
Putative LysM Effectors Contribute to Fungal Lifestyle

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
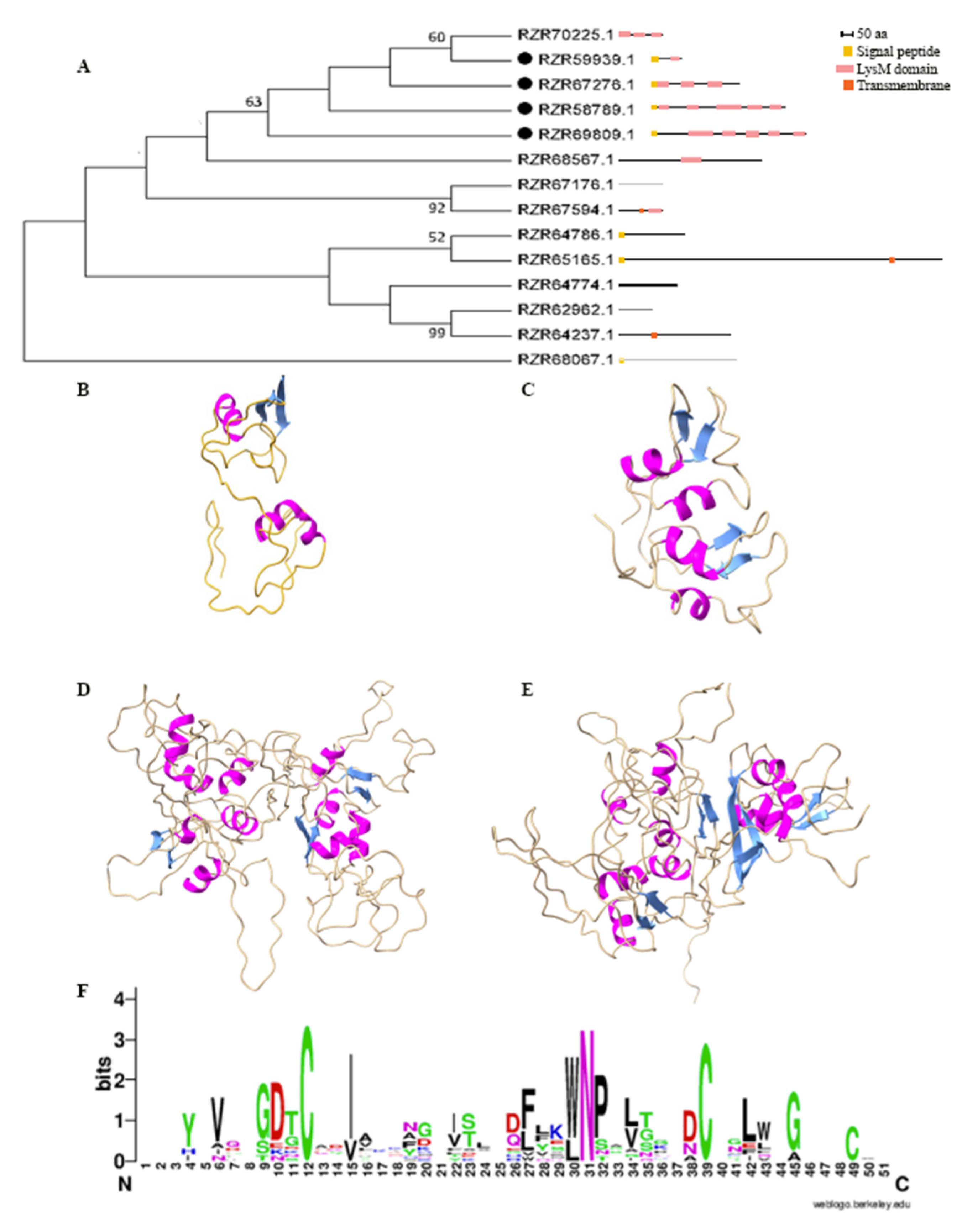
2.1. Pc123 Has Four Putative LysM Effectors
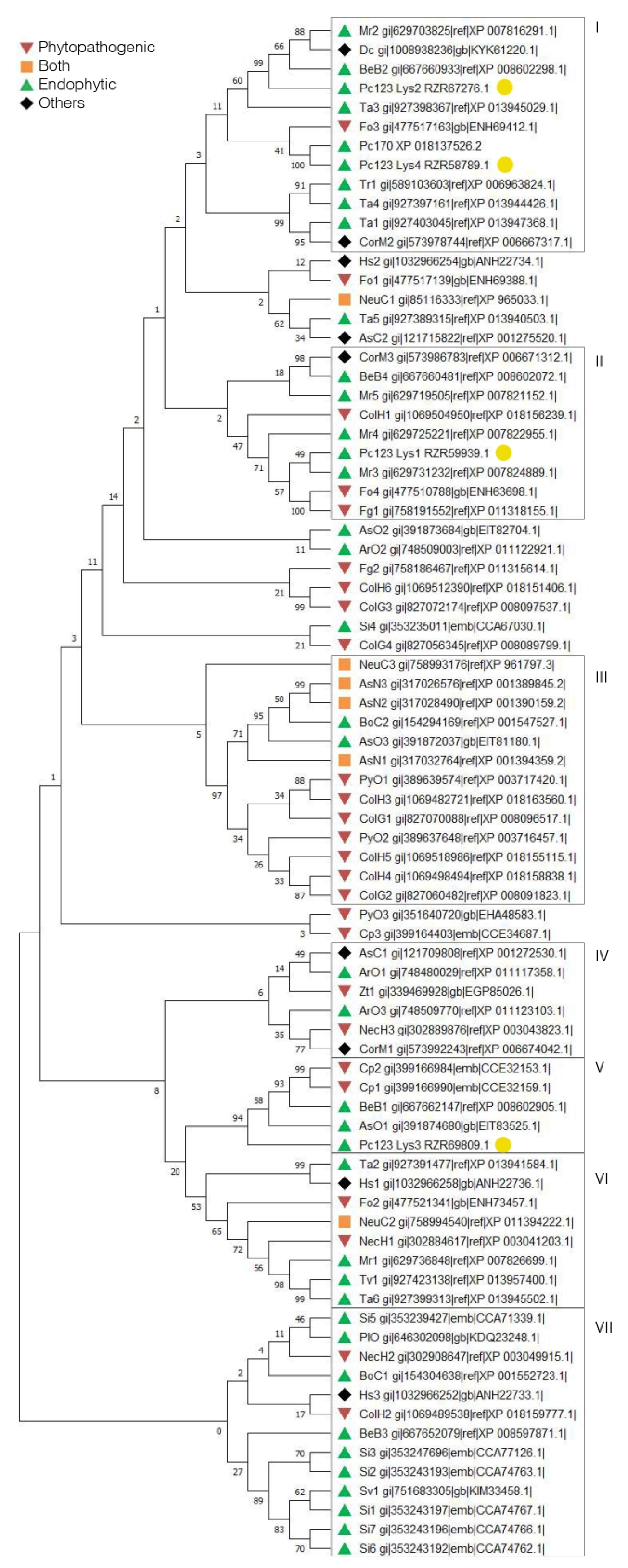
2.2. Putative LysM Effectors May Be Associated with the Lifestyle of a Fungus
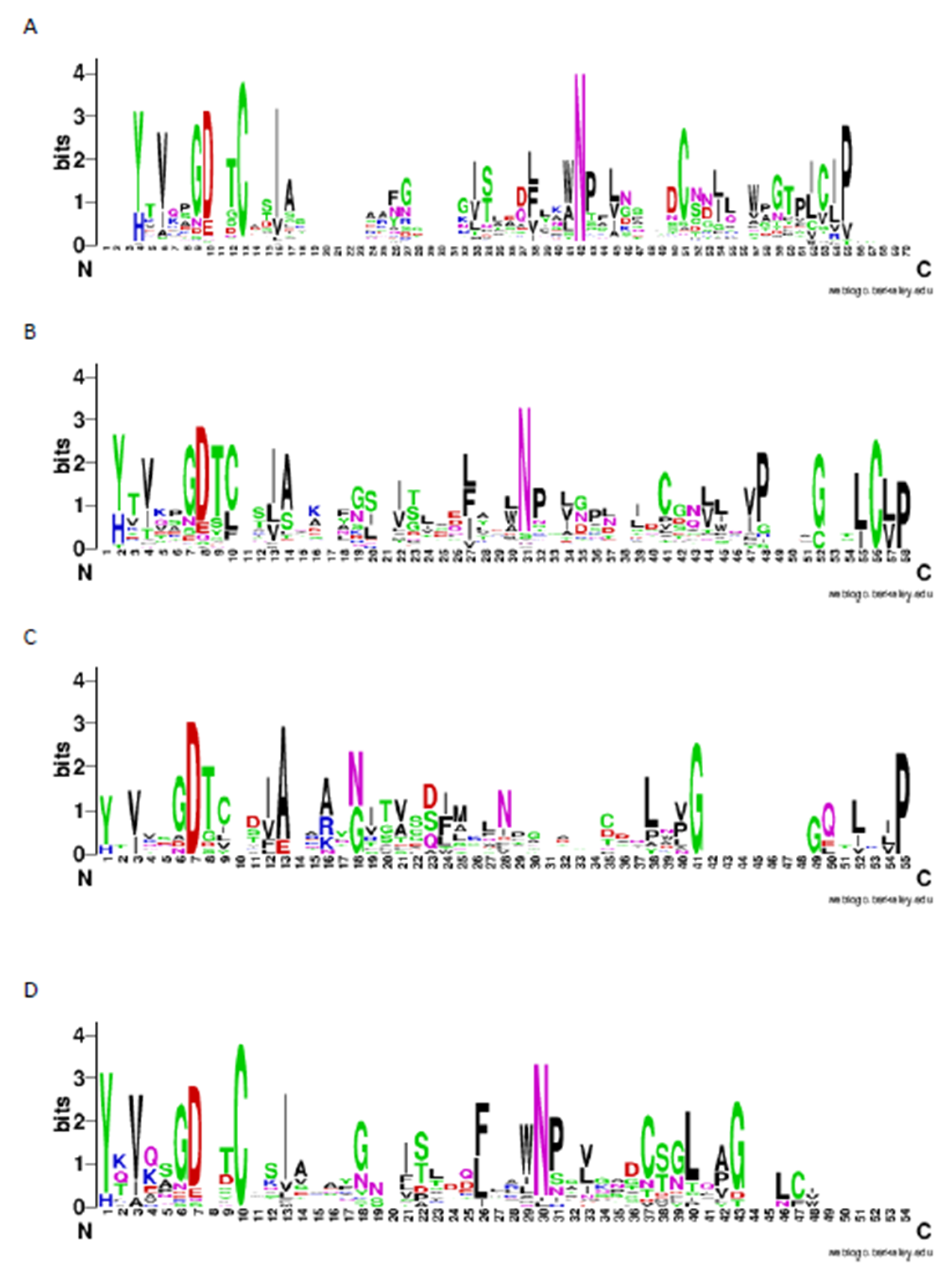
2.3. Patterns of LysM Motifs May Reflect Fungal Lifestyle
2.4. Molecular Docking
2.5. Genes Encoding Putative LysM Effectors Are Expressed
3. Discussion
4. Materials and Methods
4.1. Identification of Putative LysM Effectors
4.2. Identification of Protein Domains
4.3. Three-Dimensional Structures
4.4. Phylogenetic Analyses and Molecular Docking
4.5. qRT-PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gams, W.; Zare, R. A revision of Verticillium sect. Prostrata. III. Generic classification. Nov. Hedwig. 2001, 72, 329–337. [Google Scholar] [CrossRef]
- dos Santos, M.C.; Esteves, I.; Kerry, B.; Abrantes, I. Biology, growth parameters and enzymatic activity of Pochonia chlamydosporia isolated from potato cyst and root-knot nematodes. Nematology 2013, 15, 493–504. [Google Scholar] [CrossRef]
- Giné, A.; Bonmatí, M.; Sarro, A.; Stchiegel, A.; Valero, J.; Ornat, C.; Fernández, C.; Sorribas, F.J. Natural occurrence of fungal egg parasites of root-knot nematodes, Meloidogyne spp. in organic and integrated vegetable production systems in Spain. BioControl 2013, 58, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Kerry, B.R. Rhizosphere Interactions and the Exploitation of Microbial Agents for the Biological Control of Plant-Parasitic Nematodes. Annu. Rev. Phytopathol. 2000, 38, 423–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Llorca, L.V.; Olivares-Bernabeu, C.; Salinas, J.; Jansson, H.-B.; Kolattukudy, P.E. Pre-penetration events in fungal parasitism of nematode eggs. Mycol. Res. 2002, 106, 499–506. [Google Scholar] [CrossRef]
- Mauchline, T.H.; Kerry, B.R.; Hirsch, P.R. The biocontrol fungus Pochonia chlamydosporia shows nematode host preference at the infraspecific level. Mycol. Res. 2004, 108, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Monfort, E.; Lopez-Llorca, L.V.; Jansson, H.-B.; Salinas, J.; Park, J.O.; Sivasithamparam, K. Colonisation of seminal roots of wheat and barley by egg-parasitic nematophagous fungi and their effects on Gaeumannomyces graminis var. tritici and development of root-rot. Soil Biol. Biochem. 2005, 37, 1229–1235. [Google Scholar] [CrossRef]
- Maciá-Vicente, J.G.; Rosso, L.C.; Ciancio, A.; Jansson, H.-B.; Lopez-Llorca, L.V. Colonisation of barley roots by endophytic Fusarium equiseti and Pochonia chlamydosporia: Effects on plant growth and disease. Ann. Appl. Biol. 2009, 155, 391–401. [Google Scholar] [CrossRef]
- Escudero, N.; Lopez-Llorca, L.V. Effects on plant growth and root-knot nematode infection of an endophytic GFP transformant of the nematophagous fungus Pochonia chlamydosporia. Symbiosis 2012, 57, 33–42. [Google Scholar] [CrossRef]
- Zavala-Gonzalez, E.A.; Escudero, N.; Lopez-Moya, F.; Aranda-Martinez, A.; Exposito, A.; Ricaño-Rodríguez, J.; Naranjo-Ortiz, M.A.; Ramírez-Lepe, M.; Lopez-Llorca, L.V. Some isolates of the nematophagous fungus Pochonia chlamydosporia promote root growth and reduce flowering time of tomato. Ann. Appl. Biol. 2015, 166, 472–483. [Google Scholar] [CrossRef]
- Larriba, E.; Jaime, M.D.L.A.; Nislow, C.; Martín-Nieto, J.; Lopez-Llorca, L.V. Endophytic colonization of barley (Hordeum vulgare) roots by the nematophagous fungus Pochonia chlamydosporia reveals plant growth promotion and a general defense and stress transcriptomic response. J. Plant Res. 2015, 128, 665–678. [Google Scholar] [CrossRef]
- Ciancio, A.; Pentimone, I.; Colagiero, M.; Rosso, L. Regulatory Factors in Pochonia chlamydosporia-Induced Gene Expression. In Perspectives in Sustainable Nematode Management through Pochonia chlamydosporia Applications for Root and Rhizosphere Health; Manzanilla-López, R.H., Lopez-Llorca, L.V., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 99–109. ISBN1 978-3-319-59222-0. ISBN2 978-3-319-59224-4. [Google Scholar]
- Zavala-Gonzalez, E.A.; Rodríguez-Cazorla, E.; Escudero, N.; Aranda-Martinez, A.; Martínez-Laborda, A.; Ramírez-Lepe, M.; Vera, A.; Lopez-Llorca, L.V. Arabidopsis thaliana root colonization by the nematophagous fungus Pochonia chlamydosporia is modulated by jasmonate signaling and leads to accelerated flowering and improved yield. New Phytol. 2017, 213, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L. Molecular Mechanisms of Chitosan Interactions with Fungi and Plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.-Q.; Wang, F.-Z.; Li, J.-F. Hide-and-Seek: Chitin-Triggered Plant Immunity and Fungal Counterstrategies. Trends Plant Sci. 2020, 25, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P.H.J. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef] [Green Version]
- Kombrink, A.; Thomma, B.P.H.J. LysM Effectors: Secreted Proteins Supporting Fungal Life. PLoS Pathog. 2013, 9, e1003769. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vallet, A.; Saleem-Batcha, R.; Kombrink, A.; Hansen, G.; Valkenburg, D.-J.; Thomma, B.P.H.J.; Mesters, J.R. Fungal effector Ecp6 outcompetes host immune receptor for chitin binding through intrachain LysM dimerization. eLife 2013, 2, e00790. [Google Scholar] [CrossRef]
- Yin, H.; Du, Y.; Dong, Z. Chitin Oligosaccharide and Chitosan Oligosaccharide: Two Similar but Different Plant Elicitors. Front. Plant Sci. 2016, 7, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, R.; Kombrink, A.; Motteram, J.; Loza-Reyes, E.; Lucas, J.; Hammond-Kosack, K.E.; Thomma, B.P.H.J.; Rudd, J.J. Analysis of two in planta expressed LysM effector homologs from the fungus Mycosphaerella graminicola reveals novel functional properties and varying contributions to virulence on wheat. Plant Physiol. 2011, 156, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Cen, K.; Li, B.; Lu, Y.; Zhang, S.; Wang, C. Divergent LysM effectors contribute to the virulence of Beauveria bassiana by evasion of insect immune defenses. PLoS Pathog. 2017, 13, e1006604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kombrink, A.; Rovenich, H.; Shi-Kunne, X.; Rojas-Padilla, E.; van den Berg, G.C.M.; Domazakis, E.; de Jonge, R.; Valkenburg, D.-J.; Sánchez-Vallet, A.; Seidl, M.F.; et al. Verticillium dahliae LysM effectors differentially contribute to virulence on plant hosts: Verticillium dahliae LysM effectors. Mol. Plant Pathol. 2017, 18, 596–608. [Google Scholar] [CrossRef] [Green Version]
- Romero-Contreras, Y.J.; Ramírez-Valdespino, C.A.; Guzmán-Guzmán, P.; Macías-Segoviano, J.I.; Villagómez-Castro, J.C.; Olmedo-Monfil, V. Tal6 from Trichoderma atroviride is a LysM effector involved in mycoparasitism and plant association. Front. Microbiol. 2019, 10, 2231. [Google Scholar] [CrossRef] [PubMed]
- Akcapinar, G.B.; Kappel, L.; Sezerman, O.U.; Seidl-Seiboth, V. Molecular diversity of LysM carbohydrate-binding motifs in fungi. Curr. Genet. 2015, 61, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Buist, G.; Steen, A.; Kok, J.; Kuipers, O.P. LysM, a widely distributed protein motif for binding to (peptido) glycans. Mol. Microbiol. 2008, 68, 838–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kombrink, A.; Sánchez-Vallet, A.; Thomma, B.P.H.J. The role of chitin detection in plant–pathogen interactions. Microbes Infect. 2011, 13, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Bycroft, M. The structure of a LysM domain from E. coli membrane-bound lytic murein transglycosylase D (MltD). J. Mol. Biol. 2000, 299, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Bielnicki, J.; Devedjiev, Y.; Derewenda, U.; Dauter, Z.; Joachimiak, A.; Derewenda, Z.S. B. subtilis YkuD protein at 2.0 Å resolution: Insights into the structure and function of a novel, ubiquitous family of bacterial enzymes. Proteins Struct. Funct. Bioinform. 2006, 62, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnuma, T.; Onaga, S.; Murata, K.; Taira, T.; Katoh, E. LysM Domains from Pteris ryukyuensis Chitinase-A: A stability study and characterization of the chitin-binding site. J. Biol. Chem. 2008, 283, 5178–5187. [Google Scholar] [CrossRef] [Green Version]
- Miya, A.; Albert, P.; Shinya, T.; Desaki, Y.; Ichimura, K.; Shirasu, K.; Narusaka, Y.; Kawakami, N.; Kaku, H.; Shibuya, N. CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 19613–19618. [Google Scholar] [CrossRef] [Green Version]
- Levin, E.; Ballester, A.R.; Raphael, G.; Feigenberg, O.; Liu, Y.; Norelli, J.; Gonzalez-Candelas, L.; Ma, J.; Dardick, C.; Wisniewski, M.; et al. Identification and characterization of LysM effectors in Penicillium expansum. PLoS ONE 2017, 12, e0186023. [Google Scholar] [CrossRef] [Green Version]
- Larriba, E.; Jaime, M.D.L.A.; Carbonell-Caballero, J.; Conesa, A.; Dopazo, J.; Nislow, C.; Martín-Nieto, J.; Lopez-Llorca, L.V. Sequencing and functional analysis of the genome of a nematode egg-parasitic fungus, Pochonia chlamydosporia. Fungal Genet. Biol. 2014, 65, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi; Reprint de.; IHW-Verl: Eching, Germany, 1993; ISBN 978-3-9803083-8-0. [Google Scholar]
- Zare, R.; Gams, W.; Evans, H.C. A revision of Verticillium section Prostrata. V. The genus Pochonia, with notes on Rotiferophthora. Nova Hedwigia 2001, 73, 51–86. [Google Scholar] [CrossRef]
- Mesnage, S.; Dellarole, M.; Baxter, N.J.; Rouget, J.-B.; Dimitrov, J.D.; Wang, N.; Fujimoto, Y.; Hounslow, A.M.; Lacroix-Desmazes, S.; Fukase, K.; et al. Molecular basis for bacterial peptidoglycan recognition by LysM domains. Nat. Commun. 2014, 5, 4269. [Google Scholar] [CrossRef] [PubMed]
- Onaga, S.; Taira, T. A new type of plant chitinase containing LysM domains from a fern (Pteris ryukyuensis): Roles of LysM domains in chitin binding and antifungal activity. Glycobiology 2008, 18, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Qin, F.; Shen, B.; Shi, Q.; Liu, C.; Zhang, X.; Jiao, Y.; Lu, J.; Gao, Y.; Suarez-Fernandez, M.; et al. Genome and secretome analysis of Pochonia chlamydosporia provide new insight into egg-parasitic mechanisms. Sci. Rep. 2018, 8, 1123. [Google Scholar] [CrossRef]
- Olivares, C.M.; López-Llorca, L.V. Fungal egg-parasites of plant-parasitic nematodes from Spanish soils. Rev. Iberoam. Micol. 2002, 19, 104–110. [Google Scholar]
- Barron, G.L. The Nematode-destroying Fungi; Topics in Mycobiology; Canadian Biological Publications: Guelph, ON, Canada, 1977; ISBN 978-0-920370-00-1. [Google Scholar]
- Lebrigand, K.; He, L.D.; Thakur, N.; Arguel, M.-J.; Polanowska, J.; Henrissat, B.; Record, E.; Magdelenat, G.; Barbe, V.; Raffaele, S.; et al. Comparative Genomic Analysis of Drechmeria coniospora Reveals Core and Specific Genetic Requirements for Fungal Endoparasitism of Nematodes. PLoS Genet. 2016, 12, e1006017. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Llorca, L.V.; Jansson, H.-B.; Vicente, J.G.M.; Salinas, J. Nematophagous Fungi as Root Endophytes. In Microbial Root Endophytes; Schulz, B.J.E., Boyle, C.J.C., Sieber, T.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 9, pp. 191–206. ISBN 978-3-540-33525-2. [Google Scholar]
- Nordbring-Hertz, B.; Jansson, H.-B.; Tunlid, A. Nematophagous Fungi. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2001; p. a0000374. ISBN 978-0-470-01590-2. [Google Scholar]
- Zhang, Y.; Qiao, M.; Xu, J.; Cao, Y.; Zhang, K.-Q.; Yu, Z.-F. Genetic diversity and recombination in natural populations of the nematode-trapping fungus Arthrobotrys oligospora from China. Ecol. Evol. 2013, 3, 312–325. [Google Scholar] [CrossRef]
- Bharagava, R.N.; Chowdhary, P. Emerging and Eco-Friendly Approaches for Waste Management; Springer: Berlin/Heidelberg, Germany, 2018; ISBN 978-981-10-8669-4. [Google Scholar]
- Sasan, R.K.; Bidochka, M.J. The insect-pathogenic fungus Metarhizium robertsii (Clavicipitaceae) is also an endophyte that stimulates plant root development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Baek, J.-M.; Howell, C.R.; Kenerley, C.M. The role of an extracellular chitinase from Trichoderma virens Gv29-8 in the biocontrol of Rhizoctonia solani. Curr. Genet. 1999, 35, 41–50. [Google Scholar] [CrossRef]
- Onofre, S.B.; Bonfante, T.; dos Santos, Z.M.Q.; Moura, M.C.; Cardoso, A.F. Cellulase Production by Endophytic Strains of Trichoderma reesei from Baccharis dracunculifolia; D. C. (Asteraceae). Adv. Microbiol. 2014, 04, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. The root endophytic fungus Trichoderma atroviride induces foliar herbivory resistance in maize plants. Appl. Soil Ecol. 2018, 124, 45–53. [Google Scholar] [CrossRef]
- Suarez-Fernandez, M.; Sambles, C.; Lopez-Moya, F.; Nueda, M.J.; Studholme, D.J.; Lopez-Llorca, L.V. Chitosan modulates Pochonia chlamydosporia gene expression during nematode egg parasitism. Environ. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- del Barrio-Duque, A.; Ley, J.; Samad, A.; Antonielli, L.; Sessitsch, A.; Compant, S. Beneficial Endophytic Bacteria-Serendipita indica Interaction for Crop Enhancement and Resistance to Phytopathogens. Front. Microbiol. 2019, 10, 2888. [Google Scholar] [CrossRef] [PubMed]
- De-la-Peña, C.; Badri, D.V.; Lei, Z.; Watson, B.S.; Brandão, M.M.; Silva-Filho, M.C.; Sumner, L.W.; Vivanco, J.M. Root Secretion of Defense-related Proteins Is Development-dependent and Correlated with Flowering Time. J. Biol. Chem. 2010, 285, 30654–30665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.-T.; Akutse, K.S.; Xia, X.-F.; Chen, J.-H.; Ai, X.; Tang, Y.; Wang, Q.; Feng, B.-W.; Goettel, M.S.; You, M.-S. Endophytic effects of Aspergillus oryzae on radish (Raphanus sativus) and its herbivore, Plutella xylostella. Planta 2018, 248, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Elfita, E.; Muharni, M.; Munawar, M.; Aryani, S. Secondary metabolite from endophytic fungi Aspergillus niger of the stem bark of kandis gajah (Garcinia griffithii). Indones. J. Chem. 2012, 12, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.-C.; Hui, S.; Choi, J.; Asiegbu, F.O.; Valkonen, J.P.T.; Lee, Y.-H. Secret lifestyles of Neurospora crassa. Sci. Rep. 2015, 4, 5135. [Google Scholar] [CrossRef]
- Pawar, N.V.; Patil, V.B.; Kamble, S.S.; Dixit, G.B. First Report of Aspergillus niger as a Plant Pathogen on Zingiber officinale from India. Plant Dis. 2008, 92, 1368. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef] [Green Version]
- de Jonge, R.; Thomma, B.P.H.J. Fungal LysM effectors: Extinguishers of host immunity? Trends Microbiol. 2009, 17, 151–157. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Sillitoe, I.; Dawson, N.; Lewis, T.E.; Das, S.; Lees, J.G.; Ashford, P.; Tolulope, A.; Scholes, H.M.; Senatorov, I.; Bujan, A.; et al. CATH: Expanding the horizons of structure-based functional annotations for genome sequences. Nucleic Acids Res. 2018, 47, D280–D284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haft, D.H.; Selengut, J.D.; Richter, R.A.; Harkins, D.; Basu, M.K.; Beck, E. TIGRFAMs and Genome Properties in 2013. Nucleic Acids Res. 2012, 41, D387–D395. [Google Scholar] [CrossRef] [Green Version]
- Pandurangan, A.P.; Stahlhacke, J.; Oates, M.E.; Smithers, B.; Gough, J. The SUPERFAMILY 2.0 database: A significant proteome update and a new webserver. Nucleic Acids Res. 2018, 47, D490–D494. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Nikolskaya, A.; Huang, H.; Yeh, L.L.; Natale, D.A.; Vinayaka, C.R.; Hu, Z.; Mazumder, R.; Kumar, S.; Kourtesis, P.; et al. PIRSF: Family classification system at the Protein Information Resource. Nucleic Acids Res. 2004, 32, D112–D114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, F.; Patricio, M.; Muffato, M.; Pignatelli, M.; Bateman, A. TreeFam v9: A new website, more species and orthology-on-the-fly. Nucleic Acids Res. 2013, 42, D922–D925. [Google Scholar] [CrossRef] [Green Version]
- Fiser, A.; Sali, A. Modeller: Generation and refinement of homology-based protein structure models. Methods Enzymol. 2003, 374, 461–491. [Google Scholar] [CrossRef]
- Ohnuma, T.; Taira, T.; Umemoto, N.; Kitaoku, Y.; Sørlie, M.; Numata, T.; Fukamizo, T. Crystal structure and thermodynamic dissection of chitin oligosaccharide binding to the LysM module of chitinase-A from Pteris ryukyuensis. Biochem. Biophys. Res. Commun. 2017, 494, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Inamine, S.; Onaga, S.; Ohnuma, T.; Fukamizo, T.; Taira, T. Purification, cDNA cloning, and characterization of LysM-containing plant chitinase from horsetail (Equisetum arvense). Biosci. Biotechnol. Biochem. 2015, 79, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Liu, Y.; Wu, G.; Zeng, S.; Tran Thi, T.G.; Liang, L.; Liang, Y.; Dong, Z.; She, D.; Wang, H.; et al. Identification of a Candidate Gene for Panicle Length in Rice (Oryza sativa L.) Via Association and Linkage Analysis. Front. Plant Sci. 2016, 7, 596. [Google Scholar] [CrossRef] [Green Version]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [Green Version]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C.; Word, M.J.; De Bakker, P. Structure validation by Cα geometry: ϕ,ψ and Cβ deviation. Proteins Struct. Funct. Bioinform. 2003, 50, 437–450. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, E.; Kerry, B.R.; Manzanilla-López, R.H.; Mutua, G.; Devonshire, J.; Kimenju, J.; Hirsch, P.R. The Pochonia chlamydosporia Serine Protease Gene vcp1 is Subject to Regulation by Carbon, Nitrogen and pH: Implications for Nematode Biocontrol. PLoS ONE 2012, 7, e35657. [Google Scholar] [CrossRef] [Green Version]
- Aranda-Martinez, A.; Lenfant, N.; Escudero, N.; Zavala-Gonzalez, E.A.; Henrissat, B.; Lopez-Llorca, L.V. CAZyme content of Pochonia chlamydosporia reflects that chitin and chitosan modification are involved in nematode parasitism: CAZome of Pochonia chlamydosporia. Environ. Microbiol. 2016, 18, 4200–4215. [Google Scholar] [CrossRef] [Green Version]
- Rosso, L.C.; Colagiero, M.; Salatino, N.; Ciancio, A. Observations on the effect of trophic conditions on Pochonia chlamydosporia gene expression. Ann. Appl. Biol. 2014, 164, 232–243. [Google Scholar] [CrossRef]
- Escudero, N.; Ferreira, S.R.; Lopez-Moya, F.; Naranjo-Ortiz, M.A.; Marin-Ortiz, A.I.; Thornton, C.R.; Lopez-Llorca, L.V. Chitosan enhances parasitism of Meloidogyne javanica eggs by the nematophagous fungus Pochonia chlamydosporia. Fungal Biol. 2016, 120, 572–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Sequence Name | Length | %Cys | LysM Domains | Name Putative Effector |
|---|---|---|---|---|---|
| Pochonia chlamydosporia 123 | gi|1576959602|gb|RZR59939.1| | 123 | 4.88 | 1 | Pc123_Lys-1 |
| gi|1576967440|gb|RZR67276.1| | 377 | 5.57 | 3 | Pc123_Lys-2 | |
| gi|1576970205|gb|RZR69809.1| | 665 | 4.96 | 6 | Pc123_Lys-3 | |
| gi|1576958349|gb|RZR58789.1| | 577 | 3.81 | 5 | Pc123_Lys-4 | |
| Pochonia chlamydosporia 170 | gi|1240655211|ref|XP_018137526.2| | 601 | 4.16 | 5 | Pc170-1 |
| gi|1069526848|ref|XP_018144528.1| | 123 | 4.88 | 1 | Pc170-2 | |
| Arthrobotrys oligospora ATCC 24927 | gi|748480029|ref|XP_011117358.1| | 677 | 5.02 | 6 | ArO-1 |
| gi|748509003|ref|XP_011122921.1| | 387 | 4.13 | 4 | ArO-2 | |
| gi|748509770|ref|XP_011123103.1| | 701 | 4.71 | 5 | ArO-3 | |
| Aspergillus clavatus NRRL 1 | gi|121709808|ref|XP_001272530.1| | 527 | 4.93 | 5 | AsC-1 |
| gi|121715822|ref|XP_001275520.1| | 343 | 4.66 | 4 | AsC-2 | |
| Aspergillus niger CBS 513.88 | gi|317032764|ref|XP_001394359.2| | 235 | 2.55 | 4 | AsN-1 |
| gi|317028490|ref|XP_001390159.2| | 223 | 3.59 | 3 | AsN-2 | |
| gi|317026576|ref|XP_001389845.2| | 228 | 3.51 | 3 | AsN-3 | |
| Aspergillus oryzae 3.042 | gi|391874680|gb|EIT83525.1| | 488 | 3.89 | 2 | AsO-1 |
| gi|391873684|gb|EIT82704.1| | 400 | 3.50 | 2 | AsO-2 | |
| gi|391872037|gb|EIT81180.1| | 228 | 3.51 | 3 | AsO-3 | |
| Beauveria bassiana ARSEF 2860 | gi|667662147|ref|XP_008602905.1| | 563 | 4.80 | 4 | BeB-1 |
| gi|667660933|ref|XP_008602298.1| | 384 | 5.73 | 3 | BeB-2 | |
| gi|667652079|ref|XP_008597871.1| | 258 | 2.33 | 2 | BeB-3 | |
| gi|667660481|ref|XP_008602072.1| | 167 | 7.19 | 2 | BeB-4 | |
| Botryotinia fuckeliana = Botrytis cinerea B05.10 | gi|154304638|ref|XP_001552723.1| | 239 | 1.67 | 1 | BoC-1 |
| gi|154294169|ref|XP_001547527.1| | 227 | 3.52 | 4 | BoC-2 | |
| Colletotrichum graminicola M1.001 | gi|827070088|ref|XP_008096517.1| | 153 | 4.58 | 2 | ColG-1 |
| gi|827060482|ref|XP_008091823.1| | 154 | 3.90 | 2 | ColG-2 | |
| gi|827072174|ref|XP_008097537.1| | 96 | 4.17 | 1 | ColG-3 | |
| gi|827056345|ref|XP_008089799.1| | 262 | 4.58 | 2 | ColG-4 | |
| Colletotrichum higginsianum IMI 349063 | gi|1069504950|ref|XP_018156239.1| | 686 | 4.52 | 4 | ColH-1 |
| gi|1069489538|ref|XP_018159777.1| | 335 | 5.37 | 2 | ColH-2 | |
| gi|1069482721|ref|XP_018163560.1| | 170 | 3.53 | 2 | ColH-3 | |
| gi|1069498494|ref|XP_018158838.1| | 176 | 3.98 | 2 | ColH-4 | |
| gi|1069518986|ref|XP_018155115.1| | 164 | 1.22 | 1 | ColH-5 | |
| gi|1069512390|ref|XP_018151406.1| | 93 | 4.30 | 1 | ColH-6 | |
| Cordyceps militaris CM01 | gi|573992243|ref|XP_006674042.1| | 455 | 4.84 | 3 | CorM-1 |
| gi|573978744|ref|XP_006667317.1| | 541 | 4.25 | 6 | CorM-2 | |
| gi|573986783|ref|XP_006671312.1| | 187 | 6.95 | 2 | CorM-3 | |
| Drechmeria coniospora | gi|1008938236|gb|KYK61220.1| | 407 | 5.90 | 4 | Dc-1 |
| Fusarium graminearum PH-1 | gi|758191552|ref|XP_011318155.1| | 221 | 4.52 | 3 | Fg-1 |
| gi|758186467|ref|XP_011315614.1| | 178 | 6.18 | 2 | Fg-2 | |
| Fusarium oxysporum (FOCTR1) | gi|477517139|gb|ENH69388.1| | 298 | 4.03 | 3 | Fo-1 |
| gi|477521341|gb|ENH73457.1| | 423 | 5.67 | 3 | Fo-2 | |
| gi|477517163|gb|ENH69412.1| | 265 | 4.53 | 3 | Fo-3 | |
| gi|477510788|gb|ENH63698.1| | 218 | 4.59 | 2 | Fo-4 | |
| Metarhizium anisopliae = Metarhizium robertsii ARSEF 23 | gi|629736848|ref|XP_007826699.1| | 588 | 4.42 | 2 | Mr-1 |
| gi|629703825|ref|XP_007816291.1| | 403 | 5.46 | 4 | Mr-2 | |
| gi|629731232|ref|XP_007824889.1| | 125 | 4.80 | 2 | Mr-3 | |
| gi|629725221|ref|XP_007822955.1| | 127 | 4.72 | 1 | Mr-4 | |
| gi|629719505|ref|XP_007821152.1| | 175 | 5.14 | 2 | Mr-5 | |
| Fusarium solani = Nectria hematococca mpVI77-13-4 | gi|302884617|ref|XP_003041203.1| | 434 | 5.07 | 3 | NecH-1 |
| gi|302908647|ref|XP_003049915.1| | 354 | 5.37 | 2 | NecH-2 | |
| gi|302889876|ref|XP_003043823.1| | 453 | 4.86 | 2 | NecH-3 | |
| Neurospora crassa OR74A | gi|85116333|ref|XP_965033.1| | 265 | 4.53 | 3 | NeuC-1 |
| gi|758994540|ref|XP_011394222.1| | 460 | 4.78 | 2 | NeuC-2 | |
| gi|758993176|ref|XP_961797.3| | 540 | 2.22 | 3 | NeuC-3 | |
| Magnaporthe oryzae = Pyricularia oryzae 70–15 | gi|389639574|ref|XP_003717420.1| | 162 | 3.70 | 2 | PyO-1 |
| gi|389637648|ref|XP_003716457.1| | 285 | 2.11 | 2 | PyO-2 | |
| gi|351640720|gb|EHA48583.1| | 276 | 4.71 | 1 | PyO-3 | |
| Piriformospora indica = Serendipita indica | gi|353243197|emb|CCA74767.1| | 527 | 3.80 | 5 | Si-1 |
| gi|353243193|emb|CCA74763.1| | 418 | 5.74 | 4 | Si-2 | |
| gi|353247696|emb|CCA77126.1| | 163 | 4.29 | 2 | Si-3 | |
| gi|353235011|emb|CCA67030.1| | 171 | 4.68 | 1 | Si-4 | |
| gi|353239427|emb|CCA71339.1| | 174 | 5.17 | 2 | Si-5 | |
| gi|353243192|emb|CCA74762.1| | 654 | 4.59 | 8 | Si-6 | |
| gi|353243196|emb|CCA74766.1| | 361 | 4.43 | 4 | Si-7 | |
| Pleurotus ostreatus | gi|646302098|gb|KDQ23248.1| | 133 | 6.77 | 2 | PlO-1 |
| Trichoderma atroviride IMI 206040 | gi|927403045|ref|XP_013947368.1| | 746 | 4.29 | 5 | Ta-1 |
| gi|927391477|ref|XP_013941584.1| | 544 | 4.78 | 2 | Ta-2 | |
| gi|927398367|ref|XP_013945029.1| | 345 | 4.64 | 3 | Ta-3 | |
| gi|927397161|ref|XP_013944426.1| | 511 | 4.11 | 3 | Ta-4 | |
| gi|927389315|ref|XP_013940503.1| | 241 | 5.39 | 3 | Ta-5 | |
| gi|927399313|ref|XP_013945502.1| | 443 | 4.97 | 2 | Ta-6 | |
| Trichoderma reesei QM6a | gi|589103603|ref|XP_006963824.1| | 473 | 4.44 | 4 | Tr-1 |
| Trichoderma virens Gv29-8 | gi|927423138|ref|XP_013957400.1| | 440 | 5.00 | 2 | Tv-1 |
| Serendipita vermifera MAFF 305830 | gi|751683305|gb|KIM33458.1| | 141 | 4.96 | 2 | Sv-1 |
| Zymoseptoria tritici IPO323 | gi|339469928|gb|EGP85026.1| | 97 | 4.12 | 1 | Zt-1 |
| Hypocrella siamensis | gi|1032966258|gb|ANH22736.1| | 312 | 4.81 | 2 | Hs-1 |
| gi|1032966254|gb|ANH22734.1| | 323 | 3.72 | 3 | Hs-2 | |
| gi|1032966252|gb|ANH22733.1| | 86 | 4.65 | 1 | Hs-3 | |
| Claviceps purpurea 20.1 | gi|399166990|emb|CCE32159.1| | 689 | 5.22 | 6 | Cp-1 |
| gi|399166984|emb|CCE32153.1| | 688 | 5.23 | 5 | Cp-2 | |
| gi|399164403|emb|CCE34687.1| | 90 | 2.22 | 1 | Cp-3 |
| Name | Bind.Energy [kcal/mol] | Dissoc. Constant [pM] | Contacting Receptor Residues |
|---|---|---|---|
| Pc123 Lys1 | >−5.9760 | >41,642,224.00 | >GLN 13 LEU 14 THR 15 ALA 16 VAL 17 VAL 18 LYS 98 TRP 99 PRO 101 GLY 102 |
| >Pc123 Lys2 | >−6.3100 | >23,697,836.00 | >CYS 83 GLY 84 ASN 85 THR 111 THR 112 SER 113 GLN 114 LYS 115 LEU 157 GLN 218 CYS 269 THR 270 GLY 271 PHE 288 ASP 289 THR 290 GLN 309 |
| >Pc123 Lys3 | >−5.6040 | >78,021,360.00 | >VAL 428 THR 429 THR 590 ASN 591 THR 596 ALA 597 THR 598 GLY 601 GLY 602 PRO 604 |
| Pc123 Lys4 | −6.4030 | 20,255,304.00 | LEU 283 GLN 284 TYR 361 GLN 362 THR 492 ILE 493 GLN 494 THR 495 SER 497 PRO 498 ILE 499 MET 500 PRO 501 GLY 502 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suarez-Fernandez, M.; Aragon-Perez, A.; Lopez-Llorca, L.V.; Lopez-Moya, F. Putative LysM Effectors Contribute to Fungal Lifestyle. Int. J. Mol. Sci. 2021, 22, 3147. https://doi.org/10.3390/ijms22063147
Suarez-Fernandez M, Aragon-Perez A, Lopez-Llorca LV, Lopez-Moya F. Putative LysM Effectors Contribute to Fungal Lifestyle. International Journal of Molecular Sciences. 2021; 22(6):3147. https://doi.org/10.3390/ijms22063147
Chicago/Turabian StyleSuarez-Fernandez, Marta, Ana Aragon-Perez, Luis Vicente Lopez-Llorca, and Federico Lopez-Moya. 2021. "Putative LysM Effectors Contribute to Fungal Lifestyle" International Journal of Molecular Sciences 22, no. 6: 3147. https://doi.org/10.3390/ijms22063147
APA StyleSuarez-Fernandez, M., Aragon-Perez, A., Lopez-Llorca, L. V., & Lopez-Moya, F. (2021). Putative LysM Effectors Contribute to Fungal Lifestyle. International Journal of Molecular Sciences, 22(6), 3147. https://doi.org/10.3390/ijms22063147