
The Impact of Selected Bacterial Sexually Transmitted Diseases on Pregnancy and Female Fertility

 ,
,  ,
, 
Abstract
:1. Introduction
2. Epidemiology
2.1. General Epidemiology (Including Epidemiology in Women and/or Pregnant Women)
2.2. Genotyping and Molecular Epidemiology
3. Pathogenesis (in Women)
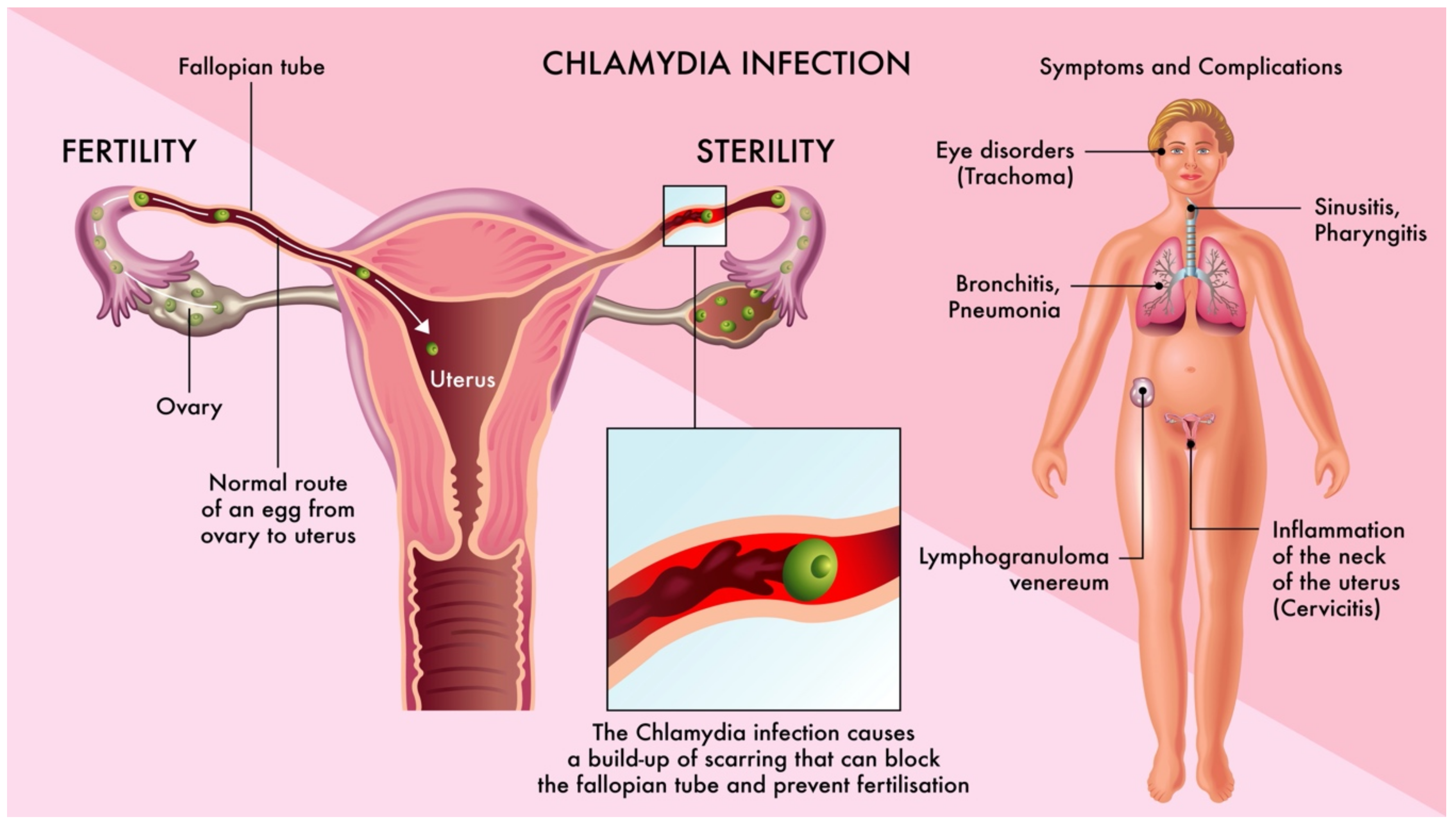
3.1. The Pathogenesis of Fallopian Tube Damage
3.2. The Pathogenesis of Infections during Pregnancy
4. Symptoms in Women with Special Attention to the Impact on Pregnancy and Infertility
4.1. Neisseria gonorrhoeae
4.2. Chlamydia trachomatis
4.3. Mycoplasma genitalium
5. Molecular Diagnostics
6. Therapy (with Special Attention to Therapy in Pregnant Women)
6.1. Neisseria gonnorhoeae
6.2. Chlamydia trachomatis
6.3. Mycoplasma genitalium
7. Antibiotic Resistance
7.1. Neisseria gonorrhoeae
7.2. Chlamydia trachomatis
7.3. Mycoplasma genitalium
8. New Antibiotics
8.1. Marketing Authorization Already Approved
8.2. Investigational Drugs
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| STIs | Sexually transmitted infections |
| PID | Pelvic inflammatory disease |
| TFI | Tubal factor infertility |
| CDC | Centers for Disease Control and Prevention |
| LGV | lymphogranuloma venereum |
| WGS | Whole genome sequencing |
| NG-MAST | Neisseria gonorrhoeae multiantigen sequence typing |
| MLST | Multilocus sequence typing |
| NG-STAR | Neisseria gonorrhoeae sequence typing for antimicrobial resistance |
| MOMP | Major outer membrane protein |
| MSM | Men who have sex with men |
| CT | Chlamydia trachomatis |
| TLR-4 | Toll-like receptor 4 |
| SGA | Small for gestational age |
| LBW | Low birth weight |
| NAATs | Nucleic acid amplification tests |
| EIA | Enzyme immunoassay |
| DIF | Direct immunofluorescence |
| SDA | Strand displacement amplification |
| TMAnvCT | Transcription-mediated amplificationNew variant C. trachomatis |
| SNP | Single-nucleotide polymorphism |
| HPA | Hybridization protection assay |
| Euro-GASPMIC | European Gonococcal Antimicrobial Surveillance ProgrammeMinimum inhibitory concentration |
| ETFs | Electron transfer proteins |
| MG | Mycoplasma genitalium |
References
- Pellati, D.; Mylonakis, I.; Bertoloni, G.; Fiore, C.; Andrisani, A.; Ambrosini, G.; Armanini, D. Genital tract infections and infertility. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 140, 3–11. [Google Scholar] [CrossRef]
- Paavonen, J. Chlamydia trachomatis infections of the female genital tract: State of the art. Ann. Med. 2012, 44, 18–28. [Google Scholar] [CrossRef]
- Tsevat, D.G.; Wiesenfeld, H.C.; Parks, C.; Peipert, J.F. Sexually transmitted diseases and infertility. Am. J. Obstet. Gynecol. 2017, 216, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuchardt, L.; Rupp, J. Chlamydia trachomatis as the Cause of Infectious Infertility: Acute, Repetitive or Persistent Long-Term Infection? Curr. Top. Microbiol. Immunol. 2018, 412, 159–182. [Google Scholar] [CrossRef] [PubMed]
- Weström, L. Effect of acute pelvic inflammatory disease on fertility. Am. J. Obstet. Gynecol. 1975, 121, 707–713. [Google Scholar] [CrossRef]
- Mårdh, P.A. Tubal factor infertility, with special regard to chlamydial salpingitis. Curr. Opin. Infect. Dis. 2004, 17, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Olaleye, A.O.; Babah, O.A.; Osuagwu, C.S.; Ogunsola, F.T.; Afolabi, B.B. Sexually transmitted infections in pregnancy—An update on Chlamydia trachomatis and Neisseria gonorrhoeae. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 255, 1–12. [Google Scholar] [CrossRef]
- Peuchant, O.; Le Roy, C.; Desveaux, C.; Paris, A.; Asselineau, J.; Maldonado, C.; Chêne, G.; Horovitz, J.; Dallay, D.; de Barbeyrac, B.; et al. Screening for Chlamydia trachomatis, Neisseria gonorrhoeae, and Mycoplasma genitalium should it be integrated into routine pregnancy care in French young pregnant women? Diagn. Microbiol. Infect. Dis. 2015, 82, 14–19. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Sexually Transmitted Infections in Europe 2013; ECDC: Stockholm, Sweden, 2015. [Google Scholar]
- World Health Organization. Report on Global Sexually Transmitted Infection Surveillance; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- US Centers for Disease Control and Prevention. Sexually Transmitted Disease Surveillance, 2017; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018. [Google Scholar]
- Chinese Center for Disease Control and Prevention. Overview of the National Legal Infectious Disease Epidemic in 2018; CDC China: Beijing, China, 2019. Available online: http://www.nhc.gov.cn/jkj/new_index.shtml (accessed on 30 September 2019).
- Manavi, K. A review on infection with Chlamydia trachomatis. Best. Pract. Res. Clin. Obstet. Gynaecol. 2006, 20, 941–951. [Google Scholar] [CrossRef] [PubMed]
- CDC. Sexually Transmitted Disease Surveillance, 2018; Department of Health and Human Services: Atlanta, GA, USA, 2019. [Google Scholar]
- McGowin, C.L.; Anderson-Smits, C. Mycoplasma genitalium: An emerging cause of sexually transmitted disease in women. PLoS Pathog. 2011, 7, e1001324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banhart, S.; Jansen, K.; Buder, S.; Tamminga, T.; Calvignac-Spencer, S.; Pilz, T.; Martini, A.; Dudareva, S.; Nikisins, S.; Dehmel, K.; et al. Molecular epidemiological typing of Neisseria gonorrhoeae isolates identifies a novel association between genogroup G10557 (G7072) and decreased susceptibility to cefixime, Germany, 2014 to 2017. Euro Surveill. 2020, 25, 1900648. [Google Scholar] [CrossRef]
- Shaskolskiy, B.; Dementieva, E.; Kandinov, I.; Chestkov, A.; Kubanov, A.; Deryabin, D.; Gryadunov, D. Genetic diversity of Neisseria gonorrhoeae multi-antigen sequence types in Russia and Europe. Int. J. Infect. Dis. 2020, 93, 1–8. [Google Scholar] [CrossRef]
- Harris, S.R.; Cole, M.J.; Spiteri, G.; Sánchez-Busó, L.; Golparian, D.; Jacobsson, S.; Goater, R.; Abudahab, K.; Yeats, C.A.; Bercot, B.; et al. Public health surveillance of multidrug-resistant clones of Neisseria gonorrhoeae in Europe: A genomic survey. Lancet Infect. Dis. 2018, 18, 758–768. [Google Scholar] [CrossRef]
- Kenyon, C.; Laumen, J.; Van Dijck, C.; De Baetselier, I.; Abdelatti, S.; Manoharan-Basil, S.S.; Unemo, M. Gonorrhoea treatment combined with population-level general cephalosporin and quinolone consumption may select for Neisseria gonorrhoeae antimicrobial resistance at the levels of NG-MAST genogroup: An ecological study in Europe. J. Glob. Antimicrob. Resist. 2020, 23, 377–384. [Google Scholar] [CrossRef]
- Golparian, D.; Sánchez-Busó, L.; Cole, M.; Unemo, M. Neisseria gonorrhoeae sequence typing for antimicrobial resistance (NG-STAR) clonal complexes are consistent with genomic phylogeny and provide simple nomenclature, rapid visualization and antimicrobial resistance (AMR) lineage predictions. J. Antimicrob. Chemother. 2021. [Google Scholar] [CrossRef]
- Thapa, J.; Watanabe, T.; Isoba, M.; Okubo, T.; Abe, K.; Minami, K.; Yamaguchi, H. Chlamydia trachomatis isolated from cervicovaginal samples in Sapporo, Japan, reveals the circulation of genetically diverse strains. BMC Infect. Dis. 2020, 20, 53. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.; Gomes, J.P. Evolution, phylogeny, and molecular epidemiology of Chlamydia. Infect. Genet. Evol. 2014, 23, 49–64. [Google Scholar] [CrossRef]
- Smolarczyk, K.; Mlynarczyk-Bonikowska, B.; Majewski, S.; Rudnicka, E.; Unemo, M.; Fiedor, P. Lymphogranuloma venereum (LGV)—An emerging problem in Poland. Postepy Dermatol. Alergol. 2020. [Google Scholar] [CrossRef]
- Patiño, L.H.; Camargo, M.; Muñoz, M.; Ríos-Chaparro, D.I.; Patarroyo, M.A.; Ramírez, J.D. Unveiling the multilocus sequence typing (MLST) schemes and core genome phylogenies for genotyping Chlamydia Trach. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Borges, V.; Cordeiro, D.; Salas, A.I.; Lodhia, Z.; Correia, C.; Isidro, J.; Fernandes, C.; Rodrigues, A.M.; Azevedo, J.; Alves, J.; et al. Chlamydia trachomatis: When the virulence-associated genome backbone imports a prevalence-associated major antigen signature. Microb. Genom. 2019, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjorth, S.V.; Björnelius, E.; Lidbrink, P.; Falk, L.; Dohn, B.; Berthelsen, L.; Ma, L.; Martin, D.H.; Jensen, J.S. Sequence-based typing of Mycoplasma genitalium reveals sexual transmission. J. Clin. Microbiol. 2006, 44, 2078–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumke, R.; Rust, M.; Glaunsinger, T. MgpB types among Mycoplasma genitalium strains from men who have sex with men in Berlin, Germany, 2016–2018. Pathogens 2020, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fookes, M.C.; Hadfield, J.; Harris, S.; Parmar, S.; Unemo, M.; Jensen, J.S.; Thomson, N.R. Mycoplasma genitalium: Whole genome sequence analysis, recombination and population structure. BMC Genom. 2017, 18, 993. [Google Scholar] [CrossRef] [PubMed]
- Beatty, W.L.; Morrison, R.P.; Byrne, G.I. Persistent chlamydiae: From cell culture to a paradigm for chlamydial pathogenesis. Microbiol. Rev. 1994, 58, 686–699. [Google Scholar] [CrossRef]
- Linhares, I.M.; Witkin, S.S. Immunopathogenic consequences of Chlamydia trachomatis 60 kDa heat shock protein expression in the female reproductive tract. Cell Stress Chaperones 2010, 15, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Akande, V.A.; Hunt, L.P.; Cahill, D.J.; Caul, E.O.; Ford, W.C.; Jenkins, J.M. Tubal damage in infertile women: Prediction using chlamydia serology. Hum. Reprod. 2003, 18, 1841–1847. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Briones, M.; Chiou, J.; Lei, L.; Patton, M.J.; Ma, L.; McClarty, G.; Caldwell, H.D. Chlamydia trachomatis lipopolysaccharide evades the canonical and concanonical inflammatory pathways to subvert innate immunity. mBio 2019, 10, e00595-19. [CrossRef] [Green Version]
- Hughes, G.; Fenton, K.A. Recent trends in gonorrhoea—An emerging public health issue? Euro Surveill. 2000, 5, 12. [Google Scholar] [CrossRef]
- Maisey, K.; Nardocci, G.; Imarai, M.; Cardenas, H.; Rios, M.; Croxatto, H.B.; Heckels, J.E.; Christodoulides, M.; Velasquez, L.A. Expression of proinflammatory cytokines and receptors by human fallopian tubes in organ culture following challenge with Neisseria gonorrhoeae. Infect. Immun. 2003, 71, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quillin, S.J.; Seifert, H.S. Neisseria gonorrhoeae host adaptation and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 226–240. [Google Scholar] [CrossRef]
- Lenz, J.D.; Dillard, J.P. Pathogenesis of Neisseria gonorrhoeae and the host defense in ascending infections of human Fallopian tube. Front. Immunol. 2018, 9, 2710. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.R.; Manhart, L.E.; Bukusi, E.A.; Astete, S.; Brunham, R.C.; Holmes, K.K.; Sinei, S.K.; Bwayo, J.J.; Totten, P.A. Association between Mycoplasma genitalium and acute endometritis. Lancet 2002, 359, 765–766. [Google Scholar] [CrossRef]
- McGowin, C.L.; Ma, L.; Martin, D.H.; Pyles, R.B. Mycoplasma genitalium-encoded MG309 activates NF-kappaB via Toll-like receptors 2 and 6 to elicit proinflammatory cytokine secretion from human genital epithelial cells. Infect. Immun. 2009, 77, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- McGowin, C.L.; Popov, V.L.; Pyles, R.B. Intracellular Mycoplasma genitalium infection of human vaginal and cervical epithelial cells elicits distinct patterns of inflammatory cytokine secretion and provides a possible survival niche against macrophage-mediated killing. BMC Microbiol. 2009, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowin, C.L.; Totten, P.A. The unique microbiology and molecular pathogenesis of Mycoplasma genitalium. J. Infect. Dis. 2017, 216 (Suppl. 2), S382–S388. [Google Scholar] [CrossRef] [Green Version]
- Baczynska, A.; Funch, P.; Fedder, J.; Knudsen, H.J.; Birkelund, S.; Christiansen, G. Morphology of human Fallopian tubes after infection with Mycoplasma genitalium and Mycoplasma hominis--in vitro organ culture study. Hum. Reprod. 2007, 22, 968–979. [Google Scholar] [CrossRef] [Green Version]
- Paavonen, J.; Eggert-Kruse, W. Chlamydia trachomatis: Impact on human reproduction. Hum. Reprod. Update 1999, 5, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Equils, O.; Lu, D.; Gatter, M.; Witkin, S.S.; Bertolotto, C.; Arditi, M.; McGregor, J.A.; Simmons, C.F.; Hobel, C.J. Chlamydia heat shock protein 60 induces trophoblast apoptosis through TLR4. J. Immunol. 2006, 177, 1257–1263. [Google Scholar] [CrossRef] [Green Version]
- Lis, R.; Rowhani-Rahbar, A.; Manhart, L.E. Mycoplasma genitalium infection and female reproductive tract disease: A meta-analysis. Clin. Infect. Dis. 2015, 61, 418–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unemo, M.; Ross, J.; Serwin, A.B.; Gomberg, M.; Cusini, M.; Jensen, J.S. 2020 European guideline for the diagnosis and treatment of gonorrhoea in adults. Int. J. STD AIDS 2020. [Google Scholar] [CrossRef] [PubMed]
- Heumann, C.L.; Quilter, L.A.S.; Eastment, M.C.; Heffron, R.; Hawes, S.E. Adverse birth outcomes and maternal Neisseria gonorrhoeae infection: A population-based cohort study in Washington state. Sex. Transm. Dis. 2017, 44, 266–271. [Google Scholar] [CrossRef] [Green Version]
- Weström, L.; Joesoef, R.; Reynolds, G.; Hagdu, A.; Thompson, S.E. Pelvic inflammatory disease and fertility. A cohort study of 1,844 women with laparoscopically verified disease and 657 control women with normal laparoscopic results. Sex. Transm. Dis. 1992, 19, 185–192. [Google Scholar] [CrossRef]
- Haggerty, C.L.; Gottlieb, S.L.; Taylor, B.D.; Low, N.; Xu, F.; Ness, R.B. Risk of sequelae after Chlamydia trachomatis genital infection in women. J. Infect. Dis. 2010, 201, S134–S155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor-Robinson, D. Mycoplasma genitalium—An up-date. Int. J. STD AIDS 2002, 13, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Rajkumari, N.; Kaur, H.; Roy, A.; Gupta, N.; Dhaliwal, L.K.; Sethi, S. Association of Mycoplasma genitalium with infertility in North Indian women. Indian J. Sex. Transm. Dis. AIDS 2015, 36, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.S.; Cusini, M.; Gomberg, M.; Moi, H. 2016 European guideline on Mycoplasma genitalium infections. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1650–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, D.; Regan, L.; Greub, G. Emerging role of Chlamydia and Chlamydia-like organisms in adverse pregnancy outcomes. Curr. Opin. Infect. Dis. 2008, 21, 70–76. [Google Scholar] [CrossRef]
- Baud, D.; Goy, G.; Jaton, K.; Osterheld, M.-C.; Blumer, S.; Borel, N.; Vial, Y.; Hohlfeld, P.; Pospischil, A.; Greub, G. Role of Chlamydia trachomatis in miscarriage. Emerg. Infect. Dis. 2011, 17, 1630–1635. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.J.P.M.D.A.; Florêncio, G.L.D.; Gabiatti, J.R.E.; Amaral, R.L.D.; Eleutério Júnior, J.; Gonçalves, A.K.D.S. Perinatal morbidity and mortality associated with chlamydial infection: A meta-analysis study. Braz. J. Infect. Dis. 2011, 15, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Likitnukul, S.; Kusmiesz, H.; Nelson, J.D.; McCracken, G.H., Jr. Role of genital mycoplasmas in young infants with suspected sepsis. J. Pediatr. 1986, 109, 971–974. [Google Scholar] [CrossRef]
- Temmerman, M.; Plummer, F.A.; Kiragu, D.; Farah, A.; Wamola, I.A.; Brunham, R.C.; Piot, P. Gonorrhoea in pregnancy. J. Obstet. Gynecol. 1992, 12, 162–166. [Google Scholar] [CrossRef]
- Ismail, M.A.; Moawad, A.H.; Poon, E.; Henderson, C. Role of Chlamydia trachomatis in postpartum endometritis. J. Reprod. Med. 1987, 32, 280–284. [Google Scholar]
- Rours, G.I.J.G.; Duijts, L.; Moll, H.A.; Arends, L.R.; de Groot, R.; Jaddoe, V.W.; Hofman, A.; Steegers, E.A.P.; Mackenbach, J.P.; Ott, A.; et al. Chlamydia trachomatis infection during pregnancy associated with preterm delivery: A population-based prospective cohort study. Eur. J. Epidemiol. 2011, 26, 493–502. [Google Scholar] [CrossRef]
- Labbé, A.C.; Frost, E.; Deslandes, S.; Mendonça, A.P.; Alves, A.C.; Pépin, J. Mycoplasma genitalium is not associated with adverse outcomes of pregnancy in Guinea-Bissau. Sex. Transm. Infect. 2002, 78, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Kacerovský, M.; Pavlovský, M.; Tosner, J. Preterm premature rupture of the membranes and genital mycoplasmas. Acta Med. 2009, 52, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Bignell, C.; Unemo, M. 2012 European guideline on the diagnosis and treatment of gonorrhoea in adults. Int. J. STD AIDS 2013, 24, 85–92. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Recommendations for the laboratory-based detection of Chlamydia trachomatis and Neisseria gonorrhoeae-2014. MMWR Recomm. Rep. 2014, 63, 1–19. [Google Scholar]
- Lanjouw, E.; Ouburg, S.; de Vries, H.J.; Stary, A.; Radcliffe, K.; Unemo, M. 2015 European guideline on the management of Chlamydia trachomatis infections. Int. J. STD AIDS 2016, 27, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Trembizki, E.; Costa, A.-M.G.; Tabrizi, S.N.; Whiley, D.M.; Twin, J. Opportunities and pitfalls of molecular testing for detecting sexually transmitted pathogens. Pathology 2015, 47, 219–226. [Google Scholar] [CrossRef]
- Tabrizi, S.N.; Unemo, M.; Limnios, A.E.; Hogan, T.R.; Hjelmevoll, S.O.; Garland, S.M.; Tapsall, J. Evaluation of six commercial nucleic acid amplification tests for detection of Neisseria gonorrhoeae and other Neisseria species. J. Clin. Microbiol. 2011, 49, 3610–3615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unemo, M.; Hansen, M.; Hadad, R.; Puolakkainen, M.; Westh, H.; Rantakokko-Jalava, K.; Thilesen, C.; Cole, M.J.; Boiko, I.; Lan, P.T.; et al. Sensitivity, specificity, inclusivity and exclusivity of the updated Aptima Combo 2 assay, which provides detection coverage of the new diagnostic-escape Chlamydia trachomatis variants. BMC Infect. Dis. 2020, 20, 419. [Google Scholar] [CrossRef] [PubMed]
- Gaydos, C.A.; Manhart, L.E.; Taylor, S.N.; Lillis, R.A.; Hook, E.W., 3rd; Klausner, J.D.; Remillard, C.V.; Love, M.; McKinney, B.; Getman, D.K. Molecular testing for Mycoplasma genitalium in the United States: Results from the AMES prospective multicenter clinical study. J. Clin. Microbiol. 2019, 57, e01125-19. [Google Scholar] [CrossRef] [Green Version]
- Drud, S.T.; Njuguna, P.; Ebeyan, S.; Erskine, S.; Holm, M.; Johansson, S.C.; Tan, L.Y.; Jensen, J.S. Evaluation of the resistancePlus MG FleXible assay for detection of wild-type and 23S rRNA-mutated Mycoplasma genitalium strains. J. Clin. Microbiol. 2020, 58, e01900-19. [Google Scholar] [CrossRef] [Green Version]
- Harding-Esch, E.M.; Cousins, E.C.; Chow, S.L.C.; Phillips, L.T.; Hall, C.L.; Cooper, N.; Fuller, S.S.; Nori, A.V.; Patel, R.; Thomas-William, S.; et al. A 30-min nucleic acid amplification point-of-care test for genital Chlamydia trachomatis infection in women: A prospective, multi-center study of diagnostic accuracy. EBioMedicine 2018, 28, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, S.; Boiko, I.; Golparian, D.; Blondeel, K.; Kiarie, J.; Toskin, I.; Peeling, R.W.; Unemo, M. WHO laboratory validation of Xpert(®) CT/NG and Xpert(®) TV on the GeneXpert system verifies high performances. APMIS 2018, 126, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Workowski, K.A.; Bolan, G.A. Sexually transmitted diseases treatment guidelines, 2015. MMWR 2015, 64, 1–137. [Google Scholar] [PubMed]
- Cole, M.J.; Quaye, N.; Jacobsson, S.; Day, M.; Fagan, E.; Ison, C.; Pitt, R.; Seaton, S.; Woodford, N.; Stary, A.; et al. Ten years of external quality assessment (EQA) of Neisseria gonorrhoeae antimicrobial susceptibility testing in Europe elucidate high reliability of data. BMC Infect. Dis. 2019, 19, 281. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.J.; Quinten, C.; Jacobsson, S.; Day, M.; Amato-Gauci, A.J.; Woodford, N.; Spiteri, G.; Unemo, M.; Euro-GASPnetwork. The European gonococcal antimicrobial surveillance programme (Euro-GASP) appropriately reflects the antimicrobial resistance situation for Neisseria gonorrhoeae in the European Union/European Economic Area. BMC Infect. Dis. 2019, 19, 1040. [Google Scholar] [CrossRef] [PubMed]
- Unemo, M.; Del Rio, C.; Shafer, W.M. Antimicrobial resistance expressed by Neisseria gonorrhoeae: A major global public health problem in the 21st century. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Demczuk, W.; Sidhu, S.; Unemo, M.; Whiley, D.M.; Allen, V.G.; Dillon, J.R.; Cole, M.; Seah, C.; Trembizki, E.; Trees, D.L.; et al. Neisseria gonorrhoeae sequence typing for antimicrobial resistance, a novel antimicrobial resistance multilocus typing scheme for tracking global dissemination of N. gonorrhoeae strains. J. Clin. Microbiol. 2017, 55, 1454–1468. [Google Scholar] [CrossRef] [Green Version]
- Mlynarczyk-Bonikowska, B.; Majewska, A.; Malejczyk, M.; Mlynarczyk, G.; Majewski, S. Multiresistant Neisseria gonorrhoeae: A new threat in second decade of the XXI century. Med. Microbiol. Immunol. 2020, 209, 95–108. [Google Scholar] [CrossRef]
- Shaskolskiy, B.; Dementieva, E.; Kandinov, I.; Filippova, M.; Petrova, N.; Plakhova, X.; Chestkov, A.; Kubanov, A.; Deryabin, D.; Gryadunov, D. Resistance of Neisseria gonorrhoeae isolates to beta-lactam antibiotics (benzylpenicillin and ceftriaxone) in Russia, 2015–2017. PLoS ONE 2019, 14, e0220339. [Google Scholar] [CrossRef] [Green Version]
- Shaskolskiy, B.; Dementieva, E.; Leinsoo, A.; Petrova, N.; Chestkov, A.; Kubanov, A.; Deryabin, D.; Gryadunov, D. Tetracycline resistance of Neisseria gonorrhoeae in Russia, 2015–2017. Infect. Genet. Evol. 2018, 63, 236–242. [Google Scholar] [CrossRef]
- Gascoyne, D.M.; Heritage, J.; Hawkey, P.M.; Turner, A.; van Klingeren, B. Molecular evolution of tetracycline-resistance plasmids carrying TetM found in Neisseria gonorrhoeae from different countries. J. Antimicrob. Chemother. 1991, 28, 173–183. [Google Scholar] [CrossRef]
- Deguchi, T.; Hatazaki, K.; Ito, S.; Kondo, H.; Horie, K.; Nakane, K.; Mizutani, K.; Tsuchiya, T.; Yasuda, M.; Yokoi, S.; et al. Macrolide and fluoroquinolone resistance is uncommon in clinical strains of Chlamydia trachomatis. J. Infect. Chemother. 2018, 24, 610–614. [Google Scholar] [CrossRef]
- Dugan, J.; Rockey, D.D.; Jones, L.; Andersen, A.A. Tetracycline resistance in Chlamydia suis mediated by genomic islands inserted into the chlamydial inv-like gene. Antimicrob. Agents Chemother. 2004, 48, 3989–3995. [Google Scholar] [CrossRef] [Green Version]
- Mestrovic, T.; Ljubin-Sternak, S. Molecular mechanisms of Chlamydia trachomatis resistance to antimicrobial drugs. Front. Biosci. (Landmark. Ed.) 2018, 23, 656–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, C.S.; Jensen, J.S.; Waites, K.B. New horizons in Mycoplasma genitalium treatment. J. Infect. Dis. 2017, 216, S412–S419. [Google Scholar] [CrossRef] [PubMed]
- Tagg, K.A.; Jeoffreys, N.J.; Couldwell, D.L.; Donald, J.A.; Gilbert, G.L. Fluoroquinolone and macrolide resistance-associated mutations in Mycoplasma genitalium. J. Clin. Microbiol. 2013, 51, 2245–2249. [Google Scholar] [CrossRef] [Green Version]
- Murray, G.L.; Bodiyabadu, K.; Danielewski, J.; Garland, S.M.; Machalek, D.A.; Fairley, C.K.; Jensen, J.S.; Williamson, D.A.; Tan, L.Y.; Mokany, E.; et al. Moxifloxacin and sitafloxacin treatment failure in Mycoplasma genitalium infection: Association with parC mutation G248T (S83I) and concurrent gyrA mutations. J. Infect. Dis. 2020, 221, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Machalek, D.A.; Tao, Y.; Shilling, H.; Jensen, J.S.; Unemo, M.; Murray, G.; Chow, E.P.F.; Low, N.; Garland, S.M.; Vodstrcil, L.A.; et al. Prevalence of mutations associated with resistance to macrolides and fluoroquinolones in Mycoplasma genitalium: A systematic review and meta-analysis. Lancet Infect. Dis. 2020, 20, 1302–1314. [Google Scholar] [CrossRef]
- Chen, M.Y.; McNulty, A.; Avery, A.; Whiley, D.; Tabrizi, S.N.; Hardy, D.; Das, A.F.; Nenninger, A.; Fairley, C.K.; Hocking, J.S.; et al. Solithromycin versus ceftriaxone plus azithromycin for the treatment of uncomplicated genital gonorrhoea (SOLITAIRE-U): A randomised phase 3 non-inferiority trial. Lancet Infect. Dis. 2019, 19, 833–842. [Google Scholar] [CrossRef]
- Mendes, R.E.; Paukner, S.; Doyle, T.B.; Gelone, S.P.; Flamm, R.K.; Sader, H.S. Low prevalence of gram-positive isolates showing elevated lefamulin MIC results during the SENTRY surveillance program for 2015–2016 and characterization of resistance mechanisms. Antimicrob. Agents Chemother. 2019, 63, e02158-18. [Google Scholar] [CrossRef] [Green Version]
- Bradford, P.A.; Miller, A.A.; O’Donnell, J.; Mueller, J.P. Zoliflodacin: An oral spiropyrimidinetrione antibiotic for the treatment of Neisseria gonorrheae, including multi-drug-resistant isolates. ACS Infect. Dis. 2020, 6, 1332–1345. [Google Scholar] [CrossRef]
- Scangarella-Oman, N.E.; Hossain, M.; Dixon, P.B.; Ingraham, K.; Min, S.; Tiffany, C.A.; Perry, C.R.; Raychaudhuri, A.; Dumont, E.F.; Huang, J.; et al. Microbiological analysis from a phase 2 randomized study in adults evaluating single oral doses of gepotidacin in the treatment of uncomplicated urogenital gonorrhea caused by Neisseria gonorrhoeae. Antimicrob. Agents Chemother. 2018, 62, e01221-18. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, S.; Mason, C.; Khan, N.; Meo, P.; Unemo, M. In vitro activity of the novel oral antimicrobial SMT-571, with a new mechanism of action, against MDR and XDR Neisseria gonorrhoeae: Future treatment option for gonorrhoea? J. Antimicrob. Chemother. 2019, 74, 1591–1594. [Google Scholar] [CrossRef] [PubMed]
- Jacobsson, S.; Mason, C.; Khan, N.; Meo, P.; Unemo, M. High in vitro activity of DIS-73285, a novel antimicrobial with a new mechanism of action, against MDR and XDR Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2020, 75, 3244–3247. [Google Scholar] [CrossRef]
- Riedel, S.; Vijayakumar, D.; Berg, G.; Kang, A.D.; Smith, K.P.; Kirby, J.E. Evaluation of apramycin against spectinomycin-resistant and -susceptible strains of Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2019, 74, 1311–1316. [Google Scholar] [CrossRef]
- Bruhn, D.F.; Waidyarachchi, S.L.; Madhura, D.B.; Shcherbakov, D.; Zheng, Z.; Liu, J.; Abdelrahman, Y.M.; Singh, A.P.; Duscha, S.; Rathi, C.; et al. Aminomethyl spectinomycins as therapeutics for drug-resistant respiratory tract and sexually transmitted bacterial infections. Sci. Transl. Med. 2015, 7. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Neisseria gonorrhoeae | Chlamydia trachomatis | Mycoplasma genitalium |
| Ectopic pregnancy ⇐⇐ pelvic inflammatory disease (PID) ⇒⇒ infertility [3,44,52] | ||
| Miscarriage [53] | ||
| Increased perinatal mortality | Increased perinatal mortality [54] | Recurrent pregnancy loss [53] |
| Neonatal infection (conjunctivitis) | Neonatal infection (conjunctivitis, pneumonia) | Neonatal infection [55] |
| Postpartum endometritis [56] | Postpartum endometritis –ve [54]/+ve [57] | Postpartum endometritis |
| Low birth weight (LBW) [43] | ||
| Preterm birth [44,52] | ||
| Small for gestational age (SGA) [43] | Not small for gestational age (SGA) [58] | Not small for gestational age (SGA) [59] |
| Premature rupture of membranes [52,60] | ||
| NAAT | Producer | Detected Pathogen | Methods Used | Target |
|---|---|---|---|---|
| Aptima Combo 2 assay | Hologic Inc. | CT/NG | TMA | NG, specific regions in 16S rRNA CT, specific regions in 23Sr RNA |
| Aptima CT assay | Hologic Inc. | CT | TMA | Specific regions in 16S rRNA |
| Aptima NG assay | Hologic Inc. | NG | TMA | Specific regions in 16S rRNA |
| Abbott RealTime CT/NG | Abbott | CT/NG | real-time PCR | CT, two specific regions in cryptic plasmid NG, specific sequence in OPA gene |
| cobas 4800 CT/NG Test | Roche Diagnostics | CT/NG | PCR | CT, two targets, one in cryptic plasmid and one in chromosome NG, direct repeat (DR) 9 specific regions |
| ProbeTec ET CT/GC Amplified DNA assay | Becton Dickinson | CT/NG | SDA | CT, specific region in cryptic plasmid (ORF) NG, pilin gene inverting protein homolog |
| ProbeTec CT QX Amplified DNA assay | Becton Dickinson | CT | SDA | Specific region in cryptic plasmid (ORF) |
| ProbeTec NG QX Amplified DNA assay | Becton Dickinson | NG | SDA | Specific region in pilin gene |
| Xpert CT/NG | Cepheid | CT/NG | real-time PCR | CT, specific chromosomal DNA sequence NG, two specific chromosomal DNA sequences (both should be detected for a positive result) |
| Aptima Mycoplasma genitalium Assay | Hologic Inc. | MG | TMA | Specific regions in 16s rRNA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smolarczyk, K.; Mlynarczyk-Bonikowska, B.; Rudnicka, E.; Szukiewicz, D.; Meczekalski, B.; Smolarczyk, R.; Pieta, W. The Impact of Selected Bacterial Sexually Transmitted Diseases on Pregnancy and Female Fertility. Int. J. Mol. Sci. 2021, 22, 2170. https://doi.org/10.3390/ijms22042170
Smolarczyk K, Mlynarczyk-Bonikowska B, Rudnicka E, Szukiewicz D, Meczekalski B, Smolarczyk R, Pieta W. The Impact of Selected Bacterial Sexually Transmitted Diseases on Pregnancy and Female Fertility. International Journal of Molecular Sciences. 2021; 22(4):2170. https://doi.org/10.3390/ijms22042170
Chicago/Turabian StyleSmolarczyk, Katarzyna, Beata Mlynarczyk-Bonikowska, Ewa Rudnicka, Dariusz Szukiewicz, Blazej Meczekalski, Roman Smolarczyk, and Wojciech Pieta. 2021. "The Impact of Selected Bacterial Sexually Transmitted Diseases on Pregnancy and Female Fertility" International Journal of Molecular Sciences 22, no. 4: 2170. https://doi.org/10.3390/ijms22042170
APA StyleSmolarczyk, K., Mlynarczyk-Bonikowska, B., Rudnicka, E., Szukiewicz, D., Meczekalski, B., Smolarczyk, R., & Pieta, W. (2021). The Impact of Selected Bacterial Sexually Transmitted Diseases on Pregnancy and Female Fertility. International Journal of Molecular Sciences, 22(4), 2170. https://doi.org/10.3390/ijms22042170