.J.; Witters, L.A.; Lynch, G.S.; Kemp, B.E.; Stapleton, D. Expression of the AMP-activated protein kinase β1 and β2 subunits in skeletal muscle. FEBS Lett. 1999, 460, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Koh, H.J.; Hirshman, M.F.; He, H.; Li, Y.; Manabe, Y.; Balschi, J.A.; Goodyear, L.J. Adrenaline is a critical mediator of acute exercise-induced AMP-activated protein kinase activation in adipocytes. Biochem. J. 2007, 403, 473–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, S.; Hiroi, Y.; Zou, Y.; Aikawa, R.; Toko, H.; Shibasaki, F.; Yazaki, Y.; Nagai, R.; Komuro, I. β-adrenergic pathway induces apoptosis through calcineurin activation in cardiac myocytes. J. Biol. Chem. 2000, 275, 34528–34533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, Z.; Steinberg, R.A. Cyclic AMP stimulates dephosphorylation of specific proteins in intact S49 mouse lymphoma cells. FEBS Lett. 1985, 180, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Chruscinski, A.J.; Singh, H.; Chan, S.M.; Utz, P.J. Broad-scale phosphoprotein profiling of beta adrenergic receptor (β-AR) signaling reveals novel Phosphorylation and Dephosphorylation events. PLoS ONE 2013, 8, e82164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamori, T.; Hayakawa, T.; Suzuki, M.; Titani, K. Identification of two 17-kDa rat parotid gland phosphoproteins, subjects for dephosphorylation upon β-adrenergic stimulation, as destrin- and cofilin-like proteins. J. Biol. Chem. 1995, 270, 8061–8067. [Google Scholar] [CrossRef] [Green Version]
- Weeks, K.L.; Ranieri, A.; Karaś, A.; Bernardo, B.C.; Ashcroft, A.S.; Molenaar, C.; McMullen, J.R.; Avkiran, M. β-Adrenergic Stimulation Induces Histone Deacetylase 5 (HDAC5) Nuclear Accumulation in Cardiomyocytes by B55α-PP2A-Mediated Dephosphorylation. J. Am. Heart Assoc. 2017, 6, e004861. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Chu, Z.; Han, J.; Zhang, Q.; Zhang, D.; Dang, Y.; Ren, J.; Chan, H.C.; Zhang, J.; Huang, Y. Phosphorylation-dependent mitochondrial translocation of MAP4 is an early step in hypoxia-induced apoptosis in cardiomyocytes. Cell Death Dis. 2014, 5, e1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhang, Q.; Zhang, X.; Zhang, J.; Wang, X.; Ren, J.; Jia, J.; Zhang, D.; Jiang, X.; Zhang, J.; et al. Microtubule associated protein 4 phosphorylation leads to pathological cardiac remodeling in mice. EBioMedicine 2018, 37, 221–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yang, F.; Gritsenko, M.A.; Wang, Y.; Clauss, T.; Liu, T.; Shen, Y.; Monroe, M.E.; Lopez-Ferrer, D.; Reno, T.; et al. Reversed-phase chromatography with multiple fraction concatenation strategy for proteome profiling of human MCF10A cells. Proteomics 2011, 11, 2019–2026. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Colaert, N.; Helsens, K.; Martens, L.; Vandekerckhove, J.; Gevaert, K. Improved visualization of protein consensus sequences by iceLogo. Nat. Methods 2009, 6, 786–787. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Güran, A.; Ji, Y.; Fang, P.; Pan, K.-T.; Urlaub, H.; Avkiran, M.; Lenz, C. Quantitative Analysis of the Cardiac Phosphoproteome in Response to Acute β-Adrenergic Receptor Stimulation In Vivo. Int. J. Mol. Sci. 2021, 22, 12584. https://doi.org/10.3390/ijms222212584
Güran A, Ji Y, Fang P, Pan K-T, Urlaub H, Avkiran M, Lenz C. Quantitative Analysis of the Cardiac Phosphoproteome in Response to Acute β-Adrenergic Receptor Stimulation In Vivo. International Journal of Molecular Sciences. 2021; 22(22):12584. https://doi.org/10.3390/ijms222212584
Chicago/Turabian StyleGüran, Alican, Yanlong Ji, Pan Fang, Kuan-Ting Pan, Henning Urlaub, Metin Avkiran, and Christof Lenz. 2021. "Quantitative Analysis of the Cardiac Phosphoproteome in Response to Acute β-Adrenergic Receptor Stimulation In Vivo" International Journal of Molecular Sciences 22, no. 22: 12584. https://doi.org/10.3390/ijms222212584
APA StyleGüran, A., Ji, Y., Fang, P., Pan, K. -T., Urlaub, H., Avkiran, M., & Lenz, C. (2021). Quantitative Analysis of the Cardiac Phosphoproteome in Response to Acute β-Adrenergic Receptor Stimulation In Vivo. International Journal of Molecular Sciences, 22(22), 12584. https://doi.org/10.3390/ijms222212584