
β3-Adrenoreceptors as ROS Balancer in Hematopoietic Stem Cell Transplantation


{kind=link}
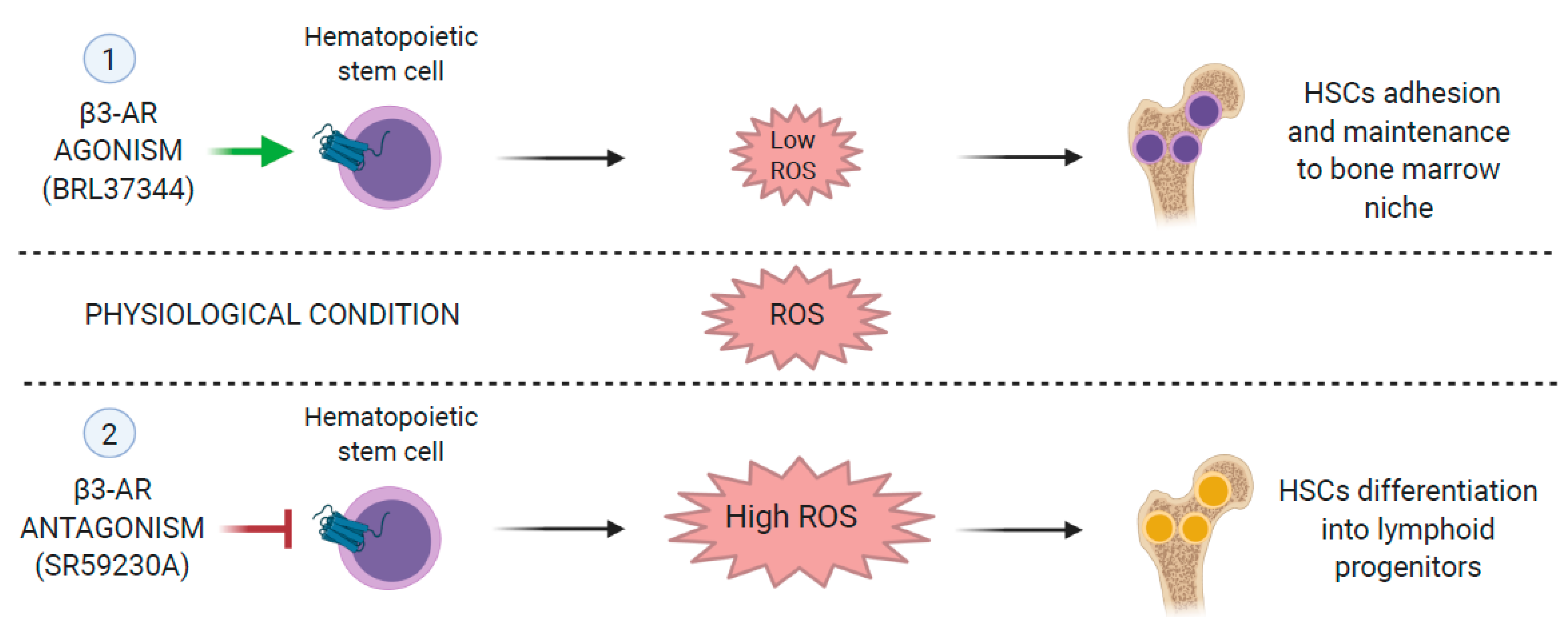{kind=link}
Abstract
:1. Introduction
Hematopoietic Stem Cell Transplantation
2. Reactive Oxygen Species
HSCT and ROS
3. β3-Adrenoreceptor
3.1. β3-ARs and ROS
3.2. β3-ARs and HSCs
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BM | Bone marrow |
| β3-AR | β3-adrenoreceptor |
| GSH | Glutathione |
| GvHD | Graft-versus-host disease |
| HSCs | Hematopoietic stem cells |
| HSCT | Hematopoietic stem cell transplantation |
| HSPCs | Hematopoietic progenitor stem cells |
| MDSC | Myeloid-derived suppressor cell |
| mtROS | Mitochondrial ROS |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NK | Natural killer |
| OS | Oxidative stress |
| PB | Peripheral blood |
| PBCs | Peripheral blood cells |
| PBSCT | Peripheral blood stem cells transplantation |
| ROS | Reactive oxygen species |
| UCP2 | Uncoupling protein-2 |
References
- Ludin, A.; Gur-Cohen, S.; Golan, K.; Kaufmann, K.B.; Itkin, T.; Medaglia, C.; Lu, X.; Ledergor, G.; Kollet, O.; Lapidot, T. Reactive Oxygen Species Regulate Hematopoietic Stem Cell Self-Renewal, Migration and Development, as well as Their Bone Marrow Microenvironment. Antioxid. Redox Signal. 2014, 21, 1605–1619. [Google Scholar] [CrossRef] [Green Version]
- Mahla, R.S. Stem Cells Applications in Regenerative Medicine and Disease Therapeutics. Int. J. Cell Biol. 2016, 2016, 6940283. [Google Scholar] [CrossRef] [Green Version]
- Lapidot, T.; Dar, A.; Kollet, O. How do stem cells find their way home? Blood 2005, 106, 1901–1910. [Google Scholar] [CrossRef] [Green Version]
- Fagioli, F.; Zecca, M.; Locatelli, F.; Lanino, E.; Uderzo, C.; Di Bartolomeo, P.; Berger, M.; Favre, C.; Rondelli, R.; Andrea, P.; et al. Allogeneic stem cell transplantation for children with acute myeloid leukemia in second complete remission. J. Pediatr. Hematol. Oncol. 2008, 30, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felfly, H.; Haddad, G.G. Hematopoietic stem cells: Potential new applications for translational medicine. J. Stem Cells 2014, 9, 163–197. [Google Scholar]
- Park, B.; Yoo, K.H.; Kim, C. Hematopoietic stem cell expansion and generation: The ways to make a breakthrough. Blood Res. 2015, 50, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Angheben, A.; Giaconi, E.; Menconi, M.; Casazza, G.; Najajreh, M.; Anselmi, M.; Gobbi, F.; Bisoffi, Z.; Tascini, C.; Favre, C. Reactivation of Chagas disease after a bone marrow transplant in Italy: First case report. Blood Transfus. 2012, 10, 542–544. [Google Scholar]
- Center for International Blood and Marrow Transplant Research. CIBMTR Summary Slides I. Archived from the Original; Center for International Blood and Marrow Transplant Research: Milwaukee, WI, USA, 2012. [Google Scholar]
- Langereis, E.J.; den Os, M.M.; Breen, C.; Jones, S.A.; Knaven, O.C.; Mercer, J.; Weston, M.; Paula, M.K.; Jim, K.; Tyler, K.; et al. Progression of Hip Dysplasia in Mucopolysaccharidosis Type I Hurler After Successful Hematopoietic Stem Cell Transplantation. J. Bone Jt. Surg. Am. Vol. 2016, 98, 386–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemi Taheri, A.P.; Radmard, A.R.; Kooraki, S.; Behfar, M.; Pak, N.; Hamidieh, A.A.; Ghavamzadeh, A. Radiologic resolution of malignant infantile osteopetrosis skeletal changes following hematopoietic stem cell transplantation. Pediatr. Blood Cancer 2015, 62, 1645–1649. [Google Scholar] [CrossRef]
- Cutler, C.; Antin, J.H. Peripheral blood stem cells for allogeneic transplantation: A review. Stem Cells 2001, 19, 108–117. [Google Scholar] [CrossRef]
- Storek, J.; Joseph, A.; Espino, G.; Dawson, M.A.; Douek, D.C.; Sullivan, K.M.; Flowers, M.E.; Martin, P.; Mathioudakis, G.; Nash, R.A.; et al. Immunity of patients surviving 20 to 30 years after allogeneic or syngeneic bone marrow transplantation. Blood 2001, 98, 3505–3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saba, N.; Flaig, T. Bone Marrow Transplantation for Nonmalignant Diseases. J. Hematother. Stem Cell Res. 2002, 11, 377–387. [Google Scholar] [CrossRef]
- Khaddour, K.; Hana, C.K.; Mewawalla, P. Hematopoietic Stem Cell Transplantation (Bone Marrow Transplant); StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Levine, D.; Navarro, O.M.; Chaudry, G.; Doyle, J.J.; Susan, I. Blaser Imaging the complications of bone marrow transplantation in children. Radiographics 2007, 27, 307–324. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J. Worldwide bone marrow transplantation activity in the last decade. In New Strategies in Bone Marrow Transplantation; Wiley-Liss: New York, NY, USA, 1991; pp. 1–6. [Google Scholar]
- Buckley, R.H. A historical review of bone marrow transplantation for immunodeficiencies. J. Allergy Clin. Immunol. 2004, 113, 793–800. [Google Scholar] [CrossRef]
- Myers, L.A.; Patel, D.D.; Puck, J.M.; Buckley, R.H. Hematopoietic stem cell transplantation for severe combined immunodeficiency in the neonatal period leads to superior thymic output and improved survival. Blood 2002, 99, 872–878. [Google Scholar] [CrossRef] [Green Version]
- Woolfrey, A.E.; Anasetti, C.; Storer, B.; Doney, K.; Milner, L.A.; Sievers, A.L.; Carpenter, P.; Martin, P.; Petersdorf, E.; Appelbaum, F.R.; et al. Factors associated with outcome after unrelated marrow transplantation for treatment of acute lymphoblastic leukemia in children. Blood 2002, 99, 2002–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtick, U.; Albrecht, M.; Chemnitz, J.M.; Theurich, S.; Skoetz, N.; Scheid, C.; von Bergwelt-Baildon, M. Bone marrow versus peripheral blood allogeneic haematopoietic stem cell transplantation for haematological malignancies in adults. Cochrane Database Syst. Rev. 2014, 20, CD010189. [Google Scholar] [CrossRef] [PubMed]
- Stem Cell Trialists’ Collaborative Group: Allogeneic Peripheral Blood Stem-Cell Compared with Bone Marrow Transplantation in the Management of Hematologic Malignancies: An Individual Patient Data Meta-Analysis of Nine Randomized Trials. J. Clin. Oncol. 2005, 23, 5074–5087. [CrossRef] [PubMed]
- Cutler, C.; Antin, J.H. Stem cell sources: Peripheral blood stem cells and bone marrow for allogeneic transplantation. In Stem Cell Transplantation Hematologic Disorders; Soiffer, R.J., Ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 337–356. [Google Scholar]
- Mohty, M.; Kuentz, M.; Michallet, M.; Bourhis, J.H.; Milpied, N.; Sutton, L.; Jouet, J.P.; Attal, M.; Bordigoni, P.; Cahn, J.Y.; et al. Chronic graft-versus-host disease after allogeneic blood stem cell transplantation: Long-term results of a randomized study. Blood 2002, 100, 3128–3134. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.N.D.; Lima, L.C. The association between physical exercise and reactive oxygen species (ROS) production. J. Sports Med. Doping Stud. 2015, 5, 152. [Google Scholar]
- Willems, P.H.; Rossignol, R.; Dieteren, C.E.; Murphy, M.P.; Koopman, W.J. Redox homeostasis and mitochondrial dynamics. Cell Metab. 2015, 22, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, C.N.; Ito, K. DNA damage response, redox status and hematopoiesis. Blood Cells Mol. Dis. 2014, 52, 12–18. [Google Scholar] [CrossRef] [Green Version]
- Yahata, T.; Takanashi, T.; Muguruma, Y.; Ibrahim, A.A.; Matsuzawa, H.; Uno, T.; Sheng, Y.; Onizuka, M.; Ito, M.; Kato, S.; et al. Accumulation of oxidative DNA damage restricts the self-renewal capacity of human hematopoietic stem cells. Blood 2011, 118, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Santoro, M.M. ROS homeostasis and metabolism: A dangerous liaison in cancer cells. Cell Death Dis. 2017, 7, e2253. [Google Scholar] [CrossRef]
- Morry, J.; Ngamcherdtrakul, W.; Yantasee, W. Oxidative stress in cancer and fibrosis: Opportunity for therapeutic intervention with antioxidant compounds, enzymes, and nanoparticles. Redox Biol. 2017, 11, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Chen, J.W.; Dodia, C.; Feinstein, S.I.; Jain, M.K.; Fisher, A.B. 1-Cys peroxiredoxin, a bifunctional enzyme with gluthatione peroxidase and phospholipase A2 activities. J. Biol. Chem. 2000, 275, 28421–28427. [Google Scholar] [CrossRef] [Green Version]
- Day, A.M.; Brown, J.D.; Taylor, S.R.; Rand, J.D.; Morgan, B.A.; Veal, E.A. Inactivation of a peroxiredoxin by hydrogen peroxide is critical for thioredoxin-mediated repair of oxidized proteins and cell survival. Mol. Cell 2012, 45, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Calvani, M.; Comito, G.; Giannoni, E.; Chiarugi, P. Timedependent stabilization of hypoxia inducible factor-1α by different intracellular sources of reactive oxygen species. PLoS ONE 2012, 7, e38388. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Mendoza, N.; Morales-González, A.; Madrigal-Santillán, E.O.; Madrigal-Bujaidar, E.; Álvarez-González, I.; Garcia-Melo, R.F.; Anguiano-Robledo, L.; Fregoso-Aguilar, T.; Morales-Gonzalez, J.A. Antioxidant and adaptative response mediated by Nrf2 during physical exercise. Antioxidants 2019, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Laurenti, E.; Gottgens, B. From haematopoietic stem cells to complex differentiation landscapes. Nature 2018, 553, 418–426. [Google Scholar] [CrossRef]
- Wilson, E.; Laurenti, E.; Trumpp, A. Balancing dormant and self-renewing hematopoietic stem cells. Curr. Opin. Gen. Dev. 2009, 19, 461–468. [Google Scholar] [CrossRef]
- Itkin, T.; Gur-Cohen, S.; Spencer, J.A.; Schajnovitz, A.; Ramasamy, S.K.; Kusumbe, A.P.; Ledergor, G.; Jung, Y.; Milo, I.; Poulos, M.G.; et al. Distinct bone marrow blood vessels differentially regulate haematopoiesis. Nature 2016, 532, 323–328. [Google Scholar] [CrossRef]
- Suda, T.; Takubo, K.; Semenza, G.L. Metabolic regulation of hematopoietic stem cells in the hypoxic niche. Cell Stem Cell 2011, 9, 298–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takubo, K.; Nagamatsu, G.; Kobayashi, C.I.; Nakamura-Ishizu, A.; Kobayashi, H.; Ikeda, E.; Goda, N.; Rahimi, Y.; Johnson, R.S.; Soga, T.; et al. Regulation of glycolysis by Pdk functions as a metabolic checkpoint for cell cycle quiescence in hematopoietic stem cells. Cell Stem Cell 2013, 12, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samimi, A.; Kalantari, H.; Lorestani, M.Z.; Shirzad, R.; Saki, N. Oxidative stress in normal hematopoietic stem cells and leukemia. Apmis 2018, 126, 284–294. [Google Scholar] [CrossRef]
- Bai, L.; Best, G.; Xia, W.; Peters, L.; Wong, K.; Ward, C.; Greenwood, M. Expression of Intracellular Reactive Oxygen Species in Hematopoietic Stem Cells Correlates with Time to Neutrophil and Platelet Engraftment in Patients Undergoing Autologous Bone Marrow Transplantation. Biol. Blood Marrow Transpl. 2018, 24, 1997–2002. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Song, Y.; Hu, Y.; Shi, M.-M.; Wang, Y.-T.; Wang, Y.; Zhang, X.-H.; Xu, L.-P.; Liu, K.-Y.; Deng, H.-K.; et al. Increased reactive oxygen species and exhaustion of quiescent CD34-positive bone marrow cells may contribute to poor graft function after allotransplants. Oncotarget 2016, 7, 30892–30906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, A.; Imai, Y.; Nakagawa, M.; Takahashi, T.; Ichikawa, M.; Kurokawa, M. Intracellular reactive oxygen species mark and influence the megakaryocyte-erythrocyte progenitor fate of common myeloid progenitors. Stem Cells 2014, 32, 548–557. [Google Scholar] [CrossRef]
- Lewandowski, D.; Barroca, V.; Ducongé, F.; Bayer, J.; Van Nhieu, J.T.; Pestourie, C.; Fouchet, P.; Tavitian, B.; Roméo, P.H. In vivo cellular imaging pinpoints the role of reactive oxygen species in the early steps of adult hematopoietic reconstitution. Blood 2010, 115, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.M.; Esain, V.; Frechette, G.M. Glucose metabolism impacts the spatio-temporal onset and magnitude of HSC induction in vivo. Blood 2013, 121, 2483–2493. [Google Scholar] [CrossRef] [Green Version]
- Linping, H.; Zhang, Y.; Miao, W.; Cheng, T. Reactive Oxygen Species and Nrf2: Functional and Transcriptional Regulators of Hematopoiesis. Oxid. Med. Cell. Longev. 2019, 2019, 5153268. [Google Scholar]
- Hole, P.S.; Zabkiewicz, J.; Munje, C. Overproduction of NOX derived ROS in AML promotes proliferation and disassociated with defective oxidative stress signaling. Blood 2013, 122, 3322–3330. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Han, Y.C.; Zou, Y.R. CXCR4 is required for the quiescence of primitive hematopoietic cells. J. Exp. Med. 2008, 205, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Yu, H.; Liang, P.H.; Cheng, H.; XuFeng, H.; Youan, Y.; Zhang, P.; Smith, C.A.; Cheng, T. An acute negative bystander effect of γ-irradiated recipients on transplanted hematopoietic stem cells. Blood 2012, 119, 3629–3637. [Google Scholar] [CrossRef]
- Mantel, C.; Messina-Graham, S.; Moh, A.; Cooper, S.; Hangoc, G.; Fu, X.Y.; Broxmeyer, H.E. Mouse hematopoietic cell-targeted STAT3 deletion: Stem/progenitor cell defects, mitochondrial dysfunction, ROS overproduction, and a rapid aging-like phenotype. Blood 2012, 120, 2589–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Erp, A.C.; Hoeksma, D.; Rebolledo, R.A.; Ottens, P.J.; Jochmans, I.; Monbaliu, D.; Pirenne, J.; Henri, G.; Leuvenink, D.; Decuypere, J.P. The Crosstalk between ROS and Autophagy in the Field of Transplantation Medicine. Oxid. Med. Cell. Longev. 2017, 2017, 7120962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, S.; Massi, D.; Farini, V.; Baroni, G.; Parri, M.; Innocenti, S.; Cecchi, R.; Chiarugi, P. Beta-adrenoceptors are upregulated in human melanoma and their activation releases pro-tumorigenic cytokines and metalloproteases in melanoma cell lines. Lab. Investig. 2013, 93, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jockers, R.; Da Silva, A.; Strosberg, A.D.; Bouvier, M.; Marullo, S. New molecular and structural determinants involved in β2-adrenergic receptor desensitization and sequestration. Delineation using chimeric β3/β2-adrenergic receptors. J. Biol. Chem. 1996, 271, 9355–9362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emorine, L.J.; Marullo, S.; Briend-Sutren, M.M.; Patey, G.; Tate, K.; Delavier-Klutchko, C.; Strosberg, A.D. Molecular characterization of the human beta 3-adrenergic receptor. Science 1989, 245, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Nahmias, C.; Blin, N.; Elalouf, J.M.; Mattei, M.G.; Strosberg, A.D.; Emorine, L.J. Molecular characterization of the mouse β3-adrenergic receptor: Relationship with the atypical receptor of adipocytes. EMBO J. 1991, 10, 3721–3727. [Google Scholar] [CrossRef]
- Bartness, T.J.; Vaughan, C.H.; Song, C.K. Sympathetic and sensory innervation of brown adipose tissue. Int. J. Obes. 2010, 34 (Suppl. 1), S36–S42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessy, C.; Balligand, J.L. Beta3-adrenergic receptors in cardiac and vascular tissues emerging concepts and therapeutic perspectives. Adv. Pharmacol. 2010, 59, 135–163. [Google Scholar]
- Igawa, Y.; Michel, M.C. Pharmacological profile of β3- adrenoceptor agonists in clinical development for the treatment of overactive bladder syndrome. Naunyn-Schmiedeberg Arch. Pharmacol. Vol. 2013, 386, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Perrone, M.G.; Notarnicola, M.; Caruso, M.G.; Tutino, V.; Scilimati, A. Upregulation of beta3-adrenergic receptor mrna in human colon cancer: A preliminary study. Oncology 2008, 75, 224–229. [Google Scholar] [CrossRef]
- Dal Monte, M.; Casini, G.; Filippi, L.; Nicchia, G.P.; Svelto, M.; Bagnoli, P. Functional involvement of β3-adrenergic receptors in melanoma growth and vascularization. J. Mol. Med. 2013, 91, 1407–1419. [Google Scholar] [CrossRef]
- Calvani, M.; Pelon, F.; Comito, G.; Taddei, M.L.; Moretti, S.; Innocenti, S.; Nassini, R.; Gerlini, G.; Borgognoni, L.; Bambi, F.; et al. Norepinephrine promotes tumor microenvironment reactivity through β3-adrenoreceptors during melanoma progression. Oncotarget 2015, 6, 4615–4632. [Google Scholar] [CrossRef] [Green Version]
- Montoya, A.; Amaya, C.N.; Belmont, A.; Diab, N.; Trevino, R.; Villanueva, G.; Rains, S.; Sanchez, L.A.; Badri, N.; Otoukesh, S.; et al. Use of non-selective beta-blockers is associated with decreased tumor proliferative indices in early-stage breast cancer. Oncotarget 2017, 8, 6446–6460. [Google Scholar] [CrossRef] [Green Version]
- Michel, M.C.; Gravas, S. Safety and tolerability of β3-adrenoceptor agonists in the treatment of overactive bladder syndrome—Insight from transcriptosome and experimental studies. Expert Opin. Drug Saf. 2016, 15, 647–657. [Google Scholar] [CrossRef]
- Cernecka, H.; Sand, C.; Michel, M.C. The odd sibling: Features of β3-adrenoceptor pharmacology. Mol. Pharmacol. 2014, 86, 479–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvani, M.; Bruno, G.; Dal Monte, M.; Nassini, R.; Fontani, F.; Casini, A.; Cavallini, L.; Becatti, M.; Bianchini, F.; DeLogu, F.; et al. β3-Adrenoceptor as a potential immuno-suppressor agent in melanoma. Br. J. Pharmacol. 2019, 176, 2509–2524. [Google Scholar] [CrossRef]
- Cole, S.W.; Sood, A.K. Molecular pathways: Beta-adrenergic signaling in cancer. Clin. Cancer Res. 2012, 18, 1201–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjosted, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, V.; Grazzini, M.; Benemei, S.; Marchionni, N.; Botteri, E.; Pennacchioli, E.; Geppetti, P.; Gandini, S. Propranolol for off-label treatment of patients with melanoma: Results from a cohort study. JAMA Oncol. 2018, 4, e172908. [Google Scholar] [CrossRef]
- Hwa, Y.L.; Shi, Q.; Kumar, S.K.; Lacy, M.A.Q.; Gertz, M.A.; Kapoor, P.; Buadi, F.K.; Leung, N.; Dingli, D.; Go, R.S.; et al. Beta-blockers improve survival outcomes in patients with multiple myeloma: A retrospective evaluation. Am. J. Hematol. 2017, 92, 50–55. [Google Scholar] [CrossRef]
- Jin, J.; Miao, C.; Wang, Z.; Zhang, W.; Zhang, X.; Xie, X.; Lu, W. Design and synthesis of aryloxypropanolamine as β3-adrenergic receptor antagonist in cancer and lipolysis. Eur. J. Med. Chem. 2018, 150, 757–770. [Google Scholar] [CrossRef]
- Andersson, K.E.; Martin, N.; Nitti, V. Selective β3-adrenoceptor agonists for the treatment of overactive bladder. J. Urol. 2013, 190, 1173–1180. [Google Scholar] [CrossRef]
- Perrone, G.M.; Scilimati, A. β(3)-Adrenoceptor agonists and (antagonists as) inverse agonists history, perspective, constitutive activity, and stereospecific binding. Methods Enzymol. 2010, 484, 197–230. [Google Scholar]
- Schena, G.; Caplan, M.J. Everything You Always Wanted to Know about β3-AR * (* But Were Afraid to Ask). Cells 2019, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Hadi, T.; Douhard, R.; Dias, A.M.M.; Wendremaire, M.; Pezzè, M. Beta3 adrenergic receptor stimulation in human macrophages inhibits NADPH oxidase activity and induces catalase expression via PPAR γ activation. Biochim. Biophys. Acta 2017, 1854, 1769–1784. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Kadoi, H.; Yamamuro, A.; Ishimaru, Y.; Maeda, S. Noradrenaline increases intracellular glutathione in human astrocytoma U-251 MG cells by inducing glutamate-cysteine ligase protein via β3- adrenoceptor stimulation. Eur. J. Pharmacol. 2015, 772, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Calvani, M.; Cavallini, L.; Tondo, A.; Spinelli, V.; Ricci, L.; Pasha, A.; Bruno, G.; Buonvicino, D.; Bigagli, E.; Vignoli, M.; et al. β3-Adrenoreceptors Control Mitochondrial Dormancy in Melanoma and Embryonic Stem Cells. Oxid. Med. Cell. Longev. 2018, 2018, 6816508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvani, M.; Favre, C. Antioxidant Nutraceutical approach to Ewing Sarcoma: Where is the Trap? Biomed. J. Sci. Tech. Res. 2019, 17, 12805–12814. [Google Scholar]
- Pasha, A.; Vignoli, M.; Subbiani, A.; Nocentini, A.; Selleri, S.; Gratteri, P.; DaBraio, A.; Casini, T.; Filippi, L.; Fotzi, I.; et al. β3-Adrenoreceptor Activity Limits Apigenin Efficacy in Ewing Sarcoma Cells: A Dual Approach to Prevent Cell Survival. Int. J. Mol. Sci. 2019, 20, 2149. [Google Scholar] [CrossRef] [Green Version]
- Bogacka, I.; Gettys, T.W.; da Jonge, L.; Nguyen, T.; Smith, J.M.; Xie, H.; Greenway, F.; Smith, S.R. The Effect of β-Adrenergic and Peroxisome Proliferator–Activated Receptor-γ Stimulation on Target Genes Related to Lipid Metabolism in Human Subcutaneous Adipose Tissue. Diabetes Care 2007, 30, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Chiarelli, F.; Di Marzio, D. Peroxisome proliferator-activated receptor-gamma agonists and diabetes: Current evidence and future perspectives. Vasc. Health Risk Manag. 2008, 4, 297–304. [Google Scholar]
- Shi, S.T.; Li, Y.F.; Guo, Y.Q.; Wang, Z.H. Effect of beta-3 adrenoceptor stimulation on the levels of ApoA-I, PPARα, and PPARγ in apolipoprotein e-deficient mice. J Cardiov. Pharmacol. 2014, 65, 407–411. [Google Scholar] [CrossRef]
- Maestroni, G.J.M. Adrenergic Modulation of Hematopoiesis. J. Neuroimmune Pharmacol. 2019, 15, 82–92. [Google Scholar] [CrossRef]
- Freeman, J.G.; Ryan, J.J.; Shelburne, C.P.; Bailey, D.P.; Bouton, L.A.; Narasimhachari, N.; Domen, J.; Siméon, N.; Couderc, F.; Stewart, J.K. Catecholamines in murine bone marrow derived mast cells. J. Neuroimmunol. 2001, 119, 231–238. [Google Scholar] [CrossRef]
- Maestroni, G.J.; Conti, A.; Pedrinis, E. Effect of adrenergic agents on hematopoiesis after syngeneic bone marrow transplantation in mice. Blood 1992, 80, 1178–1182. [Google Scholar] [CrossRef] [Green Version]
- Marino, F.; Cosentino, M.; Bombelli, R.; Ferrari, M.; Maestroni, G.J.; Conti, A.; Lecchini, S. Measurement of catecholamines in mouse bone marrow by means of HPLC with electrochemical detection. Haematological 1997, 82, 392–394. [Google Scholar]
- Hasan, S.; Johnson, N.B.; Mosier, M.J.; Shankar, R.; Conrad, P.; Szilagyi, A.; Gamelli, R.L.; Muthumalaiappan, K. Myelo-erythroid commitment after burn injury is under β-adrenergic control via MafB regulation. Am. J. Physiol. Cell Physiol. 2017, 312, C286–C301. [Google Scholar] [CrossRef] [PubMed]
- Massberg, S.; Schaerli, P.; Knezevic-Maramica, I.; Köllnberger, M.; Tubo, N.; Moseman, E.A.; Huff, I.V.; Junt, T.; Wagers, A.J.; Mazo, I.B.; et al. Physiological recirculation of hematopoietic stem and progenitor cells through blood, lymph and extramedullary tissues. Cell 2007, 131, 994–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asri, A.; Sabour, J.; Akashi, A.; Soleimani, M. Homing in hematopoietic stem cells: Focus on regulatory role of CXCR7 on SDF1a/CXCR4 axis. EXCLI J. 2016, 15, 134–143. [Google Scholar] [PubMed]
- Katayama, Y.; Battista, M.; Kao, W.M.; Hidalgo, A.; Peired, A.J.; Thomas, S.A.; Frenette, P.S. Signals from the sympathetic nervous system regulate hematopoietic stem cell egress from bone marrow. Cell 2006, 124, 407–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendez-Ferrer, S.; Lucas, D.; Battista, M.; Frenette, P.S. Haematopoietic stem cell release is regulated by circadian oscillations. Nature 2008, 452, 442–447. [Google Scholar] [CrossRef]
- Jang, Y.Y.; Sharkis, S.J. A low level of reactive oxygen species selects for primitive hematopoietic stem cells that may reside in the low-oxygenic niche. Blood 2007, 110, 3056–3063. [Google Scholar] [CrossRef] [Green Version]
- Marino, F.; Cosentino, M. Adrenergic modulation of immune cells: An update. Amino Acids 2013, 45, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Facciabene, A.; Peng, X.; Hagemann, I.S.; Balint, K.; Barchetti, A.; Wang, L.; Gimotty, P.G.; Gilks, C.B.; Lal, P.; Zhang, L.; et al. Tumor hypoxia promotes tolerance and angiogenesis via CCL28 and T (reg) cells. Nature 2011, 475, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Laukova, M.; Vargovic, P.; Csaderova, L.; Chovanova, L.; Vlcek, M.; Imrich, R.; Krizanova, O.; Kvetnansky, R. Acute stress differently modulates β1, β2 and β3 adrenoceptors in T cells, but not in B cells, from the rat spleen. Neuroimmunmod 2012, 19, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Calvani, M.; Bruno, G.; Dabraio, A.; Subbiani, A.; Bianchini, F.; Fontani, F.; Casazza, G.; Vignoli, M.; De Logu, F.; Frenos, S.; et al. β3-Adrenoreceptor Blockade Induces Stem Cells Differentiation in Melanoma Microenvironment. Int. J. Mol. Sci. 2020, 21, 1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasha, A.; Calvani, M.; Favre, C. β3-Adrenoreceptors as ROS Balancer in Hematopoietic Stem Cell Transplantation. Int. J. Mol. Sci. 2021, 22, 2835. https://doi.org/10.3390/ijms22062835
Pasha A, Calvani M, Favre C. β3-Adrenoreceptors as ROS Balancer in Hematopoietic Stem Cell Transplantation. International Journal of Molecular Sciences. 2021; 22(6):2835. https://doi.org/10.3390/ijms22062835
Chicago/Turabian StylePasha, Amada, Maura Calvani, and Claudio Favre. 2021. "β3-Adrenoreceptors as ROS Balancer in Hematopoietic Stem Cell Transplantation" International Journal of Molecular Sciences 22, no. 6: 2835. https://doi.org/10.3390/ijms22062835
APA StylePasha, A., Calvani, M., & Favre, C. (2021). β3-Adrenoreceptors as ROS Balancer in Hematopoietic Stem Cell Transplantation. International Journal of Molecular Sciences, 22(6), 2835. https://doi.org/10.3390/ijms22062835