
The Importance of Therapeutically Targeting the Binary Toxin from Clostridioides difficile

 and
and 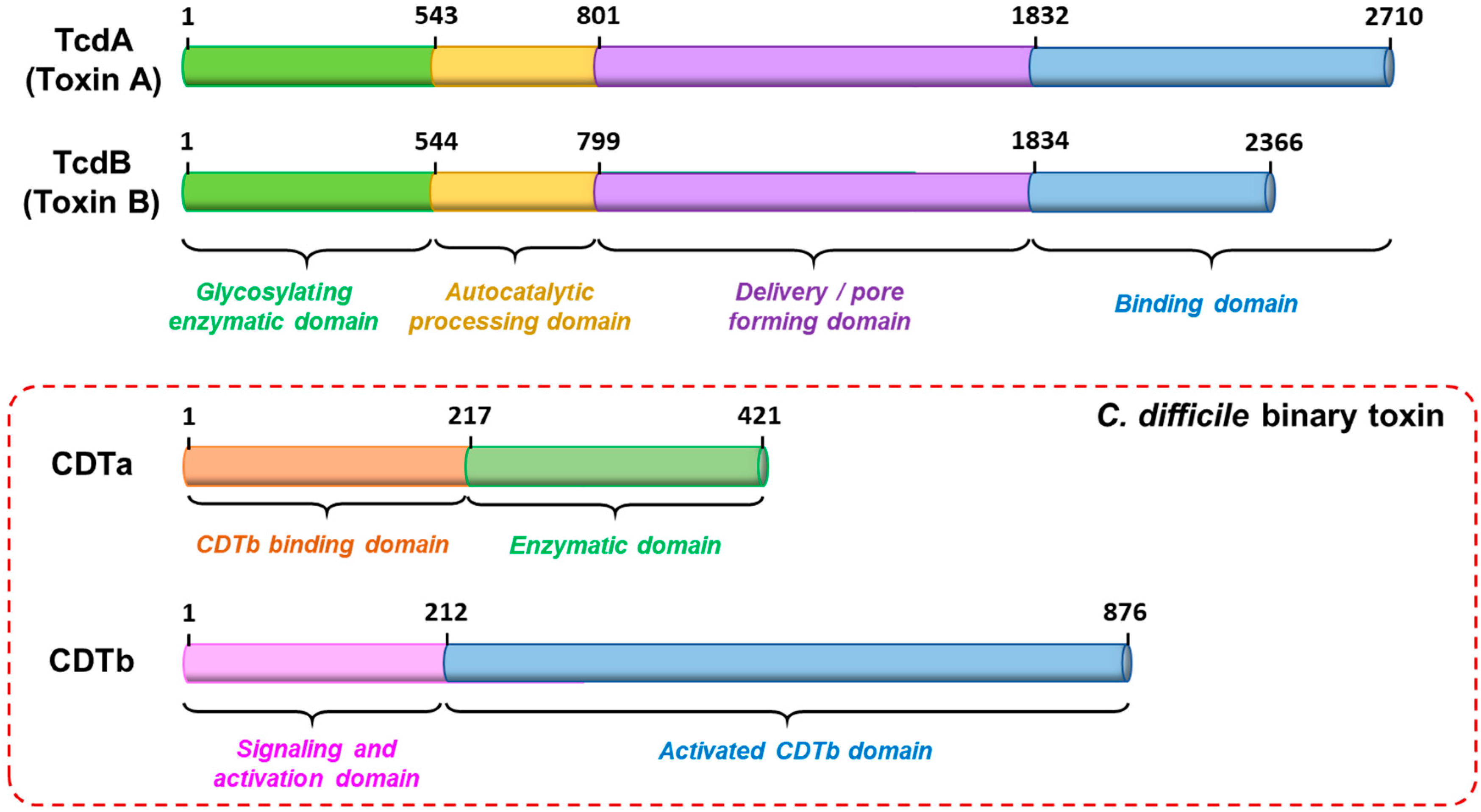{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. CDT Epidemiology
3. CDT Structure and Mechanism of Action
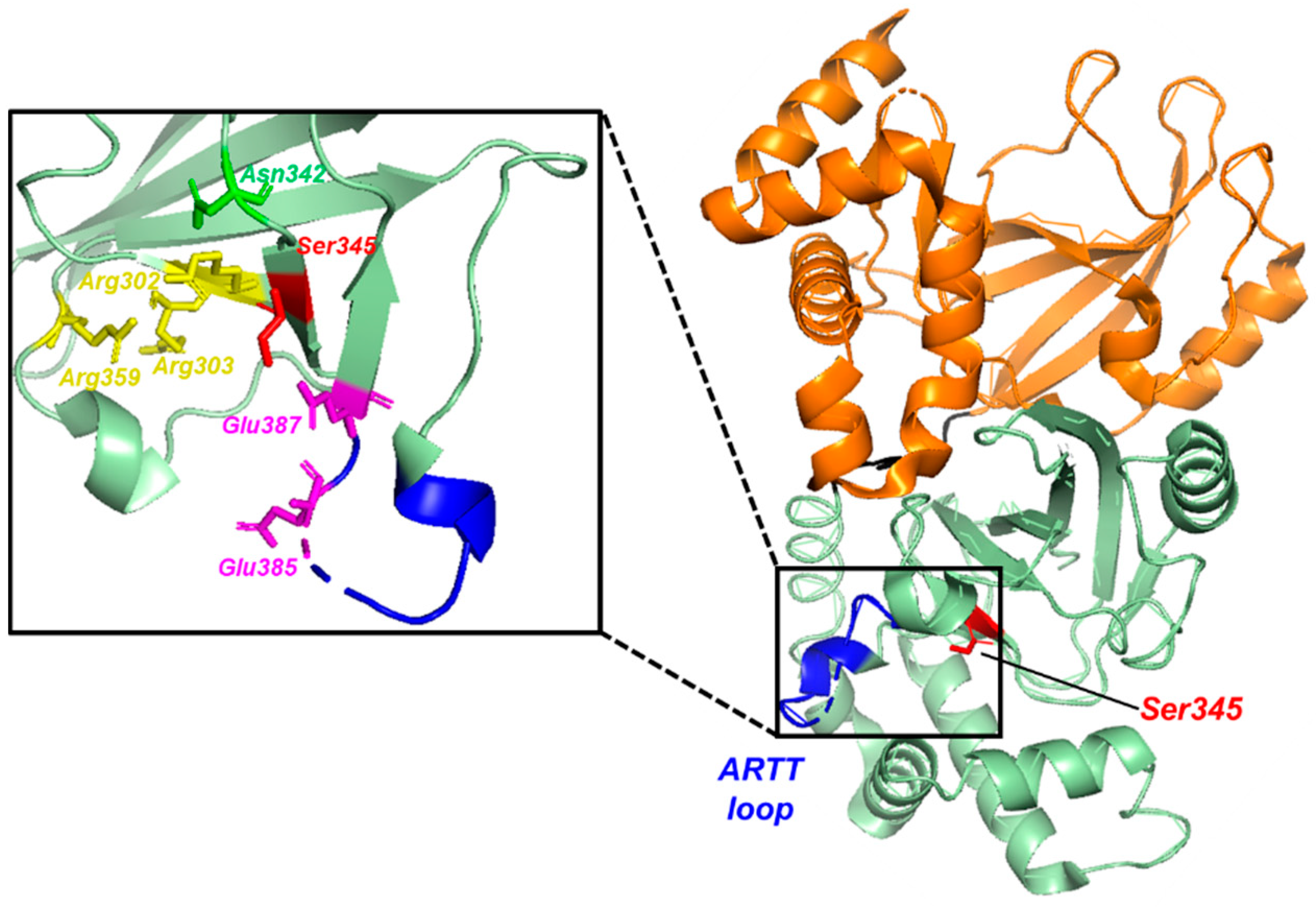
3.1. Structure of the CDTa Enzymatic Subunit
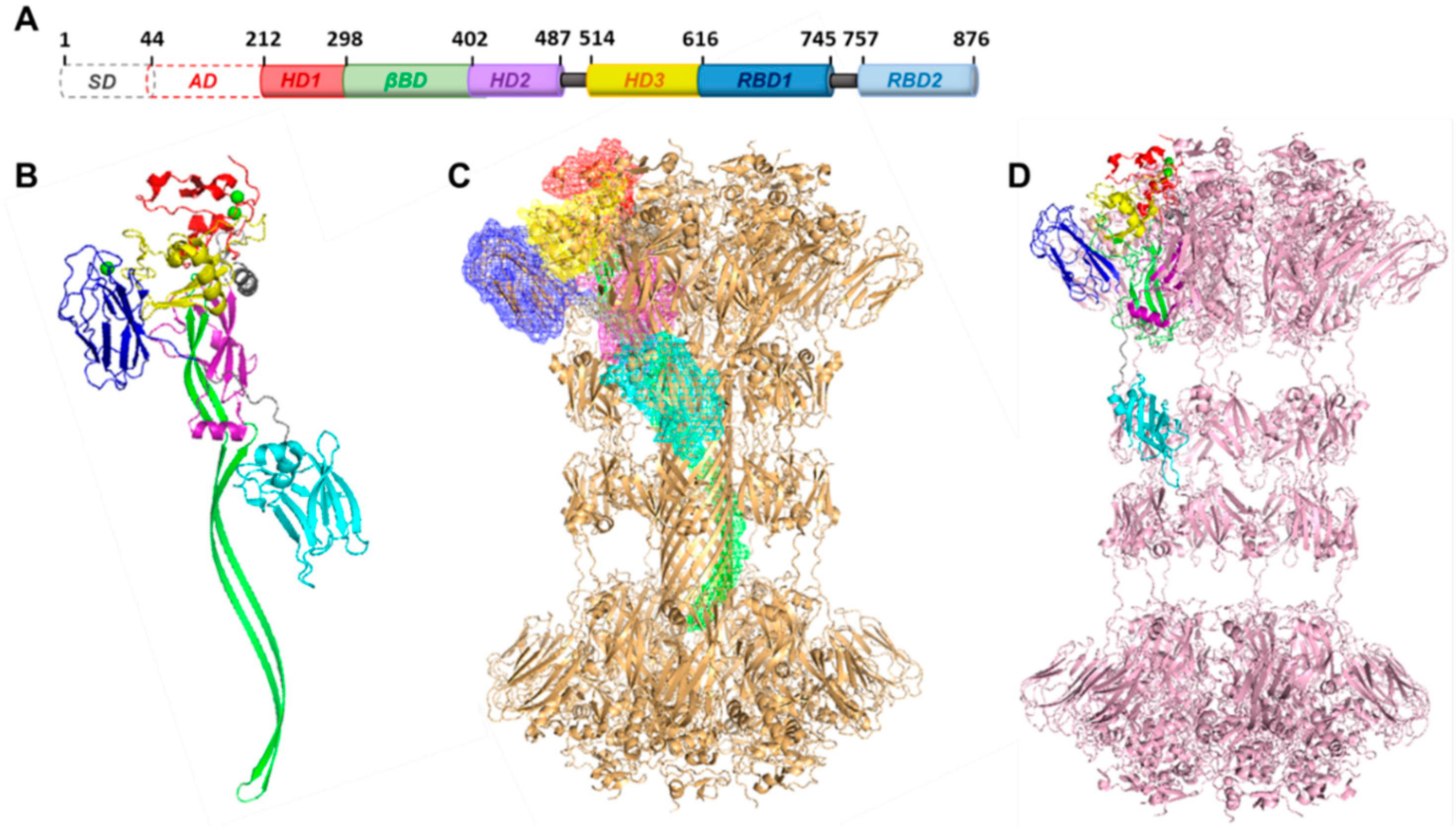
3.2. Structure of the Binding Domain
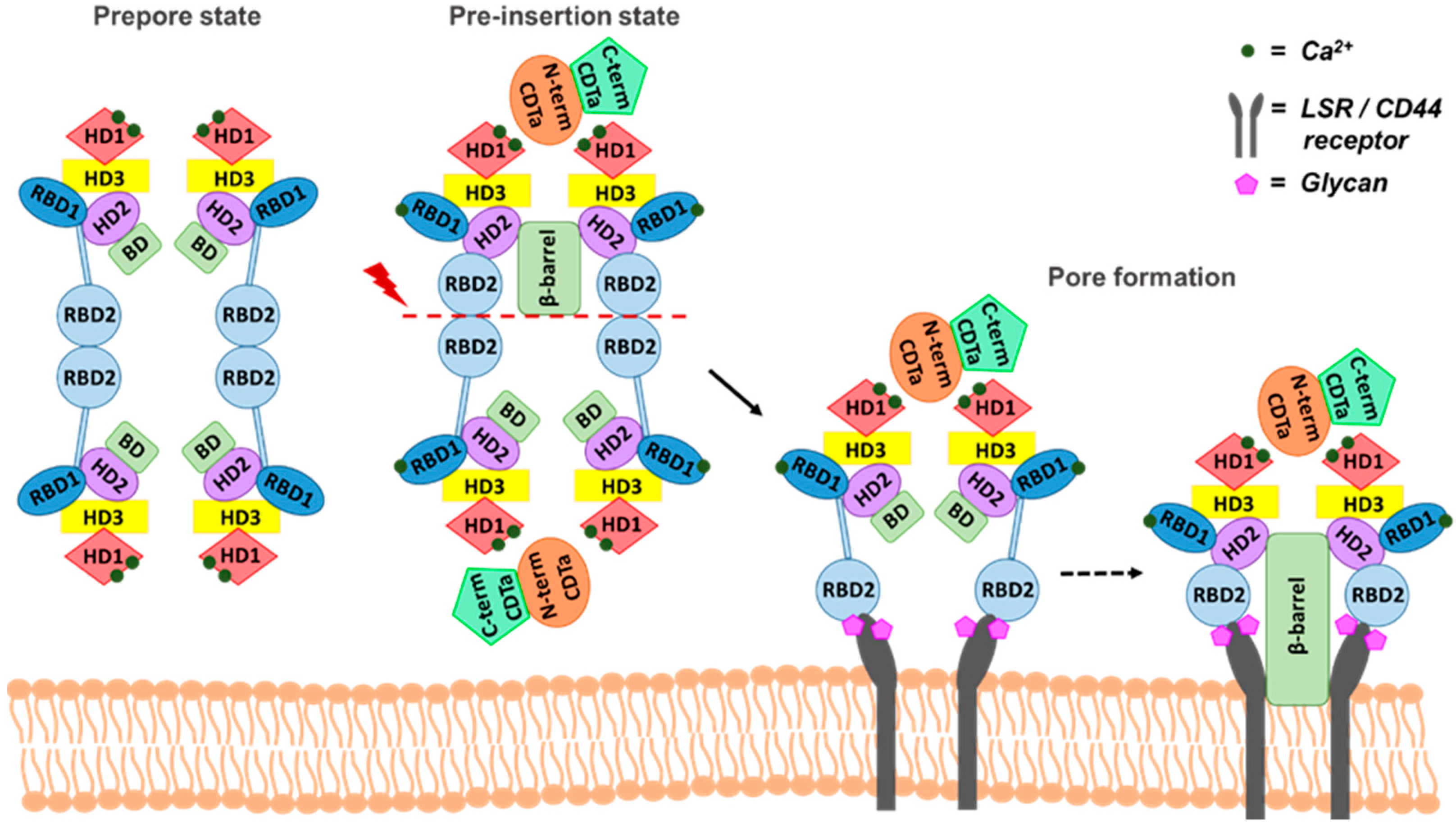
4. Receptor Interaction and a Plausible Molecular Mechanism
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- Davies, A.H.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. Super toxins from a super bug: Structure and function of Clostridium difficile toxins. Biochem. J. 2011, 436, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popoff, M.R. “Bacterial Toxins” Section in the Journal Toxins: A Fantastic Multidisciplinary Interplay between Bacterial Pathogenicity Mechanisms, Physiological Processes, Genomic Evolution, and Subsequent Development of Identification Methods, Efficient Treatment, and Prevention of Toxigenic Bacteria. Toxins 2018, 10, 44. [Google Scholar]
- Di Bella, S.; Ascenzi, P.; Siarakas, S.; Petrosillo, N.; di Masi, A. Clostridium difficile Toxins A and B: Insights into Pathogenic Properties and Extraintestinal Effects. Toxins 2016, 8, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekaran, R.; Lacy, D.B. The role of toxins in Clostridium difficile infection. FEMS Microbiol. Rev. 2017, 41, 723–750. [Google Scholar] [CrossRef] [Green Version]
- Aktories, K.; Papatheodorou, P.; Schwan, C. Binary Clostridium difficile toxin (CDT)—A virulence factor disturbing the cytoskeleton. Anaerobe 2018, 53, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R.; Rubin, E.J.; Gill, D.M.; Boquet, P. Actin-specific ADP-ribosyltransferase produced by a Clostridium difficile strain. Infect. Immun. 1988, 56, 2299–2306. [Google Scholar] [CrossRef] [Green Version]
- Perelle, S.; Gibert, M.; Bourlioux, P.; Corthier, G.; Popoff, M.R. Production of a complete binary toxin (actin-specific ADP-ribosyltransferase) by Clostridium difficile CD196. Infect. Immun. 1997, 65, 1402–1407. [Google Scholar] [CrossRef] [Green Version]
- Rupnik, M.; Janezic, S. An Update on Clostridium difficile Toxinotyping. J. Clin. Microbiol. 2016, 54, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupnik, M. Heterogeneity of large clostridial toxins: Importance of Clostridium difficile toxinotypes. FEMS Microbiol. Rev. 2008, 32, 541–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbut, F.; Decre, D.; Lalande, V.; Burghoffer, B.; Noussair, L.; Gigandon, A.; Espinasse, F.; Raskine, L.; Robert, J.; Mangeol, A.; et al. Clinical features of Clostridium difficile-associated diarrhoea due to binary toxin (actin-specific ADP-ribosyltransferase)-producing strains. J. Med. Microbiol. 2005, 54, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Eckert, C.; Emirian, A.; Le Monnier, A.; Cathala, L.; De Montclos, H.; Goret, J.; Berger, P.; Petit, A.; De Chevigny, A.; Jean-Pierre, H.; et al. Prevalence and pathogenicity of binary toxin-positive Clostridium difficile strains that do not produce toxins A and B. New Microbes New Infect. 2015, 3, 12–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, L.C.; Killgore, G.E.; Thompson, A.; Owens, R.C., Jr.; Kazakova, S.V.; Sambol, S.P.; Johnson, S.; Gerding, D.N. An epidemic, toxin gene-variant strain of Clostridium difficile. N. Engl. J. Med. 2005, 353, 2433–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popoff, M.R.; Boquet, P. Clostridium spiroforme toxin is a binary toxin which ADP-ribosylates cellular actin. Biochem. Biophys. Res. Commun. 1988, 152, 1361–1368. [Google Scholar] [CrossRef]
- Barth, H.; Aktories, K.; Popoff, M.R.; Stiles, B.G. Binary bacterial toxins: Biochemistry, biology, and applications of common Clostridium and Bacillus proteins. Microbiol. Mol. Biol. Rev. 2004, 68, 373–402. [Google Scholar] [CrossRef] [Green Version]
- Stiles, B.G.; Pradhan, K.; Fleming, J.M.; Samy, R.P.; Barth, H.; Popoff, M.R. Clostridium and bacillus binary enterotoxins: Bad for the bowels, and eukaryotic being. Toxins 2014, 6, 2626–2656. [Google Scholar] [CrossRef] [Green Version]
- Gulke, I.; Pfeifer, G.; Liese, J.; Fritz, M.; Hofmann, F.; Aktories, K.; Barth, H. Characterization of the enzymatic component of the ADP-ribosyltransferase toxin CDTa from Clostridium difficile. Infect. Immun. 2001, 69, 6004–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, N.C.; Aktories, K.; Barbieri, J.T. Novel bacterial ADP-ribosylating toxins: Structure and function. Nat. Rev. Microbiol. 2014, 12, 599–611. [Google Scholar] [CrossRef] [Green Version]
- Knapp, O.; Benz, R.; Popoff, M.R. Pore-forming activity of clostridial binary toxins. Biochim. Biophys. Acta 2016, 1858, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Godoy-Ruiz, R.; Adipietro, K.A.; Peralta, C.; Ben-Hail, D.; Varney, K.M.; Cook, M.E.; Roth, B.M.; Wilder, P.T.; Cleveland, T.; et al. Structure of the cell-binding component of the Clostridium difficile binary toxin reveals a di-heptamer macromolecular assembly. Proc. Natl. Acad. Sci. USA 2020, 117, 1049–1058. [Google Scholar] [CrossRef] [Green Version]
- Gerding, D.N.; Johnson, S.; Rupnik, M.; Aktories, K. Clostridium difficile binary toxin CDT: Mechanism, epidemiology, and potential clinical importance. Gut Microbes 2014, 5, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Finkelstein, A.; Collier, R.J. Evidence that translocation of anthrax toxin’s lethal factor is initiated by entry of its N terminus into the protective antigen channel. Proc. Natl. Acad. Sci. USA 2004, 101, 16756–16761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, T.; Yoshida, T.; Kawamoto, A.; Mitsuoka, K.; Iwasaki, K.; Tsuge, H. Cryo-EM structures reveal translocational unfolding in the clostridial binary iota toxin complex. Nat. Struct. Mol. Biol. 2020, 27, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Sheedlo, M.J.; Anderson, D.M.; Thomas, A.K.; Lacy, D.B. Structural elucidation of the Clostridioides difficile transferase toxin reveals a single-site binding mode for the enzyme. Proc. Natl. Acad. Sci. USA 2020, 117, 6139–6144. [Google Scholar] [CrossRef]
- Sundriyal, A.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. Structural basis for substrate recognition in the enzymatic component of ADP-ribosyltransferase toxin CDTa from Clostridium difficile. J. Biol. Chem. 2009, 284, 28713–28719. [Google Scholar] [CrossRef] [Green Version]
- Roth, B.M.; Varney, K.M.; Rustandi, R.R.; Weber, D.J. (1)H(N), (13)C, and (15)N resonance assignments of the CDTb-interacting domain (CDTaBID) from the Clostridium difficile binary toxin catalytic component (CDTa, residues 1-221). Biomol. NMR Assign. 2016, 10, 335–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, L.A.; Tatge, H.; Schneider, C.; Ruschig, M.; Hust, M.; Barton, J.; Thiemann, S.; Fühner, V.; Russo, G.; Gerhard, R. The Binary Toxin CDT of. Toxins 2018, 10, 225. [Google Scholar] [CrossRef] [Green Version]
- van Damme, J.; Jung, M.; Hofmann, F.; Just, I.; Vandekerckhove, J.; Aktories, K. Analysis of the catalytic site of the actin ADP-ribosylating Clostridium perfringens iota toxin. FEBS Lett. 1996, 380, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, H.; Nagahama, M.; Oda, M.; Iwamoto, S.; Utsunomiya, H.; Marquez, V.E.; Katunuma, N.; Nishizawa, M.; Sakurai, J. Structural basis of actin recognition and arginine ADP-ribosylation by Clostridium perfringens iota-toxin. Proc. Natl. Acad. Sci. USA 2008, 105, 7399–7404. [Google Scholar] [CrossRef] [Green Version]
- Holbourn, K.P.; Shone, C.C.; Acharya, K.R. A family of killer toxins. Exploring the mechanism of ADP-ribosylating toxins. FEBS J. 2006, 273, 4579–4593. [Google Scholar] [CrossRef]
- Roth, B.M.; Godoy-Ruiz, R.; Varney, K.M.; Rustandi, R.R.; Weber, D.J. 1H, 13C, and 15N resonance assignments of an enzymatically active domain from the catalytic component (CDTa, residues 216-420) of a binary toxin from Clostridium difficile. Biomol. NMR Assign. 2016, 10, 213–217. [Google Scholar] [CrossRef]
- Davies, A.H.; McGlashan, J.; Posner, M.G.; Roberts, A.K.; Shone, C.C.; Acharya, K.R. Functional significance of active site residues in the enzymatic component of the Clostridium difficile binary toxin. Biochem. Biophys. Rep. 2016, 8, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustandi, R.R.; Hamm, M. Development of an ADP-ribosylation assay for residual toxicity in C. difficile binary toxin CDTa using automated capillary western blot. J. Pharm. Biomed. Anal. 2020, 182, 113125. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Rustandi, R.R.; Lancaster, C.; Hong, L.G.; Thiriot, D.S.; Xie, J.; Secore, S.; Kristopeit, A.; Wang, S.C.; Heinrichs, J.H. Toxicity assessment of Clostridium difficile toxins in rodent models and protection of vaccination. Vaccine 2016, 34, 1319–1323. [Google Scholar] [CrossRef] [PubMed]
- Secore, S.; Wang, S.; Doughtry, J.; Xie, J.; Miezeiewski, M.; Rustandi, R.R.; Horton, M.; Xoconostle, R.; Wang, B.; Lancaster, C.; et al. Development of a Novel Vaccine Containing Binary Toxin for the Prevention of Clostridium difficile Disease with Enhanced Efficacy against NAP1 Strains. PLoS ONE 2017, 12, e0170640. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Horton, M.; Zorman, J.; Antonello, J.M.; Zhang, Y.; Arnold, B.A.; Secore, S.; Xoconostle, R.; Miezeiewski, M.; Wang, S.; et al. Development and optimization of a high-throughput assay to measure neutralizing antibodies against Clostridium difficile binary toxin. Clin. Vaccine Immunol. 2014, 21, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Sheedlo, M.J.; Jensen, J.L.; Lacy, D.B. Structural insights into the transition of Clostridioides difficile binary toxin from prepore to pore. Nat. Microbiol. 2020, 5, 102–107. [Google Scholar] [CrossRef]
- Jiang, J.; Pentelute, B.L.; Collier, R.J.; Zhou, Z.H. Atomic structure of anthrax protective antigen pore elucidates toxin translocation. Nature 2015, 521, 545–549. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Chandra, H.; Gaur, R.; Kurupati, R.K.; Chowdhury, S.; Tandon, V.; Singh, Y.; Maithal, K. Conformational fluctuations in anthrax protective antigen: A possible role of calcium in the folding pathway of the protein. FEBS Lett. 2003, 554, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Nagahama, M.; Ohkubo, N.; Kojima, T.; Shirai, H.; Iwamoto, S.; Oda, M.; Sakurai, J. Role of Ca2+-binding motif in cytotoxicity induced by Clostridium perfringens iota-toxin. Microb. Pathog. 2008, 44, 265–270. [Google Scholar] [CrossRef]
- Gao-Sheridan, S.; Zhang, S.; Collier, R.J. Exchange characteristics of calcium ions bound to anthrax protective antigen. Biochem. Biophys. Res. Commun. 2003, 300, 61–64. [Google Scholar] [CrossRef]
- Storm, L.; Bikker, F.J.; Nazmi, K.; Hulst, A.G.; der Riet-Van Oeveren, D.V.; Veerman, E.C.I.; Hays, J.P.; Kaman, W.E. Anthrax protective antigen is a calcium-dependent serine protease. Virulence 2018, 9, 1085–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatnagar, R.; Singh, Y.; Leppla, S.H.; Friedlander, A.M. Calcium is required for the expression of anthrax lethal toxin activity in the macrophagelike cell line J774A.1. Infect. Immun. 1989, 57, 2107–2114. [Google Scholar] [CrossRef] [Green Version]
- Sturino, J.M.; Pokusaeva, K.; Carpenter, R. Effective Sequestration of Clostridium difficile Protein Toxins by Calcium Aluminosilicate. Antimicrob. Agents Chemother. 2015, 59, 7178–7183. [Google Scholar] [CrossRef] [Green Version]
- Eckhardt, M.; Barth, H.; Blocker, D.; Aktories, K. Binding of Clostridium botulinum C2 toxin to asparagine-linked complex and hybrid carbohydrates. J. Biol. Chem. 2000, 275, 2328–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boraston, A.B.; Bolam, D.N.; Gilbert, H.J.; Davies, G.J. Carbohydrate-binding modules: Fine-tuning polysaccharide recognition. Biochem J. 2004, 382, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fisher, G.J. Receptor type protein tyrosine phosphatases (RPTPs)—roles in signal transduction and human disease. J. Cell Commun. Signal. 2012, 6, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktories, K.; Schwan, C.; Jank, T. Clostridium difficile Toxin Biology. Annu. Rev. Microbiol. 2017, 71, 281–307. [Google Scholar] [CrossRef]
- Mesli, S.; Javorschi, S.; Berard, A.M.; Landry, M.; Priddle, H.; Kivlichan, D.; Smith, A.J.; Yen, F.T.; Bihain, B.E.; Darmon, M. Distribution of the lipolysis stimulated receptor in adult and embryonic murine tissues and lethality of LSR-/- embryos at 12.5 to 14.5 days of gestation. Eur. J. Biochem. 2004, 271, 3103–3114. [Google Scholar] [CrossRef]
- Hemmasi, S.; Czulkies, B.A.; Schorch, B.; Veit, A.; Aktories, K.; Papatheodorou, P. Interaction of the Clostridium difficile Binary Toxin CDT and Its Host Cell Receptor, Lipolysis-stimulated Lipoprotein Receptor (LSR). J. Biol. Chem. 2015, 290, 14031–14044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papatheodorou, P.; Carette, J.E.; Bell, G.W.; Schwan, C.; Guttenberg, G.; Brummelkamp, T.R.; Aktories, K. Lipolysis-stimulated lipoprotein receptor (LSR) is the host receptor for the binary toxin Clostridium difficile transferase (CDT). Proc. Natl. Acad. Sci. USA 2011, 108, 16422–16427. [Google Scholar] [CrossRef] [Green Version]
- Papatheodorou, P.; Hornuss, D.; Nolke, T.; Hemmasi, S.; Castonguay, J.; Picchianti, M.; Aktories, K. Clostridium difficile binary toxin CDT induces clustering of the lipolysis-stimulated lipoprotein receptor into lipid rafts. MBio 2013, 4, e00244-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, M.L.; Marvaud, J.C.; Popoff, M.R.; Stiles, B.G. Detergent-resistant membrane microdomains facilitate Ib oligomer formation and biological activity of Clostridium perfringens iota-toxin. Infect. Immun. 2004, 72, 2186–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahama, M.; Yamaguchi, A.; Hagiyama, T.; Ohkubo, N.; Kobayashi, K.; Sakurai, J. Binding and internalization of Clostridium perfringens iota-toxin in lipid rafts. Infect. Immun. 2004, 72, 3267–3275. [Google Scholar] [CrossRef] [Green Version]
- Oliferenko, S.; Paiha, K.; Harder, T.; Gerke, V.; Schwarzler, C.; Schwarz, H.; Beug, H.; Gunthert, U.; Huber, L.A. Analysis of CD44-containing lipid rafts: Recruitment of annexin II and stabilization by the actin cytoskeleton. J. Cell Biol. 1999, 146, 843–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blonder, J.; Hale, M.L.; Chan, K.C.; Yu, L.R.; Lucas, D.A.; Conrads, T.P.; Zhou, M.; Popoff, M.R.; Issaq, H.J.; Stiles, B.G.; et al. Quantitative profiling of the detergent-resistant membrane proteome of iota-b toxin induced vero cells. J. Proteome Res. 2005, 4, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Wigelsworth, D.J.; Ruthel, G.; Schnell, L.; Herrlich, P.; Blonder, J.; Veenstra, T.D.; Carman, R.J.; Wilkins, T.D.; Van Nhieu, G.T.; Pauillac, S.; et al. CD44 Promotes intoxication by the clostridial iota-family toxins. PLoS ONE 2012, 7, e51356. [Google Scholar] [CrossRef]
- Gibert, M.; Marvaud, J.C.; Pereira, Y.; Hale, M.L.; Stiles, B.G.; Boquet, P.; Lamaze, C.; Popoff, M.R. Differential requirement for the translocation of clostridial binary toxins: Iota toxin requires a membrane potential gradient. FEBS Lett. 2007, 581, 1287–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, A. Clostridium difficile toxins: Mediators of inflammation. J. Innate Immun. 2012, 4, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, C.; Stecher, B.; Tzivelekidis, T.; van Ham, M.; Rohde, M.; Hardt, W.D.; Wehland, J.; Aktories, K. Clostridium difficile toxin CDT induces formation of microtubule-based protrusions and increases adherence of bacteria. PLoS Pathog. 2009, 5, e1000626. [Google Scholar] [CrossRef] [Green Version]
- Schwan, C.; Nolke, T.; Kruppke, A.S.; Schubert, D.M.; Lang, A.E.; Aktories, K. Cholesterol- and sphingolipid-rich microdomains are essential for microtubule-based membrane protrusions induced by Clostridium difficile transferase (CDT). J. Biol. Chem. 2011, 286, 29356–29365. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abeyawardhane, D.L.; Godoy-Ruiz, R.; Adipietro, K.A.; Varney, K.M.; Rustandi, R.R.; Pozharski, E.; Weber, D.J. The Importance of Therapeutically Targeting the Binary Toxin from Clostridioides difficile. Int. J. Mol. Sci. 2021, 22, 2926. https://doi.org/10.3390/ijms22062926
Abeyawardhane DL, Godoy-Ruiz R, Adipietro KA, Varney KM, Rustandi RR, Pozharski E, Weber DJ. The Importance of Therapeutically Targeting the Binary Toxin from Clostridioides difficile. International Journal of Molecular Sciences. 2021; 22(6):2926. https://doi.org/10.3390/ijms22062926
Chicago/Turabian StyleAbeyawardhane, Dinendra L., Raquel Godoy-Ruiz, Kaylin A. Adipietro, Kristen M. Varney, Richard R. Rustandi, Edwin Pozharski, and David J. Weber. 2021. "The Importance of Therapeutically Targeting the Binary Toxin from Clostridioides difficile" International Journal of Molecular Sciences 22, no. 6: 2926. https://doi.org/10.3390/ijms22062926
APA StyleAbeyawardhane, D. L., Godoy-Ruiz, R., Adipietro, K. A., Varney, K. M., Rustandi, R. R., Pozharski, E., & Weber, D. J. (2021). The Importance of Therapeutically Targeting the Binary Toxin from Clostridioides difficile. International Journal of Molecular Sciences, 22(6), 2926. https://doi.org/10.3390/ijms22062926