
Involvement of IL-4, IL-13 and Their Receptors in Pancreatic Cancer


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
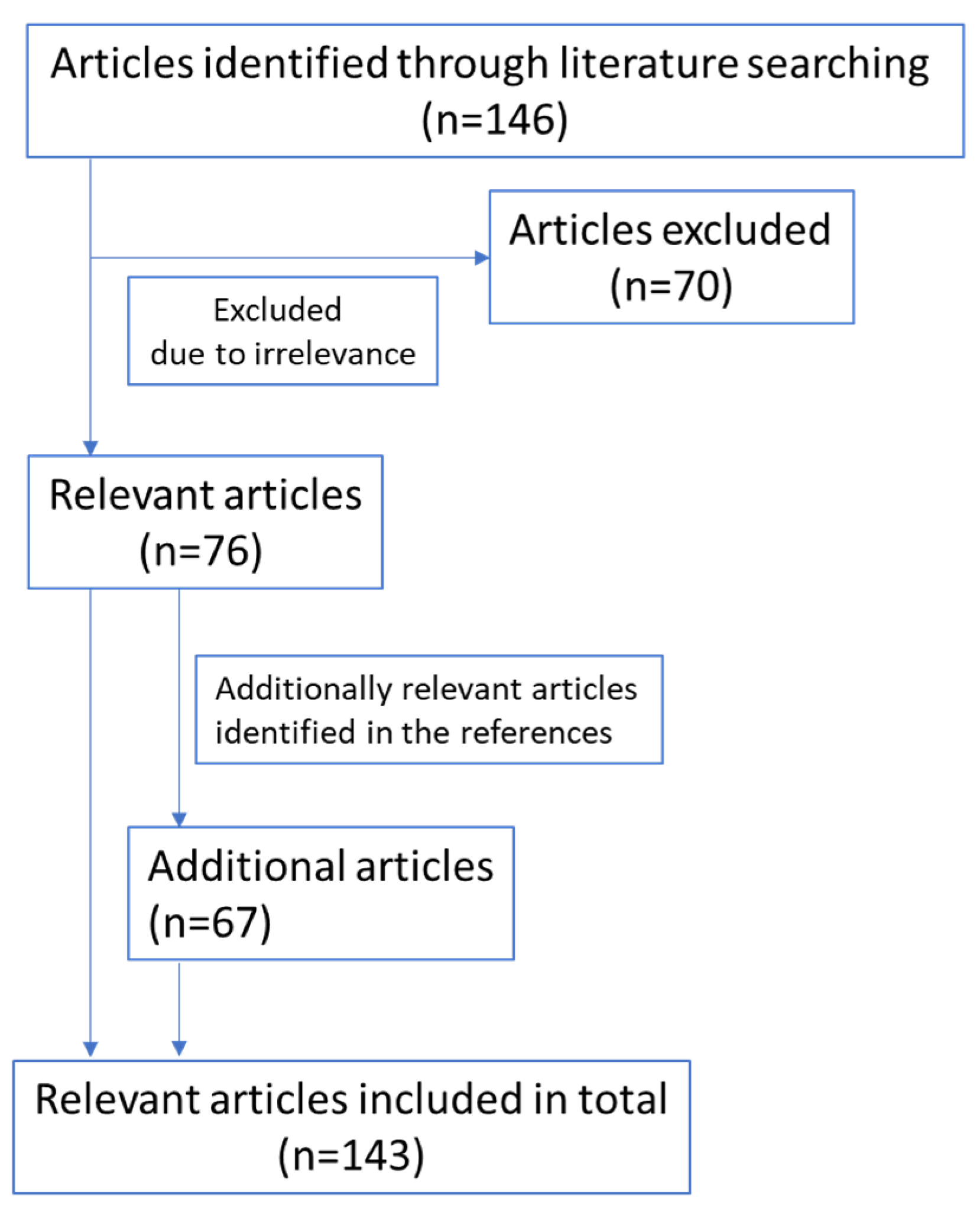
2. Methods
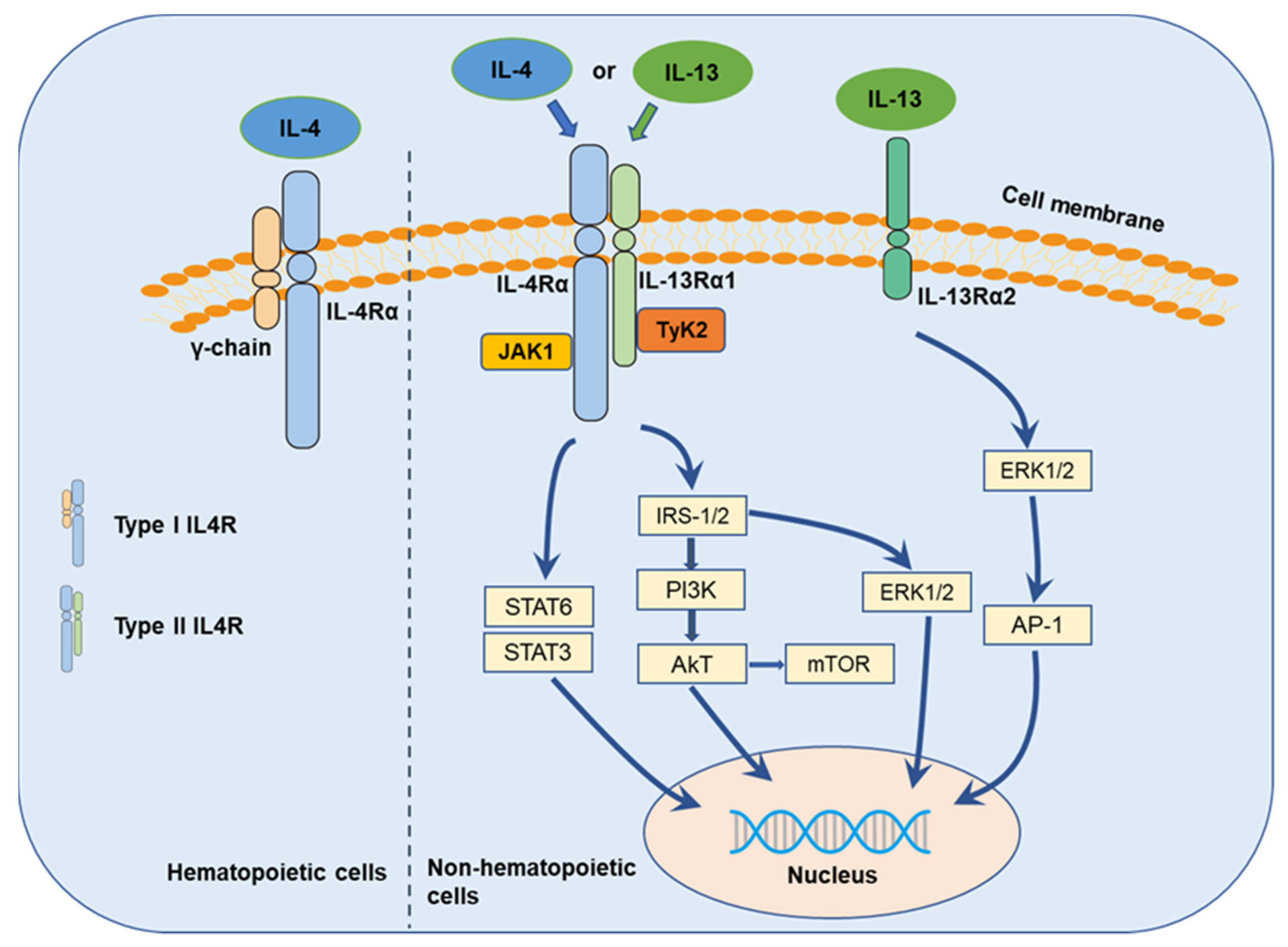
3. IL-4 and IL-13 Cytokines and Their Receptors
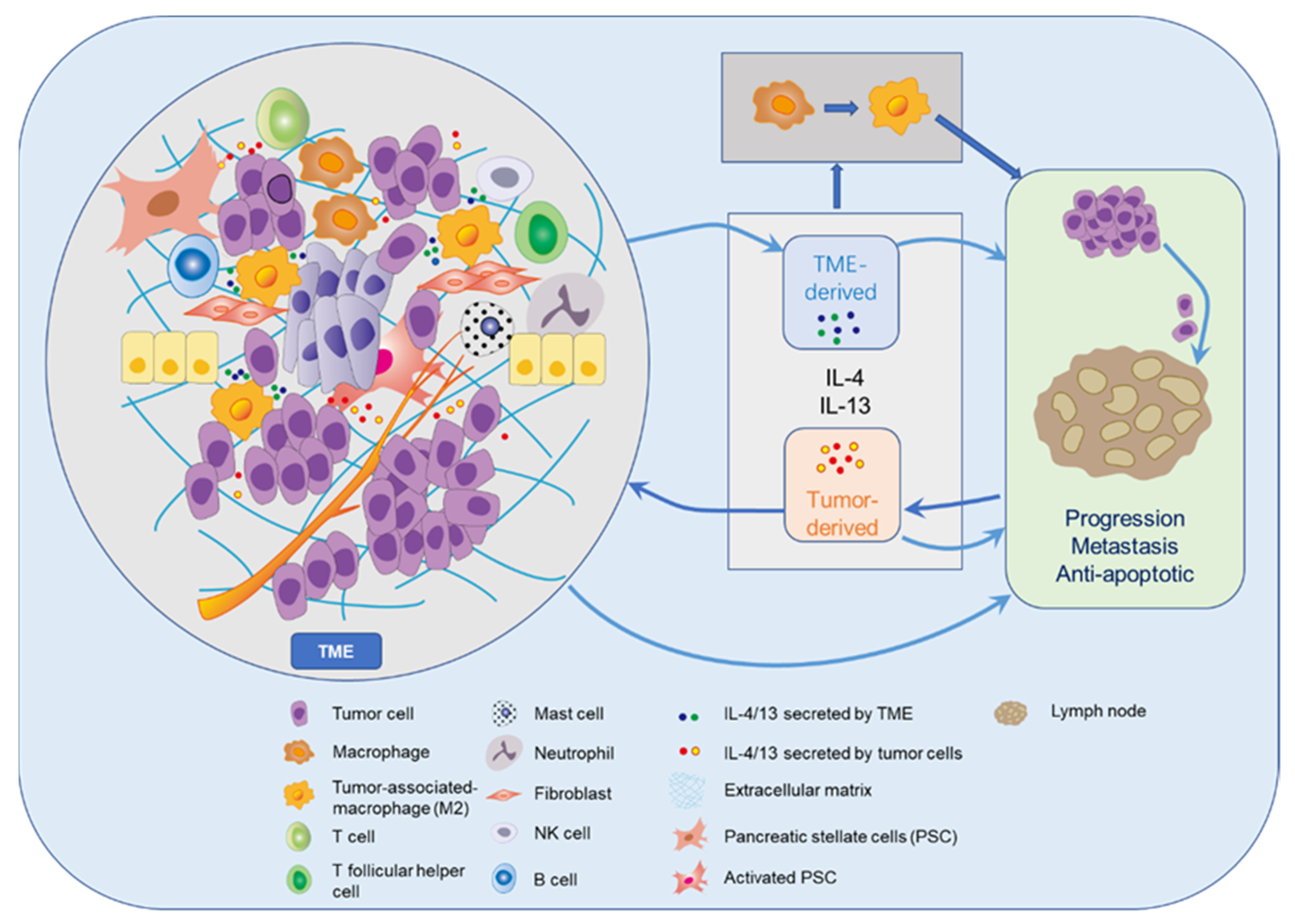
4. IL-4 in Pancreatic Cancer
5. IL-13 in Pancreatic Cancer
6. IL-4R in Pancreatic Cancer
7. IL-13R in Pancreatic Cancer
8. Future Directions of Research
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | Protein kinase B |
| AP-1 | Activator protein 1 |
| ELISA | The enzyme-linked immunosorbent assay |
| EMT | Epithelial–mesenchymal transition |
| ERK | Extracellular signal-regulated kinase |
| IL | Interleukin |
| IL-4Rα | IL-4-receptor alpha |
| IL-13Rα | IL-13-receptor alpha |
| JAK | Janus tyrosine kinase |
| JNK | c-Jun N-terminal kinase |
| MAPK | Mitogen-activated protein kinase |
| MMPs | Matrix metalloproteinases |
| mTOR | The mechanistic target of rapamycin |
| PCR | Polymerase chain reaction |
| PDAC | Pancreatic ductal adenocarcinoma |
| PE | Pseudomonas exotoxins |
| PSCs | Pancreatic stellate cells |
| Γc | Common γ chain |
| RNA | Ribonucleic acid |
| STAT | Signal transducer and activator of transcription |
| TAMs | Tumor-associated macrophages |
| TME | Tumor microenvironment |
| Tyk2 | Tyrosine kinase 2 |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Neoptolemos, J.P.; Kleeff, J.; Michl, P.; Costello, E.; Greenhalf, W.; Palmer, D.H. Therapeutic developments in pancreatic cancer: Current and future perspectives. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 333–348. [Google Scholar] [CrossRef]
- Lei, X.; Lei, Y.; Li, J.-K.; Du, W.-X.; Li, R.-G.; Yang, J.; Li, J.; Li, F.; Tan, H.-B. Immune cells within the tumor microenvironment: Biological functions and roles in cancer immunotherapy. Cancer Lett. 2020, 470, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Parayath, N.; Padmakumar, S.; Nair, S.V.; Menon, D.; Amiji, M.M. Strategies for targeting cancer immunotherapy through modulation of the tumor microenvironment. Regen. Eng. Transl. Med. 2019, 6, 29–49. [Google Scholar] [CrossRef]
- Thakkar, S.; Sharma, D.; Kalia, K.; Tekade, R.K. Tumor microenvironment targeted nanotherapeutics for cancer therapy and diagnosis: A review. Acta Biomater. 2020, 101, 43–68. [Google Scholar] [CrossRef] [PubMed]
- Farrow, B.; Albo, D.; Berger, D.H. The role of the tumor microenvironment in the progression of pancreatic cancer. J. Surg. Res. 2008, 149, 319–328. [Google Scholar] [CrossRef]
- Mbeunkui, F.; Johann, D.J. Cancer and the tumor microenvironment: A review of an essential relationship. Cancer Chemother. Pharmacol. 2008, 63, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arneth, B. Tumor microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhao, J.; Zhang, L.; Wei, F.; Lian, Y.; Wu, Y.; Gong, Z.; Zhang, S.; Zhou, J.; Cao, K.; et al. Role of tumor microenvironment in tumorigenesis. J. Cancer 2017, 8, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noy, R.; Pollard, J.W. Tumor-Associated Macrophages: From Mechanisms to Therapy. Immunity 2014, 41, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Chen, B.; Lin, Y.; Zhou, Y.; Li, X. Legumain promotes gastric cancer progression through tumor-associated macrophages in vitro and in vivo. Int. J. Biol. Sci. 2020, 16, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C.; Pathanjeli, P.; Wu, Z.; Bao, L.; Goo, L.E.; Yates, J.A.; Oliver, C.R.; Soellner, M.B.; Merajver, S.D. IL-4/IL-13 stimulated macrophages enhance breast cancer invasion via Rho-GTPase regulation of synergistic VEGF/CCL-18 signaling. Front. Oncol. 2019, 9, 456. [Google Scholar] [CrossRef] [Green Version]
- Cassetta, L.; Pollard, J.W. Targeting macrophages: Therapeutic approaches in cancer. Nat. Rev. Drug Discov. 2018, 17, 887–904. [Google Scholar] [CrossRef] [PubMed]
- DeNardo, D.G.; Barreto, J.B.; Andreu, P.; Vasquez, L.; Tawfik, D.; Kolhatkar, N.; Coussens, L.M. CD4+ T cells regulate pulmonary metastasis of mammary carcinomas by enhancing protumor properties of macrophages. Cancer Cell 2009, 16, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Li, W.; Pan, L.; Fu, E.; Xie, Y.; Chen, M.; Mu, D. Lewis lung cancer cells promote SIGNR1(CD209b)–Mediated macrophages polarization induced by IL-4 to facilitate immune evasion. J. Cell. Biochem. 2015, 117, 1158–1166. [Google Scholar] [CrossRef]
- Leonard, W.J.; Lin, J.X.; O’Shea, J.J. The gammac family of cytokines: Basic biology to therapeutic ramifications. Immunity 2019, 50, 832–850. [Google Scholar] [CrossRef] [Green Version]
- Stroud, R.M.; Wells, J.A. Mechanistic diversity of cytokine receptor signaling across cell membranes. Sci. Signal. 2004, 231, re7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraga, I.; Richter, D.; Wilmes, S.; Winkelmann, H.; Jude, K.; Thomas, C.; Suhoski, M.M.; Engleman, E.G.; Piehler, J.; Garcia, K.C. Instructive roles for cytokine-receptor binding parameters in determining signaling and functional potency. Sci. Signal. 2015, 8, ra114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autenshlyus, A.I.; Davletova, K.I.; Studenikina, A.A.; Mikhaylova, E.S.; Varaksin, N.A.; Zhurakovsky, I.P.; Proskura, A.V.; Sidorov, S.V.; Lyakhovich, V.V. Cytokine production by blood immune cells, tumor and its microenvironment, characteristics of extracellular matrix in patients with invasive ductal carcinoma of no special type. Biochem. (Moscow) Suppl. Ser. B Biomed. Chem. 2020, 14, 44–51. [Google Scholar] [CrossRef]
- Ben-Baruch, A. Host microenvironment in breast cancer development: Inflammatory cells, cytokines and chemokines in breast cancer progression: Reciprocal tumor–Microenvironment interactions. Breast Cancer Res. 2002, 5, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, K.; Sakurai, M.; Yamamoto, Y.; Suzuki, E.; Tsuda, M.; Kataoka, T.R.; Hirata, M.; Nishie, M.; Nojiri, T.; Kumazoe, M.; et al. Alteration of specific cytokine expression patterns in patients with breast cancer. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.-E.; Shirota, H.; Kasahara, Y.; Saijo, K.; Ishioka, C. IL-4 blockade alters the tumor microenvironment and augments the response to cancer immunotherapy in a mouse model. Cancer Immunol. Immunother. 2017, 66, 1485–1496. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Dou, X.Q.; Wang, F.; Zhang, J.; Wang, X.L.; Xu, G.L.; Xiang, S.S.; Gao, X.; Fu, J.; Song, H.F. IL-4Ralpha aptamer-liposome-CpG oligodeoxynucleotides suppress tumour growth by targeting the tumour microenvironment. J. Drug Target. 2017, 25, 275–283. [Google Scholar] [CrossRef]
- Braddock, M.; Hanania, N.A.; Sharafkhaneh, A.; Colice, G.; Carlsson, M. Potential risks related to modulating Interleukin-13 and Interleukin-4 signalling: A systematic review. Drug Saf. 2018, 41, 489–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyfizadeh, N.; Seyfizadeh, N.; Gharibi, T.; Babaloo, Z. Interleukin-13 as an important cytokine: A review on its roles in some human diseases. Acta Microbiol. Immunol. Hung. 2015, 62, 341–378. [Google Scholar] [CrossRef] [Green Version]
- Hallett, M.A.; Venmar, K.T.; Fingleton, B. Cytokine stimulation of epithelial cancer cells: The similar and divergent functions of IL-4 and IL-13. Cancer Res. 2012, 72, 6338–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwasniak, K.; Czarnik-Kwasniak, J.; Maziarz, A.; Aebisher, D.; Zielinska, K.; Karczmarek-Borowska, B.; Tabarkiewicz, J. Scientific reports concerning the impact of Interleukin 4, Interleukin 10 and transforming growth factor beta on cancer cells. Cent. Eur. J. Immunol. 2019, 44, 190–200. [Google Scholar] [CrossRef]
- Wilson, C.B.; Rowell, E.; Sekimata, M. Epigenetic control of T-helper-cell differentiation. Nat. Rev. Immunol. 2009, 9, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Heeb, L.E.M.; Egholm, C.; Boyman, O. Evolution and function of Interleukin-4 receptor signaling in adaptive immunity and neutrophils. Genes Immun. 2020, 21, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Moran, A.; Pavord, I.D. Anti-IL-4/IL-13 for the treatment of asthma: The story so far. Expert Opin. Biol. Ther. 2020, 20, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Moy, F.J.; Diblasio, E.; Wilhelm, J.; Powers, R. Solution structure of human IL-13 and implication for receptor binding. J. Mol. Biol. 2001, 310, 219–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurawski, S.M.; Vega, F.; Huyghe, B., Jr.; Zurawski, G. Receptors for Interleukin-13 and Interleukin-4 are complex and share a novel component that functions in signal transduction. EMBO J. 1993, 12, 2663–2670. [Google Scholar] [CrossRef] [PubMed]
- McLeod, J.J.; Baker, B.; Ryan, J.J. Mast cell production and response to IL-4 and IL-13. Cytokine 2015, 75, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Cosmi, L.; Maggi, L.; Santarlasci, V.; Capone, M.; Cardilicchia, E.; Frosali, F.; Querci, V.; Angeli, R.; Matucci, A.; Fambrini, M.; et al. Identification of a novel subset of human circulating memory CD4+ T cells that produce both IL-17A and IL-4. J. Allergy Clin. Immunol. 2010, 125, 222–230.e4. [Google Scholar] [CrossRef]
- Brown, K.D.; Zurawski, S.M.; Mosmann, T.R.; Zurawski, G. A family of small inducible proteins secreted by leukocytes are members of a new superfamily that includes leukocyte and fibroblast-derived inflammatory agents, growth factors, and indicators of various activation processes. J. Immunol. 1989, 142, 679–687. [Google Scholar]
- Seder, R.A.; Paul, W.E. Acquisition of lymphokine-producing phenotype by CD4+ T cells. Annu. Rev. Immunol. 1994, 12, 635–673. [Google Scholar] [CrossRef] [PubMed]
- Nelms, K.; Keegan, A.D.; Zamorano, J.; Ryan, J.J.; Paul, W.E. The IL-4 RECEPTOR: Signaling mechanisms and biologic functions. Annu. Rev. Immunol. 1999, 17, 701–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minty, A.; Chalon, P.; Derocq, J.M.; Dumont, X.; Guillemot, J.C.; Kaghad, M.; Labit, C.; Leplatois, P.; Liauzun, P.; Miloux, B.; et al. Interleukin-13 is a new human lymphokine regulating inflammatory and immune responses. Nature 1993, 362, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Geskin, L.J.; Viragova, S.; Stolz, D.B.; Fuschiotti, P. Interleukin-13 is overexpressed in cutaneous T-cell lymphoma cells and regulates their proliferation. Blood 2015, 125, 2798–2805. [Google Scholar] [CrossRef] [Green Version]
- Skinnider, B.F.; Kapp, U.; Mak, T.W. Interleukin 13: A growth factor in hodgkin lymphoma. Int. Arch. Allergy Immunol. 2001, 126, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Camp, B.J.; Dyhrman, S.T.; A Memoli, V.; A Mott, L.; Barth, R.J. In situ cytokine production by breast cancer tumor-infiltrating lymphocytes. Ann. Surg. Oncol. 1996, 3, 176–184. [Google Scholar] [CrossRef]
- Srabovici, N.; Mujagic, Z.; Mujanovic-Mustedanagic, J.; Muminovic, Z.; Softic, A.; Begic, L. Interleukin 13 expression in the primary breast cancer tumour tissue. Biochem. Med. (Zagreb) 2011, 21, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wang, S.; Sun, M.; Zhang, C.; Wei, C.; Yang, C.; Doi, R.; Liu, Q.; Xiong, B. miR-195-5p/NOTCH2-mediated EMT modulates IL-4 secretion in colorectal cancer to affect M2-like TAM polarization. J. Hematol. Oncol. 2019, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Formentini, A.; Braun, P.; Fricke, H.; Link, K.-H.; Henne-Bruns, D.; Kornmann, M. Expression of Interleukin-4 and Interleukin-13 and their receptors in colorectal cancer. Int. J. Color. Dis. 2012, 27, 1369–1376. [Google Scholar] [CrossRef]
- Aziz, S.; Ahmed, S.S.; Ali, A.; Khan, F.A.; Zulfiqar, G.; Iqbal, J.; Khan, A.A.; Shoaib, M. Salivary Immunosuppressive Cytokines IL-10 and IL-13 are significantly elevated in oral squamous cell carcinoma patients. Cancer Investig. 2015, 33, 318–328. [Google Scholar] [CrossRef]
- Kornmann, M.; Kleeff, J.; Debinski, W.; Korc, M. Pancreatic cancer cells express interleukin-13 and -4 receptors, and their growth is inhibited by Pseudomonas exotoxin coupled to interleukin-13 and -4. Anticancer. Res. 1999, 19, 125–131. [Google Scholar]
- Surana, R.; Wang, S.; Xu, W.; Jablonski, S.A.; Weiner, L.M. IL4 Limits the efficacy of tumor-targeted antibody therapy in a murine model. Cancer Immunol. Res. 2014, 2, 1103–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankaitis, K.V.; Fingleton, B. Targeting IL4/IL4R for the treatment of epithelial cancer metastasis. Clin. Exp. Metastasis 2015, 32, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.; Lombardo, Y.; Francipane, M.G.; Alea, M.P.; Cammareri, P.; Iovino, F.; Di Stefano, A.B.; Di Bernardo, C.; Agrusa, A.; Condorelli, G.; et al. Apoptosis resistance in epithelial tumors is mediated by tumor-cell-derived interleukin-4. Cell Death Differ. 2008, 15, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Prokopchuk, O.; Liu, Y.; Hennebruns, D.; Kornmann, M. Interleukin-4 enhances proliferation of human pancreatic cancer cells: Evidence for autocrine and paracrine actions. Br. J. Cancer 2005, 92, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Härtlova, A.; Gierliński, M.; Prescott, A.; Castellvi, J.; Losa, J.H.; Petersen, S.K.; Wenzel, U.A.; Dill, B.D.; Emmerich, C.H.; et al. Triggering MSR1 promotes JNK-mediated inflammation in IL-4-activated macrophages. EMBO J. 2019, 38, e100299. [Google Scholar] [CrossRef]
- Erb, H.H.H.; Guggenberger, F.; Santer, F.R.; Culig, Z. Interleukin-4 induces a CD44high/CD49bhighPC3 subpopulation with tumor-initiating characteristics. J. Cell. Biochem. 2018, 119, 4103–4112. [Google Scholar] [CrossRef] [Green Version]
- Todaro, M.; Alea, M.P.; Di Stefano, A.B.; Cammareri, P.; Vermeulen, L.; Iovino, F.; Tripodo, C.; Russo, A.; Gulotta, G.; Medema, J.P.; et al. Colon cancer stem cells dictate tumor growth and resist cell death by production of Interleukin-4. Cell Stem Cell 2007, 1, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Venmar, K.T.; Kimmel, D.W.; Cliffel, D.E.; Fingleton, B. IL4 receptor α mediates enhanced glucose and glutamine metabolism to support breast cancer growth. Biochim. Biophys. Acta (BBA) Bioenerg. 2015, 1853, 1219–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koller, F.L.; Hwang, D.G.; Dozier, E.; Fingleton, B. Epithelial interleukin-4 receptor expression promotes colon tumor growth. Carcinogenesis 2010, 31, 1010–1017. [Google Scholar] [CrossRef]
- Roca, H.; Craig, M.J.; Ying, C.; Varsos, Z.S.; Czarnieski, P.; Alva, A.S.; Hernandez, J.; Fuller, D.; Daignault, S.; Healy, P.N.; et al. IL-4 induces proliferation in prostate cancer PC3 cells under nutrient-depletion stress through the activation of the JNK-pathway and survivin upregulation. J. Cell. Biochem. 2011, 113, 1569–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.H.; Yang, X.Z.; Li, P.D.; Yuan, Q.; Liu, X.H.; Yuan, J.; Zhang, W.J. IL-4/Stat6 activities correlate with apoptosis and metastasis in colon cancer cells. Biochem. Biophys. Res. Commun. 2008, 369, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Venmar, K.T.; Carter, K.J.; Hwang, D.G.; Dozier, E.A.; Fingleton, B. IL4 receptor ILR4α regulates metastatic colonization by mammary tumors through multiple signaling pathways. Cancer Res. 2014, 74, 4329–4340. [Google Scholar] [CrossRef] [Green Version]
- Hilton, D.J.; Zhang, J.G.; Metcalf, D.; Alexander, W.S.; Nicola, N.A.; Willson, T.A. Cloning and characterization of a binding subunit of the interleukin 13 receptor that is also a component of the interleukin 4 receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 497–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obiri, N.I.; Debinski, W.; Leonard, W.J.; Puri, R.K. Receptor for interleukinInteraction with interleukin 4 by a mechanism that does not involve the common gamma chain shared by receptors for interleukins 2, 4, 7, 9, and 15. J. Biol. Chem. 1995, 270, 8797–8804. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; I Obiri, N.; Puri, R.K. Structure of and signal transduction through interleukin-4 and interleukin-13 receptors. Int. J. Mol. Med. 1998, 1, 551–558. [Google Scholar] [CrossRef] [PubMed]
- LaPorte, S.L.; Juo, Z.S.; Vaclavikova, J.; Colf, L.A.; Qi, X.; Heller, N.M.; Keegan, A.D.; Garcia, K.C. Molecular and structural basis of cytokine receptor pleiotropy in the interleukin-4/13 system. Cell 2008, 132, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, A.-L.; Nordgren, I.K.; Campbell-Harding, G.; Holloway, J.W.; Holgate, S.T.; Davies, D.E.; Tavassoli, A. The association of the cytoplasmic domains of interleukin 4 receptor alpha and interleukin 13 receptor alpha 2 regulates interleukin 4 signaling. Mol. BioSyst. 2013, 9, 3009–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mentink-Kane, M.M.; Wynn, T.A. Opposing roles for IL-13 and IL-13 receptor alpha2 in health and disease. Immunol. Rev. 2004, 202, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Kelly-Welch, A.; Hanson, E.M.; Keegan, A.D. Interleukin-13 (IL-13) pathway. Sci. Signal. 2005, 2005, cm8. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Taguchi, J.; Murata, T.; Puri, R.K. The interleukin-13 receptor α2 chain: An essential component for binding and internalization but not for interleukin-13–induced signal transduction through the STAT6 pathway. Blood 2001, 97, 2673–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly-Welch, A.E.; Hanson, E.M.; Boothby, M.R.; Keegan, A.D. Interleukin-4 and Interleukin-13 signaling connections maps. Science 2003, 300, 1527–1528. [Google Scholar] [CrossRef]
- Chomarat, P.; Banchereau, J. Interleukin-4 and lnterleukin-13: Their similarities and discrepancies. Int. Rev. Immunol. 1998, 17, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Leonard, W.J.; Lin, J.-X. Cytokine receptor signaling pathways. J. Allergy Clin. Immunol. 2000, 105, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Jiang, L.; Hao, S.; Liu, Z.; Ding, S.; Zhang, W.; Yang, X.; Li, S. Activation of the IL-4/STAT6 signaling pathway promotes lung cancer progression by increasing M2 myeloid cells. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.D.; Baik, J.S.; Lee, J.-H.; Mun, S.-W.; Yi, J.M.; Park, M.-T. The malignancy of liver cancer cells is increased by IL-4/ERK/AKT signaling axis activity triggered by irradiated endothelial cells. J. Radiat. Res. 2020, 61, 376–387. [Google Scholar] [CrossRef]
- Sheng, J.; Yang, Y.; Cui, Y.; He, S.; Wang, L.; Liu, L.; He, Q.; Lv, T.; Han, W.; Yu, W.; et al. M2 macrophage-mediated interleukin-4 signalling induces myofibroblast phenotype during the progression of benign prostatic hyperplasia. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wills-Karp, M.; Finkelman, F.D. Untangling the complex web of IL-4- and IL-13-mediated signaling pathways. Sci. Signal. 2008, 1, pe55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Joshi, B.H.; Puri, R.K. IL-13 regulates cancer invasion and metastasis through IL-13Ralpha2 via ERK/AP-1 pathway in mouse model of human ovarian cancer. Int. J. Cancer 2012, 131, 344–356. [Google Scholar] [CrossRef]
- Song, X.; Traub, B.; Shi, J.; Kornmann, M. Possible roles of Interleukin-4 and -13 and their receptors in gastric and colon cancer. Int. J. Mol. Sci. 2021, 22, 727. [Google Scholar] [CrossRef] [PubMed]
- Yako, Y.Y.; Brand, M.; Smith, M.; Kruger, D. Inflammatory cytokines and angiogenic factors as potential biomarkers in South African pancreatic ductal adenocarcinoma patients: A preliminary report. Pancreatology 2017, 17, 438–444. [Google Scholar] [CrossRef]
- Chen, J.; Du, M.; Li, W.-B.; Lei, C.; Yang, B.; Tang, H.-K.; Zhai, Z.-M.; Ma, J.-L.; Li, Q. The relationship between CD4(-) CD8(-) T cells in the peripheral blood of patients with pancreatic carcinoma and IL-4, IFN-gamma levels. Zhonghua Wai Ke Za Zhi [Chinese J. Surg.] 2009, 47, 995–998. [Google Scholar] [PubMed]
- Piro, G.; Simionato, F.; Carbone, C.; Frizziero, M.; Malleo, G.; Zanini, S.; Casolino, R.; Santoro, R.; Mina, M.M.; Zecchetto, C.; et al. A circulating T(H)2 cytokines profile predicts survival in patients with resectable pancreatic adenocarcinoma. Oncoimmunology 2017, 6, e1322242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gocheva, V.; Wang, H.-W.; Gadea, B.B.; Shree, T.; Hunter, K.E.; Garfall, A.L.; Berman, T.; Joyce, J.A. IL-4 induces cathepsin protease activity in tumor-associated macrophages to promote cancer growth and invasion. Genes Dev. 2010, 24, 241–255. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Xu, J.Y.; Shi, X.Y.; Huang, W.; Ruan, T.Y.; Xie, P.; Ding, J.L. M2-polarized tumor-associated macrophages promoted epithelial–mesenchymal transition in pancreatic cancer cells, partially through TLR4/IL-10 signaling pathway. Lab. Investig. 2013, 93, 844–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Konaté, M.M.; Lu, J.; Makhlouf, H.; Chuaqui, R.; Antony, S.; Meitzler, J.L.; Difilippantonio, M.J.; Liu, H.; Juhasz, A.; et al. IL-4 and IL-17A cooperatively promote hydrogen peroxide production, oxidative DNA damage, and upregulation of dual oxidase 2 in human colon and pancreatic cancer cells. J. Immunol. 2019, 203, 2532–2544. [Google Scholar] [CrossRef] [PubMed]
- Edderkaoui, M.; Xu, S.; Chheda, C.; Morvaridi, S.; Hu, R.W.; Grippo, P.J.; Mascariñas, E.; Principe, D.R.; Knudsen, B.; Xue, J.; et al. HDAC3 mediates smoking-induced pancreatic cancer. Oncotarget 2016, 7, 7747–7760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokopchuk, O.; Steinacker, J.M.; Nitsche, U.; Otto, S.; Bachmann, J.; Schubert, E.C.; Friess, H.; Martignoni, M.E. IL-4 mRNA is downregulated in the liver of pancreatic cancer patients suffering from cachexia. Nutr. Cancer 2016, 69, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Costamagna, D.; Duelen, R.; Penna, F.; Neumann, D.; Costelli, P.; Sampaolesi, M. Interleukin-4 administration improves muscle function, adult myogenesis, and lifespan of colon carcinoma-bearing mice. J. Cachex Sarcopenia Muscle 2020, 11, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Kornmann, M.; Maruyama, H.; Bergmann, U.; Tangvoranuntakul, P.; Beger, H.G.; White, M.F.; Korc, M. Enhanced expression of the insulin receptor substrate-2 docking protein in human pancreatic cancer. Cancer Res. 1998, 58, 4250–4254. [Google Scholar]
- Traub, B.; Sun, L.; Padmanabhan, J.; Xu, P.; Lemke, J.; Paschke, S.; Henne-Bruns, D.; Knippschild, U.; Kornmann, M. Endogenously expressed IL-4Rα promotes the malignant phenotype of human pancreatic cancer in vitro and in vivo. Int. J. Mol. Sci. 2017, 18, 716. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Hutzen, B.; Li, P.-K.; Ball, S.; Zuo, M.; DeAngelis, S.; Foust, E.; Sobo, M.; Friedman, L.; Bhasin, D.; et al. A novel small molecule, LLL12, inhibits STAT3 phosphorylation and activities and exhibits potent growth-suppressive activity in human cancer cells. Neoplasia 2010, 12, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabitass, R.F.; Annels, N.E.; Stocken, D.D.; Pandha, H.A.; Middleton, G.W. Elevated myeloid-derived suppressor cells in pancreatic, esophageal and gastric cancer are an independent prognostic factor and are associated with significant elevation of the Th2 cytokine interleukin-13. Cancer Immunol. Immunother. 2011, 60, 1419–1430. [Google Scholar] [CrossRef] [Green Version]
- Formentini, A.; Prokopchuk, O.; Sträter, J.; Kleeff, J.; Grochola, L.F.; Leder, G.; Henne-Bruns, D.; Korc, M.; Kornmann, M. Interleukin-13 exerts autocrine growth-promoting effects on human pancreatic cancer, and its expression correlates with a propensity for lymph node metastases. Int. J. Color. Dis. 2008, 24, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Feurino, L.W.; Li, F.; Wang, H.; Zhai, Q.; Fisher, W.E.; Chen, C.; Yao, Q. Thymosinα1 stimulates cell proliferation by activating ERK1/2, JNK, and increasing cytokine secretion in human pancreatic cancer cells. Cancer Lett. 2007, 248, 58–67. [Google Scholar] [CrossRef]
- Ma, Y.; Hwang, R.F.; Logsdon, C.D.; Ullrich, S.E. Dynamic mast cell-stromal cell interactions promote growth of pancreatic cancer. Cancer Res. 2013, 73, 3927–3937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fichtner-Feigl, S.; Strober, W.; Kawakami, K.; Puri, R.K.; Kitani, A. IL-13 signaling through the IL-13alpha2 receptor is involved in induction of TGF-beta1 production and fibrosis. Nat. Med. 2006, 12, 99–106. [Google Scholar] [CrossRef]
- Joshi, B.H.; Hogaboam, C.; Dover, P.; Husain, S.R.; Puri, R.K. Role of Interleukin-13 in cancer, pulmonary fibrosis, and other TH2-type diseases. Vitam. Horm. 2006, 74, 479–504. [Google Scholar] [PubMed]
- Xue, J.; Sharma, V.; Hsieh, M.H.; Chawla, A.; Murali, R.; Pandol, S.J.; Habtezion, A. Alternatively activated macrophages promote pancreatic fibrosis in chronic pancreatitis. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liou, G.-Y.; Bastea, L.; Fleming, A.; Döppler, H.; Edenfield, B.H.; Dawson, D.W.; Zhang, L.; Bardeesy, N.; Storz, P. The presence of Interleukin-13 at pancreatic ADM/PanIN lesions alters macrophage populations and mediates pancreatic tumorigenesis. Cell Rep. 2017, 19, 1322–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimamura, T.; Fujisawa, T.; Husain, S.R.; Joshi, B.; Puri, R.K. Interleukin 13 mediates signal transduction through interleukin 13 receptor alpha2 in pancreatic ductal adenocarcinoma: Role of IL-13 Pseudomonas exotoxin in pancreatic cancer therapy. Clin. Cancer Res. 2010, 16, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T.; Joshi, B.H.; Nakajima, A.; Puri, R.K. A novel role of Interleukin-13 Receptor 2 in pancreatic cancer invasion and metastasis. Cancer Res. 2009, 69, 8678–8685. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, T.; Royal, R.E.; Kioi, M.; Nakajima, A.; Husain, S.R.; Puri, R.K. Interleukin-4 cytotoxin therapy synergizes with gemcitabine in a mouse model of pancreatic ductal adenocarcinoma. Cancer Res. 2007, 67, 9903–9912. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K.; Kawakami, M.; Husain, S.R.; Puri, R.K. Targeting interleukin-4 receptors for effective pancreatic cancer therapy. Cancer Res. 2002, 62, 3575–3580. [Google Scholar]
- Olson, S.H.; Orlow, I.; Simon, J.; Tommasi, D.; Roy, P.; Bayuga, S.; Ludwig, E.; Zauber, A.G.; Kurtz, R.C. Allergies, variants in IL-4 and IL-4R alpha genes, and risk of pancreatic cancer. Cancer Detect Prev. 2007, 31, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.; Sukumaran, S.; Bajgain, P.; Watanabe, N.; Heslop, H.E.; Rooney, C.M.; Brenner, M.K.; Fisher, W.E.; Leen, A.M.; Vera, J.F. Improving chimeric antigen receptor-modified T cell function by reversing the immunosuppressive tumor microenvironment of pancreatic cancer. Mol. Ther. 2017, 25, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Horibe, T.; Kohno, M.; Haramoto, M.; Ohara, K.; Puri, R.K.; Kawakami, K. Targeting Interleukin-4 receptor α with hybrid peptide for effective cancer therapy. Mol. Cancer Ther. 2012, 11, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, K.; Kawakami, M.; Snoy, P.J.; Husain, S.R.; Puri, R.K. In vivo overexpression of IL-13 receptor α2 chain inhibits tumorigenicity of human breast and pancreatic tumors in immunodeficient mice. J. Exp. Med. 2001, 194, 1743–1754. [Google Scholar] [CrossRef]
- Fujisawa, T.; Joshi, B.H.; Puri, R.K. Histone modification enhances the effectiveness of IL-13 receptor targeted immunotoxin in murine models of human pancreatic cancer. J. Transl. Med. 2011, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Shimamura, T.; Goto, K.; Nakagawa, R.; Muroyama, R.; Ino, Y.; Horiuchi, H.; Endo, I.; Maeda, S.; Harihara, Y.; et al. A novel role of Interleukin 13 receptor alpha2 in perineural invasion and its association with poor prognosis of patients with pancreatic ductal adenocarcinoma. Cancers 2020, 12, 1294. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Joshi, B.H.; Puri, R.K. Sensitization of cancer cells to Interleukin 13-pseudomona sexotoxin-induced cell death by gene transfer of Interleukin 13 receptor α chain. Hum. Gene Ther. 2000, 11, 1829–1835. [Google Scholar] [CrossRef]
- Kawakami, K.; Husain, S.R.; Bright, R.K.; Puri, R.K. Gene transfer of interleukin 13 receptor alpha2 chain dramatically enhances the antitumor effect of IL-13 receptor-targeted cytotoxin in human prostate cancer xenografts. Cancer Gene Ther. 2001, 8, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K.; Kawakami, M.; Puri, R.K. IL-13 receptor-targeted cytotoxin cancer therapy leads to complete eradication of tumors with the aid of phagocytic cells in nude mice model of human cancer. J. Immunol. 2002, 169, 7119–7126. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K.; Kawakami, M.; Joshi, B.H.; Puri, R.K. Interleukin-13 receptor-targeted cancer therapy in an immunodeficient animal model of human head and neck cancer. Cancer Res. 2001, 61, 6194–6200. [Google Scholar]
- Kawakami, K.; Kawakami, M.; Puri, R.K. Specifically targeted killing of interleukin-13 (IL-13) receptor-expressing breast cancer by IL-13 fusion cytotoxin in animal model of human disease. Mol. Cancer Ther. 2004, 3, 137–147. [Google Scholar]
- Kawakami, K.; Husain, S.R.; Puri, R.K.; Kawakami, M. Potent antitumor activity of IL-13 cytotoxin in human pancreatic tumors engineered to express IL-13 receptor α2 chain in vivo. Gene Ther. 2003, 10, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Nakashima, H.; Nakajima, A.; Joshi, B.H.; Puri, R.K. Targeting IL-13Ralpha2 in human pancreatic ductal adenocarcinoma with combination therapy of IL-13-PE and gemcitabine. Int. J. Cancer 2011, 128, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Stish, B.J.; Vickers, S.M.; Buchsbaum, D.J.; Saluja, A.K.; Vallera, D.A. A new drug delivery method of bispecific ligand-directed toxins, which reduces toxicity and promotes efficacy in a model of orthotopic pancreatic cancer. Pancreas 2010, 39, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallera, D.A.; Stish, B.J.; Shu, Y.; Chen, H.; Saluja, A.K.; Buchsbaum, D.J.; Vickers, S.M. Genetically designing a more potent antipancreatic cancer agent by simultaneously co-targeting human IL13 and EGF receptors in a mouse xenograft model. Gut 2008, 57, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Puri, R.K.; Tran, S.E.F.; Holmström, T.H.; Ahonen, M.; Kähäri, V.-M.; Eriksson, J.E. Characterization of a powerful high affinity antagonist that inhibits biological activities of human Interleukin-13. J. Biol. Chem. 2001, 276, 15185–15191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kioi, M.; Kawakami, K.; Puri, R.K. Mechanism of action of interleukin-13 antagonist (IL-13E13K) in cells expressing various types of IL-4R. Cell. Immunol. 2004, 229, 41–51. [Google Scholar] [CrossRef] [PubMed]
- König, A.; Vilsmaier, T.; Rack, B.; Friese, K.; Janni, W.; Jeschke, U.; Andergassen, U.; Trapp, E.; Jückstock, J.; Jäger, B.; et al. Determination of Interleukin-4, 5, 6, 8 and 13 in serum of patients with breast cancer before treatment and its correlation to circulating tumor cells. Anticancer. Res. 2016, 36, 3123–3130. [Google Scholar]
- Lin, C.; Liu, H.; Zhang, H.; He, H.; Li, H.; Shen, Z.; Qin, J.; Qin, X.; Xu, J.; Sun, Y. Interleukin-13 receptor α2 is associated with poor prognosis in patients with gastric cancer after gastrectomy. Oncotarget 2016, 7, 49281–49288. [Google Scholar] [CrossRef] [Green Version]
- Mondello, P.; Cuzzocrea, S.; Arrigo, C.; Pitini, V.; Mian, M.; Bertoni, F. STAT6 activation correlates with cerebrospinal fluid IL-4 and IL-10 and poor prognosis in primary central nervous system lymphoma. Hematol. Oncol. 2020, 38, 106–110. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wang, Z.; Zhang, S. Correlation analysis of IL-4, IL-10 and APN levels with postoperative infection of colorectal cancer. Oncol. Lett. 2018, 17, 1603–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashidi, M.; Bazi, A.; Shiran, M.R.; Bagheri, A.; Mehrabadi, A.R.; Kalantar, H.; Ghafouri, Z.; Hosseini, S.M. Tropisetron attenuates tumor growth and progression in an experimental model of mouse lung cancer. J. Cell. Biochem. 2020, 121, 1610–1622. [Google Scholar] [CrossRef]
- Shirota, H.; Klinman, D.M.; Ito, S.-E.; Ito, H.; Kubo, M.; Ishioka, C. IL4 from T follicular helper cells downregulates antitumor immunity. Cancer Immunol. Res. 2016, 5, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Shen, W.; Yu, J.; Wang, L. TBX21 predicts prognosis of patients and drives cancer stem cell maintenance via the TBX21–IL-4 pathway in lung adenocarcinoma. Stem Cell Res. Ther. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Prevost-Blondel, A.; Richard, Y. Interleukin 4-Induced Gene 1 as an emerging regulator of B-cell biology and its role in cutaneous melanoma. Crit. Rev. Immunol. 2019, 39, 39–57. [Google Scholar] [CrossRef]
- Guo, C.; Ouyang, Y.; Cai, J.; Xiong, L.; Chen, Y.; Zeng, X.; Liu, A. High Expression of IL-4R enhances proliferation and invasion of hepatocellular carcinoma cells. Int. J. Biol. Markers 2017, 32, 384–390. [Google Scholar] [CrossRef]
- Hsi, L.C.; Kundu, S.; Palomo, J.; Xu, B.; Ficco, R.; Vogelbaum, M.A.; Cathcart, M.K. Silencing IL-13Ralpha2 promotes glioblastoma cell death via endogenous signaling. Mol. Cancer Ther. 2011, 10, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Zhang, L.; He, M.; Patterson, E.E.; Nilubol, N.; Fojo, A.T.; Joshi, B.; Puri, R.; Kebebew, E. Interleukin-13 receptor alpha2 is a novel therapeutic target for human adrenocortical carcinoma. Cancer 2012, 118, 5698–5708. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.J.; Li, B.H.; Yang, X.Z.; Li, P.D.; Yuan, Q.; Liu, X.H.; Xu, S.B.; Zhang, Y.; Yuan, J.; Gerhard, G.S.; et al. IL-4-induced Stat6 activities affect apoptosis and gene expression in breast cancer cells. Cytokine 2008, 42, 39–47. [Google Scholar] [CrossRef]
- Kreitman, R.J.; Puri, R.K.; Pastan, I. A circularly permuted recombinant interleukin 4 toxin with increased activity. Proc. Natl. Acad. Sci. USA 1994, 91, 6889–6893. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, K.; Puri, R.K.; Kawakami, M. Overexpressed cell surface interleukin-4 receptor molecules can be successfully targeted for antitumor cytotoxin therapy. Crit. Rev. Immunol. 2001, 21, 12. [Google Scholar] [CrossRef]
- Kawakami, M.; Kawakami, K.; A Stepensky, V.; A Maki, R.; Robin, H.; Muller, W.; Husain, S.R.; Puri, R.K. Interleukin 4 receptor on human lung cancer: A molecular target for cytotoxin therapy. Clin. Cancer Res. 2002, 8, 3503–3511. [Google Scholar] [PubMed]
- Kim, E.S.; Choi, Y.E.; Hwang, S.J.; Han, Y.-H.; Park, M.-J.; Bae, I.H. IL-4, a direct target of miR-340/429, is involved in radiation-induced aggressive tumor behavior in human carcinoma cells. Oncotarget 2016, 7, 86836–86856. [Google Scholar] [CrossRef] [Green Version]
- Garland, L.; Gitlitz, B.; Ebbinghaus, S.; Pan, H.; de Haan, H.; Puri, R.K.; Von Hoff, D.; Figlin, R. Phase I trial of intravenous IL-4 pseudomonas exotoxin protein (NBI-3001) in patients with advanced solid tumors that express the IL-4 receptor. J. Immunother. 2005, 28, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Balyasnikova, I.V.; Wainwright, D.A.; Solomaha, E.; Lee, G.; Han, Y.; Thaci, B.; Lesniak, M.S. Characterization and immunotherapeutic implications for a novel antibody targeting interleukin (IL)-13 receptor alpha2. J. Biol. Chem. 2012, 287, 30215–30227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenouchi, M.; Hirai, S.; Sakuragi, N.; Yagita, H.; Hamada, H.; Kato, K. Epigenetic modulation enhances the therapeutic effect of anti-IL-13R(alpha)2 antibody in human mesothelioma xenografts. Clin. Cancer Res. 2011, 17, 2819–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartolome, R.A.; Jaen, M.; Casal, J.I. An IL13Ralpha2 peptide exhibits therapeutic activity against metastatic colorectal cancer. Br. J. Cancer 2018, 119, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Natoli, A.; Lüpertz, R.; Merz, C.; Müller, W.W.; Köhler, R.; Krammer, P.H.; Li-Weber, M. Targeting the IL-4/IL-13 signaling pathway sensitizes Hodgkin lymphoma cells to chemotherapeutic drugs. Int. J. Cancer 2013, 133, 1945–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rand, R.W.; Kreitman, R.J.; Patronas, N.; Varricchio, F.; Pastan, I.; Puri, R.K. Intratumoral administration of recombinant circularly permuted interleukin-4-Pseudomonas exotoxin in patients with high-grade glioma. Clin. Cancer Res. 2000, 6, 2157–2165. [Google Scholar]
- Liu-Chittenden, Y.; Jain, M.; Kumar, P.; Patel, D.; Aufforth, R.; Neychev, V.; Sadowski, S.; Gara, S.K.; Joshi, B.H.; Cottle-Delisle, C.; et al. Phase I trial of systemic intravenous infusion of interleukin-13-Pseudomonas exotoxin in patients with metastatic adrenocortical carcinoma. Cancer Med. 2015, 4, 1060–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, J.; Song, X.; Traub, B.; Luxenhofer, M.; Kornmann, M. Involvement of IL-4, IL-13 and Their Receptors in Pancreatic Cancer. Int. J. Mol. Sci. 2021, 22, 2998. https://doi.org/10.3390/ijms22062998
Shi J, Song X, Traub B, Luxenhofer M, Kornmann M. Involvement of IL-4, IL-13 and Their Receptors in Pancreatic Cancer. International Journal of Molecular Sciences. 2021; 22(6):2998. https://doi.org/10.3390/ijms22062998
Chicago/Turabian StyleShi, Jingwei, Xujun Song, Benno Traub, Michael Luxenhofer, and Marko Kornmann. 2021. "Involvement of IL-4, IL-13 and Their Receptors in Pancreatic Cancer" International Journal of Molecular Sciences 22, no. 6: 2998. https://doi.org/10.3390/ijms22062998
APA StyleShi, J., Song, X., Traub, B., Luxenhofer, M., & Kornmann, M. (2021). Involvement of IL-4, IL-13 and Their Receptors in Pancreatic Cancer. International Journal of Molecular Sciences, 22(6), 2998. https://doi.org/10.3390/ijms22062998