
Ivabradine Induces Cardiac Protection against Myocardial Infarction by Preventing Cyclophilin-A Secretion in Pigs under Coronary Ischemia/Reperfusion

 ,
,  , ,
, ,  ,
,  and
and 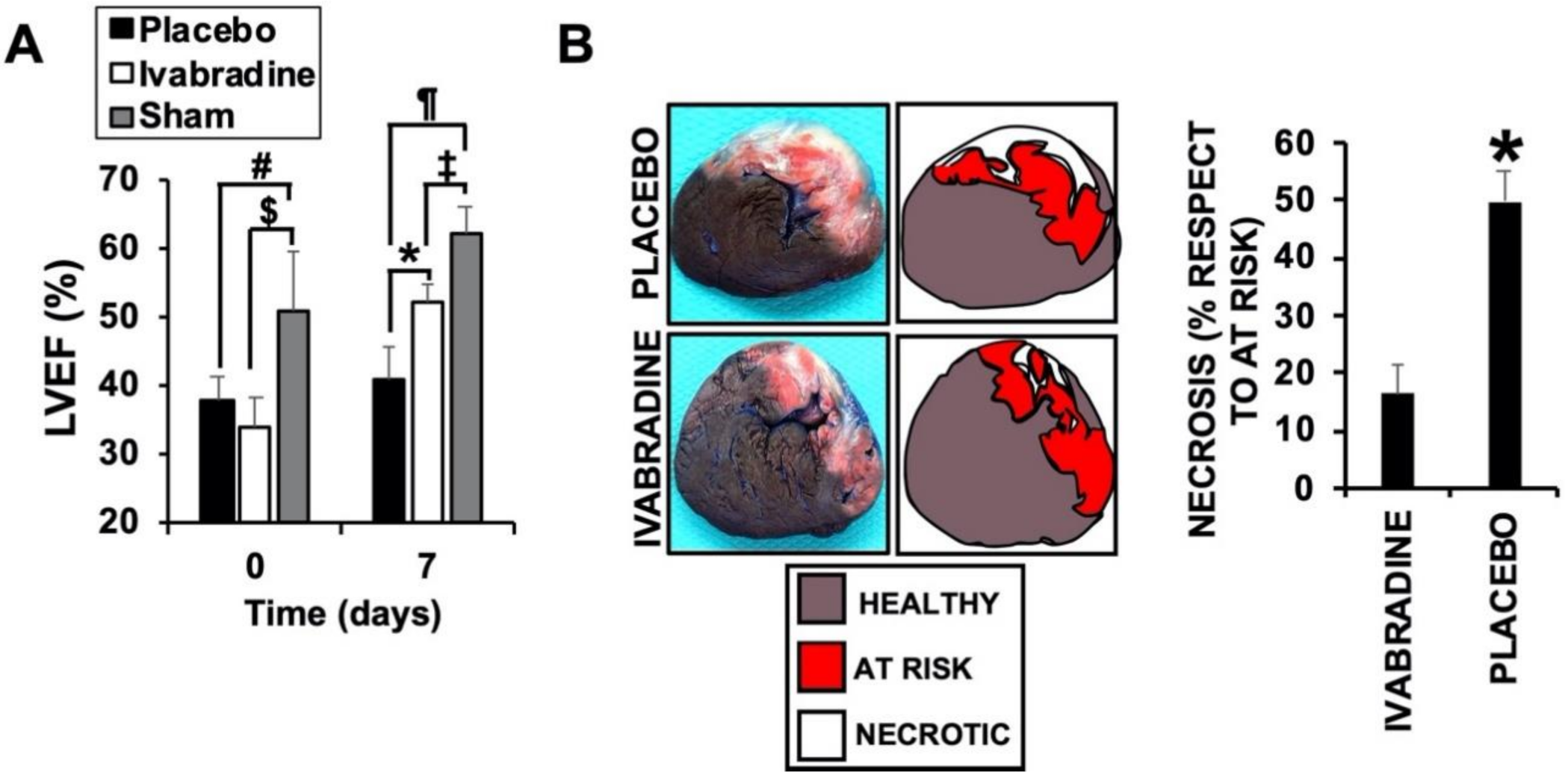{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ivabradine Induces Cardiac Protection of Pigs against IR
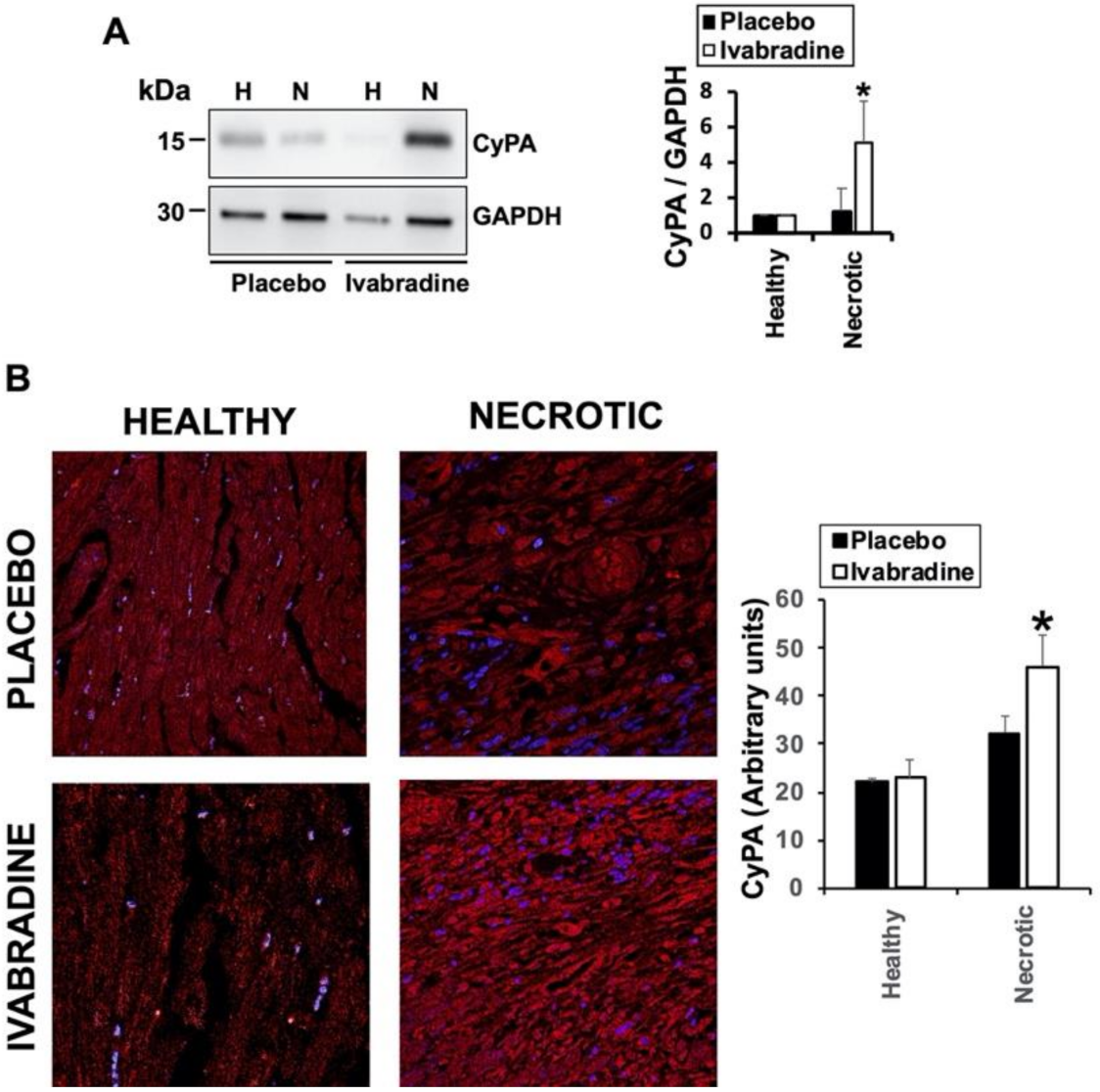
2.2. Ivabradine Increases the Expression of Cyclophilin-A in the Hearts of Pigs after IR
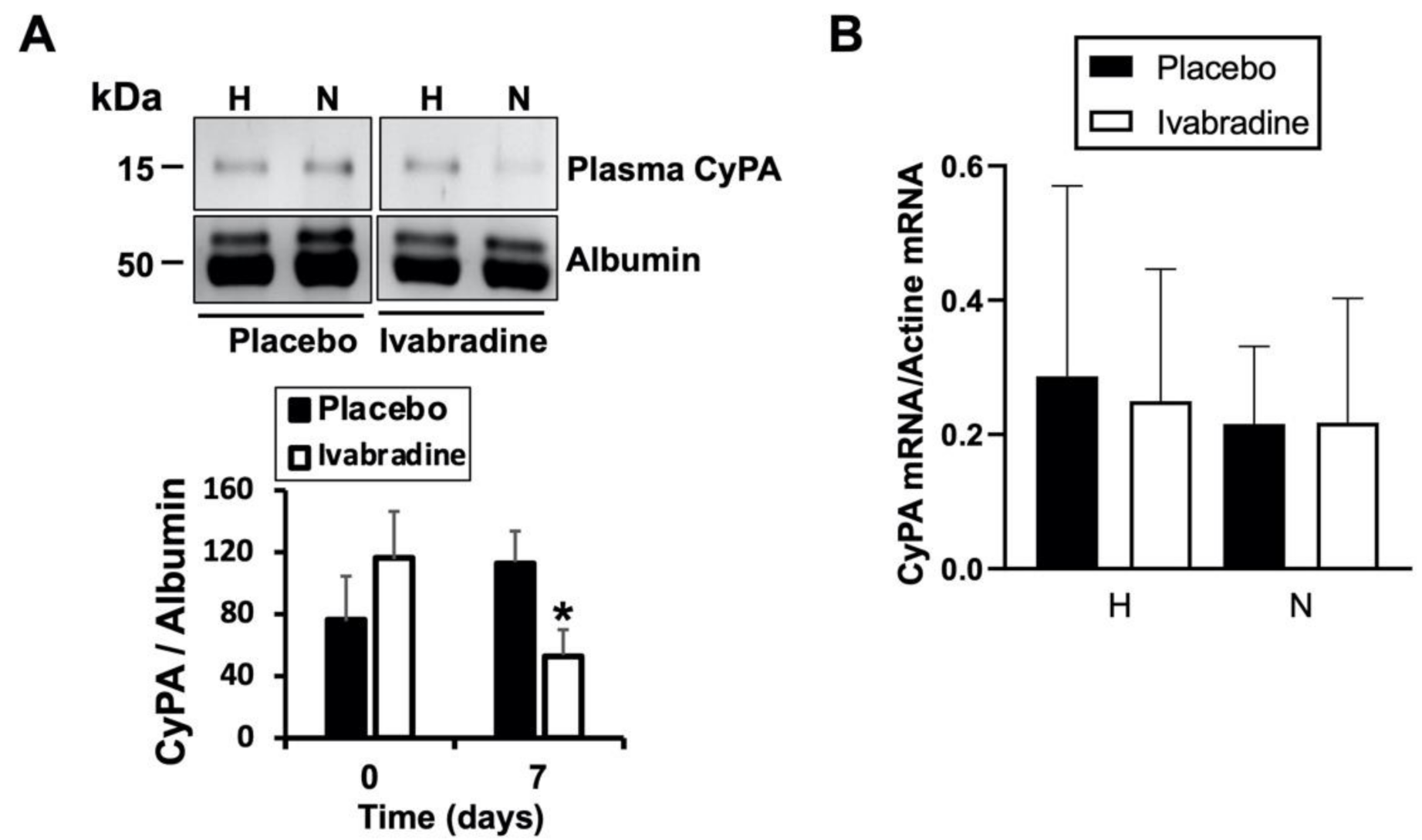
2.3. Ivabradine Reduces Plasma CyPA Levels after IR
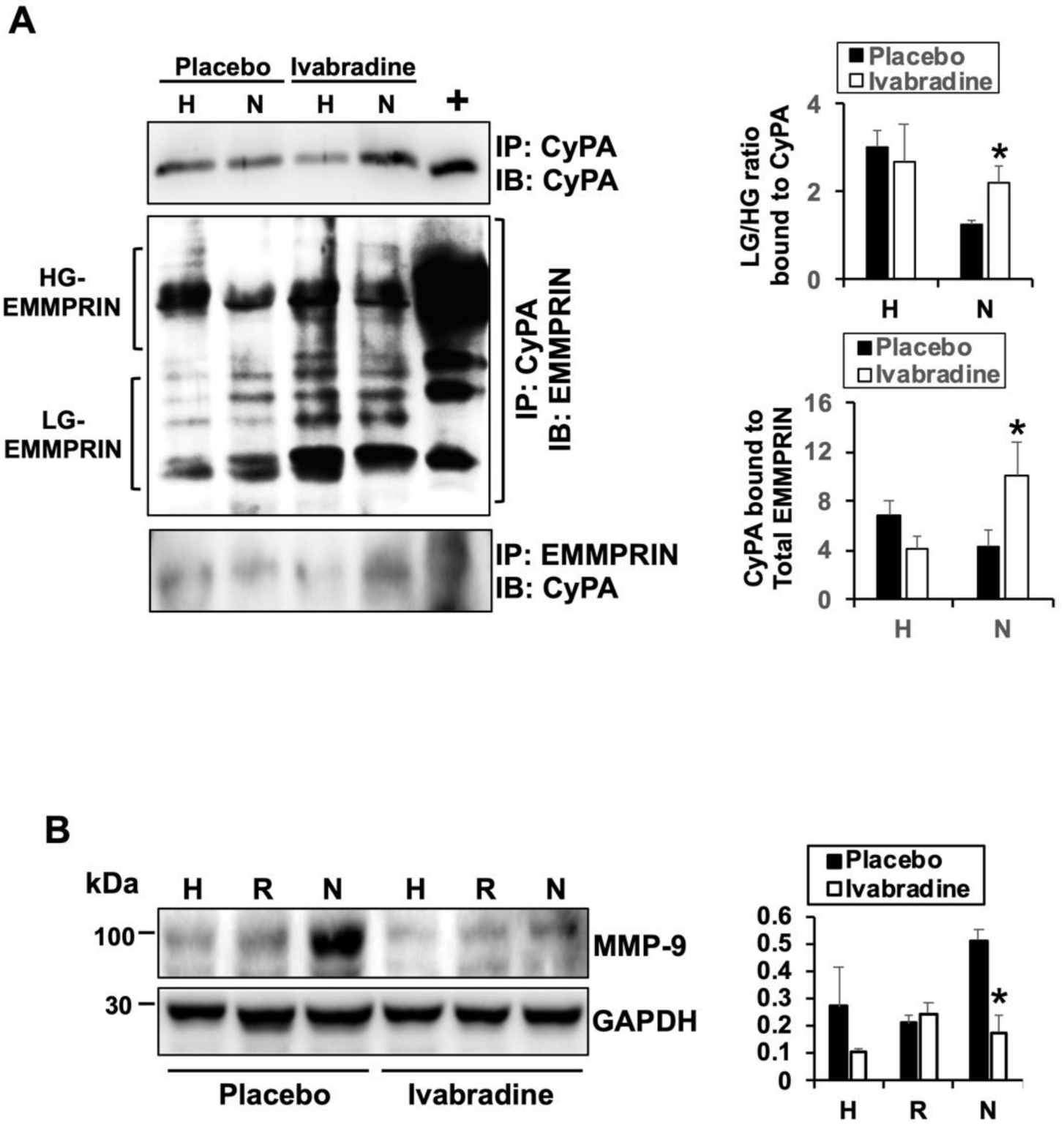
2.4. Ivabradine Increases the Binding between CyPA and Low-Glycosylated forms of EMMPRIN
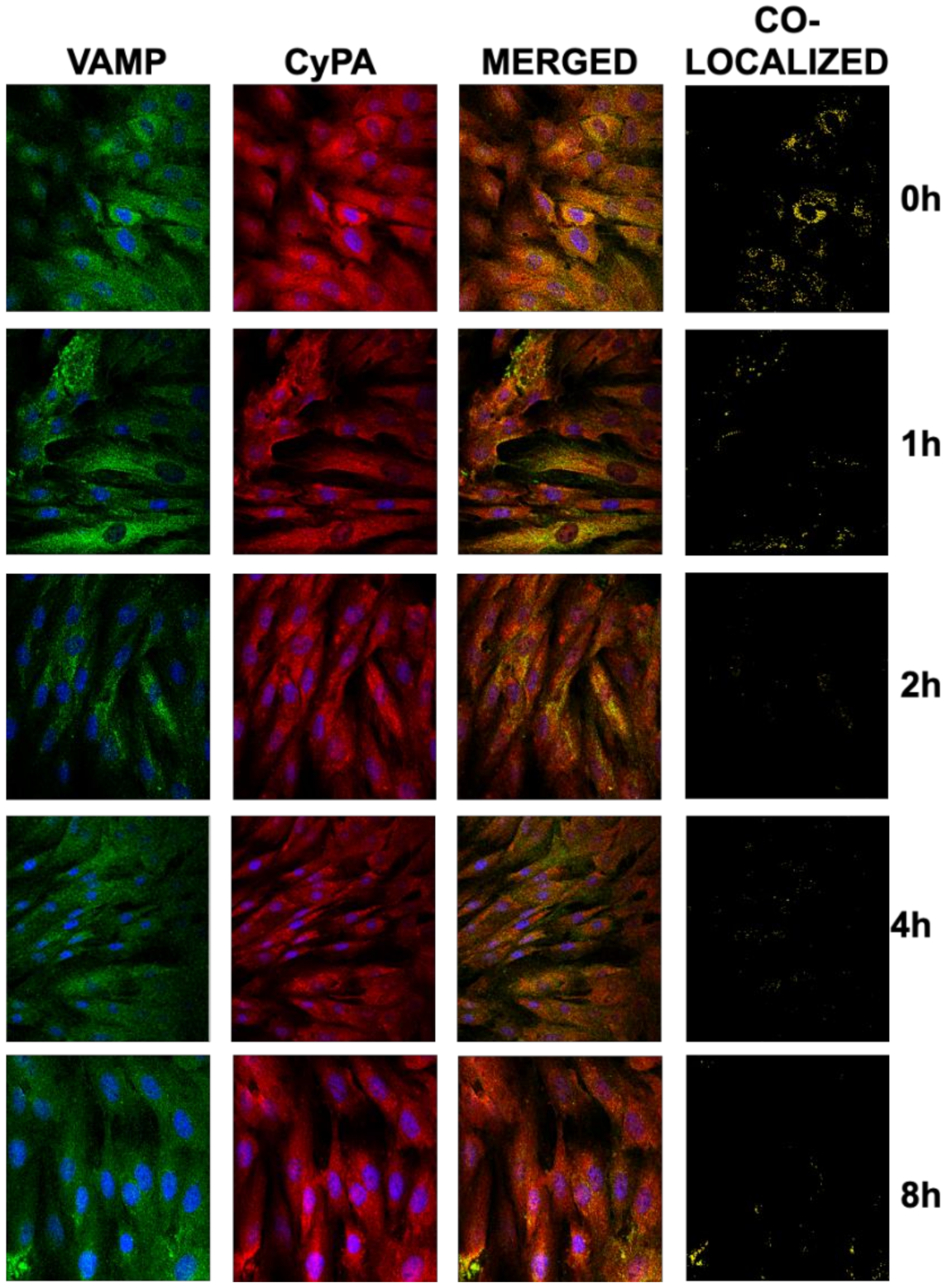
2.5. Ivabradine Reduces the Secretion of CyPA in Cardiac Cells
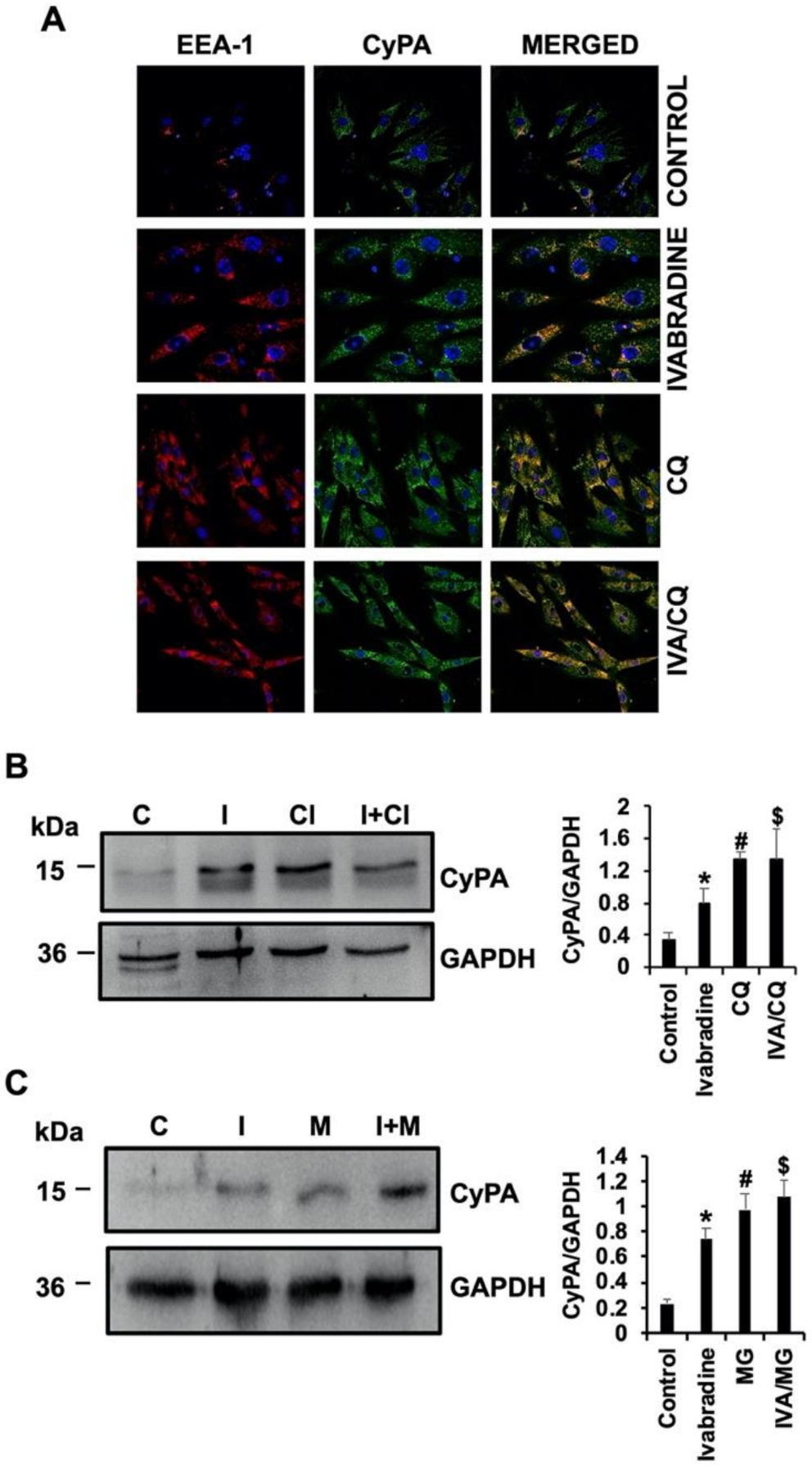
2.6. Ivabradine Decreases Lysosomal Degradation of CyPA in Cardiac Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cardiac Ischemia/Reperfusion
4.3. Echocardiography
4.4. Evans Blue/TTC Staining
4.5. RNA Isolation and RT-PCR
4.6. Confocal Microscopy
4.7. H9c2 Cell Culture and Treatment
4.8. Protein Expression Determination by Immunoblot
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ide, T.; Ohtani, K.; Higo, T.; Tanaka, M.; Kawasaki, Y.; Tsutsui, H. Ivabradine for the Treatment of Cardiovascular Diseases. Circ. J. 2019, 83, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Izco, M.; Castejón, B.; Piedras, M.J.; Zamorano, J.L.; San Martín, M.; Zaragoza, C. Efectos de la ivabradina en la frecuencia cardiaca y los parámetros hemodinámicos en un modelo porcino de shock cardiogénico. Rev. Esp. Cardiol. 2017, 70, 1139–1141. [Google Scholar] [CrossRef] [PubMed]
- Izco, M.P.; Ramírez-Carracedo, R.; Navarro, I.H.; Ruiz, Á.O.; Navarro, B.C.; Berrocal, I.C.; Aramburu, C.L.; Salinas, G.L.A.; Díez, J.; Redondo, M.S.; et al. Ivabradine in acute heart failure: Effects on heart rate and hemodynamic parameters in a randomized and controlled swine trial. Cardiol. J. 2020, 27, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Seizer, P.; Ochmann, C.; Schönberger, T.; Zach, S.; Rose, M.; Borst, O.; Klingel, K.; Kandolf, R.; Macdonald, H.R.; Nowak, R.A.; et al. Disrupting the EMMPRIN (CD147)-Cyclophilin A Interaction Reduces Infarct Size and Preserves Systolic Function After Myocardial Ischemia and Reperfusion. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1377–1386. [Google Scholar] [CrossRef] [Green Version]
- Tarin, C.; Lavin, B.; Gomez, M.; Saura, M.; Diez-Juan, A.; Zaragoza, C. The extracellular matrix metalloproteinase inducer EMMPRIN is a target of nitric oxide in myocardial ischemia/reperfusion. Free Radic. Biol. Med. 2011, 51, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, I.; Piedras, M.J.G.M.; Herruzo, I.; Turpin, M.D.C.; Castejón, B.; Reventun, P.; Martin, A.; Saura, M.; Zamorano, J.L.; Zaragoza, C. EMMPRIN-Targeted Magnetic Nanoparticles for In Vivo Visualization and Regression of Acute Myocardial Infarction. Theranostics 2016, 6, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Carracedo, R.; Tesoro, L.; Hernandez, I.; Diez-Mata, J.; Filice, M.; Toro, R.; Rodriguez-Piñero, M.; Zamorano, J.L.; Saura, M.; Zaragoza, C. Non-Invasive Detection of Extracellular Matrix Metalloproteinase Inducer EMMPRIN, a New Therapeutic Target against Atherosclerosis, Inhibited by Endothelial Nitric Oxide. Int. J. Mol. Sci. 2018, 19, 3248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Carracedo, R.; Tesoro, L.; Hernandez, I.; Diez-Mata, J.; Botana, L.; Saura, M.; Sanmartin, M.; Zamorano, J.L.; Zaragoza, C. Ivabradine-Stimulated Microvesicle Release Induces Cardiac Protection against Acute Myocardial Infarction. Int. J. Mol. Sci. 2020, 21, 6566. [Google Scholar] [CrossRef]
- Schmidt, R.; Bultmann, A.; Ungerer, M.; Joghetaei, N.; Bulbul, O.; Thieme, S.; Chavakis, T.; Toole, B.P.; Gawaz, M.; Schömig, A.; et al. Extracellular matrix metalloproteinase inducer regulates matrix metalloproteinase activity in cardiovascular cells: Implications in acute myocardial infarction. Circulation 2006, 113, 834–841. [Google Scholar] [CrossRef] [Green Version]
- Ferlito, M.; Fulton, W.B.; Zauher, M.A.; Marbán, E.; Steenbergen, C.; Lowenstein, C.J.; Zauher, A.M. VAMP-1, VAMP-2, and syntaxin-4 regulate ANP release from cardiac myocytes. J. Mol. Cell. Cardiol. 2010, 49, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Rath, D.; Ungern-Sternberg, S.; Heinzmann, D.; Sigle, M.; Monzien, M.; Horstmann, K.; Schaeffeler, E.; Winter, S.; Müller, K.; Groga-Bada, P.; et al. Platelet surface expression of cyclophilin A is associated with increased mortality in patients with symptomatic coronary artery disease. J. Thromb. Haemost. 2019, 18, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Yuan, W.; Li, J.; Zhou, H.; Xu, L.; Weng, J.; Li, X.; Zhang, X.; Wang, Z.; Yan, J. Cyclophilin A mediates the ox-LDL-induced activation and apoptosis of macrophages via au-tophagy. Int. J. Cardiol. 2017, 230, 142–148. [Google Scholar] [CrossRef]
- Su, Z.; Lin, M.; Su, Y.; Li, J.; Lin, R.; Wu, M.; Wang, X.; Huang, L.; Chen, Y.; Shu, X.; et al. Oxidized low-density lipoprotein inhibits the degradation of cyclophilin A via the lysosome in vascular smooth muscle cells. Am. J. Transl. Res. 2020, 12, 3964–3973. [Google Scholar]
- Seizer, P.; Fuchs, C.; Ungern-Sternberg, S.N.V.; Heinzmann, D.; Langer, H.; Gawaz, M.; May, A.E.; Geisler, T. Platelet-bound cyclophilin A in patients with stable coronary artery disease and acute myocardial infarction. Platelets 2015, 27, 1–4. [Google Scholar] [CrossRef]
- Suzuki, J.; Jin, Z.-G.; Meoli, D.F.; Matoba, T.; Berk, B.C. Cyclophilin A Is Secreted by a Vesicular Pathway in Vascular Smooth Muscle Cells. Circ. Res. 2006, 98, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, K.; Fukumoto, Y.; Sugimura, K.; Miura, Y.; Aoki, T.; Nochioka, K.; Tatebe, S.; Miyamichi-Yamamoto, S.; Shimizu, T.; Osaki, S.; et al. Plasma cyclophilin A is a novel biomarker for coronary artery disease. Circ. J. 2013, 77, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.H.; Chang, C.C.; Kuo, C.L.; Huang, C.S.; Lin, C.S.; Liu, C.S. Decrease in plasma cyclophilin A concentration at 1 month after myocardial infarction predicts better left ventricular performance and synchronicity at 6 months: A pilot study in patients with ST elevation myocardial infarction. Int. J. Biol. Sci. 2015, 11, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Dawar, F.U.; Wu, J.; Zhao, L.; Khattak, M.N.K.; Mei, J.; Lin, L. Updates in understanding the role of cyclophilin A in leukocyte chemotaxis. J. Leukoc. Biol. 2017, 101, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Sowden, M.P.; Berk, B.C. Extracellular and Intracellular Cyclophilin A, Native and Post-Translationally Modified, Show Diverse and Specific Pathological Roles in Diseases. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Daneri-Becerra, C.; Valeiras, B.; Gallo, L.I.; Lagadari, M.; Galigniana, M.D. Cyclophilin A is a mitochondrial factor that forms complexes with p23—Correlative evidence for an anti-apoptotic action. J. Cell Sci. 2021, 134, jcs253401. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhang, W.; Wu, Y.; Zhang, M.; Wang, L.; Wang, Y.; Wang, Y.; Liu, W. Cyclophilin A inhibits A549 cell oxidative stress and apoptosis by modulating the PI3K/Akt/mTOR signaling pathway. Biosci. Rep. 2021, 41, BSR20203219. [Google Scholar] [CrossRef] [PubMed]
- Bayon, J.; Alfonso, A.; Gegunde, S.; Alonso, E.; Alvarino, R.; Santas-Alvarez, M.; Testa-Fernandez, A.; Rios-Vazquez, R.; Botana, L.; Gonzalez-Juanatey, C. Cyclophilins in Ischemic Heart Disease: Differences Between Acute and Chronic Coronary Artery Disease Patients. Cardiol. Res. 2020, 11, 319–327. [Google Scholar] [CrossRef]
- Lv, M.; Miao, J.; Zhao, P.; Luo, X.; Han, Q.; Wu, Z.; Zhang, K.; Zhu, P. CD147-mediated chemotaxis of CD4+CD161+ T cells may contribute to local inflammation in rheumatoid arthritis. Clin. Rheumatol. 2017, 37, 59–66. [Google Scholar] [CrossRef]
- Sturhan, H.; Ungern-Sternberg, S.N.; Langer, H.; Gawaz, M.; Geisler, T.; May, A.E.; Seizer, P. Regulation of EMMPRIN (CD147) on monocyte subsets in patients with symptomatic coronary artery disease. Thromb. Res. 2015, 135, 1160–1164. [Google Scholar] [CrossRef]
- Seizer, P.; Klingel, K.; Sauter, M.; Westermann, D.; Ochmann, C.; Schönberger, T.; Schleicher, R.; Stellos, K.; Schmidt, E.-M.; Borst, O.; et al. Cyclophilin A affects inflammation, virus elimination and myocardial fibrosis in coxsackievirus B3-induced myocarditis. J. Mol. Cell. Cardiol. 2012, 53, 6–14. [Google Scholar] [CrossRef]
- Cuadrado, I.; Castejon, B.; Martin, A.M.; Saura, M.; Reventun-Torralba, P.; Zamorano, J.L.; Zaragoza, C. Nitric Oxide Induces Cardiac Protection by Preventing Extracellular Matrix Degradation through the Complex Caveolin-3/EMMPRIN in Cardiac Myocytes. PLoS ONE 2016, 11, e0162912. [Google Scholar] [CrossRef] [PubMed]
- Tesoro, L.; Ramirez-Carracedo, R.; Hernandez, I.; Diez-Mata, J.; Pascual, M.; Saura, M.; Sanmartin, M.; Zamorano, J.L.; Zaragoza, C. Ivabradine induces cardiac protection by preventing cardiogenic shock-induced extracellular matrix degradation. Rev. Esp. Cardiol. 2020, in press. [Google Scholar] [CrossRef]
- Pahk, K.; Joung, C.; Song, H.Y.; Kim, S.; Kim, W.-K. Kim SP-8356, a Novel Inhibitor of CD147-Cyclophilin A Interactions, Reduces Plaque Progression and Stabilizes Vulnerable Plaques in apoE-Deficient Mice. Int. J. Mol. Sci. 2019, 21, 95. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Chang, S.B.; Hemler, M.E. Links between CD147 function, glycosylation, and caveolin-1. Mol. Biol. Cell 2004, 15, 4043–4050. [Google Scholar] [CrossRef]
- Aquila, G.; Morelli, M.B.; Sega, F.V.D.; Fortini, F.; Nigro, P.; Caliceti, C.; Ferracin, M.; Negrini, M.; Pannuti, A.; Bonora, M.; et al. Heart rate reduction with ivabradine in the early phase of atherosclerosis is protective in the endothelium of ApoE-deficient mice. J. Physiol. Pharmacol. 2018, 69, 35–52. [Google Scholar] [PubMed]
- Bøtker, H.E.; Hausenloy, D.; Andreadou, I.; Antonucci, S.; Boengler, K.; Davidson, S.M.; Deshwal, S.; Devaux, Y.; Di Lisa, F.; Di Sante, M.; et al. Practical guidelines for rigor and reproducibility in preclinical and clinical studies on cardioprotection. Basic Res. Cardiol. 2018, 113, 1–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosello, X.; Rodriguez-Sinovas, A.; Vilahur, G.; Crisóstomo, V.; Jorge, I.; Zaragoza, C.; Zamorano, J.L.; Bermejo, J.; Ordoñez, A.; Boscá, L.; et al. CIBER-CLAP (CIBERCV Cardioprotection Large Animal Platform): A multicenter preclinical network for testing reproducibility in cardiovascular interventions. Sci. Rep. 2019, 9, 20290. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, I.; Tesoro, L.; Ramirez-Carracedo, R.; Diez-Mata, J.; Sanchez, S.; Saura, M.; Zamorano, J.L.; Zaragoza, C.; Botana, L. Ivabradine Induces Cardiac Protection against Myocardial Infarction by Preventing Cyclophilin-A Secretion in Pigs under Coronary Ischemia/Reperfusion. Int. J. Mol. Sci. 2021, 22, 2902. https://doi.org/10.3390/ijms22062902
Hernandez I, Tesoro L, Ramirez-Carracedo R, Diez-Mata J, Sanchez S, Saura M, Zamorano JL, Zaragoza C, Botana L. Ivabradine Induces Cardiac Protection against Myocardial Infarction by Preventing Cyclophilin-A Secretion in Pigs under Coronary Ischemia/Reperfusion. International Journal of Molecular Sciences. 2021; 22(6):2902. https://doi.org/10.3390/ijms22062902
Chicago/Turabian StyleHernandez, Ignacio, Laura Tesoro, Rafael Ramirez-Carracedo, Javier Diez-Mata, Sandra Sanchez, Marta Saura, Jose Luis Zamorano, Carlos Zaragoza, and Laura Botana. 2021. "Ivabradine Induces Cardiac Protection against Myocardial Infarction by Preventing Cyclophilin-A Secretion in Pigs under Coronary Ischemia/Reperfusion" International Journal of Molecular Sciences 22, no. 6: 2902. https://doi.org/10.3390/ijms22062902
APA StyleHernandez, I., Tesoro, L., Ramirez-Carracedo, R., Diez-Mata, J., Sanchez, S., Saura, M., Zamorano, J. L., Zaragoza, C., & Botana, L. (2021). Ivabradine Induces Cardiac Protection against Myocardial Infarction by Preventing Cyclophilin-A Secretion in Pigs under Coronary Ischemia/Reperfusion. International Journal of Molecular Sciences, 22(6), 2902. https://doi.org/10.3390/ijms22062902