Interleukin-1β Induced Matrix Metalloproteinase Expression in Human Periodontal Ligament-Derived Mesenchymal Stromal Cells under In Vitro Simulated Static Orthodontic Forces

 , and
, and 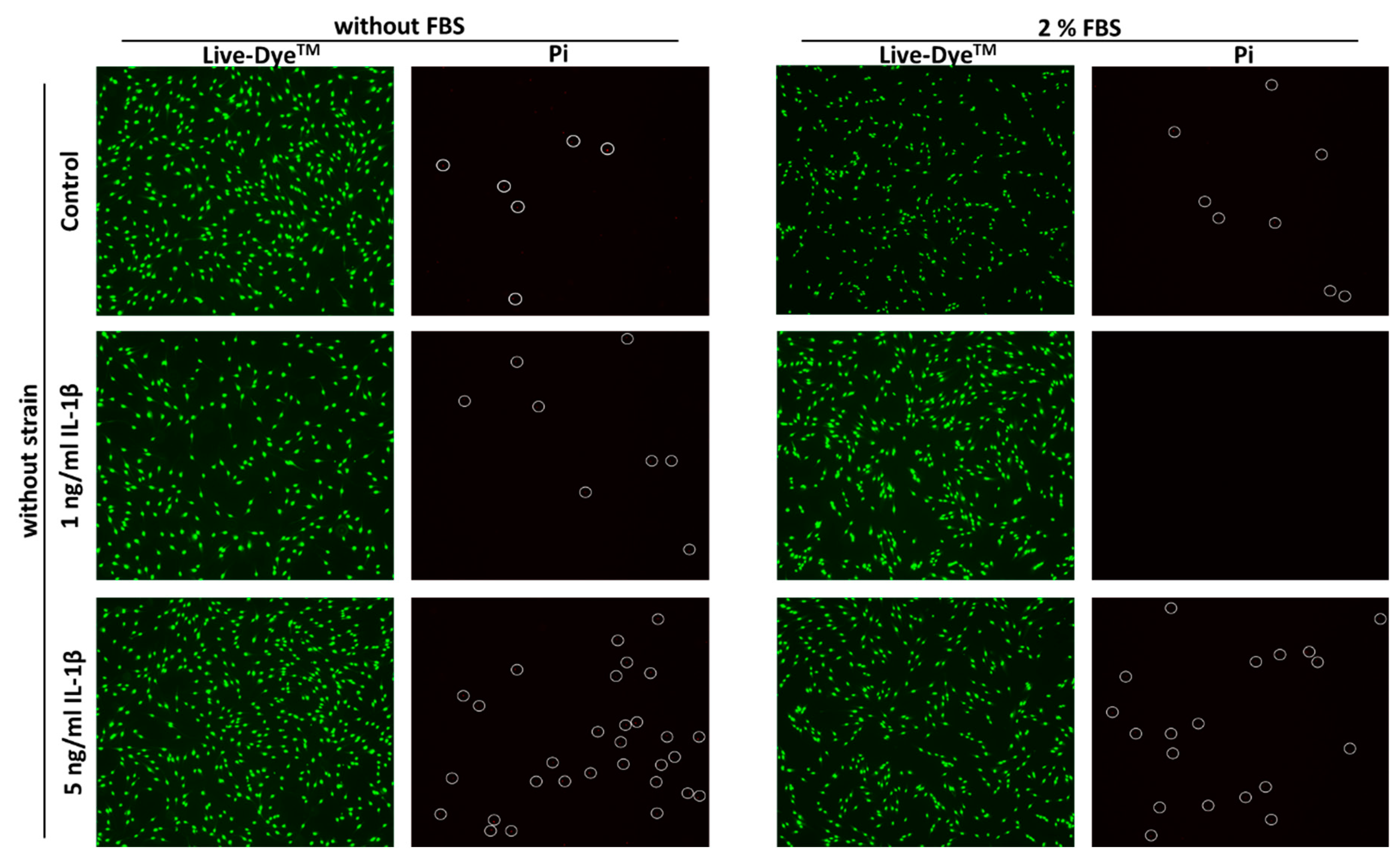{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Immunostaining of Certain Mesenchymal and Hematopoietic Surface Marker in hPDL-MSCs
2.2. Cell Viability
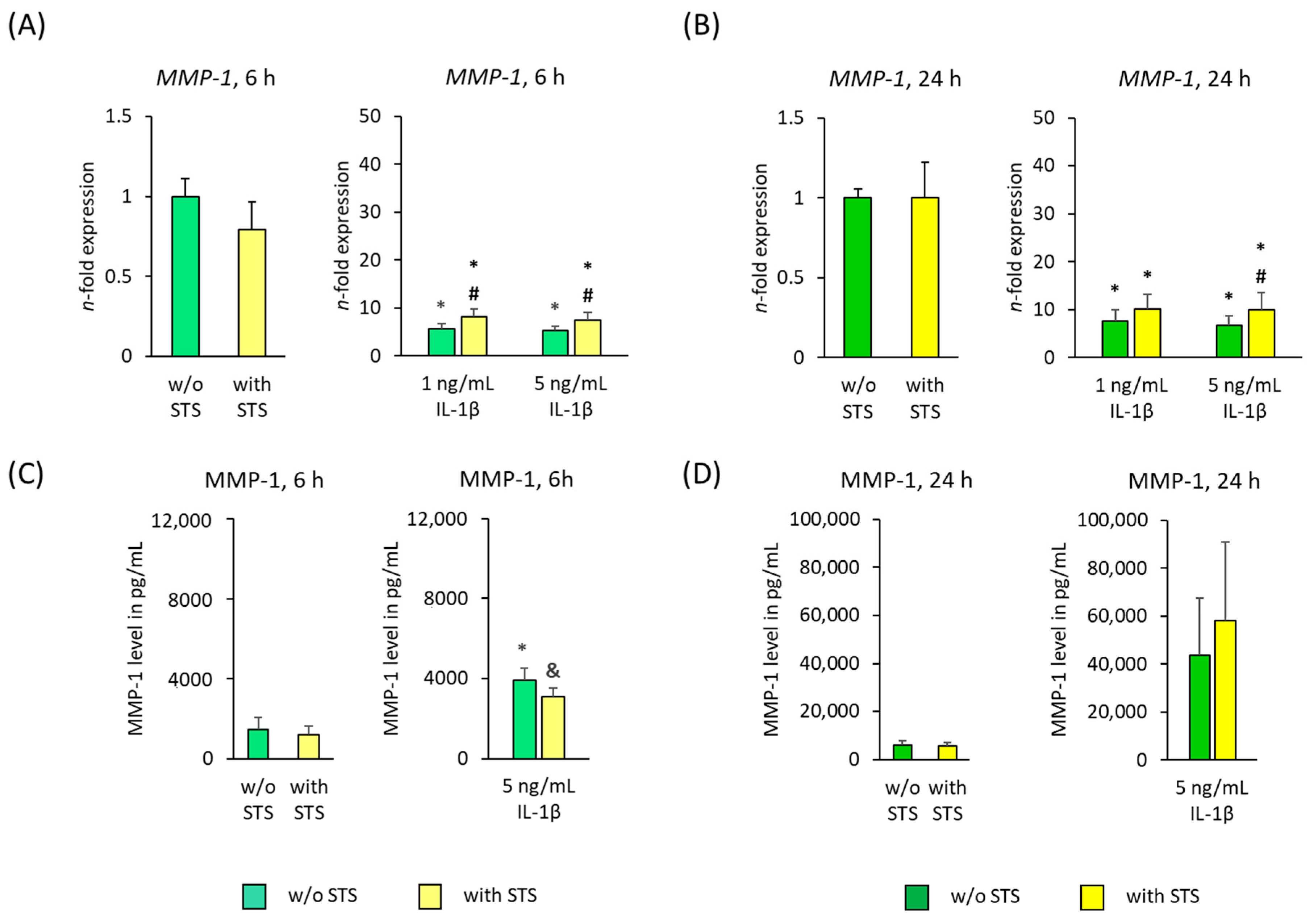
2.3. MMP-1 Expression
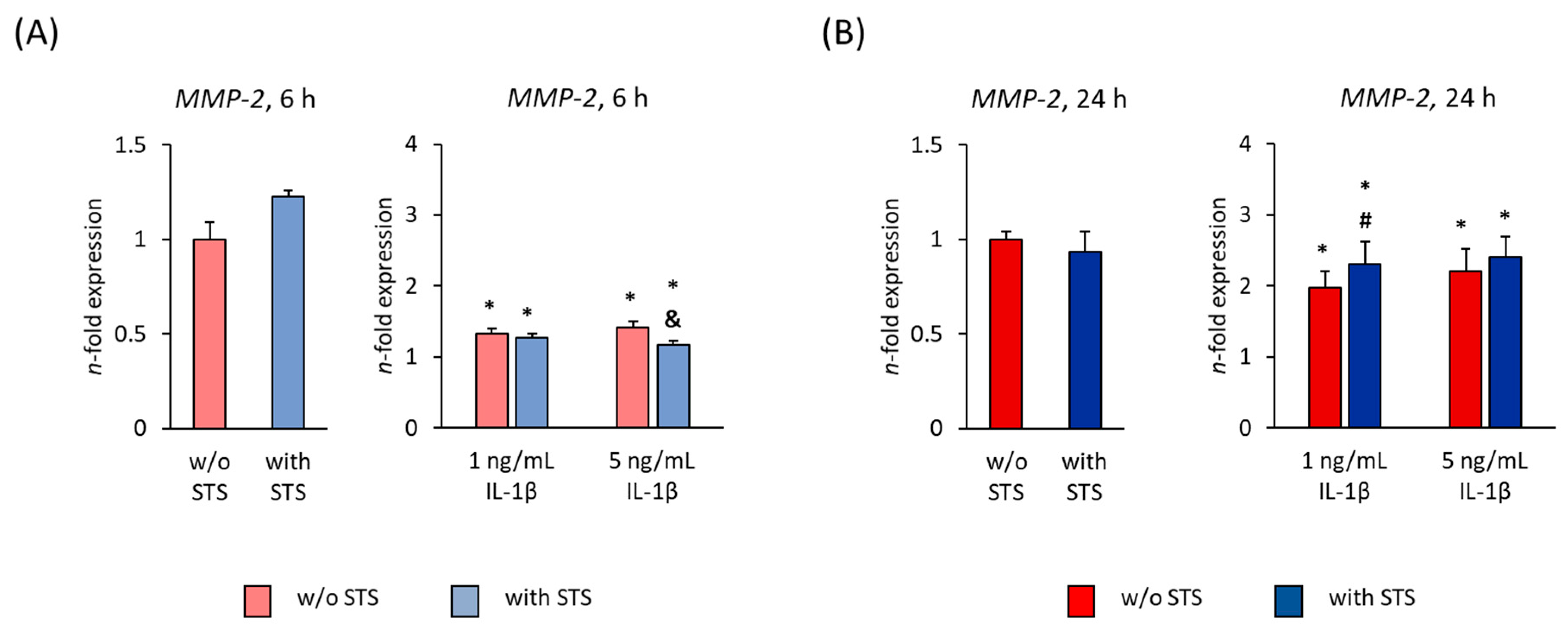
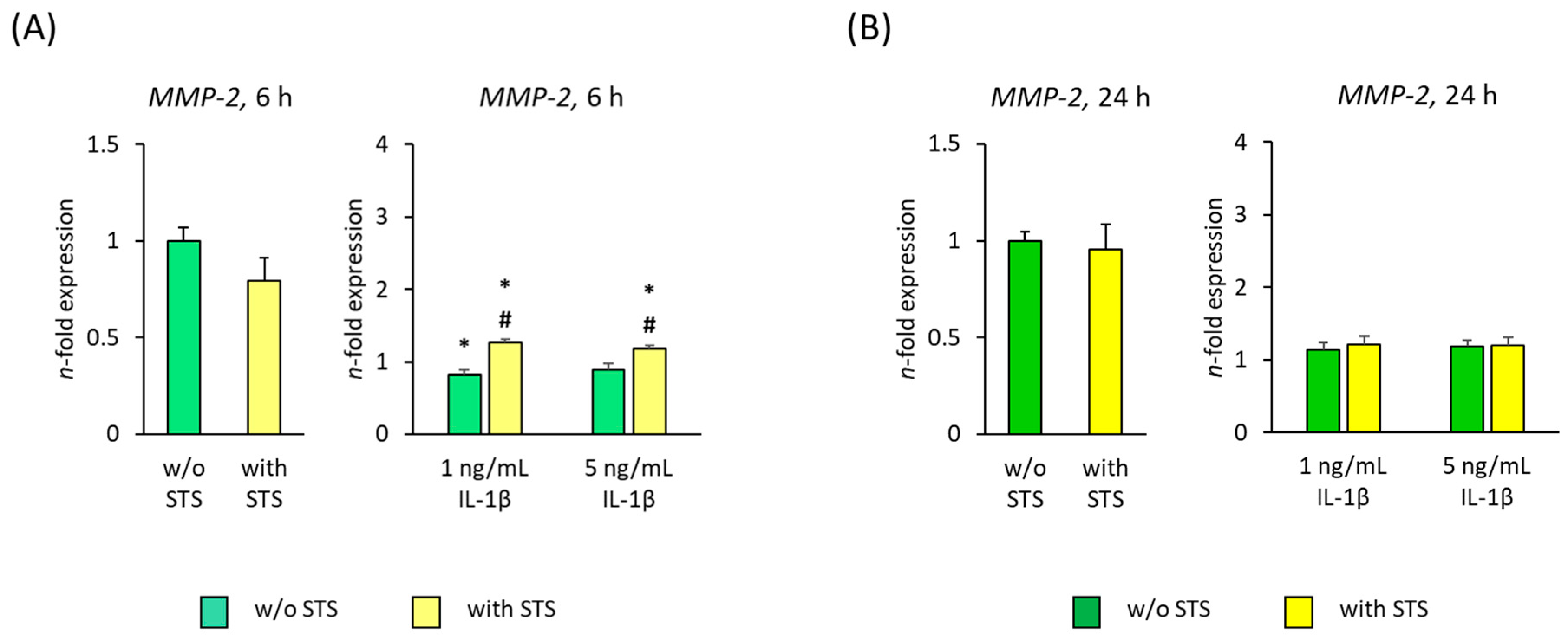
2.4. MMP-2 Expression
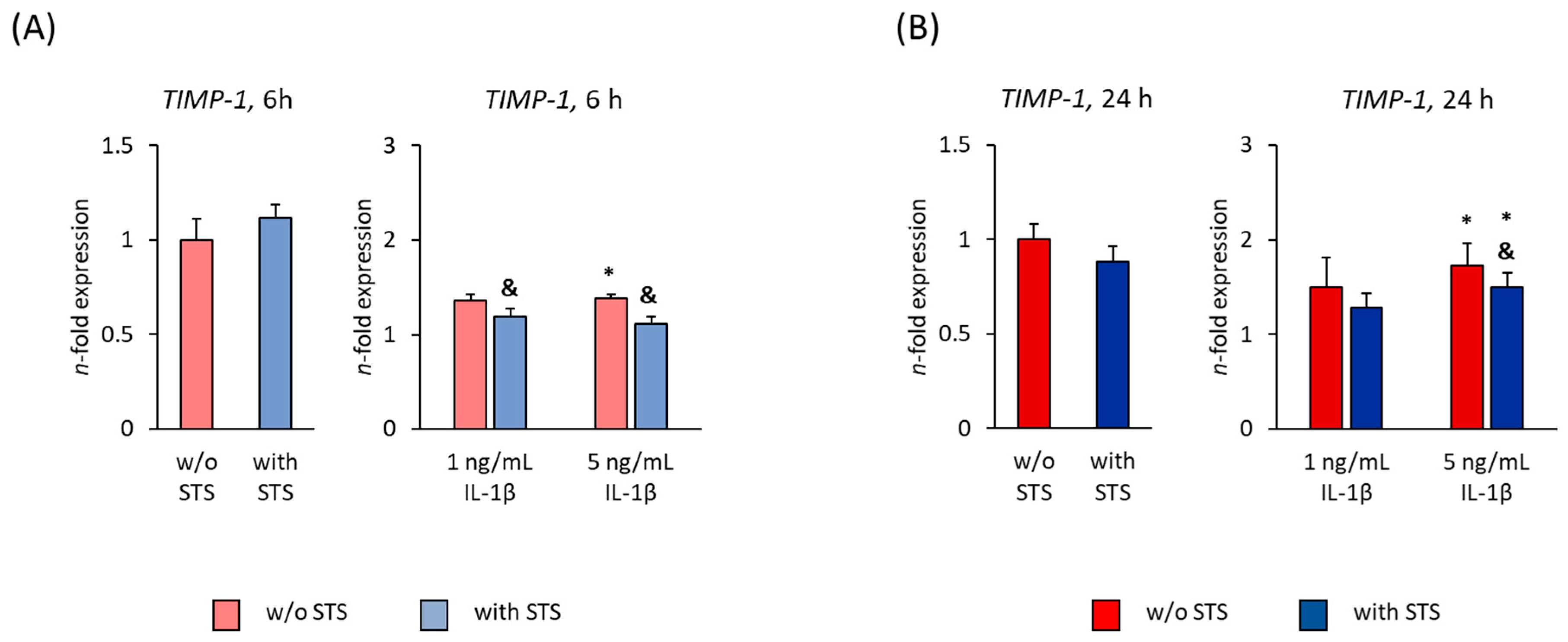
2.5. TIMP-1 Expression
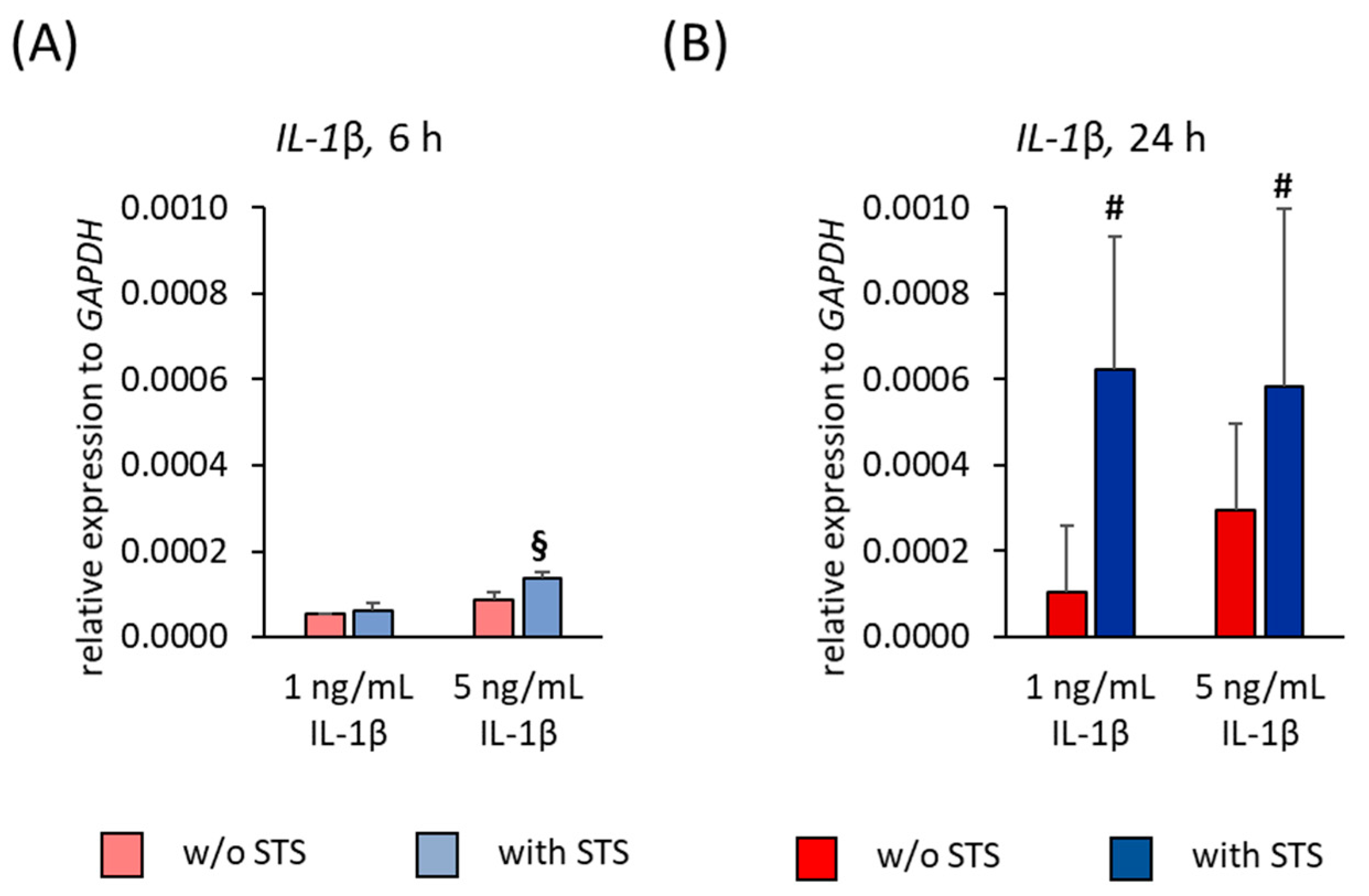
2.6. IL-1β Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Stimulation Protocol
4.3. Cell Viability
4.4. Quantitative Polymerase Chain Reaction
4.5. Immunoassay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | Extracellular matrix |
| OTM | Orthodontic tooth movement |
| PDL | Periodontal ligament |
| hPDL-MSCs | Human periodontal ligament-derived mesenchymal stromal cells |
| MMPs | Matrix metalloproteinases |
| GCF | Gingival crevicular fluid |
| TIMPs | Tissue inhibitor of matrix metalloproteinases |
| IL | Interleukin |
| STS | Static tensile strain |
| FBS | Fetal bovine serum |
| ELISA | Enzyme linked immunosorbent assay |
| S.E.M. | Standard error of the mean |
| DMEM | Dulbecco’s modified Eagles medium |
| ISCT | International Society for Cell and Gene Therapy |
| qPCR | Quantitative polymerase chain reaction |
| OD | Optical density |
| ANOVA | One-way analysis of variance |
References
- Wise, G.; King, G.J. Mechanisms of Tooth Eruption and Orthodontic Tooth Movement. J. Dent. Res. 2008, 87, 414–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.-C. Orthodontic tooth movement: The biology and clinical implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattaneo, P.M.; Dalstra, M.; Melsen, B. Strains in periodontal ligament and alveolar bone associated with orthodontic tooth movement analyzed by finite element. Orthod. Craniofac. Res. 2009, 12, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, P.; Dalstra, M.; Melsen, B. The Finite Element Method: A Tool to Study Orthodontic Tooth Movement. J. Dent. Res. 2005, 84, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Tantilertanant, Y.; Niyompanich, J.; Everts, V.; Supaphol, P.; Pavasant, P.; Sanchavanakit, N. Cyclic tensile force-upregulated IL6 increases MMP3 expression by human periodontal ligament cells. Arch. Oral Biol. 2019, 107, 104495. [Google Scholar] [CrossRef]
- Beertsen, W.; McCulloch, C.A.G.; Sodek, J. The periodontal ligament: A unique, multifunctional connective tissue. Periodontology 2000 1997, 13, 20–40. [Google Scholar] [CrossRef]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal Ligament Cells Under Mechanical Stress Induce Osteoclastogenesis by Receptor Activator of Nuclear Factor κB Ligand Up-Regulation via Prostaglandin E2 Synthesis. J. Bone Miner. Res. 2002, 17, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Kook, S.H.; Jang, Y.S.; Lee, J.C. Involvement of JNK-AP-1 and ERK-NF-κB signaling in tension-stimulated expression of Type i collagen and MMP-1 in human periodontal ligament fibroblasts. J. Appl. Physiol. 2011, 111, 1575–1586. [Google Scholar] [CrossRef] [Green Version]
- Rosselli-Murai, L.K.; Almeida, L.O.; Zagni, C.; Galindo-Moreno, P.; Padial-Molina, M.; Volk, S.L.; Murai, M.J.; Rios, H.F.; Squarize, C.H.; Castilho, R.M. Periostin Responds to Mechanical Stress and Tension by Activating the MTOR Signaling Pathway. PLoS ONE 2013, 8, e83580. [Google Scholar] [CrossRef]
- Yamashiro, K.; Myokai, F.; Hiratsuka, K.; Yamamoto, T.; Senoo, K.; Arai, H.; Nishimura, F.; Abiko, Y.; Takashiba, S. Oligonucleotide array analysis of cyclic tension-responsive genes in human periodontal ligament fibroblasts. Int. J. Biochem. Cell Biol. 2007, 39, 910–921. [Google Scholar] [CrossRef]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cellular Therapy Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J. Marrow Stromal Cells as Stem Cells for Nonhematopoietic Tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najar, M.; Bouhtit, F.; Melki, R.; Afif, H.; Hamal, A.; Fahmi, H.; Merimi, M.; Lagneaux, L. Mesenchymal Stromal Cell-Based Therapy: New Perspectives and Challenges. J. Clin. Med. 2019, 8, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Shi, S.; Gronthos, S. Perivascular Niche of Postnatal Mesenchymal Stem Cells in Human Bone Marrow and Dental Pulp. J. Bone Miner. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Yianni, V.; Sharpe, P. Perivascular-Derived Mesenchymal Stem Cells. J. Dent. Res. 2019, 98, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Yang, R.; Zhou, Y.H. Mechanobiology of periodontal ligament stem cells in orthodontic tooth movement. Stem Cells Int. 2018, 2018, 6531216. [Google Scholar] [CrossRef] [Green Version]
- Cantarella, G.; Cantarella, R.; Caltabiano, M.; Risuglia, N.; Bernardini, R.; Leonardi, R. Levels of matrix metalloproteinases 1 and 2 in human gingival crevicular fluid during initial tooth movement. Am. J. Orthod. Dentofac. Orthop. 2006, 130, 568.e11–568.e16. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page-McCaw, A.; Ewald, A.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhao, D.; Wu, Y.; Xu, C.; Zhang, F. Cyclic stretch induced gene expression of extracellular matrix and adhesion molecules in human periodontal ligament cells. Arch. Oral Biol. 2015, 60, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Huang, Y.; Song, W.; Gong, X.; Liu, M.; Jia, X.; Zhou, G.; Chen, L.; Li, A.; Fan, Y. Fluid shear stress regulates metalloproteinase-1 and 2 in human periodontal ligament cells: Involvement of extracellular signal-regulated kinase (ERK) and P38 signaling pathways. J. Biomech. 2012, 45, 2368–2375. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, C.; Walter, C.; Ziebart, T.; Grimm, S.; Meila, D.; Krieger, E.; Wehrbein, H. Induction of IL-6 and MMP-8 in human periodontal fibroblasts by static tensile strain. Clin. Oral Investig. 2013, 18, 901–908. [Google Scholar] [CrossRef]
- Narimiya, T.; Wada, S.; Kanzaki, H.; Ishikawa, M.; Tsuge, A.; Yamaguchi, Y.; Nakamura, Y. Orthodontic tensile strain induces angiogenesis via type IV collagen degradation by matrix metalloproteinase-12. J. Periodontal Res. 2017, 52, 842–852. [Google Scholar] [CrossRef]
- Saminathan, A.; Vinoth, K.J.; Wescott, D.C.; Pinkerton, M.N.; Milne, T.J.; Cao, T.; Meikle, M.C. The effect of cyclic mechanical strain on the expression of adhesion-related genes by periodontal ligament cells in two-dimensional culture. J. Periodontal Res. 2012, 47, 212–221. [Google Scholar] [CrossRef]
- Nemoto, T.; Kajiya, H.; Tsuzuki, T.; Takahashi, Y.; Okabe, K. Differential induction of collagens by mechanical stress in human periodontal ligament cells. Arch. Oral Biol. 2010, 55, 981–987. [Google Scholar] [CrossRef]
- Hacopian, N.; Nik, T.H.; Ghahremani, M.H.; Rahimi, H.R.; Ostad, S.N. Effects of continuous and interrupted forces on gene transcription in periodontal ligament cells in vitro. Acta Med. Iran. 2011, 49, 643–649. [Google Scholar]
- Bolcato-Bellemin, A.; Elkaim, R.; Abehsera, A.; Fausser, J.; Haïkel, Y.; Tenenbaum, H. Expression of mRNAs Encoding for ? and ? Integrin Subunits, MMPs, and TIMPs in Stretched Human Periodontal Ligament and Gingival Fibroblasts. J. Dent. Res. 2000, 79, 1712–1716. [Google Scholar] [CrossRef]
- Lisboa, R.A.; Lisboa, F.A.; Santos, G.D.C.; Andrade, M.; Cunha-Melo, J.R. Matrix metalloproteinase 2 activity decreases in human periodontal ligament fibroblast cultures submitted to simulated orthodontic force. In Vitro Cell. Dev. Biol. Anim. 2009, 45, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Lisboa, R.A.; Andrade, M.V.; Cunha-Melo, J.R. Zimography is an effective method for detection of matrix metalloproteinase 2 (MMP-2) activity in cultured human fibroblasts. Acta Cir. Bras. 2013, 28, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Garlet, T.P.; Coelho, U.; Silva, J.S.; Garlet, G.P. Cytokine expression pattern in compression and tension sides of the periodontal ligament during orthodontic tooth movement in humans. Eur. J. Oral Sci. 2007, 115, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Bildt, M.M.; Bloemen, M.; Kuijpers-Jagtman, A.M.; Hoff, J.W.V.D. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in gingival crevicular fluid during orthodontic tooth movement. Eur. J. Orthod. 2009, 31, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Yang, L.; Zheng, W.; Lin, T. MicroRNA-34 expression in gingival crevicular fluid correlated with orthodontic tooth movement. Angle Orthod. 2020, 90, 702–706. [Google Scholar] [CrossRef] [Green Version]
- Grimm, S.; Wolff, E.; Walter, C.; Pabst, A.M.; Mundethu, A.; Jacobs, C.; Wehrbein, H.; Jacobs, C. Influence of clodronate and compressive force on IL-1ß-stimulated human periodontal ligament fibroblasts. Clin. Oral Investig. 2019, 24, 343–350. [Google Scholar] [CrossRef]
- Nettelhoff, L.; Grimm, S.; Jacobs, C.; Walter, C.; Pabst, A.M.; Goldschmitt, J.; Wehrbein, H. Influence of mechanical compression on human periodontal ligament fibroblasts and osteoblasts. Clin. Oral Investig. 2015, 20, 621–629. [Google Scholar] [CrossRef]
- Grant, M.; Wilson, J.; Rock, P.; Chapple, I. Induction of cytokines, MMP9, TIMPs, RANKL and OPG during orthodontic tooth movement. Eur. J. Orthod. 2013, 35, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Dudic, A.; Kiliaridis, S.; Mombelli, A.; Giannopoulou, C. Composition changes in gingival crevicular fluid during orthodontic tooth movement: Comparisons between tension and compression sides. Eur. J. Oral Sci. 2006, 114, 416–422. [Google Scholar] [CrossRef]
- Jiang, C.; Li, Z.; Quan, H.; Xiao, L.; Zhao, J.; Wang, Y.; Liu, J.; Gou, Y.; An, S.; Huang, Y.; et al. Osteoimmunology in orthodontic tooth movement. Oral Dis. 2015, 21, 694–704. [Google Scholar] [CrossRef]
- Xiang, J.; Li, C.; Dong, W.; Cao, Z.; Liu, L. Expression of matrix metalloproteinase-1, matrix metalloproteinase-2 and extracellular metalloproteinase inducer in human periodontal ligament cells stimulated with interleukin-1beta. J. Periodontal Res. 2009, 44, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Murayama, R.; Kobayashi, M.; Takeshita, A.; Yasui, T.; Yamamoto, M. MAPKs, activator protein-1 and nuclear factor-κB mediate production of interleukin-1β-stimulated cytokines, prostaglandinE 2 and MMP-1 in human periodontal ligament cells. J. Periodontal Res. 2011, 46, 568–575. [Google Scholar] [PubMed]
- Long, P.; Liu, F.; Piesco, N.P.; Kapur, R.; Agarwal, S. Signaling by mechanical strain involves transcriptional regulation of proinflammatory genes in human periodontal ligament cells in vitro. Bone 2002, 30, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Isola, G.; Giudice, A.L.; Giudice, A.L.; Alibrandi, A.; Murabito, P.; Indelicato, F. Identification of the different salivary Interleukin-6 profiles in patients with periodontitis: A cross-sectional study. Arch. Oral Biol. 2021, 122, 104997. [Google Scholar] [CrossRef] [PubMed]
- Engebretson, S.P.; Hey-Hadavi, J.; Ehrhardt, F.J.; Hsu, D.; Celenti, R.S.; Grbic, J.T.; Lamster, I.B. Gingival Crevicular Fluid Levels of Interleukin-1β and Glycemic Control in Patients With Chronic Periodontitis and Type 2 Diabetes. J. Periodontol. 2004, 75, 1203–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, C.; Schramm, S.; Dirks, I.; Walter, C.; Pabst, A.; Meila, D.; Jacobs, C.; Wehrbein, H. Mechanical loading increases pro-inflammatory effects of nitrogen-containing bisphosphonate in human periodontal fibroblasts. Clin. Oral Investig. 2017, 22, 901–907. [Google Scholar] [CrossRef]
- Nokhbehsaim, M.; Deschner, B.; Winter, J.; Bourauel, C.; Jäger, A.; Jepsen, S.; Deschner, J. Anti-inflammatory effects of EMD in the presence of biomechanical loading and interleukin-1β in vitro. Clin. Oral Investig. 2011, 16, 275–283. [Google Scholar] [CrossRef]
- Brauchli, L.; Senn, C.; Ball, J.; Wichelhaus, A. Force levels of 23 nickel-titanium open-coil springs in compression testing. Am. J. Orthod. Dentofac. Orthop. 2011, 139, 601–605. [Google Scholar] [CrossRef]
- Rozman, J.; Mrvar, P.; Drevensek, M.; Pečlin, P. Evaluation of NiTi Superelastic Retraction Coil Springs for orthodontic tooth movement in rats. Bio-Med Mater. Eng. 2010, 20, 339–348. [Google Scholar] [CrossRef]
- Murabayashi, D.; Mochizuki, M.; Tamaki, Y.; Nakahara, T. Practical methods for handling human periodontal ligament stem cells in serum-free and serum-containing culture conditions under hypoxia: Implications for regenerative medicine. Hum. Cell 2017, 30, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Spitz, A.; Christovam, I.O.; Marañón-Vásquez, G.A.; Ferreira, D.M.T.P.; Adesse, D.; Maia, L.C.; Bolognese, A.M. Global gene expression profile of periodontal ligament cells submitted to mechanical loading: A systematic review. Arch. Oral Biol. 2020, 118, 104884. [Google Scholar] [CrossRef]
- Li, M.; Zhang, C.; Yang, Y. Effects of mechanical forces on osteogenesis and osteoclastogenesis in human periodontal ligament fibroblasts: A systematic review of in vitro studies. Bone Jt. Res. 2019, 8, 19–31. [Google Scholar] [CrossRef] [PubMed]
- VanSant, L.; De Llano-Pérula, M.C.; Verdonck, A.; Willems, G. Expression of biological mediators during orthodontic tooth movement: A systematic review. Arch. Oral Biol. 2018, 95, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Carano, A.; Siciliani, G. Effects of continuous and intermittent forces on human fibroblasts in vitro. Eur. J. Orthod. 1996, 18, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, P.; Hu, J.; Piesco, N.; Buckley, M.; Agarwal, S. Low Magnitude of Tensile Strain Inhibits IL-1β-dependent Induction of Pro-inflammatory Cytokines and Induces Synthesis of IL-10 in Human Periodontal Ligament Cells in vitro. J. Dent. Res. 2001, 80, 1416–1420. [Google Scholar] [CrossRef]
- Tsuji, K.; Uno, K.; Zhang, G.X.; Tamura, M. Periodontal ligament cells under intermittent tensile stress regulate mRNA expression of osteoprotegerin and tissue inhibitor of matrix metalloprotease-1 and -2. J. Bone Miner. Metab. 2004, 22, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Kawamoto, K.; Okamoto, H.; Horiuchi, N. Induction of collagenase-2 (matrix metalloproteinase-8) gene expression by interleukin-1β in human gingival fibroblasts. J. Periodontal Res. 2001, 36, 153–159. [Google Scholar] [CrossRef]
- Birkedal-Hansen, H.; Moore, W.; Bodden, M.; Windsor, L.; Decarlo, A.; Engler, J. Matrix Metalloproteinases: A Review. Crit. Rev. Oral Biol. Med. 1993, 4, 197–250. [Google Scholar] [CrossRef] [Green Version]
- Proff, P.; Reicheneder, C.; Faltermeier, A.; Kubein-Meesenburg, D.; Römer, P. Effects of mechanical and bacterial stressors on cytokine and growth-factor expression in periodontal ligament cells. Fortschr. Kieferorthop. 2014, 75, 191–202. [Google Scholar] [CrossRef]
- Hoff, J.W.V.D. Effects of mechanical tension on matrix degradation by human periodontal ligament cells cultured in collagen gels. J. Periodontal Res. 2003, 38, 449–457. [Google Scholar] [CrossRef]
- Santamaria, M.; Bagne, L.; Zaniboni, E.; Santamaria, M.P.; Jardini, M.A.N.; Felonato, M.; Dos Santos, G.M.T.; Mendonça, F.A.S.; Esquisatto, M.A.M. Diabetes mellitus and periodontitis: Inflammatory response in orthodontic tooth movement. Orthod. Craniofac. Res. 2019, 23, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Vanarsdall, R.L. Orthodontics and periodontal therapy. Periodontology 2000 1995, 9, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after Carl Sandstedt. Eur. J. Orthod. 2005, 28, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Yucel-Lindberg, T.; Båge, T. Inflammatory mediators in the pathogenesis of periodontitis. Expert Rev. Mol. Med. 2013, 15, e7. [Google Scholar] [CrossRef] [Green Version]
- Isola, G.; Polizzi, A.; Patini, R.; Ferlito, S.; Alibrandi, A.; Palazzo, G. Association among serum and salivary A. actinomycetemcomitans specific immunoglobulin antibodies and periodontitis. BMC Oral Health 2020, 20, 283. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef]
- Behm, C.; Blufstein, A.; Gahn, J.; Nemec, M.; Moritz, A.; Rausch-Fan, X.; Andrukohv, O. Cytokines Differently Define the Immunomodulation of Mesenchymal Stem Cells from the Periodontal Ligament. Cells 2020, 9, 1222. [Google Scholar] [CrossRef]
- Behm, C.; Blufstein, A.; Abhari, S.Y.; Koch, C.; Gahn, J.; Schäffer, C.; Moritz, A.; Rausch-Fan, X.; Andrukhov, O. Response of Human Mesenchymal Stromal Cells from Periodontal Tissue to LPS Depends on the Purity but Not on the LPS Source. Mediat. Inflamm. 2020, 2020, 1–17. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behm, C.; Nemec, M.; Blufstein, A.; Schubert, M.; Rausch-Fan, X.; Andrukhov, O.; Jonke, E. Interleukin-1β Induced Matrix Metalloproteinase Expression in Human Periodontal Ligament-Derived Mesenchymal Stromal Cells under In Vitro Simulated Static Orthodontic Forces. Int. J. Mol. Sci. 2021, 22, 1027. https://doi.org/10.3390/ijms22031027
Behm C, Nemec M, Blufstein A, Schubert M, Rausch-Fan X, Andrukhov O, Jonke E. Interleukin-1β Induced Matrix Metalloproteinase Expression in Human Periodontal Ligament-Derived Mesenchymal Stromal Cells under In Vitro Simulated Static Orthodontic Forces. International Journal of Molecular Sciences. 2021; 22(3):1027. https://doi.org/10.3390/ijms22031027
Chicago/Turabian StyleBehm, Christian, Michael Nemec, Alice Blufstein, Maria Schubert, Xiaohui Rausch-Fan, Oleh Andrukhov, and Erwin Jonke. 2021. "Interleukin-1β Induced Matrix Metalloproteinase Expression in Human Periodontal Ligament-Derived Mesenchymal Stromal Cells under In Vitro Simulated Static Orthodontic Forces" International Journal of Molecular Sciences 22, no. 3: 1027. https://doi.org/10.3390/ijms22031027
APA StyleBehm, C., Nemec, M., Blufstein, A., Schubert, M., Rausch-Fan, X., Andrukhov, O., & Jonke, E. (2021). Interleukin-1β Induced Matrix Metalloproteinase Expression in Human Periodontal Ligament-Derived Mesenchymal Stromal Cells under In Vitro Simulated Static Orthodontic Forces. International Journal of Molecular Sciences, 22(3), 1027. https://doi.org/10.3390/ijms22031027