
Essential Role of Non-Coding RNAs in Enterovirus Infection: From Basic Mechanisms to Clinical Prospects




Abstract
:1. Introduction
2. The Epidemiological Characteristics of EVs
3. Discovery and Concept of Non-Coding RNAs (ncRNAs)
4. Description and Biogenesis of ncRNAs
5. Acute EV Infection and ncRNA Expression Profiles
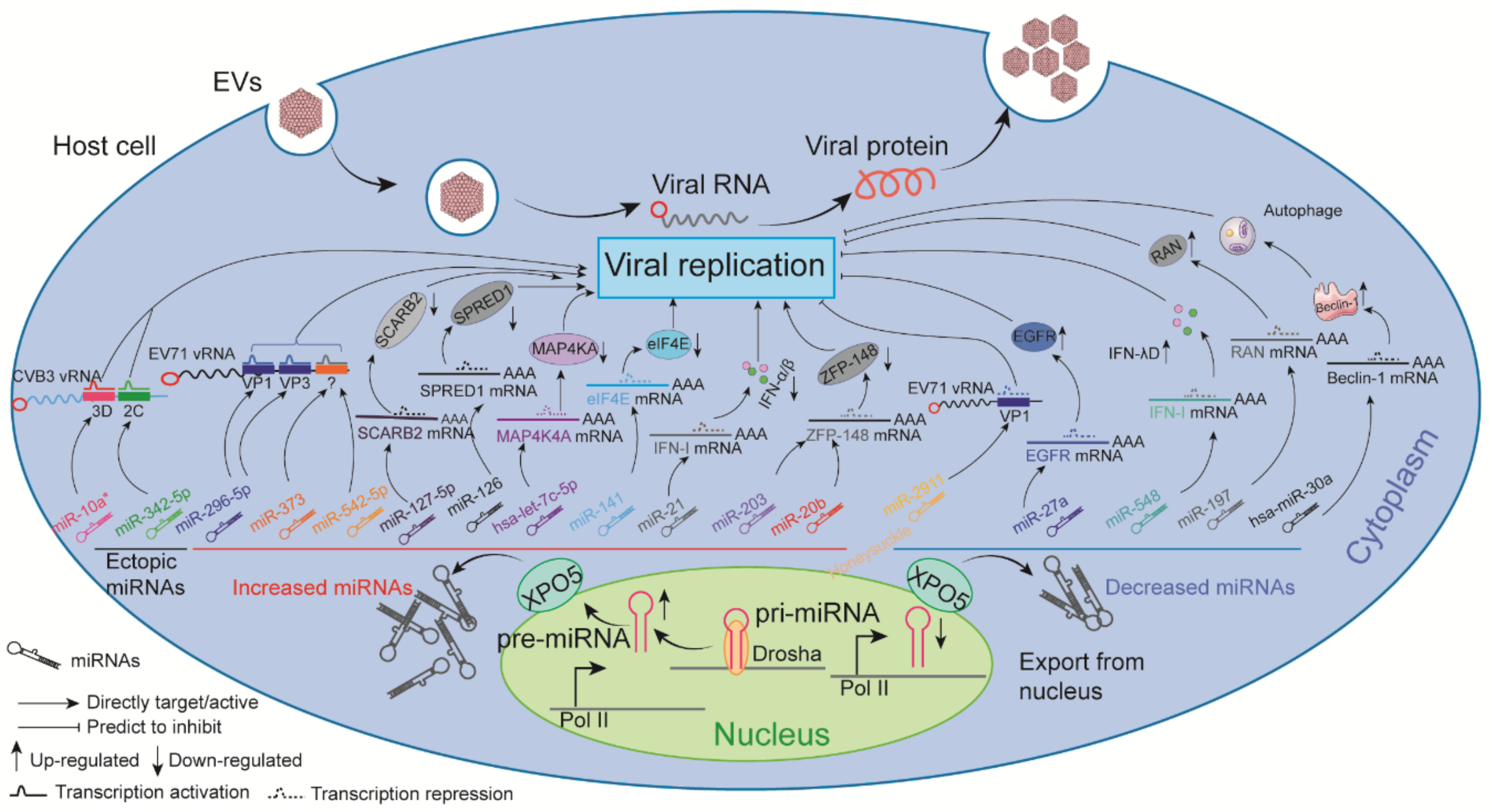
6. Known Impacts of ncRNAs on EV Replication
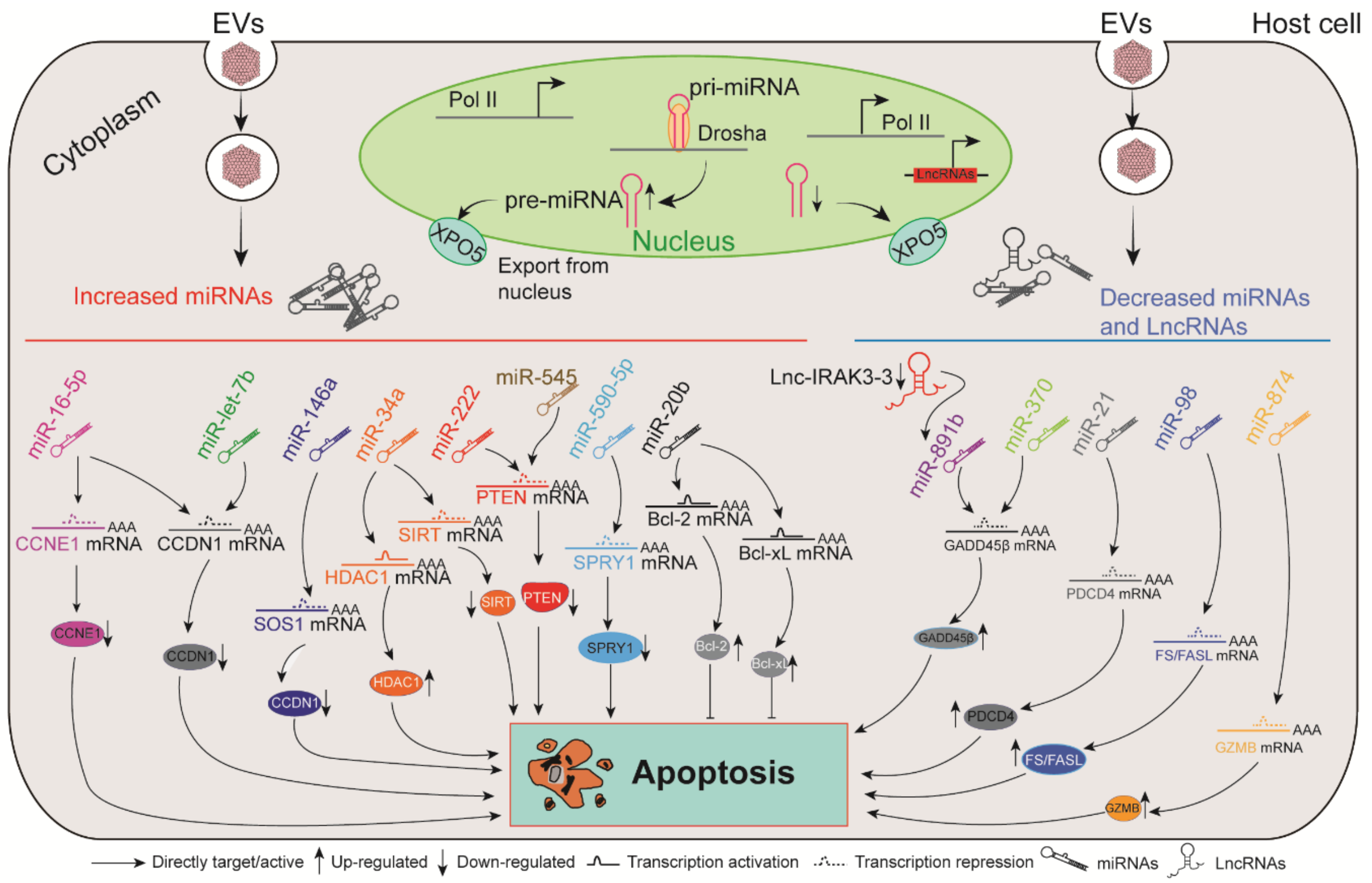
7. The Roles of ncRNAs in EV-Induced Apoptosis
8. Dysregulated ncRNAs Modulate Multiple Signaling Pathways in Response to EV Infection
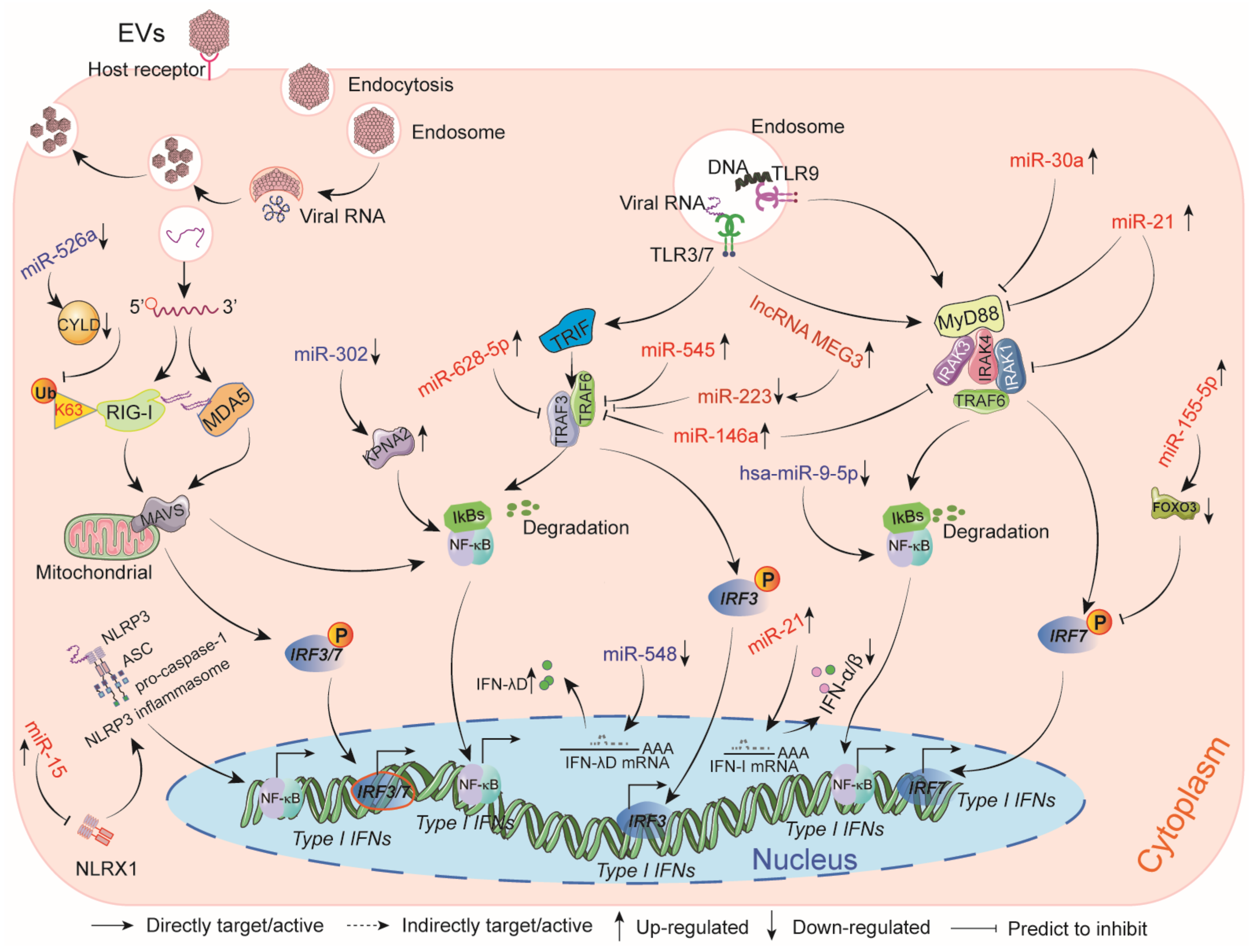
9. The Impact of ncRNAs on Host Innate Immune Responses Induced by EVs
10. Modulation of Immune Cell Function by ncRNAs during EV Infection
11. Circulating ncRNAs Provide Diagnostic Tools for EV-Associated Diseases
12. Current Status of ncRNA-Based Therapeutic Agents for EV Infection
13. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Solomon, T.; Lewthwaite, P.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 10, 778–790. [Google Scholar] [CrossRef]
- Baggen, J.; Thibaut, H.J.; Strating, J.; Van Kuppeveld, F.J.M. The life cycle of non-polio enteroviruses and how to target it. Nat. Rev. Microbiol. 2018, 16, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Pons-Salort, M.; Parker, E.P.; Grassly, N.C. The epidemiology of non-polio enteroviruses: Recent advances and outstanding questions. Curr. Opin. Infect Dis. 2015, 28, 479–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. Clusters of acute respiratory illness associated with human enterovirus 68—Asia, Europe, and United States, 2008–2010. MMWR Morb. Mortal. Wkly. Rep. 2011, 60, 1301–1304. [Google Scholar]
- Pollack, A.; Kontorovich, A.R.; Fuster, V.; Dec, G.W. Viral myocarditis—Diagnosis, treatment options, and current controversies. Nat. Rev. Cardiol. 2015, 12, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Spence-Davizon, E.; Osborne, C.; Press, C.; Schreiner, T.L.; Martin, J.; Messer, R.; Maloney, J.; Burakoff, A.; Barnes, M.; et al. Clinical characteristics of enterovirus A71 neurological disease during an outbreak in children in Colorado, USA, in 2018: An observational cohort study. Lancet Infect. Dis. 2020, 20, 230–239. [Google Scholar] [CrossRef]
- Ikeda, T.; Mizuta, K.; Abiko, C.; Aoki, Y.; Itagaki, T.; Katsushima, F.; Katsushima, Y.; Matsuzaki, Y.; Fuji, N.; Imamura, T.; et al. Acute respiratory infections due to enterovirus 68 in Yamagata, Japan between 2005 and 2010. Microbiol. Immunol. 2012, 56, 139–143. [Google Scholar] [CrossRef]
- Piralla, A.; Lilleri, D.; Sarasini, A.; Marchi, A.; Zecca, M.; Stronati, M.; Baldanti, F.; Gerna, G. Human rhinovirus and human respiratory enterovirus (EV68 and EV104) infections in hospitalized patients in Italy, 2008-2009. Diagn. Microbiol. Infect. Dis. 2012, 73, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Fuji, N.; Suzuki, A.; Tamaki, R.; Saito, M.; Aniceto, R.; Galang, H.; Sombrero, L.; Lupisan, S.; Oshitani, H. Enterovirus 68 among children with severe acute respiratory infection, the Philippines. Emerg. Infect. Dis. 2011, 17, 1430–1435. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.; Van der Sanden, S.; Snijders, B.E.; Jaramillo-Gutierrez, G.; Bont, L.; Van der Ent, C.K.; Overduin, P.; Jenny, S.L.; Jusic, E.; Van der Avoort, H.G.; et al. Emergence and epidemic occurrence of enterovirus 68 respiratory infections in The Netherlands in 2010. Virology 2012, 423, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midgley, C.M.; Jackson, M.A.; Selvarangan, R.; Turabelidze, G.; Obringer, E.; Johnson, D.; Giles, B.L.; Patel, A.; Echols, F.; Oberste, M.S.; et al. Severe respiratory illness associated with enterovirus D68—Missouri and Illinois, 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 798–799. [Google Scholar]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Beermann, J.; Piccoli, M.T.; Viereck, J.; Thum, T. Non-coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Zhang, R.; Wu, W.; Duan, G. Innate Immunity Evasion by Enteroviruses Linked to Epidemic Hand-Foot-Mouth Disease. Front. Microbiol. 2018, 9, 2422. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Schwab, R.; Palatnik, J.F.; Riester, M.; Schommer, C.; Schmid, M.; Weigel, D. Specific effects of microRNAs on the plant transcriptome. Dev. Cell 2005, 8, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siomi, H.; Siomi, M.C. Posttranscriptional regulation of microRNA biogenesis in animals. Mol. Cell 2010, 38, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denli, A.M.; Tops, B.B.; Plasterk, R.H.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, C.; Qin, L.; Li, D.; Zhou, G.; Dang, D.; Chen, S.; Sun, T.; Zhang, R.; Wu, W.; et al. Characterization of Critical Functions of Long Non-Coding RNAs and mRNAs in Rhabdomyosarcoma Cells and Mouse Skeletal Muscle Infected by Enterovirus 71 Using RNA-Seq. Viruses 2018, 10, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Guan, D.; Fan, Q.; Su, J.; Zheng, W.; Ma, W.; Ke, C. lncRNA expression signatures in response to enterovirus 71 infection. Biochem. Biophys. Res. Commun. 2013, 430, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, J.; Jia, L.; Liu, N.; Liang, Y.; Wu, F.; Liang, B.; Li, Y.; Wang, J.; Sheng, C.; et al. Analysis of miRNAs Involved in Mouse Brain Damage upon Enterovirus 71 Infection. Front. Cell. Infect. Microbiol. 2017, 7, 133. [Google Scholar] [CrossRef]
- Cui, L.; Qi, Y.; Li, H.; Ge, Y.; Zhao, K.; Qi, X.; Guo, X.; Shi, Z.; Zhou, M.; Zhu, B.; et al. Serum microRNA expression profile distinguishes enterovirus 71 and coxsackievirus 16 infections in patients with hand-foot-and-mouth disease. PLoS ONE 2011, 6, e27071. [Google Scholar] [CrossRef]
- Meng, J.; Yao, Z.; He, Y.; Zhang, R.; Yang, H.; Yao, X.; Chen, L.; Zhang, H.; Cheng, J. Long non-coding RNA expression profiles in different severity EV71-infected hand foot and mouth disease patients. Biochem. Biophys. Res. Commun. 2017, 493, 1594–1600. [Google Scholar] [CrossRef]
- Jin, J.; Li, R.; Jiang, C.; Zhang, R.; Ge, X.; Liang, F.; Sheng, X.; Dai, W.; Chen, M.; Wu, J.; et al. Transcriptome analysis reveals dynamic changes in coxsackievirus A16 infected HEK 293T cells. BMC Genom. 2017, 18, 933. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Hu, Y.; Li, J.; Zheng, H.; Wang, J.; Guo, L.; Ning, R.; Li, H.; Yang, Z.; Fan, H.; et al. Different microRNA profiles reveal the diverse outcomes induced by EV71 and CA16 infection in human umbilical vein endothelial cells using high-throughput sequencing. PLoS ONE 2017, 12, e0177657. [Google Scholar] [CrossRef]
- Hu, Y.; Song, J.; Liu, L.; Li, J.; Tang, B.; Wang, J.; Zhang, X.; Zhang, Y.; Wang, L.; Liao, Y.; et al. Different microRNA alterations contribute to diverse outcomes following EV71 and CA16 infections: Insights from high-throughput sequencing in rhesus monkey peripheral blood mononuclear cells. Int. J. Biochem. Cell Biol. 2016, 81, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Qiu, Y.; Wang, H.; Qu, Y.; Zhao, Y.; Lin, L.; Wang, Y.; Xu, W.; Zhao, W.; He, H.; et al. Expression Profile and Function Analysis of Long Non-coding RNAs in the Infection of Coxsackievirus B3. Virol Sin. 2019, 34, 618–630. [Google Scholar] [CrossRef]
- Yao, H.L.; Liu, M.; Wang, W.J.; Wang, X.L.; Song, J.; Song, Q.Q.; Han, J. Construction of miRNA-target networks using microRNA profiles of CVB3-infected HeLa cells. Sci. Rep. 2019, 9, 17876. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Tong, L.; Zhao, W.; Wang, Y.; Meng, Y.; Lin, L.; Liu, B.; Zhai, Y.; Zhong, Z.; Li, X. Microarray analysis reveals altered circulating microRNA expression in mice infected with Coxsackievirus B3. Exp. Ther Med. 2016, 12, 2220–2226. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, Z.; He, F.; Zou, J.; Wu, S.; Liu, Z. MicroRNAs regulate the pathogenesis of CVB3-induced viral myocarditis. Intervirology 2013, 56, 104–113. [Google Scholar] [CrossRef]
- Liu, Y.L.; Wu, W.; Xue, Y.; Gao, M.; Yan, Y.; Kong, Q.; Pang, Y.; Yang, F. MicroRNA-21 and -146b are involved in the pathogenesis of murine viral myocarditis by regulating TH-17 differentiation. Arch. Virol. 2013, 158, 1953–1963. [Google Scholar] [CrossRef]
- Corsten, M.F.; Papageorgiou, A.; Verhesen, W.; Carai, P.; Lindow, M.; Obad, S.; Summer, G.; Coort, S.L.; Hazebroek, M.; van Leeuwen, R.; et al. MicroRNA profiling identifies microRNA-155 as an adverse mediator of cardiac injury and dysfunction during acute viral myocarditis. Circ. Res. 2012, 111, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.W.; Ho, A.; Alshabee-Akil, A.; Hardikar, A.A.; Kay, T.W.; Rawlinson, W.D.; Craig, M.E. Coxsackievirus B5 Infection Induces Dysregulation of microRNAs Predicted to Target Known Type 1 Diabetes Risk Genes in Human Pancreatic Islets. Diabetes 2016, 65, 996–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, W.Y.; Cheung, A.C.; Tung, C.K.; Yeung, A.C.; Ngai, K.L.; Lui, V.W.; Chan, P.K.; Tsui, S.K. miR-466 is putative negative regulator of Coxsackie virus and Adenovirus Receptor. FEBS Lett 2015, 589, 246–254. [Google Scholar] [CrossRef]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA Regulation of RNA Virus Replication and Pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xiong, Y.; Zeng, L.; Peng, Z.; Liu, Z.; Zhan, H.; Yang, Z. The Role of Non-coding RNAs in Viral Myocarditis. Front. Cell Infect. Microbiol. 2020, 10, 312. [Google Scholar] [CrossRef]
- Tong, L.; Lin, L.; Wu, S.; Guo, Z.; Wang, T.; Qin, Y.; Wang, R.; Zhong, X.; Wu, X.; Wang, Y.; et al. MiR-10a* up-regulates coxsackievirus B3 biosynthesis by targeting the 3D-coding sequence. Nucleic Acids Res. 2013, 41, 3760–3771. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Qin, Y.; Tong, L.; Wu, S.; Wang, Q.; Jiao, Q.; Guo, Z.; Lin, L.; Wang, R.; Zhao, W.; et al. MiR-342-5p suppresses coxsackievirus B3 biosynthesis by targeting the 2C-coding region. Antiviral Res. 2012, 93, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.G.; Ye, X.; Zhang, H.M.; Hanson, P.J.; Liu, Z.; McManus, B.M.; Yang, D. MicroRNA-203 enhances coxsackievirus B3 replication through targeting zinc finger protein-148. Cell Mol. Life Sci. 2013, 70, 277–291. [Google Scholar] [CrossRef]
- Xu, H.F.; Gao, X.T.; Lin, J.Y.; Xu, X.H.; Hu, J.; Ding, Y.J.; Zhu, S.H. MicroRNA-20b suppresses the expression of ZFP-148 in viral myocarditis. Mol. Cell Biochem. 2017, 429, 199–210. [Google Scholar] [CrossRef]
- Ye, X.; Hemida, M.G.; Qiu, Y.; Hanson, P.J.; Zhang, H.M.; Yang, D. MiR-126 promotes coxsackievirus replication by mediating cross-talk of ERK1/2 and Wnt/beta-catenin signal pathways. Cell. Mol. Life Sci. 2013, 70, 4631–4644. [Google Scholar] [CrossRef]
- Corsten, M.F.; Heggermont, W.; Papageorgiou, A.P.; Deckx, S.; Tijsma, A.; Verhesen, W.; Van Leeuwen, R.; Carai, P.; Thibaut, H.J.; Custers, K.; et al. The microRNA-221/-222 cluster balances the antiviral and inflammatory response in viral myocarditis. Eur. Heart J. 2015, 36, 2909–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Ke, X.; Wang, M.; He, S.; Li, Q.; Zheng, C.; Zhang, Z.; Liu, Y.; Wang, H. Human microRNA hsa-miR-296-5p suppresses enterovirus 71 replication by targeting the viral genome. J. Virol. 2013, 87, 5645–5656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Tien, P. MiR373 and miR542-5p regulate the replication of enterovirus 71 in rhabdomyosarcoma cells. Sheng Wu Gong Cheng Xue Bao 2014, 30, 943–953. [Google Scholar]
- Li, X.; Huang, Y.; Sun, M.; Ji, H.; Dou, H.; Hu, J.; Yan, Y.; Wang, X.; Chen, L. Honeysuckle-encoded microRNA2911 inhibits Enterovirus 71 replication via targeting VP1 gene. Antivir. Res. 2018, 152, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Fu, Y.; Chen, D.; Wang, H.; Su, A.; Zhang, L.; Chang, L.; Zheng, N.; Wu, Z. miR-127-5p negatively regulates enterovirus 71 replication by directly targeting SCARB2. FEBS Open Bio 2017, 7, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, X.; Shi, Y.; Zhou, B.; Du, C.; Liu, Y.; Han, S.; Yin, J.; Peng, B.; He, X.; et al. miR-27a suppresses EV71 replication by directly targeting EGFR. Virus Genes 2014, 49, 373–382. [Google Scholar] [CrossRef]
- Zhou, B.; Chu, M.; Xu, S.; Chen, X.; Liu, Y.; Wang, Z.; Zhang, F.; Han, S.; Yin, J.; Peng, B.; et al. Hsa-let-7c-5p augments enterovirus 71 replication through viral subversion of cell signaling in rhabdomyosarcoma cells. Cell Biosci. 2017, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Xu, W.; Chen, D.; Feng, C.; Zhang, L.; Wang, X.; Lv, X.; Zheng, N.; Jin, Y.; Wu, Z. Enterovirus 71 induces autophagy by regulating has-miR-30a expression to promote viral replication. Antivir. Res. 2015, 124, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.F.; Huang, R.T.; Chien, K.Y.; Huang, J.Y.; Lau, K.S.; Jheng, J.R.; Chiu, C.H.; Wu, T.Y.; Chen, C.Y.; Horng, J.T. Host MicroRNA miR-197 Plays a Negative Regulatory Role in the Enterovirus 71 Infectious Cycle by Targeting the RAN Protein. J. Virol. 2016, 90, 1424–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, B.C.; Yu, S.L.; Chen, J.J.; Chang, S.Y.; Yan, B.S.; Hong, Q.S.; Singh, S.; Kao, C.L.; Chen, H.Y.; Su, K.Y.; et al. Enterovirus-induced miR-141 contributes to shutoff of host protein translation by targeting the translation initiation factor eIF4E. Cell Host Microbe 2011, 9, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Feng, L.; Li, J.; Xu, H.; Mei, X.; Feng, L.; Sun, H.; Gao, J.; Zhang, X. miR-545 promoted enterovirus 71 replication via directly targeting phosphatase and tensin homolog and tumor necrosis factor receptor-associated factor 6. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; He, X.; Zheng, Z.; Zhang, Z.; Wei, C.; Guan, K.; Hou, L.; Zhang, B.; Zhu, L.; Cao, Y.; et al. Downregulation of microRNA miR-526a by enterovirus inhibits RIG-I-dependent innate immune response. J. Virol. 2014, 88, 11356–11368. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xie, J.; Xu, X.; Wang, J.; Ao, F.; Wan, Y.; Zhu, Y. MicroRNA-548 down-regulates host antiviral response via direct targeting of IFN-lambda1. Protein Cell 2013, 4, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Ho, B.C.; Yu, I.S.; Lu, L.F.; Rudensky, A.; Chen, H.Y.; Tsai, C.W.; Chang, Y.L.; Wu, C.T.; Chang, L.Y.; Shih, S.R.; et al. Inhibition of miR-146a prevents enterovirus-induced death by restoring the production of type I interferon. Nat. Commun. 2014, 5, 3344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, N.; Zhou, Z.; Li, Y.; Zhao, L.; Xue, Z.; Lu, R.; Jia, K. Enterovirus 71-induced has-miR-21 contributes to evasion of host immune system by targeting MyD88 and IRAK1. Virus Res. 2017, 237, 27–36. [Google Scholar] [CrossRef]
- Kvansakul, M. Viral Infection and Apoptosis. Viruses 2017, 9, 356. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Chen, Y.; Cheng, X.; Cai, Z.; Qiu, S. GATA1/SP1 and miR-874 mediate enterovirus-71-induced apoptosis in a granzyme-B-dependent manner in Jurkat cells. Arch. Virol. 2020, 165, 2531–2540. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.Q.; Zhang, X.Y.; Guo, H.; Yang, Y.; An, S.; Liu, Y.; Guo, X.X.; Xu, T.R.; Hao, Q. Research progress on pathophysiological function of SOS1 protein. Sheng Li Xue Bao 2018, 70, 565–570. [Google Scholar] [PubMed]
- Feng, X.; Koh, D.W. Chapter Five—Roles of Poly(ADP-Ribose) Glycohydrolase in DNA Damage and Apoptosis. In International Review of Cell and Molecular Biology; Jeon, K.W., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 304, pp. 227–281. [Google Scholar]
- Chang, Y.L.; Ho, B.C.; Sher, S.; Yu, S.L.; Yang, P.C. miR-146a and miR-370 coordinate enterovirus 71-induced cell apoptosis through targeting SOS1 and GADD45β. Cell. Microbiol. 2015, 17, 802–818. [Google Scholar] [CrossRef]
- Liao, Y.W.; Ho, B.C.; Chen, M.H.; Yu, S.L. Host relieves lnc-IRAK3-3-sequestered miR-891b to attenuate apoptosis in Enterovirus 71 infection. Cell. Microbiol. 2019, 21, e13043. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zheng, Z.; Sun, J.; Zhang, Y.; Wei, C.; Ke, X.; Liu, Y.; Deng, L.; Wang, H. MiR-16-5p mediates a positive feedback loop in EV71-induced apoptosis and suppresses virus replication. Sci. Rep. 2017, 7, 16422. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Wang, H.; Xu, F.; Huang, Y.; Liu, Z.; Liu, T. Enterovirus 71 induces apoptosis of SHSY5Y human neuroblastoma cells through stimulation of endogenous microRNA let-7b expression. Mol. Med. Rep. 2015, 12, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Li, M.; Yu, Y.; Shi, H.; Chen, R. microRNA-34a aggravates coxsackievirus B3-induced apoptosis of cardiomyocytes through the SIRT1-p53 pathway. J. Med. Virol. 2019, 91, 1643–1651. [Google Scholar] [CrossRef]
- He, J.; Yue, Y.; Dong, C.; Xiong, S. MiR-21 confers resistance against CVB3-induced myocarditis by inhibiting PDCD4-mediated apoptosis. Clin. Invest. Med. 2013, 36, E103–E111. [Google Scholar] [CrossRef] [Green Version]
- Germano, J.F.; Sawaged, S.; Saadaeijahromi, H.; Andres, A.M.; Feuer, R.; Gottlieb, R.A.; Sin, J. Coxsackievirus B infection induces the extracellular release of miR-590-5p, a proviral microRNA. Virology 2019, 529, 169–176. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Zhao, Z.; Jin, Z. Expression of miR-98 in myocarditis and its influence on transcription of the FAS/FASL gene pair. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chen, J.; He, L.; Stiles, B.L. PTEN: Tumor Suppressor and Metabolic Regulator. Front. Endocrinol. 2018, 9, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Gao, X.; Hu, J.; Xie, Y.; Zuo, Y.; Xu, H.; Zhu, S. ADAR1p150 Forms a Complex with Dicer to Promote miRNA-222 Activity and Regulate PTEN Expression in CVB3-Induced Viral Myocarditis. Int. J. Mol. Sci. 2019, 20, 407. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, I.; Alidjinou, E.K.; Bertin, A.; Sane, F.; Hober, D. miRNAs in enterovirus infection. Crit. Rev. Microbiol. 2018, 44, 701–714. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-kappaB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [Green Version]
- Fei, Y.; Chaulagain, A.; Wang, T.; Chen, Y.; Liu, J.; Yi, M.; Wang, Y.; Huang, Y.; Lin, L.; Chen, S.; et al. MiR-146a down-regulates inflammatory response by targeting TLR3 and TRAF6 in Coxsackievirus B infection. Rna 2020, 26, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.G.; Liu, H.; Zhang, J.B.; Zhang, S.L.; Zhao, L.H.; Liang, W.Q. Upregulated microRNA-214 enhances cardiac injury by targeting ITCH during coxsackievirus infection. Mol. Med. Rep. 2015, 12, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Chen, K.H.; Dong, X.M.; Fang, Y.; Li, W.G.; Huang, G.Y.; Song, W. A role of pre-mir-10a coding region variant in host susceptibility to coxsackie virus-induced myocarditis. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3500–3507. [Google Scholar] [PubMed]
- Cao, H.; Yang, B.; Zhao, Y.; Deng, X.; Shen, X. The pro-apoptosis and pro-inflammation role of LncRNA HIF1A-AS1 in Coxsackievirus B3-induced myocarditis via targeting miR-138. Cardiovasc. Diagn. Ther. 2020, 10, 1245–1255. [Google Scholar] [CrossRef]
- Xia, K.; Zhang, Y.; Sun, D. miR-217 and miR-543 downregulation mitigates inflammatory response and myocardial injury in children with viral myocarditis by regulating the SIRT1/AMPK/NF-κB signaling pathway. Int. J. Mol. Med. 2020, 45, 634–646. [Google Scholar] [CrossRef]
- Li, B.; Zheng, J. MicroR-9-5p suppresses EV71 replication through targeting NFκB of the RIG-I-mediated innate immune response. FEBS Open Bio 2018, 8, 1457–1470. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.X.; Bai, Z.F.; Wang, W.; Wu, H.Y. Effects of microrna-93 on mouse cardiac microvascular endothelial cells injury and inflammatory response by mediating SPP1 through the NF-KappaB pathway. J. Cell. Biochem. 2019, 120, 2847–2858. [Google Scholar] [CrossRef]
- Jafari, M.; Ghadami, E.; Dadkhah, T.; Akhavan-Niaki, H. PI3k/AKT signaling pathway: Erythropoiesis and beyond. J. Cell. Physiol. 2019, 234, 2373–2385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xiong, Y.; Xu, J.; Chen, S.; Li, P.; Huang, Y.; Wang, Y.; Chen, W.X.; Wang, B. Host MicroRNA hsa-miR-494-3p Promotes EV71 Replication by Directly Targeting PTEN. Front. Cell. Infect. Microbiol. 2018, 8, 278. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paroo, Z.; Ye, X.; Chen, S.; Liu, Q. Phosphorylation of the human microRNA-generating complex mediates MAPK/Erk signaling. Cell 2009, 139, 112–122. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Xiao, Z.; Yao, H.; Li, S.; Feng, M.; Wang, W.; Liu, Z.; Liu, Z.; Wu, J. The protective role of microRNA-21 against coxsackievirus B3 infection through targeting the MAP2K3/P38 MAPK signaling pathway. J. Transl. Med. 2019, 17, 335. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Wang, Y.; Bian, L.; Liu, Q.; Long, J.E. Enterovirus 71 antagonizes the antiviral activity of host STAT3 and IL-6R with partial dependence on virus-induced miR-124. J. Gen. Virol. 2017, 98, 3008–3025. [Google Scholar] [CrossRef]
- Peng, N.; Yang, X.; Zhu, C.; Zhou, L.; Yu, H.; Li, M.; Lin, Y.; Wang, X.; Li, Q.; She, Y.; et al. MicroRNA-302 Cluster Downregulates Enterovirus 71-Induced Innate Immune Response by Targeting KPNA2. J. Immunol. 2018, 201, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.M.; Chen, M.G.; Chen, D.W.; Wu, W.F.; Liu, Y.L.; Lin, F.H. The effect of microRNA-21 on myocardial fibrosis in mice with chronic viral myocarditis. Zhonghua Xin Xue Guan Bing Za Zhi 2018, 46, 450–457. [Google Scholar] [CrossRef]
- Wilson, C.C.; Schooley, R.T. 3—Host Responses to Infection. In Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 26–39.e2. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef]
- Schenten, D.; Medzhitov, R. The control of adaptive immune responses by the innate immune system. Adv. Immunol. 2011, 109, 87–124. [Google Scholar] [CrossRef]
- Kemball, C.C.; Alirezaei, M.; Whitton, J.L. Type B coxsackieviruses and their interactions with the innate and adaptive immune systems. Future Microbiol. 2010, 5, 1329–1347. [Google Scholar] [CrossRef] [Green Version]
- Häcker, H.; Tseng, P.H.; Karin, M. Expanding TRAF function: TRAF3 as a tri-faced immune regulator. Nat. Rev. Immunol. 2011, 11, 457–468. [Google Scholar] [CrossRef]
- Li, D.; Chen, S.; Zhang, W.; Zhang, C.; Sun, T.; Du, Y.; Ding, R.; Gao, Y.; Jin, Y.; Duan, G. MicroRNA-628-5p Facilitates Enterovirus 71 Infection by Suppressing TRAF3 Signaling. Cell. Mol. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.L.; Zhang, S.X.; Zheng, C.F.; Li, Y.F.; Zhang, L.H.; Su, Q.Y.; Hao, Y.F.; Wang, S.; Li, X.W. Long non-coding RNA MEG3 inhibits M2 macrophage polarization by activating TRAF6 via microRNA-223 down-regulation in viral myocarditis. J. Cell. Mol. Med. 2020, 24, 12341–12354. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. TLR signaling pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Song, W.; Zhang, W.; Li, J.; Li, C.; Qiu, Y.; Fang, Y.; Jiang, Q.; Li, X.; et al. Exosomes from EV71-infected oral epithelial cells can transfer miR-30a to promote EV71 infection. Oral Dis. 2020, 26, 778–788. [Google Scholar] [CrossRef]
- Yang, D.; Wang, X.; Gao, H.; Chen, B.; Si, C.; Wang, S. Downregulation of miR-155-5p facilitates enterovirus 71 replication through suppression of type I IFN response by targeting FOXO3/IRF7 pathway. Cell Cycle 2020, 19, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Tong, R.; Jia, T.; Shi, R.; Yan, F. Inhibition of microRNA-15 protects H9c2 cells against CVB3-induced myocardial injury by targeting NLRX1 to regulate the NLRP3 inflammasome. Cell. Mol. Biol. Lett. 2020, 25, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, C.X.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.F.; Wei, J.; Yao, R.W.; Yang, L.; Chen, L.L. Coordinated circRNA Biogenesis and Function with NF90/NF110 in Viral Infection. Mol. Cell 2017, 67, 214–227.e217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.G.; Kim, M.V.; Chen, X.; Batista, P.J.; Aoyama, S.; Wilusz, J.E.; Iwasaki, A.; Chang, H.Y. Sensing Self and Foreign Circular RNAs by Intron Identity. Mol. Cell 2017, 67, 228–238.e225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildreth, A.D.; O’Sullivan, T.E. Tissue-Resident Innate and Innate-Like Lymphocyte Responses to Viral Infection. Viruses 2019, 11, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.Y.; Hsiung, C.A.; Lu, C.Y.; Lin, T.Y.; Huang, F.Y.; Lai, Y.H.; Chiang, Y.P.; Chiang, B.L.; Lee, C.Y.; Huang, L.M. Status of cellular rather than humoral immunity is correlated with clinical outcome of enterovirus 71. Pediatr. Res. 2006, 60, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Kong, Y.; Han, B.; Jiang, D.; Jia, H.; Zhang, L. Long Non-coding RNA Expression Profile and Functional Analysis in Children With Acute Fulminant Myocarditis. Front. Pediatr. 2019, 7, 283. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Han, B. Dysregulated CD4+ T Cells and microRNAs in Myocarditis. Front. Immunol. 2020, 11, 539. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Li, X.; Tang, Z.; Wang, X.; Zhong, M.; Suo, Q.; Zhang, Y.; Lv, K. Silencing MicroRNA-155 Attenuates Cardiac Injury and Dysfunction in Viral Myocarditis via Promotion of M2 Phenotype Polarization of Macrophages. Sci. Rep. 2016, 6, 22613. [Google Scholar] [CrossRef]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, X.; Wang, C.; Zhang, M.; Yang, H.; Lv, K. lncRNA AK085865 Promotes Macrophage M2 Polarization in CVB3-Induced VM by Regulating ILF2-ILF3 Complex-Mediated miRNA-192 Biogenesis. Mol. Ther. Nucleic Acids 2020, 21, 441–451. [Google Scholar] [CrossRef]
- Gou, W.; Zhang, Z.; Yang, C.; Li, Y. MiR-223/Pknox1 axis protects mice from CVB3-induced viral myocarditis by modulating macrophage polarization. Exp. Cell Res. 2018, 366, 41–48. [Google Scholar] [CrossRef]
- Grasedieck, S.; Schöler, N.; Bommer, M.; Niess, J.H.; Tumani, H.; Rouhi, A.; Bloehdorn, J.; Liebisch, P.; Mertens, D.; Döhner, H.; et al. Impact of serum storage conditions on microRNA stability. Leukemia 2012, 26, 2414–2416. [Google Scholar] [CrossRef]
- Zhang, X.; Hong, R.; Chen, W.; Xu, M.; Wang, L. The role of long noncoding RNA in major human disease. Bioorg. Chem. 2019, 92, 103214. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.L.; Weng, K.F.; Huang, Y.C.; Chen, C.J. Elevated expression of circulating miR876-5p is a specific response to severe EV71 infections. Sci. Rep. 2016, 6, 24149. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Pietsch, H.; Escher, F.; Schultheiss, H.P. MicroRNA profiling as a novel diagnostic tool for identification of patients with inflammatory and/or virally induced cardiomyopathies. ESC Heart Fail. 2020. [Google Scholar] [CrossRef]
- Kuehl, U.; Lassner, D.; Gast, M.; Stroux, A.; Rohde, M.; Siegismund, C.; Wang, X.; Escher, F.; Gross, M.; Skurk, C.; et al. Differential Cardiac MicroRNA Expression Predicts the Clinical Course in Human Enterovirus Cardiomyopathy. Circ. Heart Fail. 2015, 8, 605–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, L.; Tirosh-Wagner, T.; Vardi, A.; Abbas, H.; Pillar, N.; Shomron, N.; Nevo-Caspi, Y.; Paret, G. Circulating MicroRNAs: A Potential Biomarker for Cardiac Damage, Inflammatory Response, and Left Ventricular Function Recovery in Pediatric Viral Myocarditis. J. Cardiovasc. Transl. Res. 2018, 11, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, M.; Zhao, C.; Chen, C.; Kong, Q.; Cai, Z.; Li, D. MiR-1/133 attenuates cardiomyocyte apoptosis and electrical remodeling in mice with viral myocarditis. Cardiol. J. 2020, 27, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchie, A. First microRNA mimic enters clinic. Nat. Biotechnol. 2013, 31, 577. [Google Scholar] [CrossRef]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; Van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, E.J.; Hadac, E.M.; Greiner, S.; Russell, S.J. Engineering microRNA responsiveness to decrease virus pathogenicity. Nat. Med. 2008, 14, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.; Kunitomi, M.; Vignuzzi, M.; Saksela, K.; Andino, R. Harnessing endogenous miRNAs to control virus tissue tropism as a strategy for developing attenuated virus vaccines. Cell Host Microbe 2008, 4, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.F.; Ding, Y.J.; Shen, Y.W.; Xue, A.M.; Xu, H.M.; Luo, C.L.; Li, B.X.; Liu, Y.L.; Zhao, Z.Q. MicroRNA- 1 represses Cx43 expression in viral myocarditis. Mol. Cell. Biochem. 2012, 362, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xuan, L.; Liu, Y.; Shao, L.; Ge, H.; Gu, J.; Wei, C.; Zhao, M. Astragalus Root dry extract restores connexin43 expression by targeting miR-1 in viral myocarditis. Phytomedicine 2018, 46, 32–38. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Subtype | EVs | Common Diseases Caused by EVs |
|---|---|---|
| Types-A | CVA6, CVA16, EVA71, other serotypes | HFMD, Herpangina, Aseptic meningitis, Pulmonary edema, Neonatal sepsis, Acute febrile disease |
| Types-B | CVB1-B6, 7 of the original echoviruses | HFMD, Aseptic meningitis, Respiratory disorders, Meningitis, Viral myocarditis, Herpangina, Neonatal sepsis |
| Types-C | polioviruses and other serotypes | Poliomyelitis, Respiratory disorders, Muscle inflammation, Neonatal sepsis |
| Types-D | EV-D68, EV-D70, EV-D94, EV-D111, and EV-D120 | Respiratory disorders, Acute flaccid myelitis (AFM), Aseptic meningitis |
| Virus Type | Model | ncRNA | Total | Up | Down | qRT-PCR | References |
|---|---|---|---|---|---|---|---|
| EVA71 | in vitro | lncRNA | 23 | 18 | 5 | YES | [22] |
| EVA71 | in vivo | lncRNA | 104 | 72 | 32 | YES | [22] |
| EVA71 | in vitro | lncRNA | 4866 | 2990 | 1876 | YES | [23] |
| EVA71 | in vitro | miRNA | 4270 | 1718 | 2552 | NO | [23] |
| EVA71 | in vivo | miRNA | 20 | 4 | 16 | YES | [24] |
| EVA71/CVA16 | human | miRNA | 128 | 102 | 26 | YES | [25] |
| EVA71 | ex vivo | lncRNA | 8541 | - | - | YES | [26] |
| CVA16 | in vitro | miRNA | 1954 | 1825 | 129 | YES | [27] |
| EVA71/CVA16 | in vitro | miRNA | 39/92 | 14/36 | 25/56 | YES | [28] |
| EVA71/CVA16 | ex vivo | miRNA | 76/42 | 10/12 | 66/30 | YES | [29] |
| CVB3 | in vitro | lncRNA | 700 | 431 | 269 | NO | [30] |
| CVB3 | in vitro | miRNA | 597 | - | - | YES | [31] |
| CVB3 | in vivo | miRNA | 96 | 33 | 63 | YES | [32] |
| CVB3 | in vivo | miRNA | 5 | 4 | 1 | YES | [33] |
| CVB3 | in vivo | miRNA | 3 | 2 | 1 | YES | [34] |
| CVB3 | human | miRNA | 107 | - | - | YES | [35] |
| CVB5 | in vivo | miRNA | 33 | - | - | YES | [36] |
| CVB4 | in vitro | miRNA | 7 | - | - | YES | [37] |
| Signaling Pathway | Influence | NcRNAs | Expression | Viral Replication | Model | References |
|---|---|---|---|---|---|---|
| NF-κB | inhibition | miR-146a | Upregulated | Unknown | in vitro | [78] |
| inhibition | miR-217 | Upregulated | Unknown | in vivo; in vitro | [82] | |
| inhibition | miR-543 | Upregulated | Unknown | in vivo; in vitro | [82] | |
| inhibition | miR-9-5p | Downregulated | Unknown | in vivo; in vitro | [83] | |
| activation | miR-214 | Upregulated | Unknown | in vivo; | [79] | |
| activation | LncRNA HIF1A-AS1 | Upregulated | Unknown | in vivo; in vitro | [81] | |
| unknown | miR-93 | Upregulated | Unknown | in vivo; in vitro | [84] | |
| activation | miR-526a | Upregulated | Inhibition | in vitro | [56] | |
| PI3K/AKT | inhibition | miR-27a | Downregulated | Inhibition | in vitro | [50] |
| activation | miR-494-3p | Upregulated | Promotion | in vitro | [86] | |
| MAPK | inhibition | miR-21 | Upregulated | Unknown | in vivo; in vitro | [89] |
| inhibition | let-7c-5p | Upregulated | Promotion | in vitro | [51] | |
| JAK/STAT | inhibition | miR-124 | Upregulated | Promotion | in vitro | [90] |
| inhibition | miR-217 | Upregulated | Unknown | in vivo; in vitro | [82] | |
| inhibition | miR-543 | Upregulated | Unknown | in vivo; in vitro | [82] | |
| Wnt/β-catenin | inhibition | miR-126 | Upregulated | Promotion | in vitro | [44] |
| AMPK | inhibition | miR-217 | Upregulated | Unknown | in vivo; in vitro | [82] |
| inhibition | miR-543 | Upregulated | Unknown | in vivo; in vitro | [82] | |
| JNK | inhibition | miR-302 | Downregulated | Inhibition | in vivo; in vitro | [91] |
| activation | let-7c-5p | Upregulated | Promotion | in vitro | [51] | |
| TGF-β1/Smad7 | inhibition | miR-21 | Unknown | Unknown | in vivo; | [92] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, P.; Chen, S.; Zhang, W.; Duan, G.; Jin, Y. Essential Role of Non-Coding RNAs in Enterovirus Infection: From Basic Mechanisms to Clinical Prospects. Int. J. Mol. Sci. 2021, 22, 2904. https://doi.org/10.3390/ijms22062904
Zhu P, Chen S, Zhang W, Duan G, Jin Y. Essential Role of Non-Coding RNAs in Enterovirus Infection: From Basic Mechanisms to Clinical Prospects. International Journal of Molecular Sciences. 2021; 22(6):2904. https://doi.org/10.3390/ijms22062904
Chicago/Turabian StyleZhu, Peiyu, Shuaiyin Chen, Weiguo Zhang, Guangcai Duan, and Yuefei Jin. 2021. "Essential Role of Non-Coding RNAs in Enterovirus Infection: From Basic Mechanisms to Clinical Prospects" International Journal of Molecular Sciences 22, no. 6: 2904. https://doi.org/10.3390/ijms22062904
APA StyleZhu, P., Chen, S., Zhang, W., Duan, G., & Jin, Y. (2021). Essential Role of Non-Coding RNAs in Enterovirus Infection: From Basic Mechanisms to Clinical Prospects. International Journal of Molecular Sciences, 22(6), 2904. https://doi.org/10.3390/ijms22062904