
Possibility of Japanese Cedar Pollen Causing False Positives in the Deep Mycosis Test

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Comparison of the Specificities of supBGRP and mDectin-1
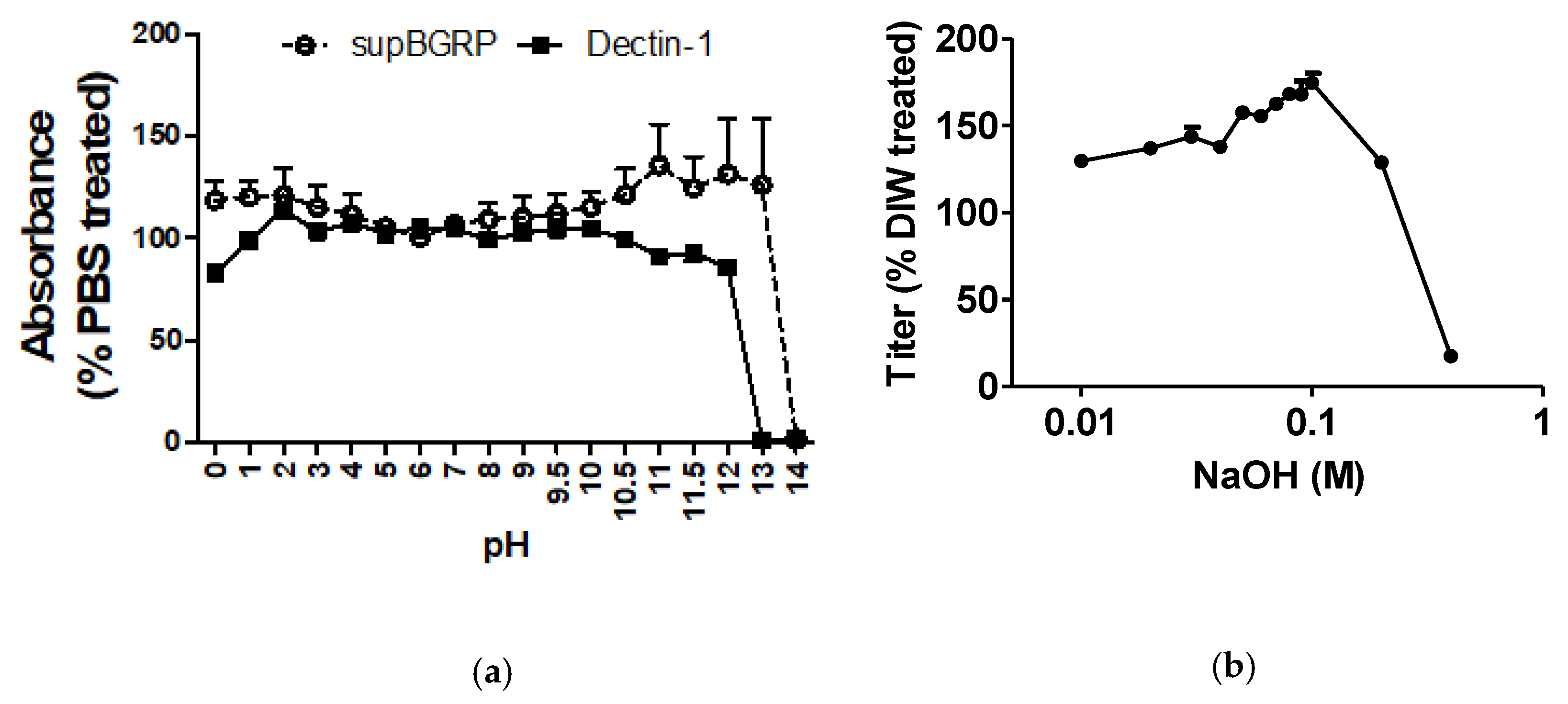
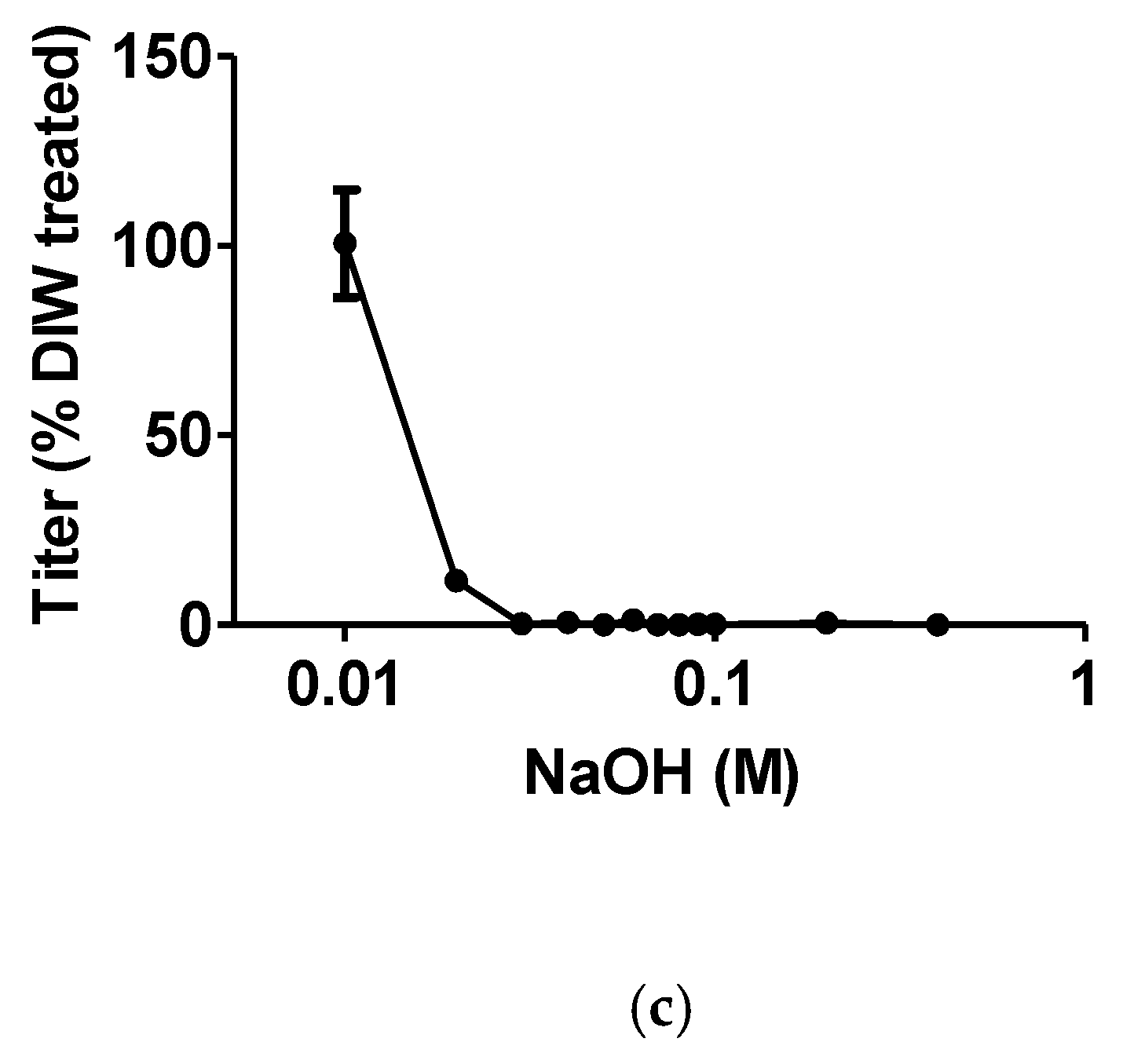
2.2. Effect of pH on the Binding Stability of supBGRP
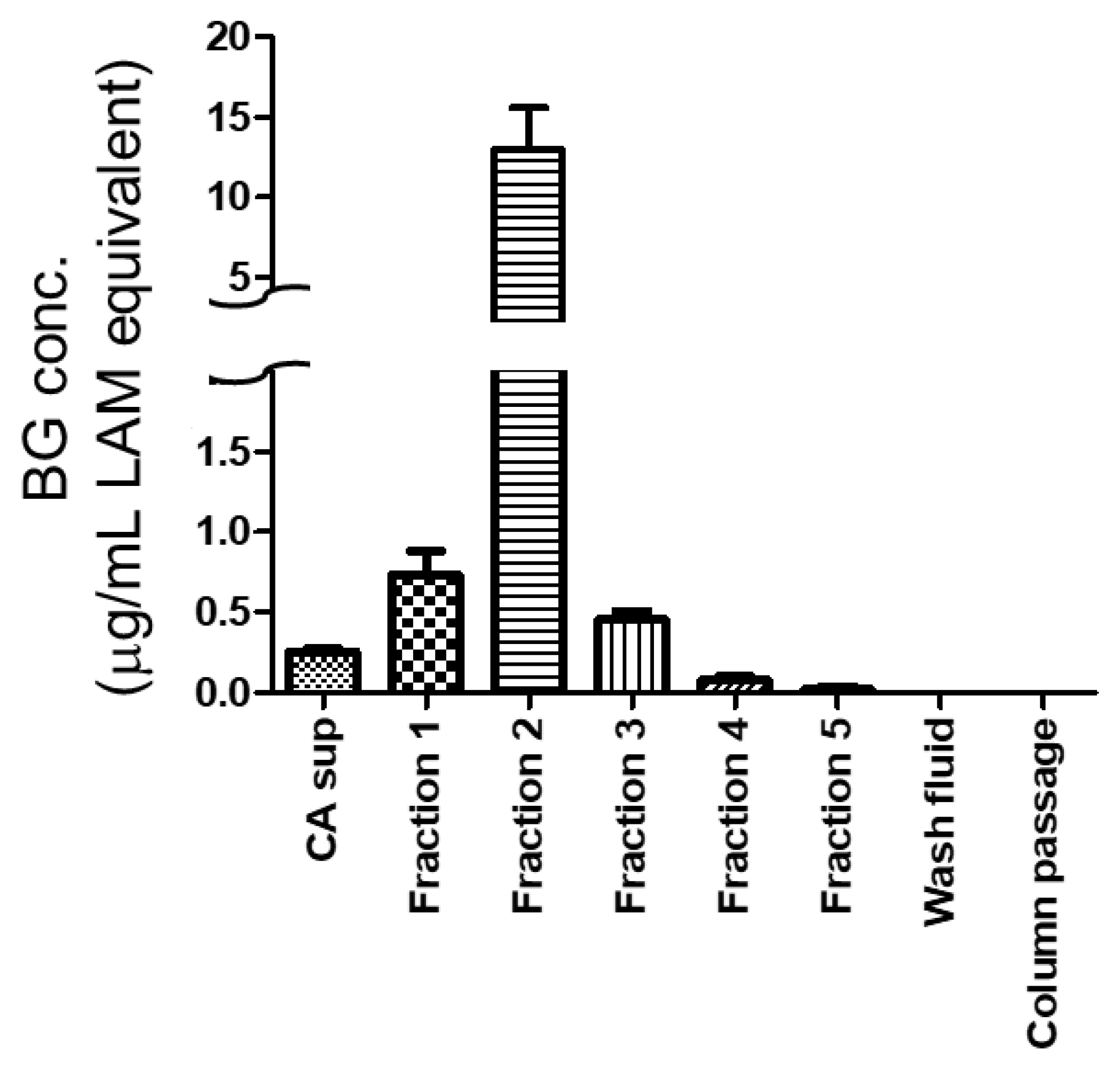
2.3. Column Concentration of Beta-1,3-d-Glucans
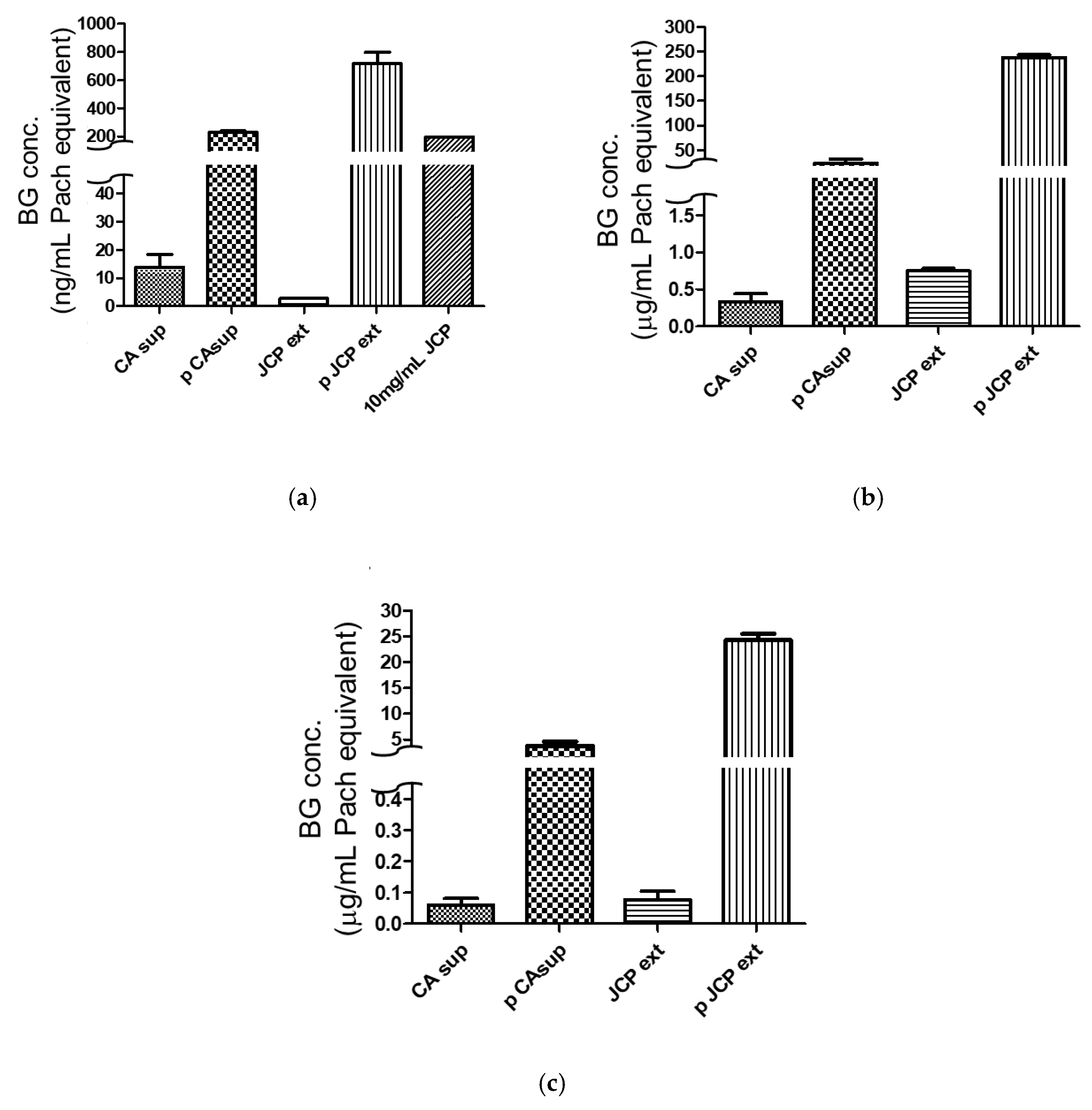
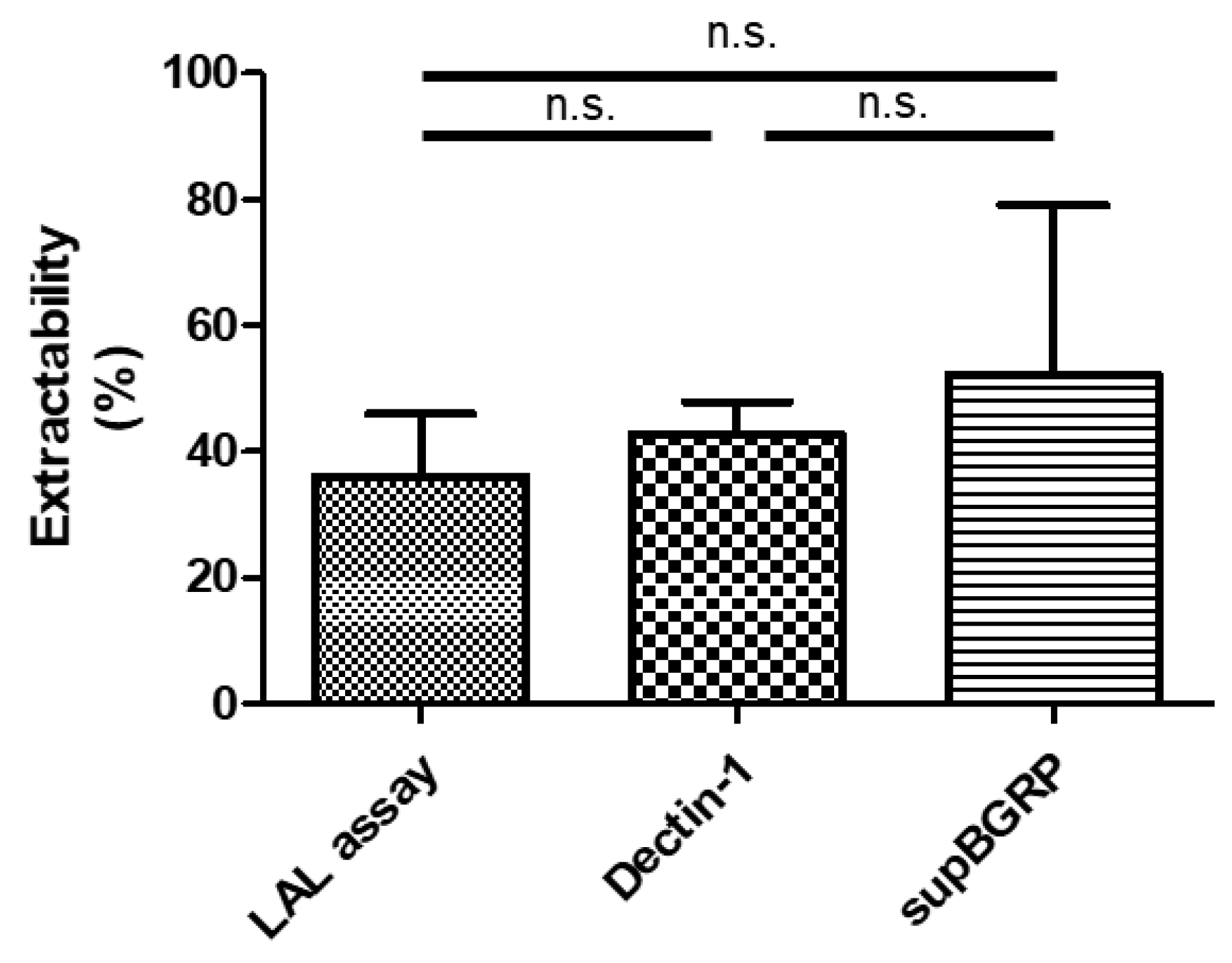
2.4. Comparison of Different Beta-Glucan Quantitative Methods
3. Discussion
4. Materials and Methods
4.1. Polysaccharide Reagents
4.2. β-Glucan Quantitative Assays
4.3. Comparing the Specificity and Binding Stability of supBGRP and Dectin-1
4.4. Sample Preparation
4.5. Beta-Glucan Recognition Protein Column Purification
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APBG | Beta-glucan from Aureobasidium pullulans |
| AP-FBG | Fermented beta-glucan from Aureobasidium pullulans |
| Bal | Barley beta-glucan |
| BG | Beta-1,3-d-glucan |
| BGRP | Beta-glucan recognition protein |
| CA | Candida albicans |
| CA sup | Candida albicans culture supernatant |
| CSBG | Candida solubilized beta-glucan |
| CMC | Carboxymethylcellulose |
| Curd | Curdlan |
| Dex | Dextran |
| DIW | Deionized water |
| EIA | Enzyme immunoassay |
| ext | Extract |
| JCP | Japanese cedar pollen |
| LAL | Limulus amebocyte lysate |
| LAM | Laminarin |
| LNT | Lentinan |
| Man | Mannan from Saccharomyces cerevisiae |
| mDectin-1 | Mouse Dectin-1 |
| Pach | Pachyman |
| Para | Paramylon |
| PBS | Phosphate-buffered saline |
| PEG | Polyethylene glycol |
| Pul | Pullulan |
| Pus | Pustulan |
| SCL | Scleroglucan |
| SPG | Schizophyllan |
| supBGRP | Artificial beta-glucan recognition protein |
| Xyl | Xylan from corn core |
References
- Taylor, P.R.; Tsoni, S.V.; Willment, J.A.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 is required for beta-glucan recognition and control of fungal infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Immune recognition. A new receptor for beta-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Reyes, C.N.; Becker, C.A.; Katsumoto, T.R.; Ma, J.; Wolf, A.J.; Bose, N.; Chan, A.S.; Magee, A.S.; Danielson, M.E.; et al. Activation of the innate immune receptor Dectin-1 upon formation of a ‘phagocytic synapse’. Nature 2011, 472, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Rossnagle, E.; Lowell, C.A.; Simmons, R.M. Dectin-1 activates Syk tyrosine kinase in a dynamic subset of macrophages for reactive oxygen production. Blood 2005, 106, 2543–2550. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Kamiya, T.; Liu, Y.; Kadoki, M.; Kakuta, S.; Oshima, K.; Hattori, M.; Takeshita, K.; Kanai, T.; Saijo, S.; et al. Inhibition of Dectin-1 Signaling Ameliorates Colitis by Inducing Lactobacillus-Mediated Regulatory T Cell Expansion in the Intestine. Cell Host Microbe 2015, 18, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahasi, K.; Ochiai, M.; Horiuchi, M.; Kumeta, H.; Ogura, K.; Ashida, M.; Inagaki, F. Solution structure of the silkworm betaGRP/GNBP3 N-terminal domain reveals the mechanism for beta-1,3-glucan-specific recognition. Proc. Natl. Acad. Sci. USA 2009, 106, 11679–11684. [Google Scholar] [CrossRef] [Green Version]
- Aketagawa, J.; Tanaka, S.; Tamura, H.; Shibata, Y.; Saitô, H. Activation of limulus coagulation factor G by several (1-->3)-beta-D-glucans: Comparison of the potency of glucans with identical degree of polymerization but different conformations. J. Biochem. 1993, 113, 683–686. [Google Scholar] [CrossRef]
- Kurone, Y.; Ishibashi, K.I.; Yamanaka, D.; Miura, N.N.; Adachi, Y.; Ohno, N. Preparation and Biological Characterization of Limulus Factor G-activating Substance of Aspergillus spp. Med. Mycol. J. 2017, 58, E121–E129. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, M.; Ohno, N.; Miura, N.N.; Adachi, Y.; Aizawa, M.W.; Tamura, H.; Tanaka, S.; Yadomae, T. Chemical and immunochemical characterization of limulus factor G-activating substance of Candida spp. FEMS Immunol. Med. Microbiol. 1999, 24, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Mitsutake, K.; Miyazaki, T.; Miyazaki, H.; Iwamoto, M.; Maesaki, S.; Hashimoto, A.; Kaku, M.; Koga, H.; Kohno, S.; Hara, K. Evaluation of (1-3)-beta-D-glucan in aspergillosis and cryptococcosis. Nihon Kyobu Shikkan Gakkai Zasshi 1994, 32, 37–41. [Google Scholar]
- Zhou, Y.; Liang, Y.; Yan, Q.; Zhang, L.; Chen, D.; Ruan, L.; Kong, Y.; Shi, H.; Chen, M.; Chen, J. The draft genome of horseshoe crab Tachypleus tridentatus reveals its evolutionary scenario and well-developed innate immunity. BMC Genom. 2020, 21, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, A.; Yasui, M.; Kawagoe, T.; Tamura, H.; Tanaka, S.; Takagi, H. False-positive endotoxemia derives from gauze glucan after hepatectomy for hepatocellular carcinoma with cirrhosis. Hepato-Gastroenterol. 1997, 44, 1413–1418. [Google Scholar]
- Ohata, A.; Usami, M.; Horiuchi, T.; Nagasawa, K.; Kinoshita, K. Release of (1→3)-beta-D-glucan from depth-type membrane filters and their in vitro effects on proinflammatory cytokine production. Artif. Organs 2003, 27, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Nakajima, M.; Yamasaki, M.; Kitano, Y.; Niki, Y.; Ohsawa, G.; Matsushima, T. Investigation of the influence of hemodialysis membranes and factors associated with hemodialysis on serum (1→3)-beta-D-glucan. Kansenshogaku Zasshi. J. Jpn. Assoc. Infect. Dis. 1998, 72, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, N.; Emori, Y.; Yadomae, T.; Saito, K.; Masuda, A.; Oikawa, S. Reactivity of Limulus amoebocyte lysate towards (1—3)-beta-D-glucans. Carbohydr. Res. 1990, 207, 311–318. [Google Scholar] [CrossRef]
- Liss, B.; Cornely, O.A.; Hoffmann, D.; Dimitriou, V.; Wisplinghoff, H. 1,3-β-D-Glucan contamination of common antimicrobials. J. Antimicrob. Chemother. 2016, 71, 913–915. [Google Scholar] [CrossRef]
- Moro, H.; Koshio, N.; Bamba, Y.; Koizumi, T.; Cho, H.; Aoki, N.; Hayashi, M.; Tsubata, C.; Sakagami, A.; Sato, M.; et al. The Effect of Intravenous Gamma-globulin Reagents on the Measurement Results of (1→3)-β-D-glucan. Kansenshogaku Zasshi. J. Jpn. Assoc. Infect. Dis. 2017, 91, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Neun, B.W.; Cedrone, E.; Potter, T.M.; Crist, R.M.; Dobrovolskaia, M.A. Detection of Beta-Glucan Contamination in Nanotechnology-Based Formulations. Molecules 2020, 25, 3367. [Google Scholar] [CrossRef]
- Wan, G.H.; Li, C.S.; Guo, S.P.; Rylander, R.; Lin, R.H. An airbone mold-derived product, beta-1,3-D-glucan, potentiates airway allergic responses. Eur. J. Immunol. 1999, 29, 2491–2497. [Google Scholar] [CrossRef]
- Ariizumi, T.; Toriyama, K. Genetic regulation of sporopollenin synthesis and pollen exine development. Annu. Rev. Plant Biol. 2011, 62, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Doblin, M.S.; De Melis, L.; Newbigin, E.; Bacic, A.; Read, S.M. Pollen tubes of Nicotiana alata express two genes from different beta-glucan synthase families. Plant Physiol. 2001, 125, 2040–2052. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, S.; Zinkl, G.M.; Swanson, R.J.; Maruyama, D.; Preuss, D. Callose (beta-1,3 glucan) is essential for Arabidopsis pollen wall patterning, but not tube growth. BMC Plant Biol. 2005, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rylander, R.; Fogelmark, B.; McWilliam, A.; Currie, A. (1→3)-beta-D-glucan may contribute to pollen sensitivity. Clin. Exp. Immunol. 1999, 115, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Finkelman, M.A.; Lempitski, S.J.; Slater, J.E. beta-Glucans in standardized allergen extracts. J. Endotoxin Res. 2006, 12, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Saito, H. Sugi pollen disease from the forester’s viewpoint with special respect to pollen production in some forests. Proct. Otol. 1995, 76, 6–19. [Google Scholar]
- Kanno, T.; Adachi, Y.; Ohashi-Doi, K.; Matsuhara, H.; Hiratsuka, R.; Ishibashi, K.I.; Yamanaka, D.; Ohno, N. Latent 1,3-β-D-glucan acts as an adjuvant for allergen-specific IgE production induced by Japanese cedar pollen exposure. Allergol. Int. 2020, 70, 105–113. [Google Scholar] [CrossRef]
- Forestry Agency. About Cedar Pollen. Available online: https://www.rinya.maff.go.jp/j/press/hozen/pdf/111227-03.pdf (accessed on 17 February 2021). (In Japanese).
- Adachi, Y.; Ishii, M.; Kanno, T.; Tetsui, J.; Ishibashi, K.I.; Yamanaka, D.; Miura, N.; Ohno, N. N-Terminal (1→3)-β-d-Glucan Recognition Proteins from Insects Recognize the Difference in Ultra-Structures of (1→3)-β-d-Glucan. Int. J. Mol. Sci. 2019, 20, 3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, N.; Uchiyama, M.; Tsuzuki, A.; Tokunaka, K.; Miura, N.N.; Adachi, Y.; Aizawa, M.W.; Tamura, H.; Tanaka, S.; Yadomae, T. Solubilization of yeast cell-wall beta-(1-->3)-D-glucan by sodium hypochlorite oxidation and dimethyl sulfoxide extraction. Carbohydr. Res. 1999, 316, 161–172. [Google Scholar] [CrossRef]
- Read, S.M.; Currie, G.; Bacic, A. Analysis of the structural heterogeneity of laminarin by electrospray-ionisation-mass spectrometry. Carbohydr. Res. 1996, 281, 187–201. [Google Scholar] [CrossRef]
- Hoffmann, G.C.; Simson, B.W.; Timell, T.E. Structure and molecular size of pachyman. Carbohydr. Res. 1971, 20, 185–188. [Google Scholar] [CrossRef]
- Tabata, K.; Ito, W.; Kojima, T.; Kawabata, S.; Misaki, A. Ultrasonic degradation of schizophyllan, an antitumor polysaccharide produced by Schizophyllum commune Fries. Carbohydr. Res. 1981, 89, 121–135. [Google Scholar] [CrossRef]
- Kono, H.; Kondo, N.; Hirabayashi, K.; Ogata, M.; Totani, K.; Ikematsu, S.; Osada, M. NMR spectroscopic structural characterization of a water-soluble β-(1→3, 1→6)-glucan from Aureobasidium pullulans. Carbohydr. Polym. 2017, 174, 876–886. [Google Scholar] [CrossRef]
- Kono, H.; Kondo, N.; Isono, T.; Ogata, M.; Hirabayashi, K. Characterization of the secondary structure and order-disorder transition of a β-(1→3, 1→6)-glucan from Aureobasidium pullulans. Int. J. Biol. Macromol. 2020, 154, 1382–1391. [Google Scholar] [CrossRef]
- Tada, R.; Tanioka, A.; Iwasawa, H.; Hatashima, K.; Shoji, Y.; Ishibashi, K.; Adachi, Y.; Yamazaki, M.; Tsubaki, K.; Ohno, N. Structural characterisation and biological activities of a unique type beta-D-glucan obtained from Aureobasidium pullulans. Glycoconj. J. 2008, 25, 851–861. [Google Scholar] [CrossRef]
- Harada, T.; Masada, M.; Fujimori, K.; Maeda, I. Production of a firm, resilient gel-forming polysaccharide by a mutant of Alcaligenes faecalis var. myxogenes 10 C3. Agric. Biol. Chem. 1966, 30, 196–198. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.E.; Stone, B.A. Structure of the paramylon from Euglena gracilis. Biochim. Biophys. Acta 1960, 44, 161–163. [Google Scholar] [CrossRef]
- Bacic, A. Chemistry and organization of aleurone cell wall components from wheat and barley. Aust. J. Plants Physiol. 1981, 8, 475–495. [Google Scholar] [CrossRef]
- Sathyanarayana, B.K.; Stevens, E.S. Theoretical study of the conformations of pustulan [(1—6)-beta-D-glucan]. J. Biomol. Struct. Dyn. 1983, 1, 947–959. [Google Scholar] [CrossRef]
- Catley, B.J.; Whelan, W.J. Observations on the structure of pullulan. Arch. Biochem. Biophys. 1971, 143, 138–142. [Google Scholar] [CrossRef]
- Okuda, M.; Goto, M.; Okubo, K. Adhesion of Japanese cedar pollen to skin, clothes and laundry. Allergy 2003, 52, 855. [Google Scholar]
- Kanno, T.; Adachi, Y.; Ishibashi, K.I.; Yamanaka, D.; Ohno, N. Quantification of β-Glucan from Culinary-Medicinal Mushrooms Using Novel Artificial β-Glucan Recognition Protein. Int. J. Med. Mushrooms 2020, 22, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Tada, R.; Adachi, Y.; Ishibashi, K.; Tsubaki, K.; Ohno, N. Binding Capacity of a Barley β-d-Glucan to the β-Glucan Recognition Molecule Dectin-1. J. Agric. Food Chem. 2008, 56, 1442–1450. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanno, T.; Kim, C.; Yamanaka, D.; Ishibashi, K.-i.; Tanaka, H.; Ohno, N.; Adachi, Y. Possibility of Japanese Cedar Pollen Causing False Positives in the Deep Mycosis Test. Int. J. Mol. Sci. 2021, 22, 2135. https://doi.org/10.3390/ijms22042135
Kanno T, Kim C, Yamanaka D, Ishibashi K-i, Tanaka H, Ohno N, Adachi Y. Possibility of Japanese Cedar Pollen Causing False Positives in the Deep Mycosis Test. International Journal of Molecular Sciences. 2021; 22(4):2135. https://doi.org/10.3390/ijms22042135
Chicago/Turabian StyleKanno, Takashi, Changmin Kim, Daisuke Yamanaka, Ken-ichi Ishibashi, Hiroshi Tanaka, Naohito Ohno, and Yoshiyuki Adachi. 2021. "Possibility of Japanese Cedar Pollen Causing False Positives in the Deep Mycosis Test" International Journal of Molecular Sciences 22, no. 4: 2135. https://doi.org/10.3390/ijms22042135
APA StyleKanno, T., Kim, C., Yamanaka, D., Ishibashi, K. -i., Tanaka, H., Ohno, N., & Adachi, Y. (2021). Possibility of Japanese Cedar Pollen Causing False Positives in the Deep Mycosis Test. International Journal of Molecular Sciences, 22(4), 2135. https://doi.org/10.3390/ijms22042135