
3D Environment Is Required In Vitro to Demonstrate Altered Bone Metabolism Characteristic for Type 2 Diabetics

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
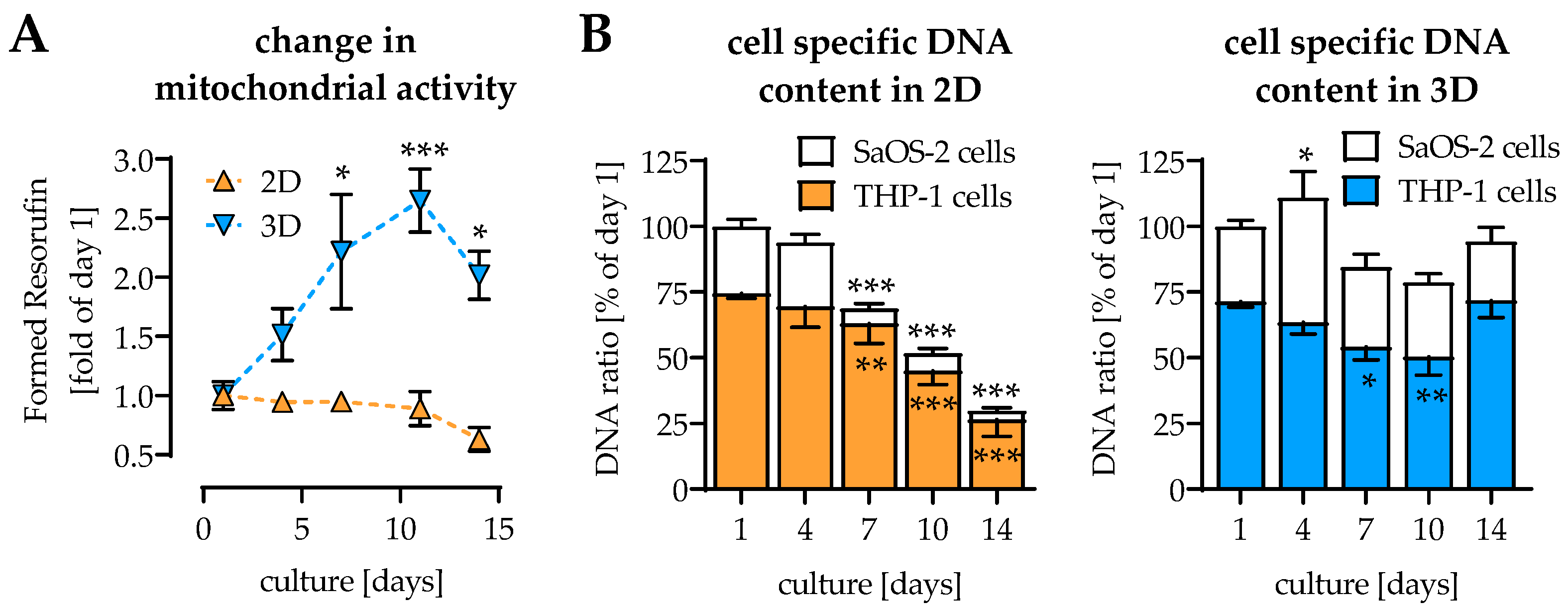
2.1. Coculture and 3D Environment Stabilize the Cell Culture
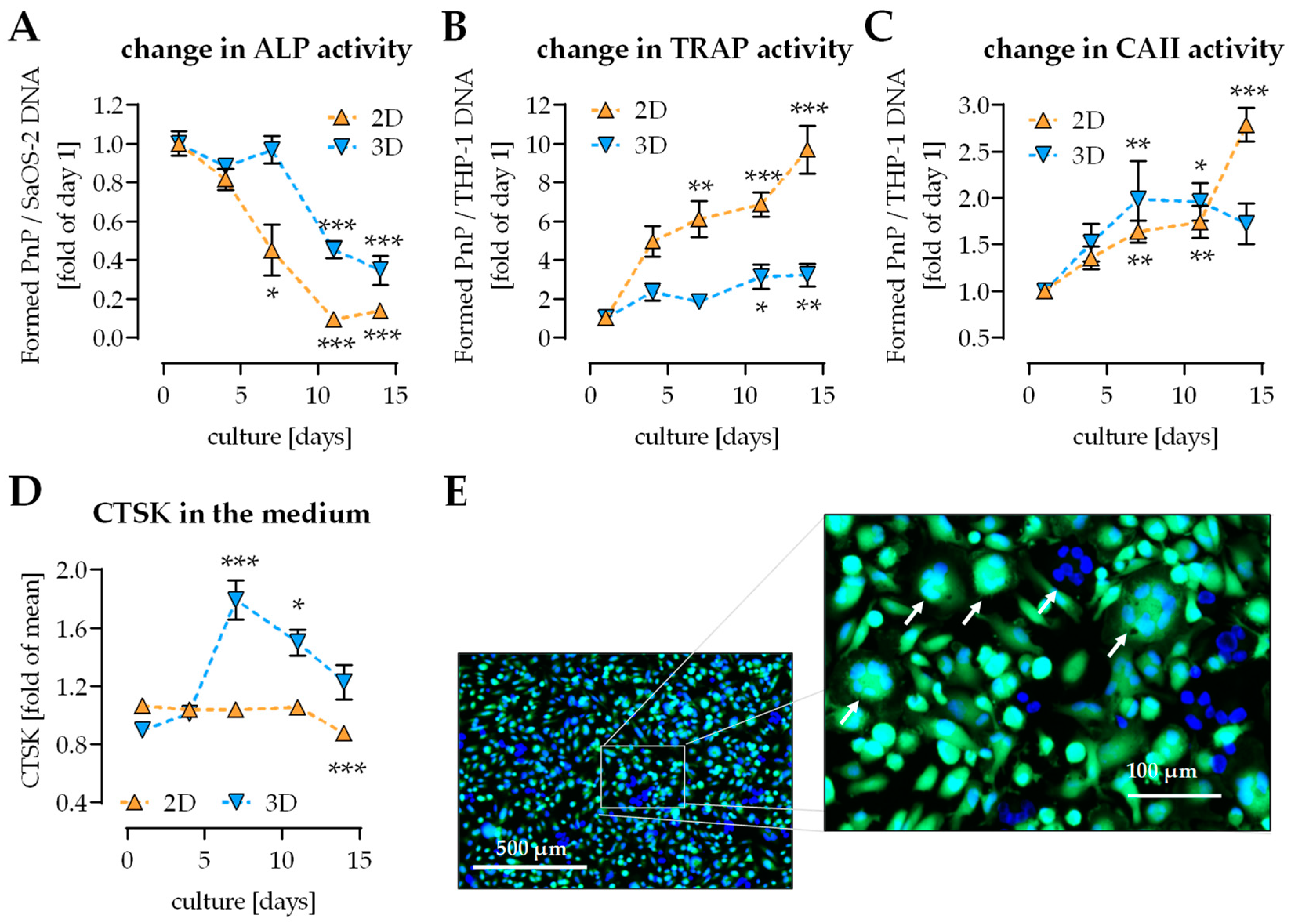
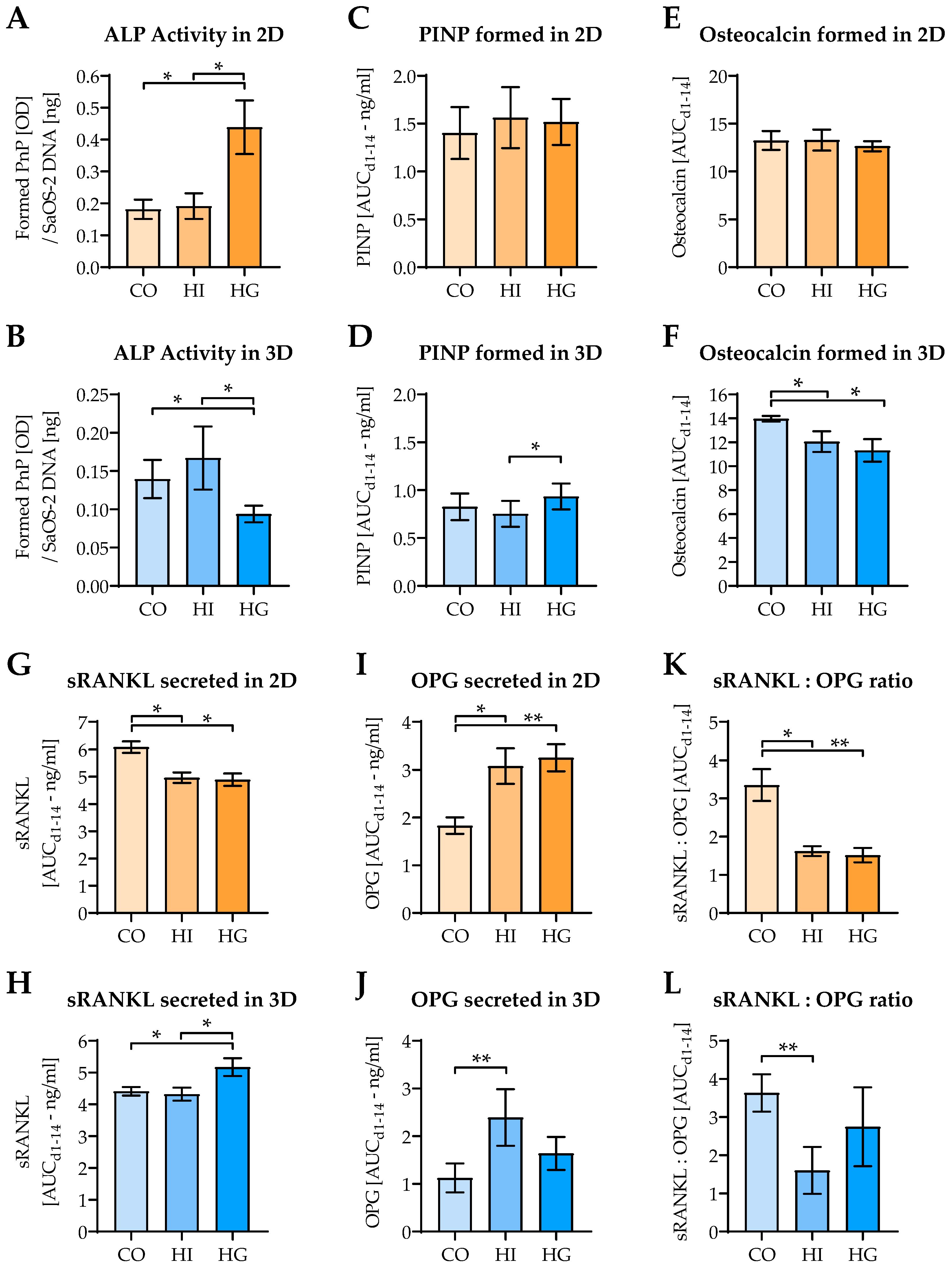
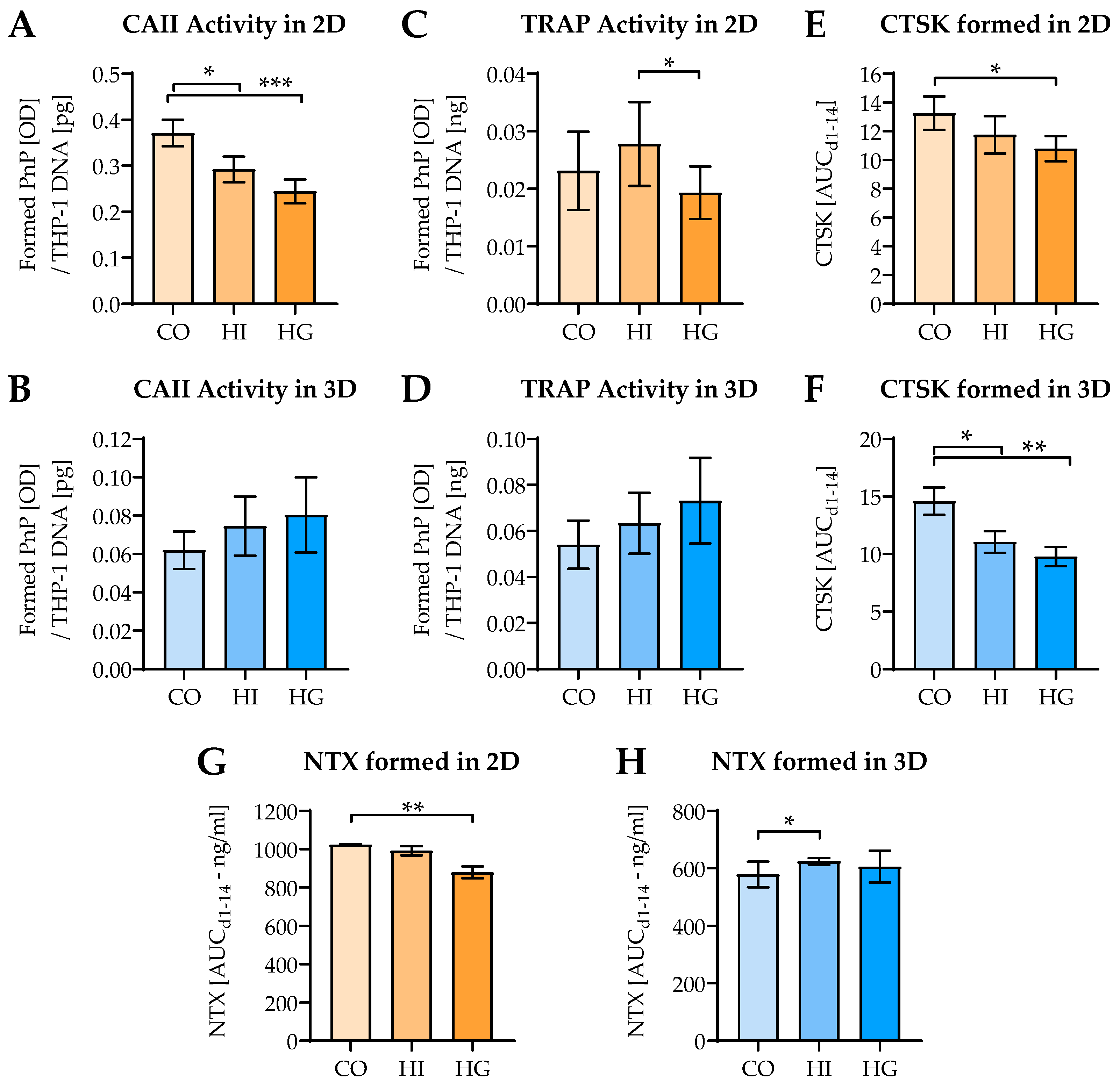
2.2. Change of Osteoblast and Osteoclast Function in 2D and 3D Coculture
2.3. Osteoclast Activity First Detectable on Day 4 of Culture
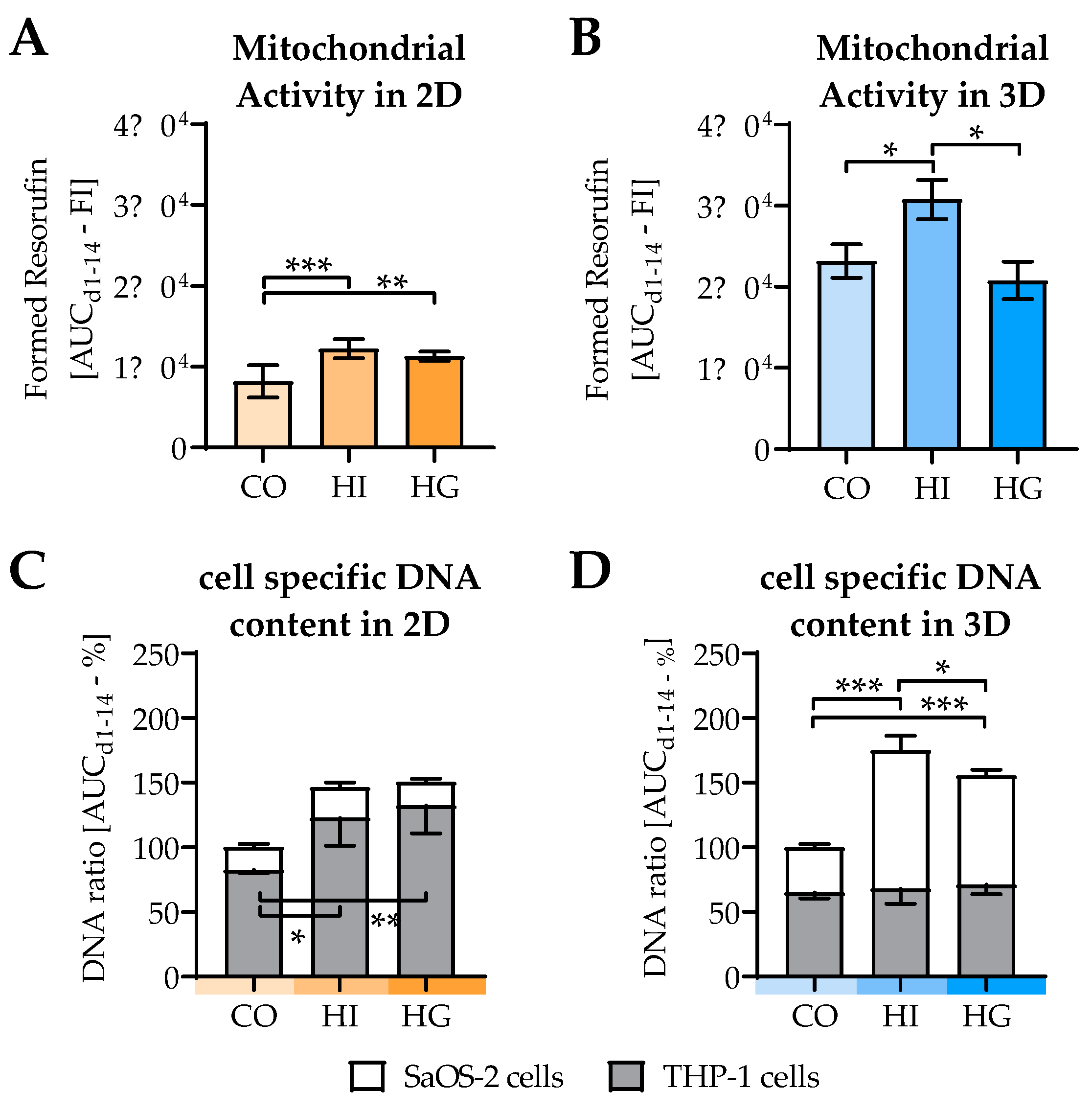
2.4. Insulin and Glucose Affect Cell Viability and Cell Composition in the Cocultures
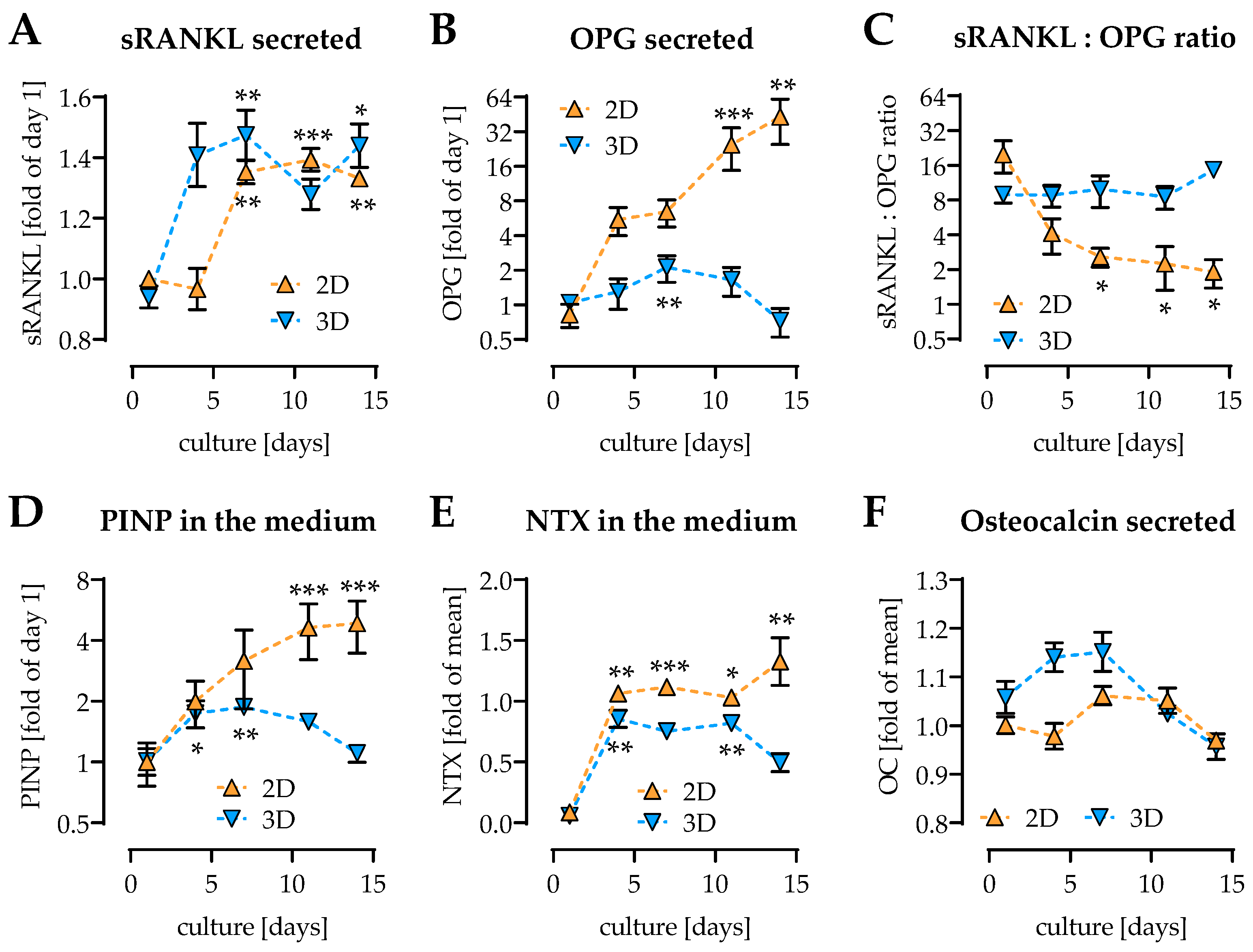
2.5. Pre-/diabetic Conditions Affect Function of Osteoblastic SaOS-2 Cells
2.6. Pre-/diabetic Conditions Affect Function of THP-1-Derived Osteoclastic Cells
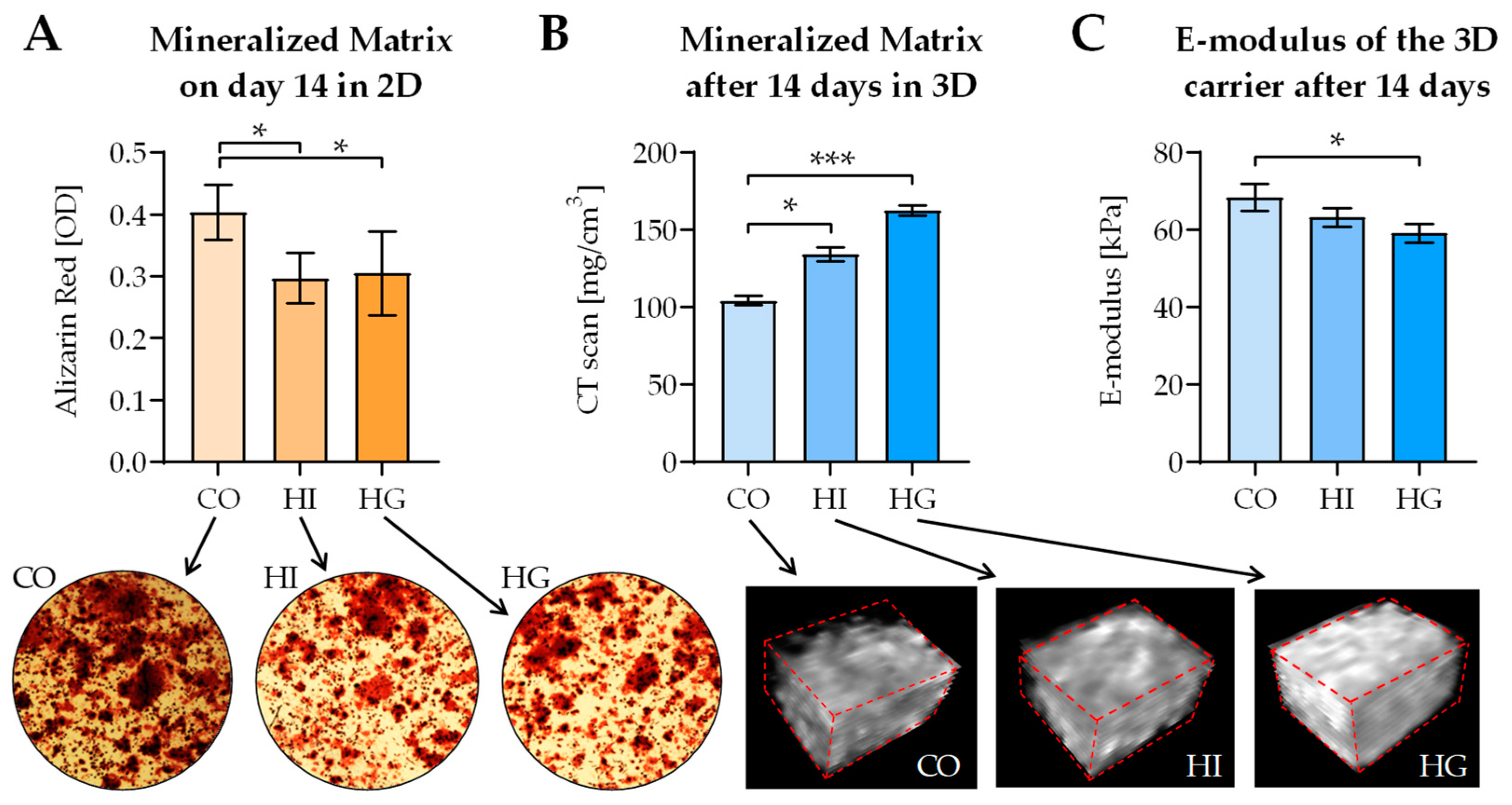
2.7. Pre-/diabetic Conditions Contrarily Affect Matrix Mineralization in 2D and 3D Coculture
3. Discussion
4. Materials and Methods
4.1. Scaffold Manufacturing and Sterilization
4.2. Cell Lines
4.3. 2D Coculture
4.4. 3D Coculture
4.5. In vitro Conditions to Simulate the Development of a Type 2 Diabetes Mellitus
- For normoglycemic control conditions (CO), 14 mM mannitol was added to the osteogenic medium (11 mM glucose) to equalize osmolality compared to hyperglycemic conditions.
- For the “prediabetic conditions” with high insulin (HI), CO medium was supplemented with 160 I.U./l insulin (Actrapid, NovoNordisk, Bagsværd, DNK).
- For the “diabetic conditions” with hyperglycemia (HG), medium was additionally supplemented with 14 mM glucose, to obtain a final glucose concentration of 25 mM.
4.6. Resazurin Conversion Assay
4.7. Isolation of Total DNA
4.8. Quantification of Cell-Specific DNA
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. Dot Blot Analysis
4.11. Alkaline Phosphatase (ALP) Activity
4.12. Tartrate-Resistant Acid Phosphatase (TRAP) Activity
4.13. Carbonic Anhydrase II (CAII) Activity
4.14. Formation of Mineralized Matrix
4.14.1. Alizarin Red Staining in 2D Cocultures
4.14.2. Mineral Content of the Scaffolds (3D Cocultures)
4.15. Scaffold Stiffness
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2D | two-dimensional |
| 3D | three-dimensional |
| ALP | alkaline phosphatase |
| APS | ammonium persulfate |
| BAAm | N,N-methylene(bis)acrylamide |
| BMD | bone mineral density |
| CAII | carbonic anhydrase II |
| DPP4 | dipeptidyl peptidase 4 |
| GLP-1 | glucagon-like peptide 1 |
| MSCs | mesenchymal stem cells |
| NTX | collagen-type I N-telopeptide |
| OPG | osteoprotegerin |
| pHEMA | poly-2-hydroxyethyl methacrylate |
| PINP | procollagen type I N-terminal propeptide |
| PRP | platelet-rich plasma |
| SGLT2 | sodium-glucose transport protein 2 |
| sRANKL | soluble receptor activator of nuclear factor kappa-Β ligand |
| TEMED | N,N,N,N-tetramethyl-ethylenediamine |
| TRAP5b | tartrate-resistant acidic phosphatase 5b |
References
- Vestergaard, P.; Rejnmark, L.; Mosekilde, L. Relative fracture risk in patients with diabetes mellitus, and the impact of insulin and oral antidiabetic medication on relative fracture risk. Diabetologia 2005, 48, 1292–1299. [Google Scholar] [CrossRef]
- Retzepi, M.; Donos, N. The effect of diabetes mellitus on osseous healing. Clin. Oral Implants Res. 2010, 21, 673–681. [Google Scholar] [CrossRef]
- Hernandez, R.K.; Do, T.P.; Critchlow, C.W.; Dent, R.E.; Jick, S.S. Patient-related risk factors for fracture-healing complications in the United Kingdom General Practice Research Database. Acta Orthop. 2012, 83, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, P. Discrepancies in bone mineral density and fracture risk in patients with type 1 and type 2 diabetes--a meta-analysis. Osteoporos. Int. 2007, 18, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Picke, A.K.; Campbell, G.; Napoli, N.; Hofbauer, L.C.; Rauner, M. Update on the impact of type 2 diabetes mellitus on bone metabolism and material properties. Endocrinol. Connect. 2019, 8, R55–R70. [Google Scholar] [CrossRef] [PubMed]
- Lauterbach, S.; Kostev, K.; Kohlmann, T. Prevalence of diabetic foot syndrome and its risk factors in the UK. J. Wound Care 2010, 19, 333–337. [Google Scholar] [CrossRef]
- Pscherer, S.; Dippel, F.W.; Lauterbach, S.; Kostev, K. Amputation rate and risk factors in type 2 patients with diabetic foot syndrome under real-life conditions in Germany. Prim Care Diabetes 2012, 6, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Kahm, K.; Laxy, M.; Schneider, U.; Rogowski, W.H.; Lhachimi, S.K.; Holle, R. Health care costs associated with incident complications in patients with type 2 diabetes in Germany. Diabetes Care 2018, 41, 971–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.B.; Groop, L. Precision medicine in type 2 diabetes. J. Intern Med. 2019, 285, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehnert, S.; Rinderknecht, H.; Aspera-Werz, R.H.; Haussling, V.; Nussler, A.K. Use of in vitro bone models to screen for altered bone metabolism, osteopathies, and fracture healing: Challenges of complex models. Arch. Toxicol. 2020, 94, 3937–3958. [Google Scholar] [CrossRef]
- Freude, T.; Braun, K.F.; Haug, A.; Pscherer, S.; Stöckle, U.; Nussler, A.K.; Ehnert, S. Hyperinsulinemia reduces osteoblast activity in vitro via upregulation of TGF-β. J. Mol. Med. 2012, 90, 1257–1266. [Google Scholar] [CrossRef]
- Ehnert, S.; Freude, T.; Ihle, C.; Mayer, L.; Braun, B.; Graeser, J.; Flesch, I.; Stockle, U.; Nussler, A.K.; Pscherer, S. Factors circulating in the blood of type 2 diabetes mellitus patients affect osteoblast maturation-description of a novel in vitro model. Exp. Cell Res. 2015, 332, 247–258. [Google Scholar] [CrossRef]
- Sreekumar, V.; Aspera-Werz, R.H.; Tendulkar, G.; Reumann, M.K.; Freude, T.; Breitkopf-Heinlein, K.; Dooley, S.; Pscherer, S.; Ochs, B.G.; Flesch, I.; et al. BMP9 a possible alternative drug for the recently withdrawn BMP7? New perspectives for (re-)implementation by personalized medicine. Arch. Toxicol. 2017, 91, 1353–1366. [Google Scholar] [CrossRef]
- Ehnert, S.; Linnemann, C.; Aspera-Werz, R.H.; Haussling, V.; Braun, B.; Weng, W.; Sheng, Z.; Ngamsri, K.C.; Nussler, A.K. Feasibility of cell lines for in vitro co-cultures models for bone metabolism. SciMed. J. 2020, 2, 157–181. [Google Scholar] [CrossRef]
- Griffith, L.G.; Swartz, M.A. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef]
- Zhao, W.; Li, X.; Liu, X.; Zhang, N.; Wen, X. Effects of substrate stiffness on adipogenic and osteogenic differentiation of human mesenchymal stem cells. Mater Sci. Eng. C Mater Biol. Appl. 2014, 40, 316–323. [Google Scholar] [CrossRef]
- Weng, W.; Haussling, V.; Aspera-Werz, R.H.; Springer, F.; Rinderknecht, H.; Braun, B.; Kuper, M.A.; Nussler, A.K.; Ehnert, S. Material-dependent formation and degradation of bone matrix-comparison of two cryogels. Bioengineering 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Haussling, V.; Deninger, S.; Vidoni, L.; Rinderknecht, H.; Ruoss, M.; Arnscheidt, C.; Athanasopulu, K.; Kemkemer, R.; Nussler, A.K.; Ehnert, S. Impact of four protein additives in cryogels on osteogenic differentiation of adipose-derived mesenchymal stem cells. Bioengineering 2019, 6, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehnert, S.; Heuberger, E.; Linnemann, C.; Nussler, A.K.; Pscherer, S. TGF-β1-Dependent downregulation of HDAC9 inhibits maturation of human osteoblasts. J. Funct. Morphol. Kinesiol. 2017, 2, 41. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhou, G.; Li, M.; Li, Y.; Yang, L.; Fu, Q.; Tian, Y. High glucose downregulates connexin 43 expression and its gap junction and hemichannel function in osteocyte-like MLO-Y4 cells through activation of the p38MAPK/ERK signal pathway. Diabetes Metab. Syndr. Obes. 2020, 13, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Ma, C.; Zhang, M. High glucose inhibits osteogenic differentiation and proliferation of MC3T3E1 cells by regulating P2X7. Mol. Med. Rep. 2019, 20, 5084–5090. [Google Scholar]
- Kopesky, P.; Tiedemann, K.; Alkekhia, D.; Zechner, C.; Millard, B.; Schoeberl, B.; Komarova, S.V. Autocrine signaling is a key regulatory element during osteoclastogenesis. Biol. Open 2014, 3, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinker, T.E.; Hammoudi, T.M.; Kemp, M.L.; Lu, H.; Temenoff, J.S. Interactions between mesenchymal stem cells, adipocytes, and osteoblasts in a 3D tri-culture model of hyperglycemic conditions in the bone marrow microenvironment. Integr. Biol. 2014, 6, 324–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalfamo, D.L.; Britten, T.M.; Storch, D.L.; Calderon, N.L.; Sorenson, H.L.; Wallet, S.M. Hyperglycemia induced and intrinsic alterations in type 2 diabetes-derived osteoclast function. Oral Dis. 2013, 19, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hygum, K.; Starup-Linde, J.; Harslof, T.; Vestergaard, P.; Langdahl, B.L. Mechanisms in endocrinology: Diabetes mellitus, a state of low bone turnover-a systematic review and meta-analysis. Eur. J. Endocrinol. 2017, 176, R137–R157. [Google Scholar] [CrossRef]
- Sassi, F.; Buondonno, I.; Luppi, C.; Spertino, E.; Stratta, E.; Di Stefano, M.; Ravazzoli, M.; Isaia, G.; Trento, M.; Passera, P.; et al. Type 2 diabetes affects bone cells precursors and bone turnover. BMC Endocrinol. Dis. 2018, 18, 55–66. [Google Scholar] [CrossRef]
- Cunha, J.S.; Ferreira, V.M.; Maquigussa, E.; Naves, M.A.; Boim, M.A. Effects of high glucose and high insulin concentrations on osteoblast function in vitro. Cell Tissue Res. 2014, 358, 249–256. [Google Scholar] [CrossRef]
- Xu, J.; Yue, F.; Wang, J.; Chen, L.; Qi, W. High glucose inhibits receptor activator of nuclear factorkappaB ligand-induced osteoclast differentiation via downregulation of vATPase V0 subunit d2 and dendritic cellspecific transmembrane protein. Mol. Med. Rep. 2015, 11, 865–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotti, T.N.; O’Sullivan, R.P.; Shen, Z.; Flannery, M.R.; Fajardo, R.J.; Ross, F.P.; Goldring, S.R.; McHugh, K.P. Bone matrix regulates osteoclast differentiation and annexin A8 gene expression. J. Cell Physiol. 2011, 226, 3413–3421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, S.E.; Shao, J.; van den Beucken, J. Combinatorial surface roughness effects on osteoclastogenesis and osteogenesis. ACS Appl Mater. Interfaces 2018, 10, 36652–36663. [Google Scholar] [CrossRef] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Dong, C.; Yang, L.; Lv, Y. 3D scaffolds with different stiffness but the same microstructure for bone tissue engineering. ACS Appl Mater. Interfaces 2015, 7, 15790–15802. [Google Scholar] [CrossRef] [PubMed]
- Shapira, L.; Halabi, A. Behavior of two osteoblast-like cell lines cultured on machined or rough titanium surfaces. Clin. Oral Implants Res. 2009, 20, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Pandit, V.; Zuidema, J.M.; Venuto, K.N.; Macione, J.; Dai, G.; Gilbert, R.J.; Kotha, S.P. Evaluation of multifunctional polysaccharide hydrogels with varying stiffness for bone tissue engineering. Tissue Eng. Part A 2013, 19, 2452–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostenuik, P.J.; Shalhoub, V. Osteoprotegerin: A physiological and pharmacological inhibitor of bone resorption. Curr. Pharm. Des. 2001, 7, 613–635. [Google Scholar] [CrossRef]
- Lotz, E.M.; Berger, M.B.; Boyan, B.D.; Schwartz, Z. Regulation of mesenchymal stem cell differentiation on microstructured titanium surfaces by semaphorin 3A. Bone 2020, 134, 115260. [Google Scholar] [CrossRef]
- Wittrant, Y.; Gorin, Y.; Woodruff, K.; Horn, D.; Abboud, H.E.; Mohan, S.; Abboud-Werner, S.L. High d(+)glucose concentration inhibits RANKL-induced osteoclastogenesis. Bone 2008, 42, 1122–1130. [Google Scholar] [CrossRef] [Green Version]
- Rucci, N.; Teti, A. The “love-hate” relationship between osteoclasts and bone matrix. Matrix Biol. 2016, 52–54, 176–190. [Google Scholar] [CrossRef]
- Costantini, S.; Conte, C. Bone health in diabetes and prediabetes. World J. Diabetes 2019, 10, 421–445. [Google Scholar] [CrossRef] [PubMed]
- Bocker, W.; Yin, Z.; Drosse, I.; Haasters, F.; Rossmann, O.; Wierer, M.; Popov, C.; Locher, M.; Mutschler, W.; Docheva, D.; et al. Introducing a single-cell-derived human mesenchymal stem cell line expressing hTERT after lentiviral gene transfer. J. Cell Mol. Med. 2008, 12, 1347–1359. [Google Scholar] [CrossRef] [Green Version]
- Ruoss, M.; Kieber, V.; Rebholz, S.; Linnemann, C.; Rinderknecht, H.; Haussling, V.; Hacker, M.; Olde Damink, L.H.H.; Ehnert, S.; Nussler, A.K. Cell-type-specific quantification of a scaffold-based 3D liver co-culture. Methods Protoc. 2019, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastelic, V.; Budowle, B.; Drobnic, K. Validation of SRY marker for forensic casework analysis. J. Forensic Sci. 2009, 54, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Koperski, K.; Schumacher, M.; Gelinsky, M. Relevance of osteoclast-specific enzyme activities in cell-based in vitro resorption assays. Eur. Cell Mater. 2017, 33, 28–42. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Protein | Role | Order # | Company |
|---|---|---|---|
| NTX (collagen-type I N-telopeptide) | Collagen degradation | E-EL-H0836 | Elabscience, Houston, TX, USA |
| OPG (osteoprotegerin) | Inhibitor for sRANKL | ABIN411341 | Antibodies-online, Aachen, Germany |
| PINP (procollagen type I N-terminal propeptide) | Collagen formation | 8003 | TecoMedical, Neufahrn, Germany |
| sRANKL (soluble receptor activator of nuclear factor kappa-Β ligand | Inducer for osteoclastogenesis | 900-K142 | Peprotech, Hamburg, Germany |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Häussling, V.; Aspera-Werz, R.H.; Rinderknecht, H.; Springer, F.; Arnscheidt, C.; Menger, M.M.; Histing, T.; Nussler, A.K.; Ehnert, S. 3D Environment Is Required In Vitro to Demonstrate Altered Bone Metabolism Characteristic for Type 2 Diabetics. Int. J. Mol. Sci. 2021, 22, 2925. https://doi.org/10.3390/ijms22062925
Häussling V, Aspera-Werz RH, Rinderknecht H, Springer F, Arnscheidt C, Menger MM, Histing T, Nussler AK, Ehnert S. 3D Environment Is Required In Vitro to Demonstrate Altered Bone Metabolism Characteristic for Type 2 Diabetics. International Journal of Molecular Sciences. 2021; 22(6):2925. https://doi.org/10.3390/ijms22062925
Chicago/Turabian StyleHäussling, Victor, Romina H. Aspera-Werz, Helen Rinderknecht, Fabian Springer, Christian Arnscheidt, Maximilian M. Menger, Tina Histing, Andreas K. Nussler, and Sabrina Ehnert. 2021. "3D Environment Is Required In Vitro to Demonstrate Altered Bone Metabolism Characteristic for Type 2 Diabetics" International Journal of Molecular Sciences 22, no. 6: 2925. https://doi.org/10.3390/ijms22062925
APA StyleHäussling, V., Aspera-Werz, R. H., Rinderknecht, H., Springer, F., Arnscheidt, C., Menger, M. M., Histing, T., Nussler, A. K., & Ehnert, S. (2021). 3D Environment Is Required In Vitro to Demonstrate Altered Bone Metabolism Characteristic for Type 2 Diabetics. International Journal of Molecular Sciences, 22(6), 2925. https://doi.org/10.3390/ijms22062925