
Intense Pulsed Light Attenuates UV-Induced Hyperimmune Response and Pigmentation in Human Skin Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. IPL Device Outputs
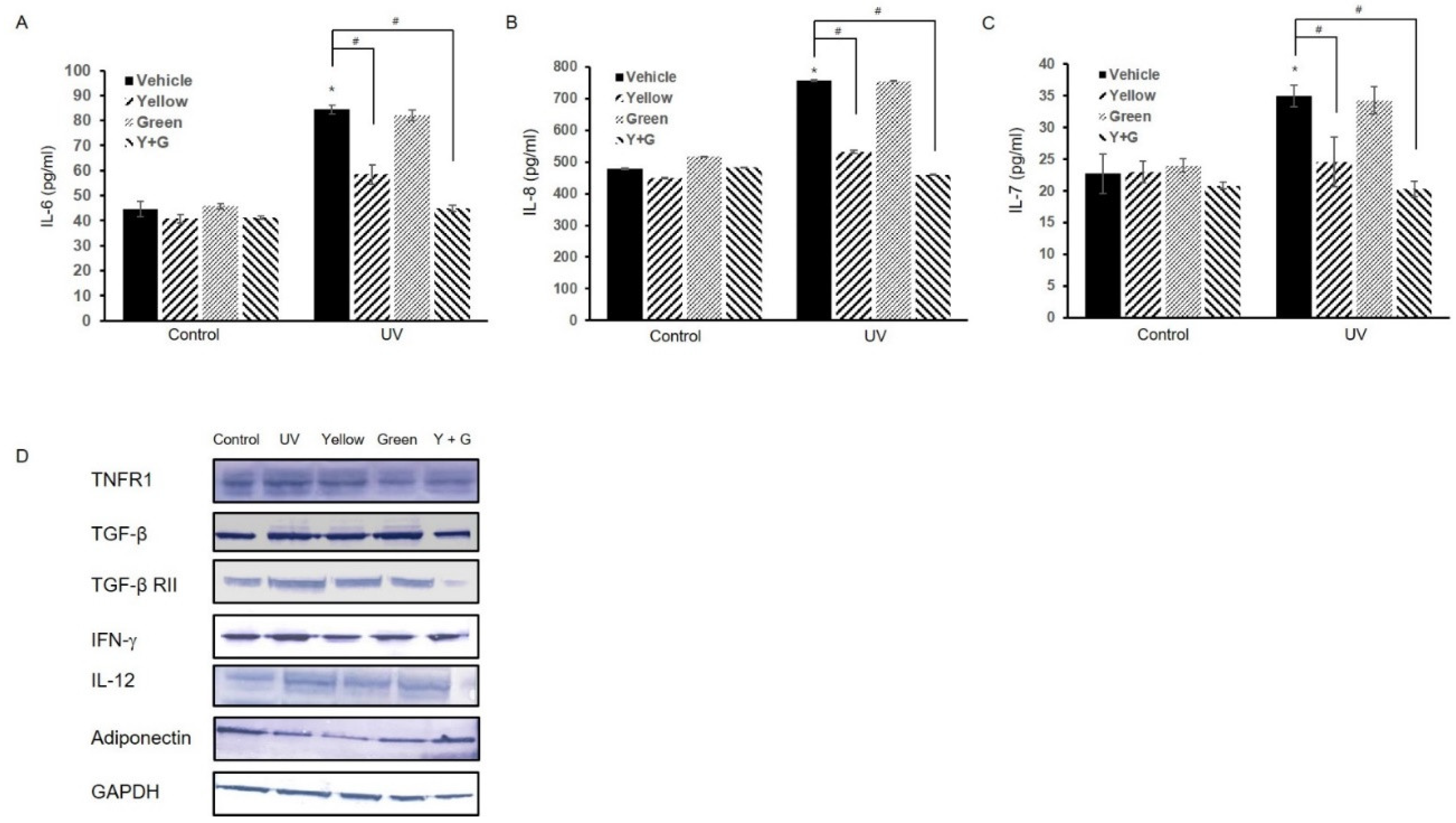
2.2. IPL Treatment Attenuates the UV-Induced Hyperimmune Reaction
2.3. IPL Treatment Reduces UV-Induced Hyperpigmentation
2.4. IPL Treatment Attenuates ROS Accumulation and Preserves Cellular Antioxidative Enzyme Capacity after UVB Irradiation
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Growth Conditions
4.3. UVB Light Apparatus
4.4. IPL Treatment
4.5. Cytokine Array and ELISA
4.6. Melanin Level Determination and Tyrosinase Enzymatic Activity Assay
4.7. RT-qPCR
4.8. Western Blot Analysis
4.9. Determination of Intracellular ROS Levels and Antioxidant Activity
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cadet, J.; Douki, T. Formation of UV-induced DNA damage contributing to skin cancer development. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2018, 17, 1816–1841. [Google Scholar] [CrossRef]
- Kimball, A.B.; Alora-Palli, M.B.; Tamura, M.; Mullins, L.A.; Soh, C.; Binder, R.L.; Houston, N.A.; Conley, E.D.; Tung, J.Y.; Annunziata, N.E.; et al. Age-induced and photoinduced changes in gene expression profiles in facial skin of Caucasian females across 6 decades of age. J. Am. Acad. Dermatol. 2018, 78, 29–39.e27. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Swalwell, H.; Latimer, J.; Haywood, R.M.; Birch-Machin, M.A. Investigating the role of melanin in UVA/UVB- and hydrogen peroxide-induced cellular and mitochondrial ROS production and mitochondrial DNA damage in human melanoma cells. Free Radic. Biol. Med. 2012, 52, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Penders, T.M.; Stanciu, C.N.; Schoemann, A.M.; Ninan, P.T.; Bloch, R.; Saeed, S.A. Bright Light Therapy as Augmentation of Pharmacotherapy for Treatment of Depression: A Systematic Review and Meta-Analysis. Prim. Care Companion CNS Disord. 2016, 18. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, M.X. A clinical review of phototherapy for psoriasis. Lasers Med. Sci. 2018, 33, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, N.S.; Lee, S.U.; Rethinasabapathy, M.; Lee, E.Z.; Cho, H.J.; Oh, S.Y.; Choe, S.R.; Kim, Y.; Hong, W.G.; Krishnan, G.; et al. Visible-light-driven dynamic cancer therapy and imaging using graphitic carbon nitride nanoparticles. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 90, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Kemény, L.; Varga, E.; Novak, Z. Advances in phototherapy for psoriasis and atopic dermatitis. Expert Rev. Clin. Immunol. 2019, 15, 1205–1214. [Google Scholar] [CrossRef]
- Mahmoud, B.H.; Hexsel, C.L.; Hamzavi, I.H.; Lim, H.W. Effects of visible light on the skin. Photochem. Photobiol. 2008, 84, 450–462. [Google Scholar] [CrossRef]
- Tsibadze, A.; Chikvaidze, E.; Katsitadze, A.; Kvachadze, I.; Tskhvediani, N.; Chikviladze, A. Visible light and human skin (review). Georgian Med. News 2015, 246, 46–53. [Google Scholar]
- Babilas, P.; Schreml, S.; Szeimies, R.M.; Landthaler, M. Intense pulsed light (IPL): A review. Lasers Surg. Med. 2010, 42, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, Y.; Wen, H.; Sun, X.; Wang, Y. Hydroxysafflor Yellow A Inhibits TNF-α-Induced Inflammation of Human Fetal Lung Fibroblasts via NF-κB Signaling Pathway. Evid. Based Complementary Altern. Med. ECAM 2019, 2019, 4050327. [Google Scholar] [CrossRef] [Green Version]
- On-Ong-Arj, P.; Wattanathorn, J.; Muchimapura, S.; Thukham-Mee, W. Yellow Laser Stimulation at GV2 Acupoint Mitigates Apoptosis, Oxidative Stress, Inflammation, and Motor Deficit in Spinal Cord Injury Rats. Evid. Based Complementary Altern. Med. ECAM 2018, 2018, 5407052. [Google Scholar] [CrossRef]
- Taylor, M.; Porter, R.; Gonzalez, M. Intense pulsed light may improve inflammatory acne through TNF-α down-regulation. J. Cosmet. Laser Ther. Off. Publ. Eur. Soc. Laser Dermatol. 2014, 16, 96–103. [Google Scholar] [CrossRef]
- Lee, M.W. Combination 532-nm and 1064-nm lasers for noninvasive skin rejuvenation and toning. Arch. Dermatol. 2003, 139, 1265–1276. [Google Scholar] [CrossRef] [Green Version]
- Vachiramon, V.; Iamsumang, W.; Triyangkulsri, K. Q-switched double frequency Nd:YAG 532-nm nanosecond laser vs. double frequency Nd:YAG 532-nm picosecond laser for the treatment of solar lentigines in Asians. Lasers Med Sci. 2018, 33, 1941–1947. [Google Scholar] [CrossRef] [PubMed]
- Park, K.Y.; Choi, S.Y.; Mun, S.K.; Kim, B.J.; Kim, M.N. Combined treatment with 578-/511-nm copper bromide laser and light-emitting diodes for post-laser pigmentation: A report of two cases. Dermatol. Ther. 2014, 27, 121–125. [Google Scholar] [CrossRef]
- Kennedy Crispin, M.; Fuentes-Duculan, J.; Gulati, N.; Johnson-Huang, L.M.; Lentini, T.; Sullivan-Whalen, M.; Gilleaudeau, P.; Cueto, I.; Suárez-Fariñas, M.; Lowes, M.A.; et al. Gene profiling of narrowband UVB-induced skin injury defines cellular and molecular innate immune responses. J. Investig. Dermatol. 2013, 133, 692–701. [Google Scholar] [CrossRef] [Green Version]
- Son, M.; Park, J.; Oh, S.; Choi, J.; Shim, M.; Kang, D.; Byun, K. Radiofrequency irradiation attenuates angiogenesis and inflammation in UVB-induced rosacea in mouse skin. Exp. Dermatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.C.; Liao, P.Y.; Hung, S.J.; Ge, J.S.; Chen, S.M.; Lai, J.C.; Hsiao, Y.P.; Yang, J.H. Topical application of glycolic acid suppresses the UVB induced IL-6, IL-8, MCP-1 and COX-2 inflammation by modulating NF-κB signaling pathway in keratinocytes and mice skin. J. Dermatol. Sci. 2017, 86, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.W.; Hung, Y.C.; Lin, T.Y.; Fang, J.Y.; Yang, P.M.; Chen, M.H.; Pan, T.L. Comparison of the Biological Impact of UVA and UVB upon the Skin with Functional Proteomics and Immunohistochemistry. Antioxidants 2019, 8, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröne, A. Keratinocytes and cytokines. Vet. Immunol. Immunopathol. 2002, 88, 1–12. [Google Scholar] [CrossRef]
- Lee, J.W.; Ryu, H.C.; Ng, Y.C.; Kim, C.; Wei, J.D.; Sabaratnam, V.; Kim, J.H. 12(S)-Hydroxyheptadeca-5Z,8E,10E-trienoic acid suppresses UV-induced IL-6 synthesis in keratinocytes, exerting an anti-inflammatory activity. Exp. Mol. Med. 2012, 44, 378–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.P.; Seo, H.S.; Shin, K.O.; Park, K.; Park, B.C.; Kim, M.H.; Park, M.; Kim, C.D.; Seo, S.J. Adiponectin Enhances Human Keratinocyte Lipid Synthesis via SIRT1 and Nuclear Hormone Receptor Signaling. J. Investig. Dermatol. 2019, 139, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Shibata, S.; Tada, Y.; Hau, C.S.; Mitsui, A.; Kamata, M.; Asano, Y.; Sugaya, M.; Kadono, T.; Masamoto, Y.; Kurokawa, M.; et al. Adiponectin regulates psoriasiform skin inflammation by suppressing IL-17 production from γδ-T cells. Nat. Commun. 2015, 6, 7687. [Google Scholar] [CrossRef] [Green Version]
- Alzyoud, J.A.M.; Khan, I.M.; Rees, S.G. In vitro studies to evaluate the effect of varying culture conditions and IPL fluencies on tenocyte activities. Lasers Med. Sci. 2017, 32, 1561–1570. [Google Scholar] [CrossRef]
- Chabert, R.; Fouque, L.; Pinacolo, S.; Garcia-Gimenez, N.; Bonnans, M.; Cucumel, K.; Domloge, N. Evaluation of light-emitting diodes (LED) effect on skin biology (in vitro study). Ski. Res. Technol. 2015, 21, 426–436. [Google Scholar] [CrossRef]
- Kobylinski, S.; Reble, C.; Schanzer, S.; Gersonde, I.; Wiora, G.; Lobo-Ploch, N.; Karrer, H.; Kolbe, L.; Khazaka, G.; Lademann, J.; et al. Non-invasive measurement of the 308 nm-LED based UVB protection factor of sunscreens. J. Biophotonics 2021. [Google Scholar] [CrossRef] [PubMed]
- Sayama, K.; Yuki, K.; Sugata, K.; Fukagawa, S.; Yamamoto, T.; Ikeda, S.; Murase, T. Carbon dioxide inhibits UVB-induced inflammatory response by activating the proton-sensing receptor, GPR65, in human keratinocytes. Sci. Rep. 2021, 11, 379. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmidt-Habel, A.; Löhberg, L.; Zajitschek, J.; Englbrecht, M.; Eichner, S.; Schuler, G. Treatment of vitiligo using targeted UVB-Intense Pulsed Light (IPL)- phototherapy: Evaluation of variables affecting therapeutic success. Eur. J. Dermatol. EJD 2014, 24, 551–559. [Google Scholar] [CrossRef]
- Horne, J.A.; Donlon, J.; Arendt, J. Green light attenuates melatonin output and sleepiness during sleep deprivation. Sleep 1991, 14, 233–240. [Google Scholar]
- Klein, R.M. Effects of green light on biological systems. Biol. Rev. Camb. Philos. Soc. 1992, 67, 199–284. [Google Scholar] [CrossRef]
- Tirico, M.; Jensen, D.; Green, C.; Ross, E.V. Short pulse intense pulsed light versus pulsed dye laser for the treatment of facial redness. J. Cosmet. Laser Ther. Off. Publ. Eur. Soc. Laser Dermatol. 2020, 22, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Wicks, N.L.; Chan, J.W.; Najera, J.A.; Ciriello, J.M.; Oancea, E. UVA phototransduction drives early melanin synthesis in human melanocytes. Curr. Biol. CB 2011, 21, 1906–1911. [Google Scholar] [CrossRef] [Green Version]
- Haltaufderhyde, K.; Ozdeslik, R.N.; Wicks, N.L.; Najera, J.A.; Oancea, E. Opsin expression in human epidermal skin. Photochem. Photobiol. 2015, 91, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofen, B.; Prado, G.; Emer, J. Melasma and Post Inflammatory Hyperpigmentation: Management Update and Expert Opinion. Ski. Ther. Lett. 2016, 21, 1–7. [Google Scholar]
- Thaysen-Petersen, D.; Lin, J.Y.; Nash, J.; Beerwerth, F.; Wulf, H.C.; Philipsen, P.A.; Haedersdal, M. The role of natural and UV-induced skin pigmentation on low-fluence IPL-induced side effects: A randomized controlled trial. Lasers Surg. Med. 2014, 46, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.R.; Vallejo, B.; Ruzgas, T.; Björklund, S. The Effect of UVB Irradiation and Oxidative Stress on the Skin Barrier-A New Method to Evaluate Sun Protection Factor Based on Electrical Impedance Spectroscopy. Sensors 2019, 19, 376. [Google Scholar] [CrossRef] [Green Version]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative stress in aging human skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [Green Version]
- Requena, M.B.; Vollet Filho, J.D.; Barboza de Nardi, A.; Escobar, A.; da Rocha Adams, R.; Bagnato, V.S.; de Menezes, P.F.C. Topical and intradermal delivery of PpIX precursors for photodynamic therapy with intense pulsed light on porcine skin model. Lasers Med. Sci. 2019, 34, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Alzyoud, J.A.M.; Joyce, N.S.; Woodward, R.D.; Khan, I.M.; Rees, S.G. In vitro tissue culture model validation-the influence of tissue culture components on IPL energy output. Lasers Med. Sci. 2019, 34, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Fishman, H.A.; Periman, L.M.; Shah, A.A. Real-Time Video Microscopy of In Vitro Demodex Death by Intense Pulsed Light. Photobiomodulation Photomed. Laser Surg. 2020, 38, 472–476. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Lee, J.; Choi, H. Intense Pulsed Light Attenuates UV-Induced Hyperimmune Response and Pigmentation in Human Skin Cells. Int. J. Mol. Sci. 2021, 22, 3173. https://doi.org/10.3390/ijms22063173
Kim J, Lee J, Choi H. Intense Pulsed Light Attenuates UV-Induced Hyperimmune Response and Pigmentation in Human Skin Cells. International Journal of Molecular Sciences. 2021; 22(6):3173. https://doi.org/10.3390/ijms22063173
Chicago/Turabian StyleKim, Juewon, Jeongin Lee, and Hyunjung Choi. 2021. "Intense Pulsed Light Attenuates UV-Induced Hyperimmune Response and Pigmentation in Human Skin Cells" International Journal of Molecular Sciences 22, no. 6: 3173. https://doi.org/10.3390/ijms22063173
APA StyleKim, J., Lee, J., & Choi, H. (2021). Intense Pulsed Light Attenuates UV-Induced Hyperimmune Response and Pigmentation in Human Skin Cells. International Journal of Molecular Sciences, 22(6), 3173. https://doi.org/10.3390/ijms22063173