1. Introduction
Maternal–fetal coordinated communications are required to promote healthy pregnancy outcomes. In addition, mothers confer natural and passive protection to their offspring through the transplacental pathway during gestation and later by lactation [
1,
2]. Cytokines (CKs), immunoglobulins (Igs), and adipokines are important components in fetal development and physiology during gestation [
3,
4,
5,
6,
7,
8]. However, the concentrations of these factors are different between populations, ethnicities, and genetic backgrounds. To date, Kee Thai Yeo and colleagues described that Kenyan neonates had higher levels of some of these factors, such as TNF-α, than United States neonates [
9].
The network of communication of the immune system is essential to maintain a tolerant and anti-inflammatory environment during pregnancy and, in turn, to protect the fetus from infections [
4]. However, in the parturition an inflammatory environment is needed [
3,
10] to lead to correct delivery [
11].
Maternal infections during birth produce higher levels of pro-inflammatory CKs in neonates [
9]. These concentration changes are very important since it has been observed that the concentrations of interleukin (IL)-6, IL-10, and C-X-C Motif Chemokine Ligand 8 (CXCL8) are associated with a higher risk of being small for gestational age (SGA) and higher concentrations of tumor necrosis factor receptor 1 (TNFRI), IL-5, IL-1, IL-8, and TNF-α are related to prematurity risk [
12,
13,
14].
Igs are glycoproteins that are divided in five classes (IgM, IgD, IgG, IgA, and IgE) according to their structure and functions [
15]. Neonates have low levels of IgM and even lower levels of IgA and IgE compared to later stages of life [
5]. Besides, IgG is mainly of maternal origin [
5], since it is the only Ig that can be transplacentally transferred.
With regard to adipokines, the adiponectin is a pleiotropic molecule that presents anti-inflammatory, anti-fibrotic, anti-apoptotic, and proangiogenic effects, in addition to its ability to increase the sensibility to insulin, which is its most known action [
16]. Recent studies have shown that the levels of this adipokine in cord blood are correlated positively with the accumulation of neonatal fat mass (FM) [
17], body mass index (BMI), and birth weight [
6], which is contrary to what it is observed in adults. Concerning leptin, there are controversial studies about pregnancy. On the one hand, it is reported that leptin levels increase in women during gestation because of their beneficial effects on the development of the fetal organs [
7]. On the other hand, it is thought that leptin concentration in breast milk and in fetal plasma can be associated with the risk of obesity later in life, and are also linked to maternal weight [
8,
18].
With regard to the maternal diet, Ellen L. Mozurkewich and colleagues demonstrated that supplements of omega-3 fatty acids (eicosapentaenoic acid, EPA; docosahexaenoic acid, DHA) in pregnant women decrease levels of proinflammatory CKs and are associated positively with those of IL-10 (anti-inflammatory CK) in maternal plasma but not in cord blood [
19]. In addition, during the gestational period the intake of potential food allergens induces tolerance in offspring in humans [
20]. Thus, this indicates that maternal diet can influence a baby’s health during pregnancy, although the exact mechanisms are rather unknown. A possible mechanism could be the influence on the maternal microbiota which, in turn, may influence the development of the neonate. In fact, there are several studies that demonstrate the impact of the diet on the maternal [
21,
22,
23,
24,
25] and infant microbiota [
26,
27,
28].
In this study, we first aimed to characterize maternal–neonatal plasma immune factor levels and their transmission in a Mediterranean birth cohort study [
29]. In a previous study with this cohort, it was determined that mothers with different diets had different intestinal microbiota compositions [
28], thus we further aimed as a secondary objective at evaluating the influence of the maternal diet and gut microbiota enterotype in this passive transmission line of immunobioactive compounds.
3. Discussion
Immune cells and immune factors participate in coordinated communications between the mother and fetus in order to support a successful pregnancy [
3,
32]. Little is known about the effect of maternal diet and maternal microbial enterotype on the composition of Igs, CKs, and adipokines in UCP. To evaluate this influence, samples from the mother–infant birth cohort in the Spanish–Mediterranean area (MAMI) were used. This knowledge may be used to improve the health and development of the neonate through nutritional interventions in pregnant women.
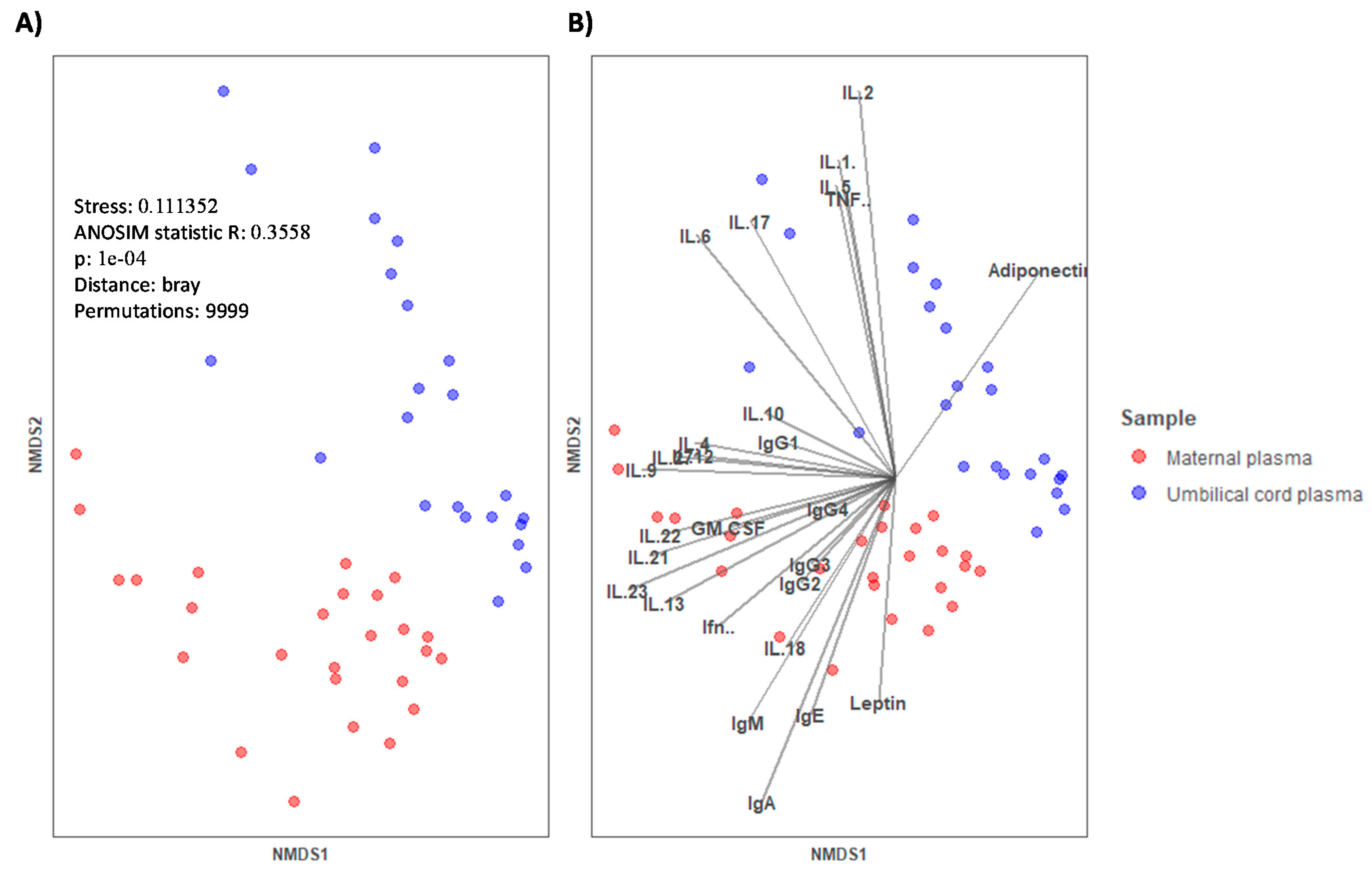
During pregnancy, the immune system must engage in maintaining tolerance towards the fetus while preserving the immune function and transferring passive immunity to the offspring. Therefore, the network of communication and trafficking of the immune system is pivotal [
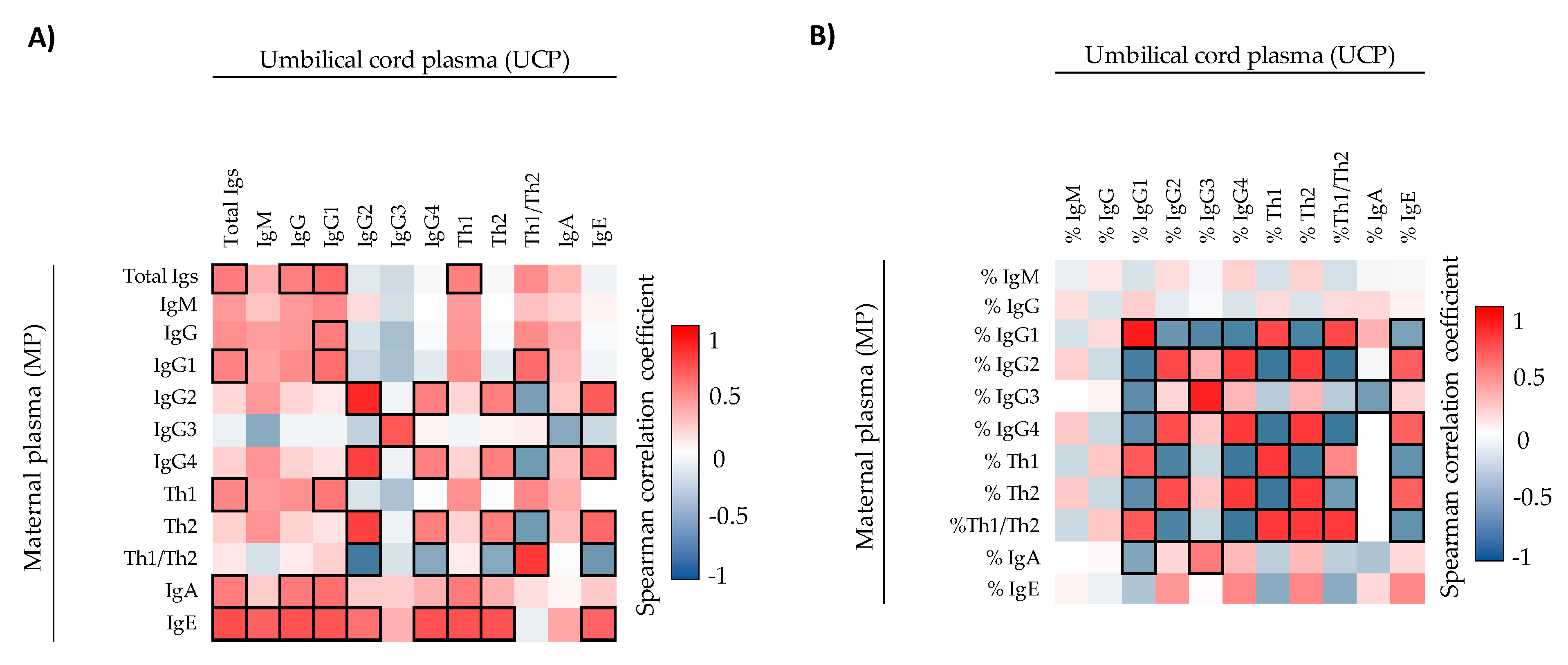
4]. In this study, we first characterized the immune and adipokine composition of UCP and MP. The overall composition of UCP was clearly different from that of MP. In general, the Ig absolute values were higher in MP than in UCP. However, %IgG and particularly %IgG1 predominated in UCP with respect to MP. These increases seemed to be due to the fact that the other percentages of classes and subclasses were lower in UCP (e.g., %IgM), causing a relative increase even though the changes in the concentrations were not pronounced. Otherwise, these increases could also be due to a real rise in relative proportions because, in addition to the transplacental transfer of IgG from the mother [
5,
33], the fetus begins to produce its own IgG haplotype from the 10
th week of gestation, and this concentration increases considerably on the delivery day [
34]. The other classes were found in lower concentrations since, to date, maternal IgG is the only known Ig to cross the placental barrier [
33], reaching the fetus by the FcRn receptors expressed by syncytiotrophoblasts [
35]. For this reason, IgM, IgA, and IgE are found in very low concentrations in infant circulation [
5]. Moreover, not all the IgG subclasses are transferred equally [
35]. This fact could explain why we found correlations between MP and UCP in IgG1, IgG2, IgG3, and IgG4 but not in total IgG. The absolute concentration of IgE also showed a correlation between MP and UCP in this study. As IgE does not cross the transplacental barrier [
33], other factors may be involved. In this regard, a study found both maternal–fetal and paternal–fetal IgE correlation [
36], suggesting that genetics and the home environment could influence the neonatal IgE concentration.
The implantation and parturition in a healthy pregnancy is mediated by pro-inflammatory responses [
3], although a tolerant and anti-inflammatory environment is generated during pregnancy [
4,
10]. A normal-term delivery has been associated with an upregulation of inflammatory CKs such as IL-1β, IL-6, IL-8, and TNF-α [
3,
11]. It is well known that these CKs are important to lead to correct parturition, such as inducing the contraction of the smooth muscle in the uterus [
11,
37]. This inflammatory environment at the delivery day could be the reason why we found higher levels of IL-1β and TNF-α in UCP than in the MP. This result is in line with a recent report stablishing that fetal-placental variables are directly associated with changes in the CKs present in UCP [
38]. A successful pregnancy needs these pro-inflammatory events, but it is well known that an overexpression of pro-inflammatory CKs can disrupt fetal and placental developmental pathways [
3] and can promote pre-term parturition or other delivery disorders [
10]. On the other hand, the under-expression of those CKs is associated with adverse pregnancy outcomes. For example, Chehroudi et al. observed lower levels of IL-1β, IL-6, and TNF-α in umbilical cord lysates at birth in mothers suffering from pre-eclampsia and gonococcus infection [
38]. They also observed a decrease in IL-10 concentration.
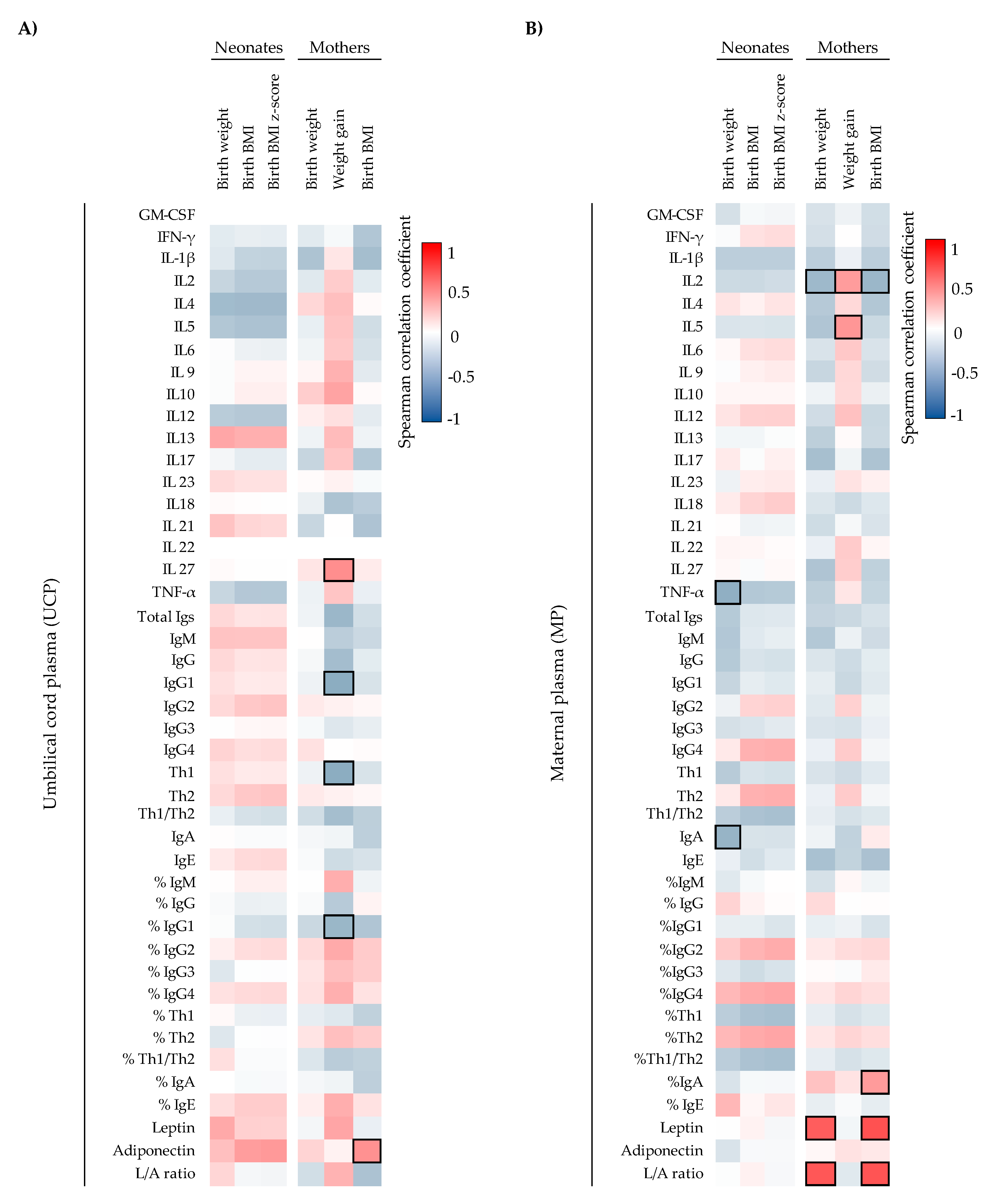
With regard to adipokine levels, we found three-fold higher levels of leptin in MP with respect to UCP, similarly to a study by Schubring et al. [
39], in which the leptin levels at 38–40 weeks of gestation were the highest in the entire studied period which corresponded to the first 6 weeks after birth. In contrast, adiponectin was higher in UCP than in MP. Moreover, we did not observe correlations between UCP and MP either for leptin or adiponectin, coinciding with results obtained by other authors [
18,
39]. Kotani et al. saw that UCP adiponectin concentration was higher than that from normal-weight adults and when the UCP adiponectin concentration was high, the mass of fetal fat also increased [
6]. Although we did not observe a correlation between UCP adiponectin and the weight of the neonate at birth, we observed a positive correlation between UCP adiponectin and the maternal BMI at the delivery day, suggesting that an increase in maternal mass fat could increase UCP adiponectin levels. Furthermore, we observed that the higher the maternal weight and BMI, the higher the maternal leptin and L/A ratio, linking with their weight gaining activity, as described previously in literature [
40].
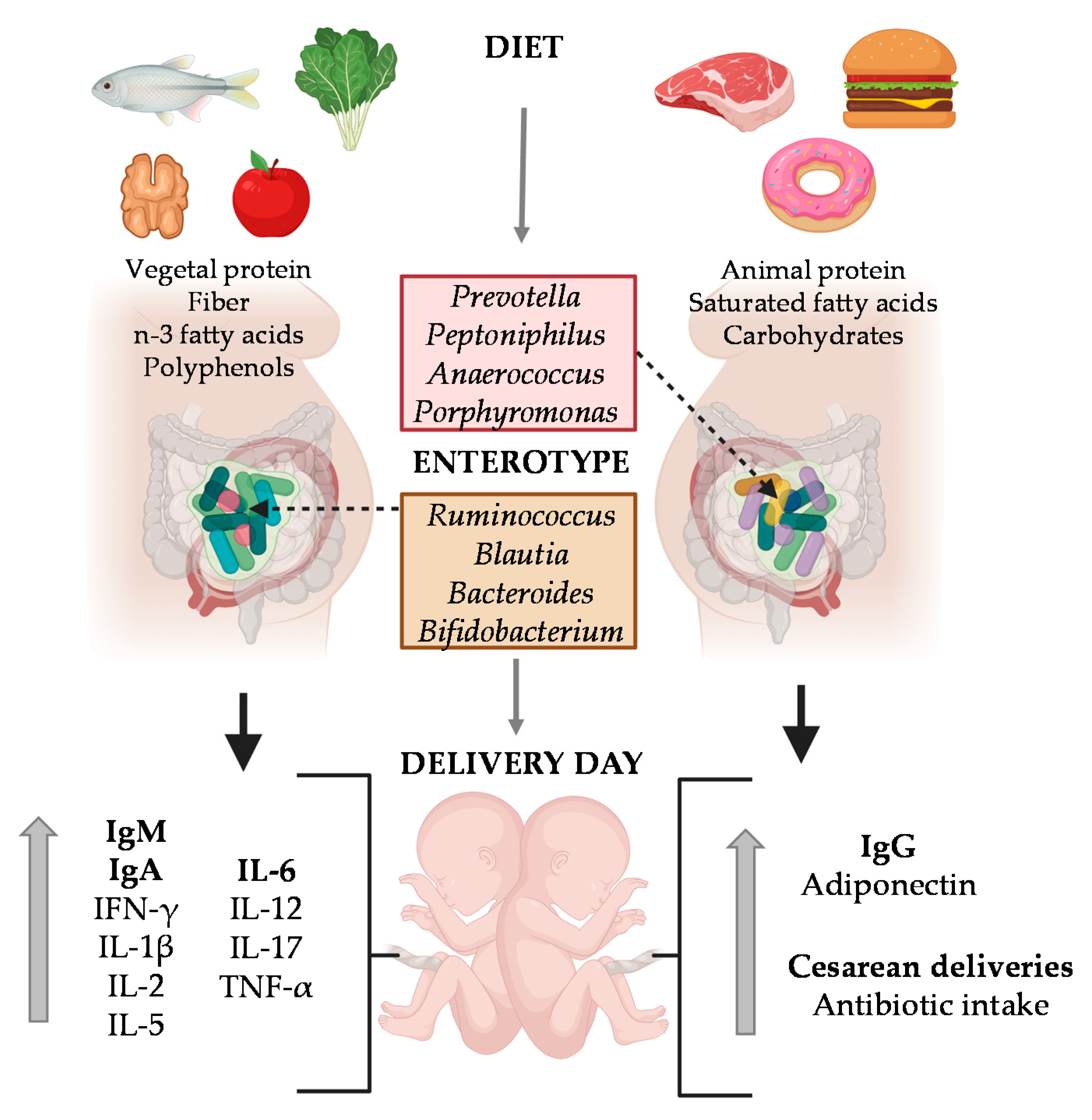
Previous studies in the same MAMI cohort showed the impact of specific dietary compounds such as dietary fiber, vegetable protein, polyphenols and lipids (mainly, the n-3 fatty acids DHA and DPA), their enterotype was characterized by predominance of the
Ruminococcaceae family (e.g.,
Ruminococcus),
Lachnospiraceae family (e.g.,
Blautia) and the genera
Bacteroides and
Bifidobacterium. On the other hand, when they biased towards higher intake of carbohydrates, saturated fatty acids and proteins (mainly animal protein) their enterotype was enriched in the genus
Prevotella,
Peptoniphilus,
Anaerococcus, and
Porphyromonas [
28].
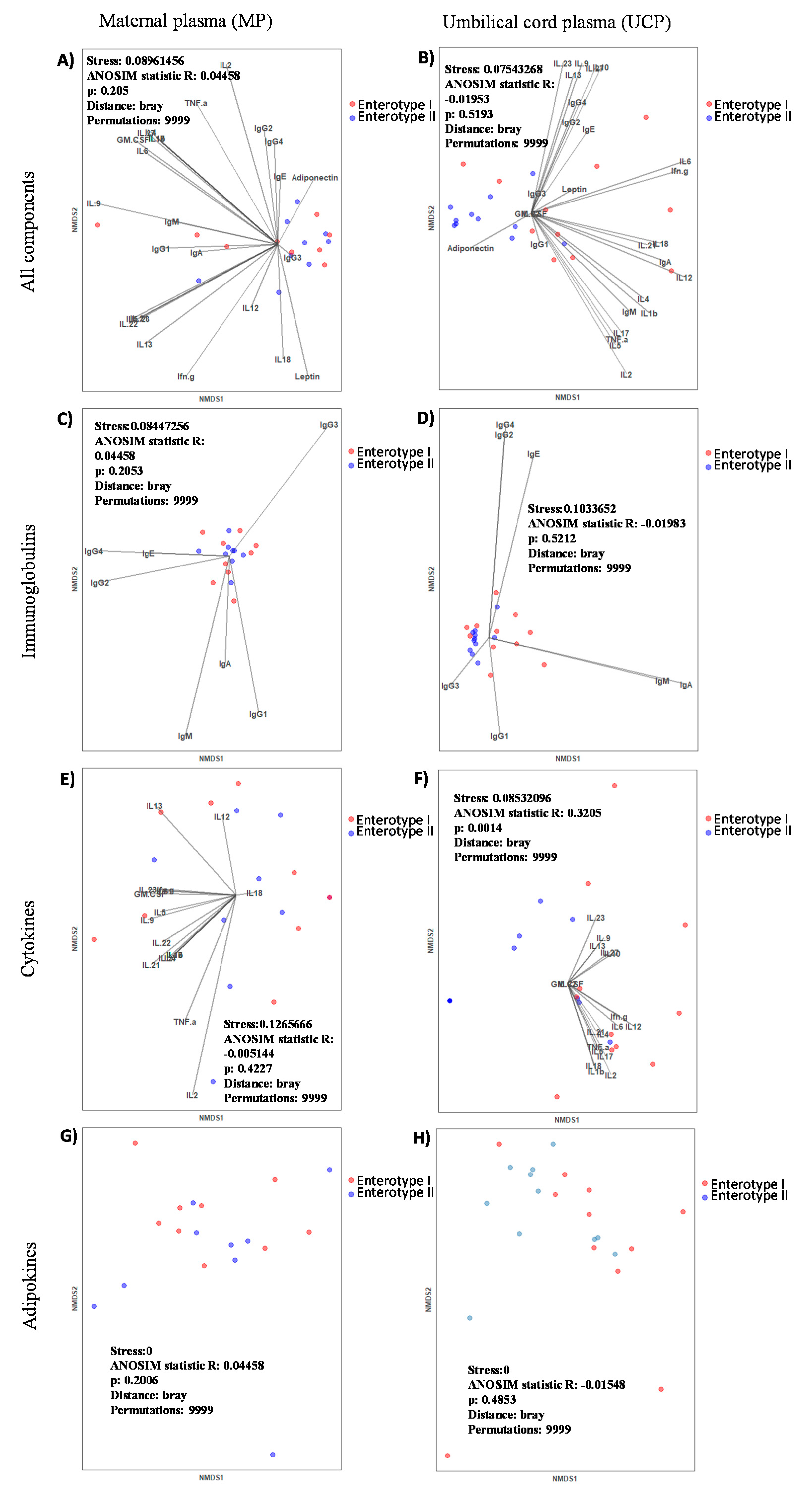
With regard to the influence of the maternal enterotype on MP composition, we can observe that the enterotype had a limited influence; only adiponectin was increased in the Enterotype II group. In contrast, we observed a great impact on UCP composition, suggesting that the gut microbial community of pregnant women affected the Ig, CK and adipokine levels of UCP, which in turn, could have had an impact during pregnancy. Moreover, mothers belonging to Enterotype II group displayed higher frequencies of antibiotic intake at birth and cesarean deliveries. Indeed, we suggest that having Enterotype II increases the susceptibility towards imbalanced immune factors, which could be influenced by the maternal diet and microbiota composition. The most important finding is that CKs, mainly the pro-inflammatory ones (IFN- γ, IL-1β, IL-6, IL-12, IL-17, and TNF-α), were decreased in the Enterotype II group at the day of delivery. These results are in line with the fact that pro-inflammatory CKs play an important role in a normal parturition [
3,
11,
38]. However, it has to be taken into account that the cesarean delivery proportion is higher in this enterotype, and in this situation, high levels of these CKs could not be required. However, the comparison of these factors between the mode of delivery for both the MP and the UCP in our limited number of samples ruled out this possible influence. Nevertheless, it would be interesting to deep into this CKs-enterotype association with higher number of participants, including similar levels of cesarean deliveries in both groups. It has also to be considered the possible influence that the intrapartum antibiotic used in the cesareans would also cause a shift in the maternal microbiota composition.
The Enterotype II group also showed lower levels of leptin and higher levels of adiponectin in MP, despite this change not being significant. In line with this, lower levels of leptin could indicate disorders during pregnancy, because it acts as an hormonal feedback loop that indicates a normal progression of pregnancy [
39]. This feedback loop can be done because the syncytiotrophoblast produces adequate amounts of placental leptin which is then secreted into the maternal circulation [
41,
42]. On the other hand, an increase in adiponectin in both MP and UCP in Enterotype II group was observed. Daryasari et al. observed differences on maternal plasma adiponectin levels depending on the mode of delivery [
43]. They found that umbilical cord blood from vaginal delivery had higher levels of adiponectin. However, in our study, we ruled out the possibility that the increase in MP adiponectin of the Enterotype II group was due to the predisposition to have cesarean delivery, because there was not a significant change comparing that component between the types of delivery for both the MP and the UCP.
Besides the maternal microbiota, the dietary components and specific nutrients have also been associated with the neonatal microbiota composition on previous studies with MAMI cohort [
27,
28]. Because in the present approach a clear association between the immune and adipokine factors and the maternal enterotype was found, the following step was to assess the impact of the maternal diet on immune and adipokine factors on MP and UCP. Surprisingly, more influence of dietary components on the immunological profile of the UCP than MP was found.
It is well established that a diet rich in vegetable protein, fish, fiber, and polyphenols improves healthy conditions both in adults and children [
44,
45,
46,
47,
48]. Moreover, a diet high in fiber meliorates an anti-inflammatory environment, and protects children from noninfectious colonic diseases and inflammations [
44], and improves the treatment of rheumatoid arthritis [
45]. The mechanism is still unknown, but fiber intake during pregnancy and lactation could be a key element in improving the infant development and promising health at short and long term. Dietary fiber components (non-digestible carbohydrates) are also broken down and oxidized incompletely by intestinal microbiota forming short-chain fatty acids (SCFA) [
49], which can reach the systemic compartment and, in turn, modulate both systemic and mucosal immune functions [
50,
51,
52] and regulate some metabolic aspects [
53]. There are evidences that the differentiation of T cells and the expression of IL-10, IL-17, and IFN-γ are supported by intestinal SCFA in mice [
54]. It is reported that n-3 PUFA supplementation is needed for pregnant women who do not eat fish, since their deficiency is associated with worse pregnancy outcomes such as low birth weight and preterm delivery [
55,
56]. Besides, Margherite Maranesi et al. suggested that reproductive performances such as fertility and pregnancy could be enhanced by PUFA supplementation [
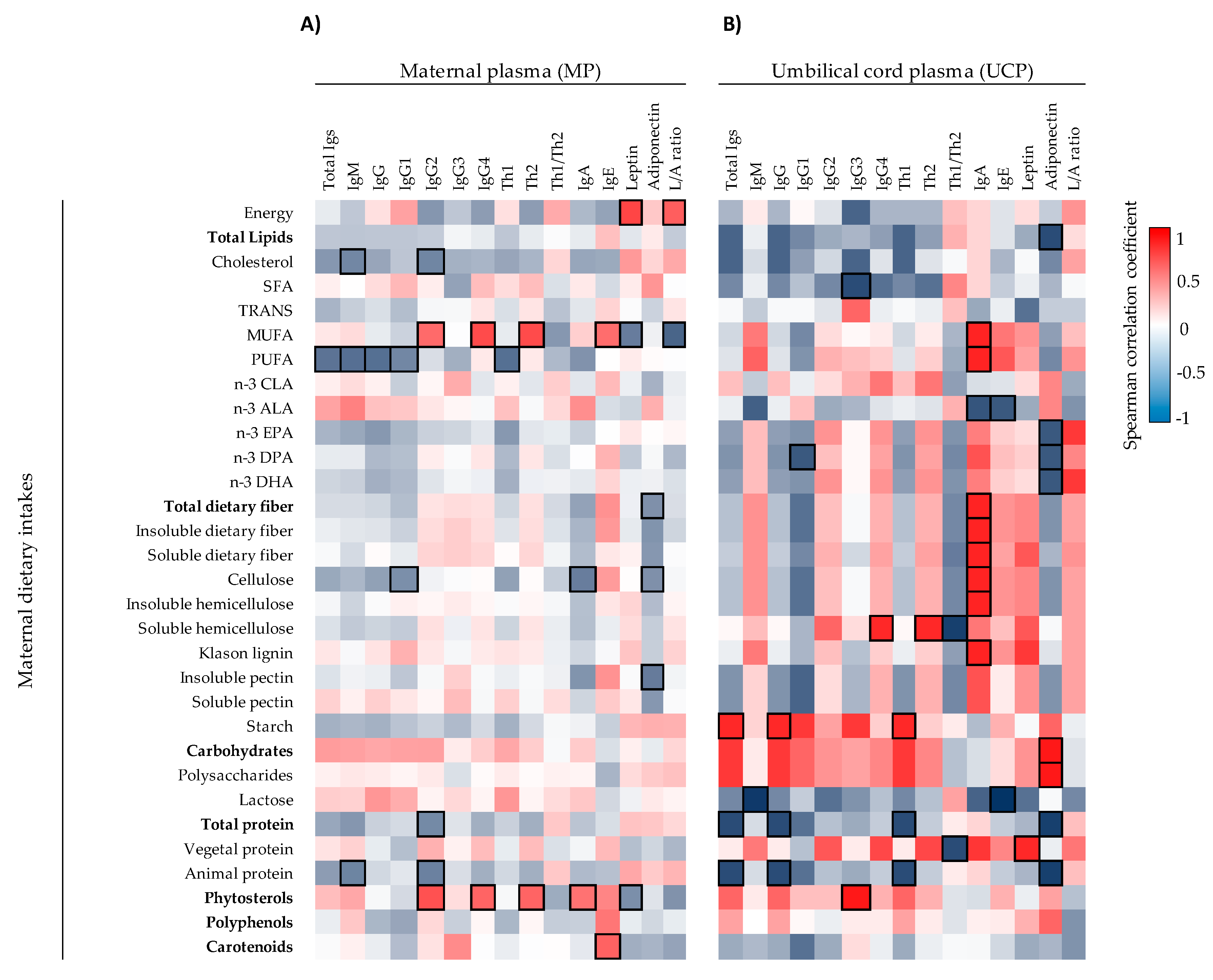
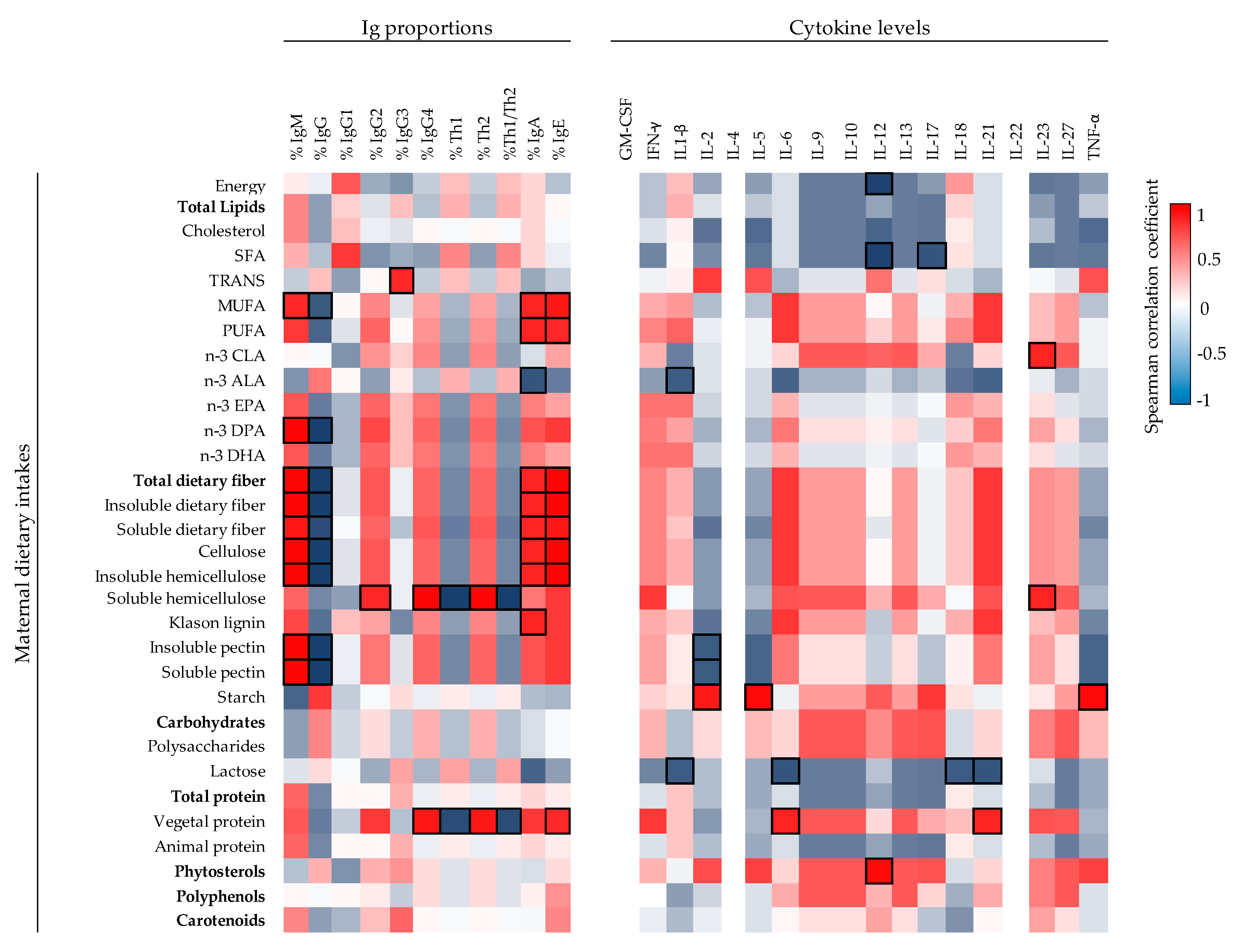
57]. There is little information available for the influence of maternal dietary components on neonatal immune factors in humans. This is the first study providing associations between maternal dietary intakes of (dietary fiber and other dietary components) with UCP immune factors. Fiber-type components (cellulose, hemicellulose, lignin, and both soluble and insoluble fiber) and MUFA and PUFA correlated positively with UCP IgA. These dietary components are related with the Enterotype I group in which there were higher levels of IgA, higher proportion of IgA and IgM, but less relative percentage of IgG on UCP. Besides this, MUFA and dietary fiber components correlated positively with UCP IgA, IgM, and IgE proportions and negatively with UCP IgG proportion. Overall, the maternal dietary fiber, possibly by raising the production and transplacental transfer of SCFA, seems to have a role in the developing mucosal immune defense of the neonate, although further studies should be performed to test this hypothesis.
Moreover, the maternal intake of lipids influences infant development due to the fatty acids fetal transfer [
58,
59]. During pregnancy, essential fatty acids (EFAs) are transported actively from the mother across the placenta, since the demand of EFAs (EPA, DHA and ALA) and long-chain polyunsaturated fatty acids (PUFAs) derivatives for fetal development is increased [
60,
61,
62]. Accordingly, our results may indicate that the lipids intake affects the immune and adipokine composition of UCP. As mentioned before, MUFA and PUFA impacted on Ig proportions. However, EFAs, specifically EPA, DPA, and DHA, were correlated negatively with adiponectin levels in UCP. This was also an interesting observation since Enterotype I group was associated with higher intake of DPA and DHA [
28] and less UCP adiponectin levels, suggesting that adiponectin could be influenced by EFAs transferred from the mothers, but also this adipokine could be influenced by maternal carbohydrates (polysaccharides), insoluble pectin and animal protein. Although UCP adiponectin has a tendency to decrease in Enterotype I group and the mothers from this group had lower animal protein intake, the association study showed a negative correlation. Another difference between the two enterotype groups was the intake of animal protein [
28]. Enterotype I group is characterized by a high intake of vegetal protein. According to the IL-6 increase observed in Enterotype I group, a positive correlation was also observed between vegetal protein and UCP IL-6 concentration, suggesting that the dietary component having an influence on the IL-6 levels in Enterotype I group could be the vegetal protein.
Among all the factors, those that coincide in the enterotype and diet analysis should receive more attention. Although more research elucidating the effect of immune factors on the development and health of the neonate is required, it is of great interest to find that the maternal diet and the maternal enterotype impact more on the levels of the UCP immune and adipokine factors than those of the MP. This is a very important fact since the neonate’s immune system is still developing and changes in these factors could determine the development, the birth, and the health later on. Mechanism of action studies that allow confirm these associations and additional comparative studies may help to provide strong information and new insights into maternal dietary interventions to improve infant development.
Finally, the limitations of this study include sample size, which could have affected the statistical power of the study and also, the low number of matched maternal-neonatal blood and fecal samples. Although the starting number of samples was low for the main objective but enough to obtain statistical differences, the sample dropouts affected the secondary objective outcomes. The details of the sample recruiting can be seen in the flow chart of
Supplementary Figure S1, in which can be observed that not all analysis were performed in all subjects. Secondly, the poor CK detectability, which is already described in the literature, also affected the quantitative interpretation of the results, which should be cautiously analyzed. Moreover, some concurring factors such as the Enterotype II and higher number of cesarean deliveries could have had and influence on the results shown here, even though we did not observe a significant effect on the immune components of the umbilical cord plasma. Despite these limitations, our study is a pioneer in providing novel and relevant data on the interactions between diet, microbiota and immunoglobulin and cytokine profiles. Our data provides the base for future studies as the topic warrants further investigation.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}