
Effect of O-Antigen Chain Length Regulation on the Immunogenicity of Shigella and Salmonella Generalized Modules for Membrane Antigens (GMMA)


 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. OAg Length Modulation through Genetic Engineering and Characterization of the Resulting GMMA
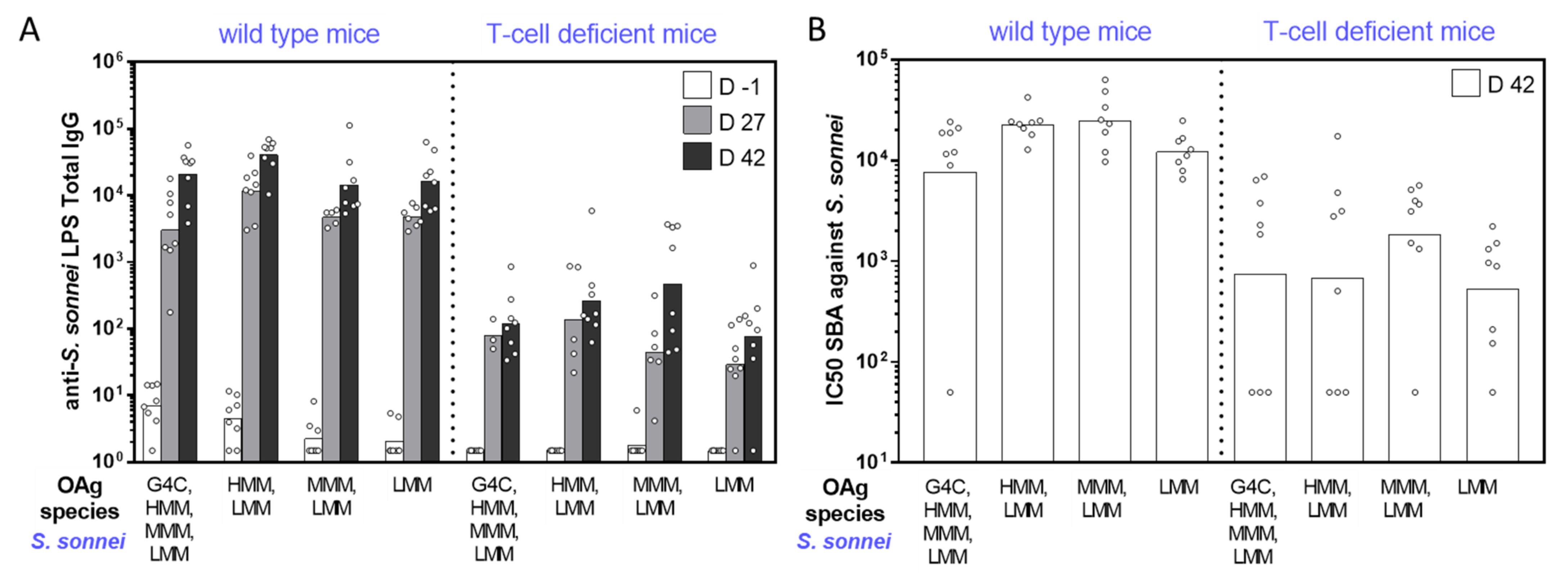
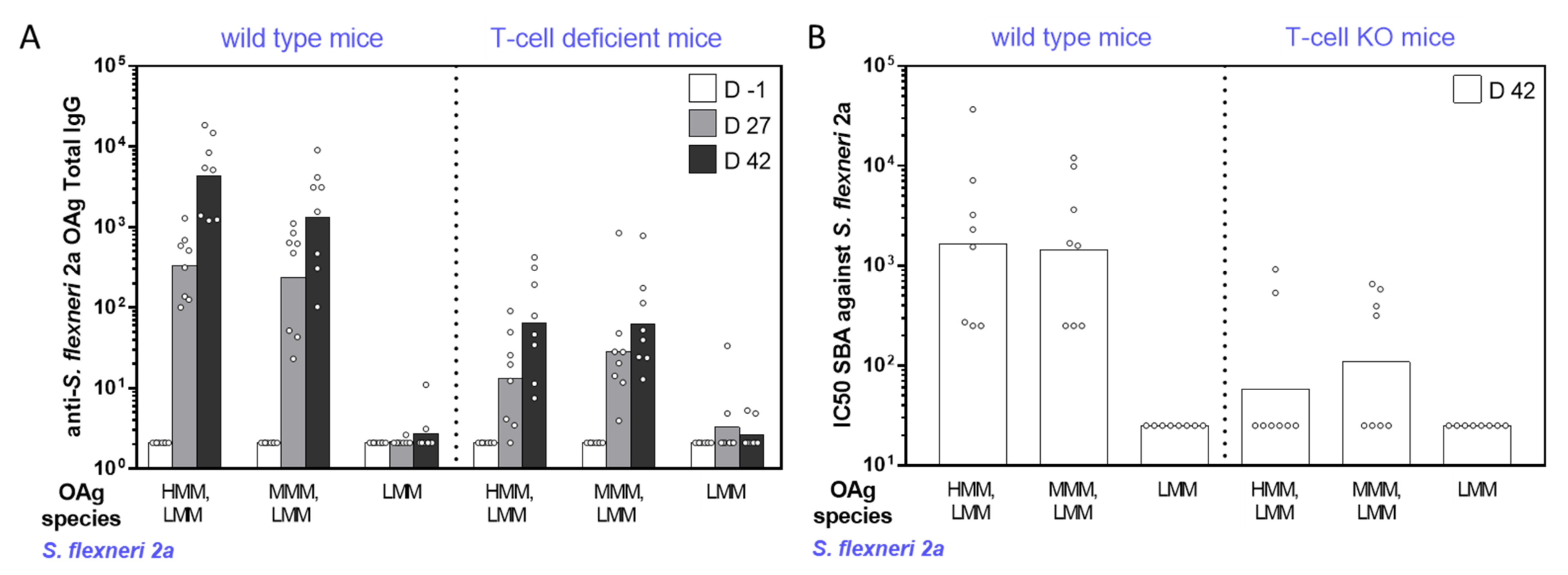
2.2. Immunogenicity Studies in Mice with GMMA Differing for OAg Length
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Generation of Mutants
4.2. GMMA Production and Characterization
4.3. Mouse Studies
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, D.; Meron-Sudai, S.; Bialik, A.; Asato, V.; Goren, S.; Ariel-Cohen, O.; Reizis, A.; Hochberg, A.; Ashkenazi, S. Serum IgG antibodies to Shigella lipopolysaccharide antigens—A correlate of protection against shigellosis. Hum. Vaccines Immunother. 2019, 15, 1401–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berti, F.; Micoli, F. Improving efficacy of glycoconjugate vaccines: From chemical conjugates to next generation constructs. Curr. Opin. Immunol. 2020, 65, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Rossi, O.; Necchi, F.; Micoli, F. OMV Vaccines and the Role of TLR Agonists in Immune Response. Int. J. Mol. Sci. 2020, 21, 4416. [Google Scholar] [CrossRef]
- Raso, M.M.; Gasperini, G.; Alfini, R.; Schiavo, F.; Aruta, M.G.; Carducci, M.; Forgione, M.C.; Martini, S.; Cescutti, P.; Necchi, F.; et al. GMMA and Glycoconjugate Approaches Compared in Mice for the Development of a Vaccine against Shigella flexneri Serotype 6. Vaccines 2020, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative immunogenicity and efficacy of equivalent outer membrane vesicle and glycoconjugate vaccines against nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef] [Green Version]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Costantino, P.; Rappuoli, R.; Berti, F. The design of semi-synthetic and synthetic glycoconjugate vaccines. Expert Opin. Drug Discov. 2011, 6, 1045–1066. [Google Scholar] [CrossRef]
- Robbins, J.B.; Kubler-Kielb, J.; Vinogradov, E.; Mocca, C.; Pozsgay, V.; Shiloach, J.; Schneerson, R. Synthesis, characterization, and immunogenicity in mice of Shigella sonnei O-specific oligosaccharide-core-protein conjugates. Proc. Natl. Acad. Sci. USA 2009, 106, 7974–7978. [Google Scholar] [CrossRef] [Green Version]
- Phalipon, A.; Tanguy, M.; Grandjean, C.; Guerreiro, C.; Bélot, F.; Cohen, D.; Sansonetti, P.J.; Mulard, L.A. A Synthetic Carbohydrate-Protein Conjugate Vaccine Candidate againstShigella flexneri2a Infection. J. Immunol. 2009, 182, 2241–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jörbeck, H.J.; Svenson, S.B.; Lindberg, A.A. Artificial Salmonella vaccines: Salmonella typhimurium O-antigen-specific oligosaccharide-protein conjugates elicit opsonizing antibodies that enhance phagocytosis. Infect. Immun. 1981, 32, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Obaro, S.K. The new pneumococcal vaccine. Clin. Microbiol. Infect. 2002, 8, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parke, J.C. Capsular polysaccharide of Haemophilus influenzae type b as a vaccine. Pediatr. Infect. Dis. J. 1987, 6, 795–798. [Google Scholar] [CrossRef]
- Blanchard-Rohner, G.; Pollard, A.J. Long-term protection after immunization with protein–polysaccharide conjugate vaccines in infancy. Expert Rev. Vaccines 2011, 10, 673–684. [Google Scholar] [CrossRef]
- Mitchell, R.; Kelly, D.F.; Pollard, A.J.; Trück, J. Polysaccharide-specific B cell responses to vaccination in humans. Hum. Vaccines Immunother. 2014, 10, 1661–1668. [Google Scholar] [CrossRef]
- Rappuoli, R. Glycoconjugate vaccines: Principles and mechanisms. Sci. Transl. Med. 2018, 10, eaat4615. [Google Scholar] [CrossRef]
- Micoli, F.; Bjarnarson, S.P.; Arcuri, M.; Pind, A.A.A.; Magnusdottir, G.J.; Necchi, F.; Di Benedetto, R.; Carducci, M.; Schiavo, F.; Giannelli, C.; et al. Short Vi-polysaccharide abrogates T-independent immune response and hyporesponsiveness elicited by long Vi-CRM197 conjugate vaccine. Proc. Natl. Acad. Sci. USA 2020, 117, 24443–24449. [Google Scholar] [CrossRef]
- Whitfield, C.; Williams, D.M.; Kelly, S.D. Lipopolysaccharide O-antigens—bacterial glycans made to measure. J. Biol. Chem. 2020, 295, 10593–10609. [Google Scholar] [CrossRef]
- Caboni, M.; Pédron, T.; Rossi, O.; Goulding, D.; Pickard, D.; Citiulo, F.; MacLennan, C.A.; Dougan, G.; Thomson, N.R.; Saul, A.; et al. An O Antigen Capsule Modulates Bacterial Pathogenesis in Shigella sonnei. PLoS Pathog. 2015, 11, e1004749. [Google Scholar] [CrossRef]
- Hoare, A.; Bravo, D.; Martinić, M.; Valvano, M.A.; Contreras, I.; Álvarez, S.A. The normal chain length distribution of the O antigen is required for the interaction of Shigella flexneri 2a with polarized Caco-2 cells. Biol. Res. 2012, 45, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Hölzer, S.U.; Schlumberger, M.C.; Jäckel, D.; Hensel, M. Effect of the O-Antigen Length of Lipopolysaccharide on the Functions of Type III Secretion Systems in Salmonella enterica. Infect. Immun. 2009, 77, 5458–5470. [Google Scholar] [CrossRef] [Green Version]
- Rana, R.; Dalal, J.; Singh, D.; Kumar, N.; Hanif, S.; Joshi, N.; Chhikara, M. Development and characterization of Haemophilus influenzae type b conjugate vaccine prepared using different polysaccharide chain lengths. Vaccine 2015, 33, 2646–2654. [Google Scholar] [CrossRef]
- Pozsgay, V.; Chu, C.; Pannell, L.; Wolfe, J.; Robbins, J.B.; Schneerson, R. Protein conjugates of synthetic saccharides elicit higher levels of serum IgG lipopolysaccharide antibodies in mice than do those of the O-specific polysaccharide from Shi-gella dysenteriae type 1. Proc. Natl. Acad. Sci. USA 1999, 96, 5194–5197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Normand, B.V.-L.; Saul, F.A.; Phalipon, A.; Belot, F.; Guerreiro, C.; Mulard, L.A.; Bentley, G.A. Structures of synthetic O-antigen fragments from serotype 2a Shigella flexneri in complex with a protective monoclonal antibody. Proc. Natl. Acad. Sci. USA 2008, 105, 9976–9981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubler-Kielb, J.; Vinogradov, E.; Mocca, C.; Pozsgay, V.; Coxon, B.; Robbins, J.B.; Schneerson, R. Immunochemical studies of Shigella flexneri 2a and 6, and Shigella dysenteriae type 1 O-specific polysaccharide-core fragments and their protein conjugates as vaccine candidates. Carbohydr. Res. 2010, 345, 1600–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesinski, G.B.; Westerink, M.A.J. Novel vaccine strategies to T-independent antigens. J. Microbiol. Methods 2001, 47, 135–149. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Launay, O.; Ndiaye, A.G.W.; Conti, V.; Loulergue, P.; Sciré, A.S.; Landre, A.M.; Ferruzzi, P.; Nedjaai, N.; Schütte, L.D.; Auerbach, J.; et al. Booster Vaccination With GVGH Shigella sonnei 1790GAHB GMMA Vaccine Compared to Single Vaccination in Unvaccinated Healthy European Adults: Results From a Phase 1 Clinical Trial. Front. Immunol. 2019, 10, 335. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Del Bino, L.; Alfini, R.; Carboni, F.; Romano, M.R.; Adamo, R. Glycoconjugate vaccines: Current approaches towards faster vaccine design. Expert Rev. Vaccines 2019, 18, 881–895. [Google Scholar] [CrossRef]
- Meloni, E.; Colucci, A.M.; Micoli, F.; Sollai, L.; Gavini, M.; Saul, A.; Di Cioccio, V.; MacLennan, C.A. Simplified low-cost production of O-antigen from Salmonella Typhimurium Generalized Modules for Membrane Antigens (GMMA). J. Biotechnol. 2015, 198, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegerle, N.; Bose, J.; Ramachandran, G.; Galen, J.E.; Levine, M.M.; Simon, R.; Tennant, S.M. Overexpression of O-polysaccharide chain length regulators in Gram-negative bacteria using the Wzx-/Wzy-dependent pathway enhances production of defined modal length O-polysaccharide polymers for use as haptens in glycoconjugate vaccines. J. Appl. Microbiol. 2018, 125, 575–585. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Ravenscroft, N.; Cescutti, P.; Stefanetti, G.; Londero, S.; Rondini, S.; MacLennan, C.A. Structural analysis of O-polysaccharide chains extracted from different Salmonella Typhimurium strains. Carbohydr. Res. 2014, 385, 1–8. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, G.; Alfini, R.; Cescutti, P.; Caboni, M.; Lanzilao, L.; Necchi, F.; Saul, A.; MacLennan, C.; Rondini, S.; Micoli, F. Characterization of O-antigen delivered by Generalized Modules for Membrane Antigens (GMMA) vaccine candidates against nontyphoidal Salmonella. Vaccine 2017, 35, 419–426. [Google Scholar] [CrossRef]
- Giannelli, C.; Raso, M.M.; Palmieri, E.; De Felice, A.; Pippi, F.; Micoli, F. Development of a Specific and Sensitive HPAEC-PAD Method for Quantification of Vi Polysaccharide Applicable to other Polysaccharides Containing Amino Uronic Acids. Anal. Chem. 2020, 92, 6304–6311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedetto, G.; Cescutti, P.; Giannelli, C.; Rizzo, R.; Micoli, F. Multiple Techniques for Size Determination of Generalized Modules for Membrane Antigens from Salmonella typhimurium and Salmonella enteritidis. ACS Omega 2017, 2, 8282–8289. [Google Scholar] [CrossRef]
- Rossi, O.; Pesce, I.; Giannelli, C.; Aprea, S.; Caboni, M.; Citiulo, F.; Valentini, S.; Ferlenghi, I.; MacLennan, C.A.; D’Oro, U.; et al. Modulation of endotoxicity of Shigella generalized modules for membrane antigens (GMMA) by genetic lipid A modifications: Relative activation of TLR4 and TLR2 pathways in different mutants. J. Biol. Chem. 2014, 289, 24922–24935. [Google Scholar] [CrossRef] [Green Version]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef]
- Lanzilao, L.; Stefanetti, G.; Saul, A.; MacLennan, C.A.; Micoli, F.; Rondini, S. Strain Selection for Generation of O-Antigen-Based Glycoconjugate Vaccines against Invasive Nontyphoidal Salmonella Disease. PLoS ONE 2015, 10, e0139847. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Rondini, S.; Gavini, M.; Pisoni, I.; Lanzilao, L.; Colucci, A.; Giannelli, C.; Pippi, F.; Sollai, L.; Pinto, V.; et al. A scalable method for O-antigen purification applied to various Salmonella serovars. Anal. Biochem. 2013, 434, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Necchi, F.; Saul, A.; Rondini, S. Development of a high-throughput method to evaluate serum bactericidal activity using bacterial ATP measurement as survival readout. PLoS ONE 2017, 12, e0172163. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Deleted | Gene Function | Resulting Phenotype |
|---|---|---|
| ept-etk | Part of the G4C operon needed for capsule assembly and export | Lack of G4C formation |
| wzz (pSS) | pSS plasmid-borne OAg co-polymerase, responsible for HMM OAg polymerization in S. sonnei | Lack of HMM OAg formation |
| fepE (pHS2) | pHS2 plasmid-borne OAg co-polymerase, responsible for HMM OAg polymerization in S. flexneri | Lack of HMM OAg formation |
| fepE | Chromosomal encoded OAg co-polymerase, responsible for HMM OAg polymerization in S. Typhimurium | Lack of HMM OAg formation |
| wzzB | Chromosomal encoded OAg co-polymerase, responsible for MMM OAg polymerization in S. sonnei, S. flexneri and S. Typhimurium | Lack of MMM OAg formation in S. sonnei and S. Typhimurium; De-regulation of OAg polymerization in S. flenxeri 2a |
| wzy | OAg polymerase, responsible for the polymerization of the repeating units in S. sonnei and S. flexneri | Formation of OAg composed by 1 single repeating unit |
| GMMA | Mutations | OAg Length | Total Sugar/Protein w/w Ratio | Size 2 × Rw nm |
|---|---|---|---|---|
| S. sonnei | ΔtolR | G4C, HMM, MMM, LMM OAg | 0.34 | 87.0 |
| ΔtolR ΔG4C ΔwzzB | HMM, LMM OAg | 0.35 | 90.3 | |
| ΔtolR ΔG4C Δwzz | MMM, LMM OAg | 0.18 | 91.4 | |
| ΔtolR ΔG4C Δwzz ΔwzzB | LMM OAg | 0.14 | 90.8 | |
| S. flexneri 2a | ΔtolR | HMM, MMM, LMM OAg | 0.58 | 89.6 |
| pHS2-cured ΔtolR | MMM, LMM OAg | 0.53 | 91.4 | |
| pHS2-cured ΔtolR ΔwzzB | De-regulated OAg | 0.40 | 93.6 | |
| pHS2-cured ΔtolR ΔwzzB Δwzy pACYC-wzy | LMM OAg | 0.17 | 81.0 | |
| S. Typhimurium | ΔtolR | HMM, MMM, LMM OAg | 1.35 | 64.4 |
| ΔtolR ΔfepE | MMM, LMM OAg | 1.42 | 66.2 | |
| ΔtolR ΔfepE ΔwzzB | LMM OAg | 0.44 | 62.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gasperini, G.; Raso, M.M.; Arato, V.; Aruta, M.G.; Cescutti, P.; Necchi, F.; Micoli, F. Effect of O-Antigen Chain Length Regulation on the Immunogenicity of Shigella and Salmonella Generalized Modules for Membrane Antigens (GMMA). Int. J. Mol. Sci. 2021, 22, 1309. https://doi.org/10.3390/ijms22031309
Gasperini G, Raso MM, Arato V, Aruta MG, Cescutti P, Necchi F, Micoli F. Effect of O-Antigen Chain Length Regulation on the Immunogenicity of Shigella and Salmonella Generalized Modules for Membrane Antigens (GMMA). International Journal of Molecular Sciences. 2021; 22(3):1309. https://doi.org/10.3390/ijms22031309
Chicago/Turabian StyleGasperini, Gianmarco, Maria Michelina Raso, Vanessa Arato, Maria Grazia Aruta, Paola Cescutti, Francesca Necchi, and Francesca Micoli. 2021. "Effect of O-Antigen Chain Length Regulation on the Immunogenicity of Shigella and Salmonella Generalized Modules for Membrane Antigens (GMMA)" International Journal of Molecular Sciences 22, no. 3: 1309. https://doi.org/10.3390/ijms22031309
APA StyleGasperini, G., Raso, M. M., Arato, V., Aruta, M. G., Cescutti, P., Necchi, F., & Micoli, F. (2021). Effect of O-Antigen Chain Length Regulation on the Immunogenicity of Shigella and Salmonella Generalized Modules for Membrane Antigens (GMMA). International Journal of Molecular Sciences, 22(3), 1309. https://doi.org/10.3390/ijms22031309