
Epigenetic Changes Governing Scn5a Expression in Denervated Skeletal Muscle


 , , and
, , and
Abstract
:1. Introduction
2. Results
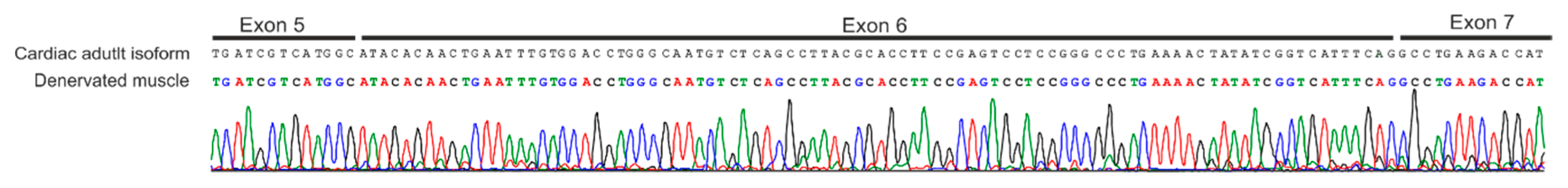
2.1. Denervated Gastrocnemius Muscle Expresses the Scn5a Adult Isoform
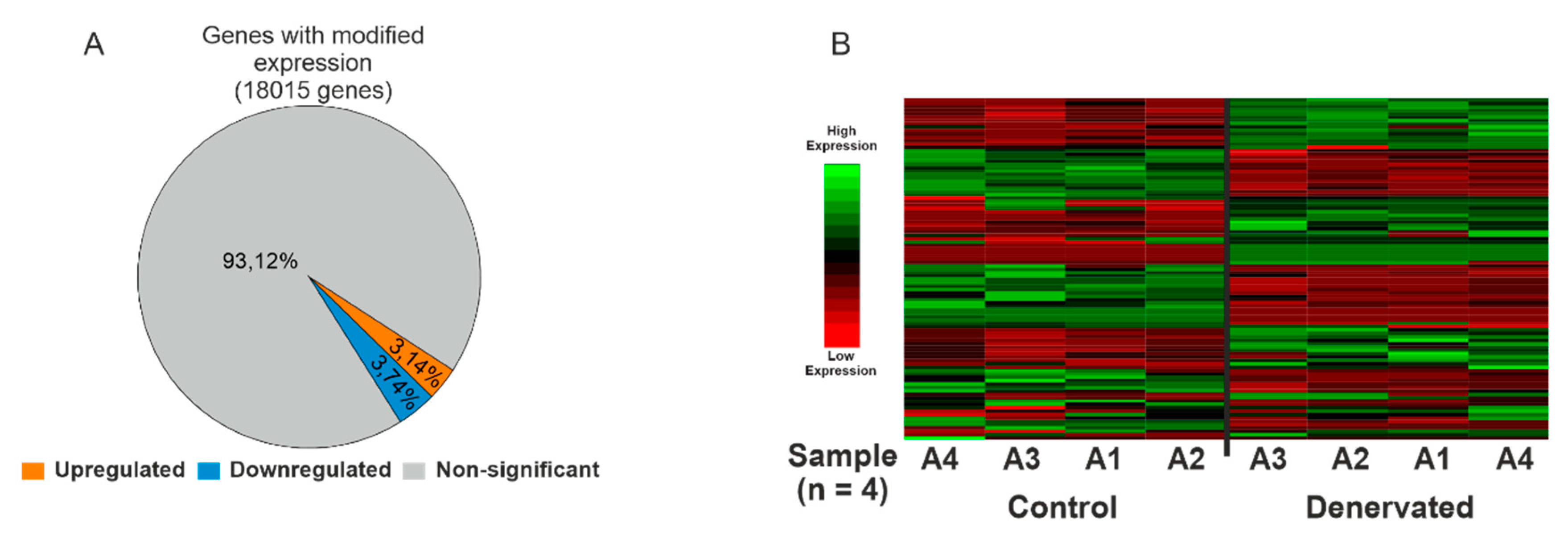
2.2. Skeletal Muscle Denervation Leads to Significant Changes in the Transcriptome
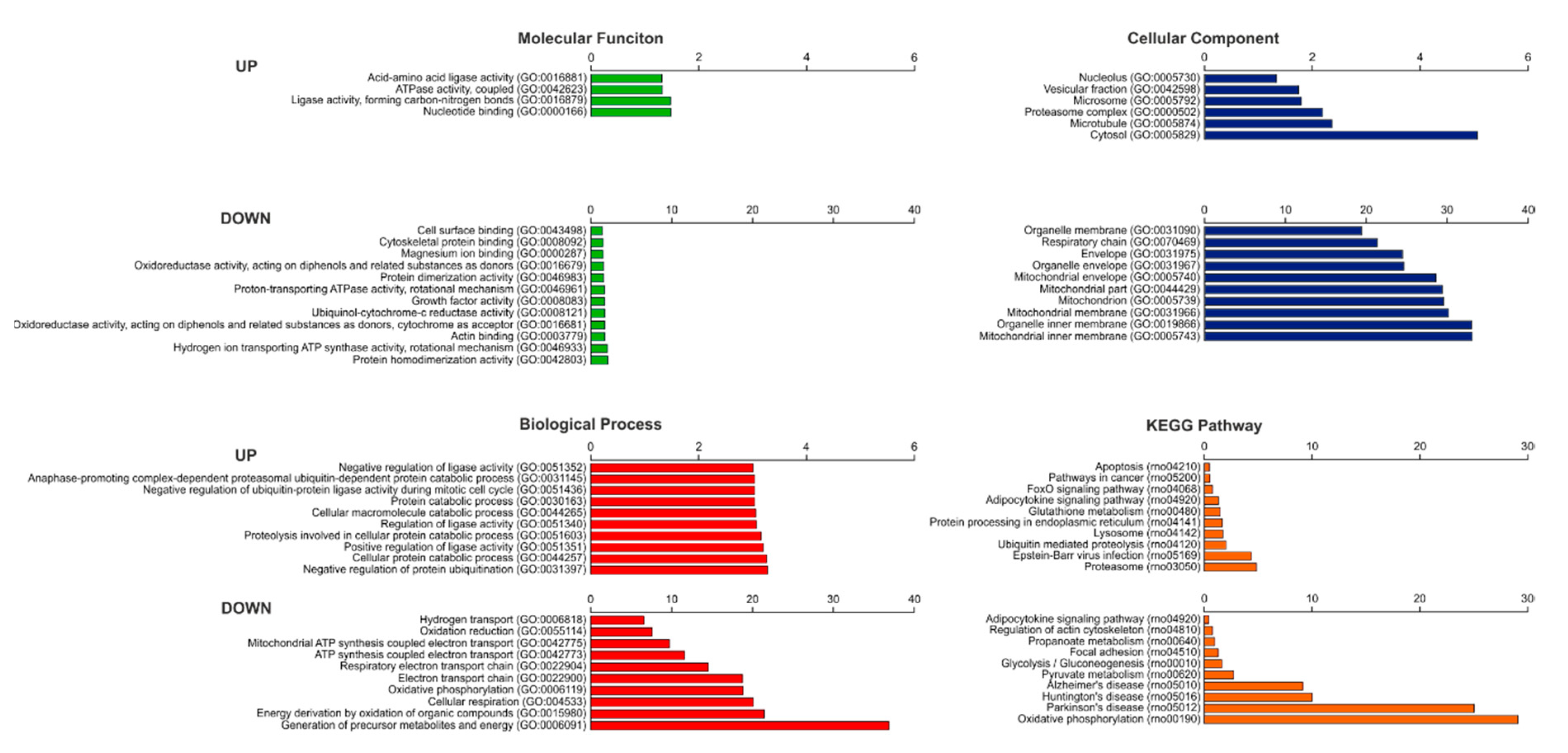
2.3. Re-Expression of Scn5a Is Not Related to any Functional Annotation Based on Gene Ontology (GO) or Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathways
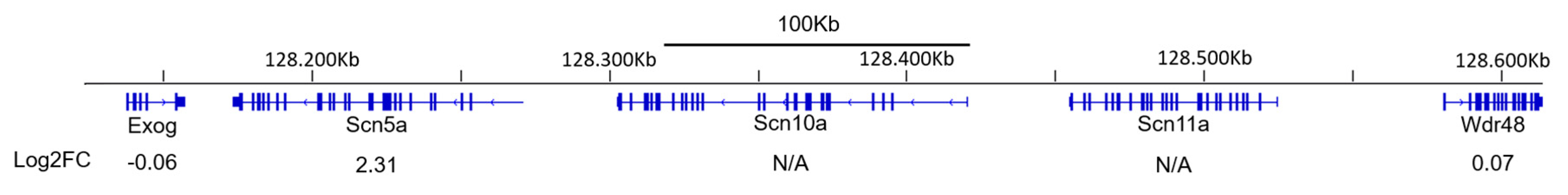
2.4. Scn5a TAD Neighboring Genes and Other Sodium Channels Are not Upregulated Upon Muscle Denervation
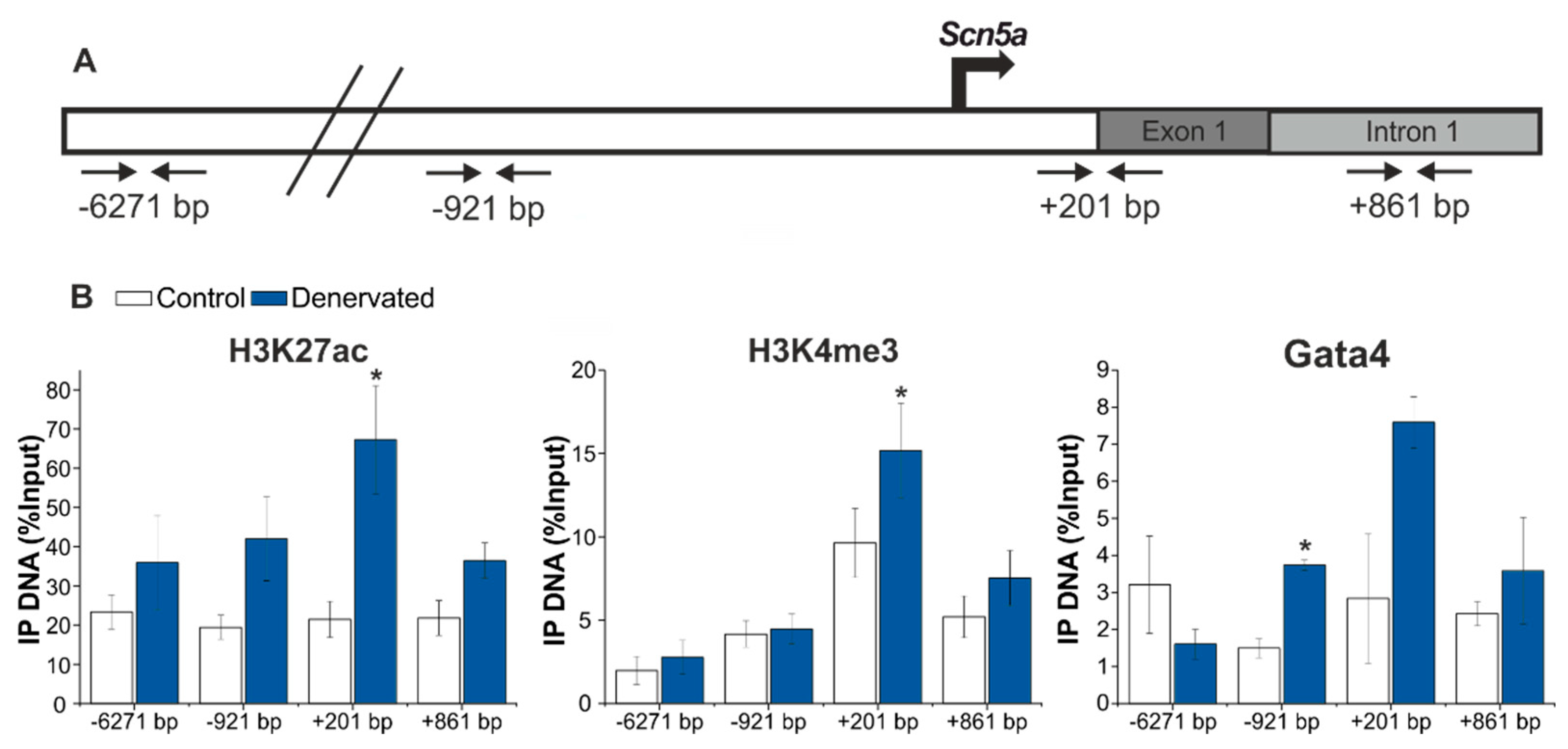
2.5. Denervation Is Associated with an Enrichment of H3K27ac and H3K4me3 and Gata4 Binding at the Scn5a Promoter Region
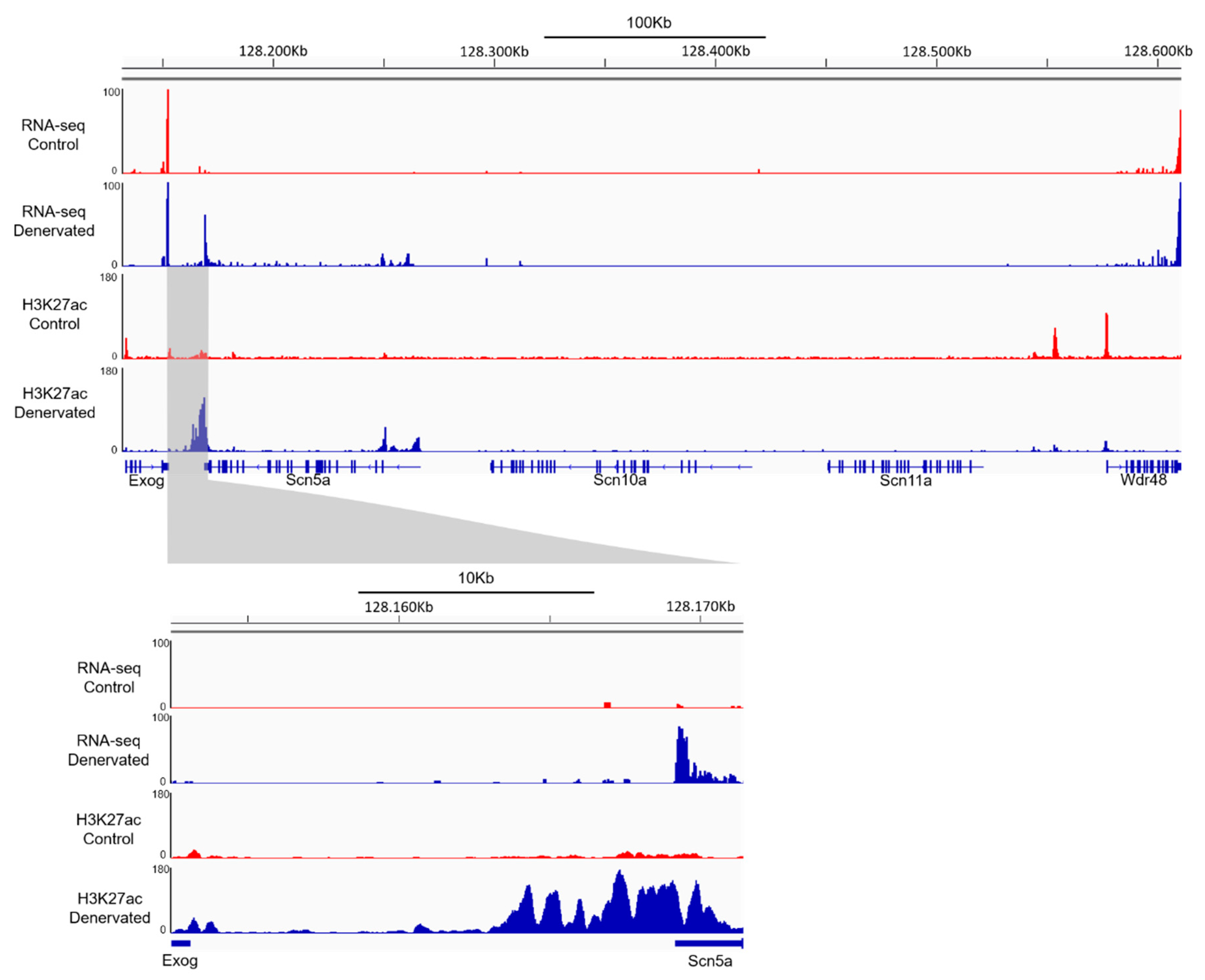
2.6. Scn5a Re-Expression in Skeletal Muscle Is Associated with Activation of a Super Enhancer Downstream of the Scn5a Gene
3. Discussion
4. Materials and Methods
4.1. Rat Skeletal Muscle Denervation and Sample Collection
4.2. RNA Isolation, cDNA Synthesis, and Scn5a Sequencing
4.3. RNA-Seq and Data Analysis
4.4. Immunoprecipitation Experiments and Western Blot
4.5. ChIP-qPCR Assays
4.6. ChIP-Seq Assays
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BP | Biological process |
| BrS | Brugada syndrome |
| CC | Cellular component |
| ChIP | Chromatin immunoprecipitation |
| DE | Differentially expressed |
| FC | Fold change |
| H3K27ac | Histone H3 lysine (K) 27 acetylation |
| H3K4me3 | Histone H3 lysine (K) 4 trimethylation |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MF | Molecular function |
| RE | Regulatory elements |
| RNA-seq | RNA sequencing |
| TAD | Topological associated domain |
| TF | Transcription factor |
| TSS | Transcription start site |
| TTX | Tetrodotoxin |
| VGSC | Voltage-gated sodium channel/s |
References
- Trimmer, J.; Cooperman, S.; Agnew, W.; Mandel, G. Regulation of Muscle Sodium Channel Transcripts during Development and in Response to Denervation. Dev. Biol. 1990, 142, 360–367. [Google Scholar] [CrossRef]
- Wood, J.N.; Iseppon, F. Sodium channels. Brain Neurosci. Adv. 2018, 2, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y.; Liu, N.; Priori, S.G. Sodium channel mutations and arrhythmias. Nat. Rev. Cardiol. 2009, 6, 337–348. [Google Scholar] [CrossRef]
- Brackenbury, W.J.; Isom, L.L. Na+ channel β subunits: Overachievers of the ion channel family. Front. Pharm. 2011, 2, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rook, M.B.; Evers, M.M.; Vos, M.A.; Bierhuizen, M.F.A. Biology of cardiac sodium channel Nav1.5 expression. Cardiovasc. Res. 2012, 93, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Renganathan, M.; Dib-Hajj, S.; Waxman, S.G. Nav1.5 Underlies the ‘Third TTX-R Sodium Current’ in Rat Small DRG Neurons. Mol. Brain Res. 2002, 106, 70–82. [Google Scholar] [CrossRef]
- Ou, Y.; Gibbons, S.J.; Miller, S.M.; Strege, P.R.; Rich, A.; Distad, M.A.; Ackerman, M.J.; Rae, J.L.; Szurszewski, J.H.; Farrugia, G. SCN5A is expressed in human jejunal circular smooth muscle cells. Neurogastroenterol. Motil. 2002, 14, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Strege, P.R.; Ou, Y.; Sha, L.; Rich, A.; Gibbons, S.J.; Szurszewski, J.H.; Sarr, M.G.; Farrugia, G. Sodium current in human intestinal interstitial cells of Cajal. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, 1111–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, H.A.; Colom, L.V.; Sutherland, M.L.; Noebels, J.L. Selective localization of cardiac SCN5A sodium channels in limbic regions of rat brain. Nat. Neurosci. 1999, 2, 593–595. [Google Scholar] [CrossRef]
- Wu, L.; Nishiyama, K.; Hollyfield, J.G.; Wang, Q. Localization of Nav1.5 sodium channel protein in the mouse brain. Neuroreport 2002, 13, 2547–2551. [Google Scholar] [CrossRef] [Green Version]
- Scornik, F.S.; Desai, M.; Brugada, R.; Guerchicoff, A.; Pollevick, G.D.; Antzelevitch, C.; Pérez, G.J. Functional expression of “cardiac-type” Nav1.5 sodium channel in canine intracardiac ganglia. Heart Rhythm 2006, 3, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallen, R.; Sheng, Z.; Yang, J.; Chen, L.; Rogart, R.; Barchi, R. Primary Structure and Expression of a Sodium Channel Characteristic of Denervated and Immature Rat Skeletal Muscle. Neuron 1990, 4, 233–242. [Google Scholar] [CrossRef]
- Redfern, P.; Lundh, H.; Thesleff, S. Tetrodotoxin resistant action potentials in denervated rat skeletal muscle. Eur. J. Pharm. 1970, 11, 263–265. [Google Scholar] [CrossRef]
- Redfern, P.; Thesleff, S. Action Potential Generation in Denervated Rat Skeletal Muscle. Acta Physiol. Scand. 1971, 82, 70–78. [Google Scholar] [CrossRef]
- Harris, J.B.; Thesleff, S. Studies on Tetrodotoxin Resistant Action Potentials in Denervated Skeletal Muscle. Acta Physiol. Scand. 1971, 83, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Pappone, P.A. Voltage-clamp experiments in normal and denervated mammalian skeletal muscle fibres. J. Physiol. 1980, 306, 377–410. [Google Scholar] [CrossRef] [Green Version]
- Rogart, R.B.; Cribbs, L.L.; Muglia, L.K.; Kephart, D.D.; Kaiser, M.W. Molecular cloning of a putative tetrodotoxin-resistant rat heart Na+ channel isoform. Proc. Nati. Acad. Sci. USA 1989, 86, 8170–8174. [Google Scholar] [CrossRef] [Green Version]
- White, M.M.; Chen, L.; Kleinfield, R.; Kallen, R.G.; Barchi, R.L. SkM2, a Na+ channel cDNA clone from denervated skeletal muscle, encodes a tetrodotoxin-insensitive Na+ channel. Mol. Pharm. 1991, 39, 604–608. [Google Scholar]
- Sellin, L.C.; Thesleff, S. Alterations in membrane electrical properties during long-term denervation of rat skeletal muscle. Acta Physiol. Scand. 1980, 108, 243–246. [Google Scholar] [CrossRef]
- Yang, J.S.; Sladky, J.T.; Kallen, R.C.; Barchi, R.L. TTX-Sensitive and TTX-Insensitive Sodium Channel mRNA Transcripts Are Independently Regulated in Adult Skeletal Muscle after Denervation. Neuron 1991, 7, 421–427. [Google Scholar] [CrossRef]
- Sheng, Z.-H.; Zhang, H.; Barchi, R.L.; Kallen, R.G. Molecular Cloning and Functional Analysis of the Promoter of Rat Skeletal Muscle Voltage-Sensitive Sodium Channel Subtype 2 (rSkM2): Evidence for Muscle-Specific Nuclear Protein Binding to the Core Promoter. DNA Cell Biol. 2009, 13, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.M.; Zheng, Z.; Messi, M.L.; Delbono, O. Extension and magnitude of denervation in skeletal muscle from ageing mice. J. Physiol. 2005, 565, 757–764. [Google Scholar] [CrossRef]
- Rowan, S.L.; Rygiel, K.; Purves-Smith, F.M.; Solbak, N.M.; Turnbull, D.M.; Hepple, R.T. Denervation causes fiber atrophy and myosin heavy chain co-expression in senescent skeletal muscle. PLoS ONE 2012, 7, e29082. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, K.; Kanda, F.; Mitsui, S.; Kohara, N.; Chihara, K. Fibrillation potentials of denervated rat skeletal muscle are associated with expression of cardiac-type voltage-gated sodium channel isoform Nav1.5. Clin. Neurophysiol. 2012, 123, 1650–1655. [Google Scholar] [CrossRef]
- Nielsen, M.W.; Holst, A.G.; Olesen, S.-P.; Olesen, M.S. The genetic component of brugada syndrome. Front. Physiol. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kapplinger, J.D.; Tester, D.J.; Alders, M.; Benito, B.; Berthet, M.; Brugada, J.; Brugada, P.; Fressart, V.; Guerchicoff, A.; Harris-Kerr, C.; et al. An international compendium of mutations in the SCN5A-encoded cardiac sodium channel in patients referred for Brugada syndrome genetic testing. Heart Rhythm 2010, 7, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, T.; Surber, R. SCN5A channelopathies—An update on mutations and mechanisms. Prog. Biophys. Mol. Biol. 2008, 98, 120–136. [Google Scholar] [CrossRef]
- Leoni, A.-L.; Gavillet, B.; Rougier, J.-S.; Marionneau, C.; Probst, V.; Le Scouarnec, S.; Schott, J.-J.; Demolombe, S.; Bruneval, P.; Huang, C.L.H.; et al. Variable Nav1.5 protein expression from the wild-type allele correlates with the penetrance of cardiac conduction disease in the Scn5a +/− mouse model. PLoS ONE 2010, 5, e9298. [Google Scholar] [CrossRef] [Green Version]
- Bezzina, C.R.; Shimizu, W.; Yang, P.; Koopmann, T.T.; Tanck, M.W.T.; Miyamoto, Y.; Kamakura, S.; Roden, D.M.; Wilde, A.A.M. Common Sodium Channel Promoter Haplotype in Asian Subjects Underlies Variability in Cardiac Conduction. Circulation 2006, 113, 338–344. [Google Scholar] [CrossRef] [Green Version]
- Van Den Boogaard, M.; Wong, L.Y.E.; Tessadori, F.; Bakker, M.L.; Dreizehnter, L.K.; Wakker, V.; Bezzina, C.R.; AC’t Hoen, P.A.C.; Bakkers, J.; Barnett, P.; et al. Genetic variation in T-box binding element functionally affects SCN5A / SCN10A enhancer. J. Clin. Investig. 2012, 122, 2519–2530. [Google Scholar] [CrossRef] [Green Version]
- Van Den Boogaard, M.; Smemo, S.; Burnicka-Turek, O.; Arnolds, D.E.; van de Werken, H.J.G.; Klous, P.; McKean, D.; Muehlschlegel, J.D.; Moosmann, J.; Toka, O.; et al. A common genetic variant within SCN10A modulates cardiac SCN5A expression. J. Clin. Investig. 2014, 124, 1844–1852. [Google Scholar] [CrossRef] [Green Version]
- Tarradas, A.; Pinsach-Abuin, M.; Mackintosh, C.; Brugada, R.; Beltran-Alvarez, P.; Pagans, S. Transcriptional regulation of the sodium channel gene (SCN5A) by GATA4 in human heart. J. Mol. Cell. Cardiol. 2017, 102, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Sayed, Z.R.; Canac, R.; Cimarosti, B.; Bonnard, C.; Gourraud, J.-B.; Hamamy, H.; Kayserili, H.; Girardeau, A.; Jouni, M.; Jacob, N.; et al. Human model of IRX5 mutations reveals key role for this transcription factor in ventricular conduction. Cardiovasc. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurman, R.E.; Rynes, E.; Humbert, R.; Vierstra, J.; Maurano, M.T.; Haugen, E.; Sheffield, N.C.; Stergachis, A.B.; Wang, H.; Vernot, B.; et al. The accessible chromatin landscape of the human genome. Nature 2012, 489, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Zaglia, T.; Dedja, A.; Candiotto, C.; Cozzi, E.; Schiaffino, S.; Ausoni, S. Cardiac interstitial cells express GATA4 and control dedifferentiation and cell cycle re-entry of adult cardiomyocytes. J. Mol. Cell. Cardiol. 2009, 46, 653–662. [Google Scholar] [CrossRef]
- Hernandez, G. (Caltech) Building a Gene Regulatory Network in Adult Mouse Skeletal Muscle Following Nerve Injury: Transcriptome Characterization and New Model for Functional cis-Regulatory Analysis in Vivo. Ph.D. Thesis, California Institute of Technology, Pasadena, CA, USA, 2013. [Google Scholar]
- Cohen, T.J.; Waddell, D.S.; Barrientos, T.; Lu, Z.; Feng, G.; Cox, G.A.; Bodine, S.C.; Yao, T.P. The histone deacetylase HDAC4 connects neural activity to muscle transcriptional reprogramming. J. Biol. Chem. 2007, 282, 33752–33759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Kupershmidt, S.; Roden, D.M. Cloning and initial characterization of the human cardiac sodium channel (SCN5A) promoter. Cardiovasc. Res. 2004, 61, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Man, J.C.K.; Mohan, R.A.; van den Boogaard, M.; Hilvering, C.R.E.; Jenkins, C.; Wakker, V.; Bianchi, V.; de Laat, W.; Barnett, P.; Boukens, B.J.; et al. An enhancer cluster controls gene activity and topology of the SCN5A-SCN10A locus in vivo. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, S.P.; Diss, J.K.J.; Chioni, A.-M.; Mycielska, M.E.; Pan, H.; Yamaci, R.F.; Pani, F.; Siwy, Z.; Krasowska, M.; Grzywna, Z.; et al. Voltage-gated sodium channel expression and potentiation of human breast cancer metastasis. Clin. Cancer Res. 2005, 11, 5381–5389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.; Yoon, B.C.; Holt, C.E. Axonal mRNA localization and local protein synthesis in nervous system assembly, maintenance and repair. Nat. Rev. Neurosci. 2012, 13, 308–324. [Google Scholar] [CrossRef] [Green Version]
- Thakor, D.; Lin, A.; Matsuka, Y.; Meyer, E.M.; Ruangsri, S.; Nishimura, I.; Spigelman, I. Increased peripheral nerve excitability and local NaV1.8 mRNA up-regulation in painful neuropathy. Mol. Pain 2009, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Van Tuyn, J.; Atsma, D.E.; Winter, E.M.; van der Velde-van Dijke, I.; Pijnappels, D.A.; Bax, N.A.M.; Knaän-Shanzer, S.; Gittenberger-de Groot, A.C.; Poelmann, R.E.; van der Laarse, A.; et al. Epicardial Cells of Human Adults Can Undergo an Epithelial-to-Mesenchymal Transition and Obtain Characteristics of Smooth Muscle Cells In Vitro. Stem Cells 2007, 25, 271–278. [Google Scholar] [CrossRef]
- Peterkin, T.; Gibson, A.; Patient, R. Redundancy and evolution of GATA factor requirements in development of the myocardium. Dev. Biol. 2007, 311, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.T.; Morrisey, E.E.; Anandappa, R.; Sigrist, K.; Lu, M.M.; Parmacek, M.S.; Soudais, C.; Leiden, J.M. GATA4 Transcription Factor Is Required for Ventral Morphogenesis and Heart Tube Formation. Genes Dev 1997, 11, 1048–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopal, S.K.; Ma, Q.; Obler, D.; Shen, J.; Manichaikul, A.; Tomita-Mitchell, A.; Boardman, K.; Briggs, C.; Garg, V.; Srivastava, D.; et al. Spectrum of heart disease associated with murine and human GATA4 mutation. J. Mol. Cell. Cardiol. 2007, 43, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munshi, N.V.; McAnally, J.; Bezprozvannaya, S.; Berry, J.M.; Richardson, J.A.; Hill, J.A.; Olson, E.N. Cx30.2 enhancer analysis identifies Gata4 as a novel regulator of atrioventricular delay. Development 2009, 136, 2665–2674. [Google Scholar] [CrossRef] [Green Version]
- He, A.; Won, S.; Ma, Q.; Pu, W.T. Co-occupancy by multiple cardiac transcription factors identifies transcriptional enhancers active in heart. Proc. Natl. Acad. Sci. USA 2011, 108, 5632–5637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, A.; Gu, F.; Hu, Y.; Ma, Q.; Yi, Y.L.; Akiyama, J.A.; Visel, A.; Pennacchio, L.A.; Pu, W.T. Dynamic GATA4 enhancers shape the chromatin landscape central to heart development and disease. Nat. Commun. 2014, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Log2 FC | p-Value | |
|---|---|---|---|
| α sodium channel subunits | Scn1a | −1.98 | 4.57 × 10−4 |
| Scn2a | −0.25 | NS | |
| Scn3a | 0.17 | NS | |
| Scn4a | 0.13 | NS | |
| Scn5a | 2.31 | 4.29 × 10−5 | |
| Scn7a | −1.08 | NS | |
| Scn8a | 0.05 | NS | |
| Scn9a | −0.25 | NS | |
| β sodium channel subunits | Scn1b | 0.34 | NS |
| Scn2b | 0.11 | NS | |
| Scn3b | −0.93 | NS | |
| Scn4b | −0.45 | NS | |
| Transcription regulators | Arx | −2.37 | 2.58 × 10−16 |
| Fos | −2.02 | 2.58 × 10−15 | |
| Foxp3 | −2.07 | 1.53 × 10−4 | |
| Gata4 | 2.36 | 2.27 × 10−5 | |
| Msx3 | −2.43 | 1.25 × 10−5 | |
| Myog | 2.93 | 3.85 × 10−61 | |
| Nfkb2 | 2.04 | 1.04 × 10−22 | |
| Nlrc5 | 2.55 | 2.06 × 10−13 | |
| Nr4a1 | −3.16 | 2.15 × 10−33 | |
| Nr4a2 | −2.4 | 1.49 × 10−9 | |
| Perm1 | −3.69 | 2.49 × 10−33 | |
| Runx1 | 2.76 | 1.16 × 10−16 | |
| Tgif1 | 2.06 | 3.16 × 10−20 | |
| Tmem229a | −2 | 5.16 × 10−4 | |
| Epigenetic modifiers | Hdac4 | 2.23 | 1.08 × 10−12 |
| Chromosome | Start | End | Gene | Log2fc ChIP-seq | Log2fc RNA-seq | p-Adj RNA-seq |
|---|---|---|---|---|---|---|
| 8 | 128,153,123 | 128,170,975 | Exog | 3.27 | - | - |
| Scn5a | 3.27 | 2.31 | 4.29 × 10−5 | |||
| 4 | 97,879,010 | 97,895,598 | 2.33 | - | - | |
| 2 | 188,844,059 | 188,877,449 | Kcnn3 | 1.89 | 2.15 | 3.32 × 10−20 |
| 2 | 34,586,496 | 34,618,138 | Adamts6 | 1.85 | 0.16 | NS |
| 9 | 119,081,166 | 119,099,933 | Dlgap1 | 1.84 | 0.75 | NS |
| LOC102555426 | 1.84 | - | - | |||
| 2 | 188,892,238 | 188,924,410 | Kcnn3 | 1.83 | 2.15 | 3.32 × 10−20 |
| 3 | 151,407,846 | 151,445,826 | Uqcc1 | 1.76 | 0.06 | NS |
| 1 | 140,978,031 | 141,044,113 | Abhd2 | 1.67 | 2.98 | 7.94 × 10−23 |
| 1 | 175,572,393 | 175,629,876 | LOC103691204 | 1.61 | - | - |
| Ampd3 | 1.61 | 1.94 | 3.46 × 10−4 | |||
| rnf141 | 1.61 | 0.33 | NS | |||
| 2 | 203,895,667 | 203,947,339 | 1.60 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreras, D.; Martinez-Moreno, R.; Pinsach-Abuin, M.; Santafe, M.M.; Gomà, P.; Brugada, R.; Scornik, F.S.; Pérez, G.J.; Pagans, S. Epigenetic Changes Governing Scn5a Expression in Denervated Skeletal Muscle. Int. J. Mol. Sci. 2021, 22, 2755. https://doi.org/10.3390/ijms22052755
Carreras D, Martinez-Moreno R, Pinsach-Abuin M, Santafe MM, Gomà P, Brugada R, Scornik FS, Pérez GJ, Pagans S. Epigenetic Changes Governing Scn5a Expression in Denervated Skeletal Muscle. International Journal of Molecular Sciences. 2021; 22(5):2755. https://doi.org/10.3390/ijms22052755
Chicago/Turabian StyleCarreras, David, Rebecca Martinez-Moreno, Mel·lina Pinsach-Abuin, Manel M. Santafe, Pol Gomà, Ramon Brugada, Fabiana S. Scornik, Guillermo J. Pérez, and Sara Pagans. 2021. "Epigenetic Changes Governing Scn5a Expression in Denervated Skeletal Muscle" International Journal of Molecular Sciences 22, no. 5: 2755. https://doi.org/10.3390/ijms22052755
APA StyleCarreras, D., Martinez-Moreno, R., Pinsach-Abuin, M., Santafe, M. M., Gomà, P., Brugada, R., Scornik, F. S., Pérez, G. J., & Pagans, S. (2021). Epigenetic Changes Governing Scn5a Expression in Denervated Skeletal Muscle. International Journal of Molecular Sciences, 22(5), 2755. https://doi.org/10.3390/ijms22052755