
Adenosine A2A Receptors Are Upregulated in Peripheral Blood Mononuclear Cells from Atrial Fibrillation Patients

 ,
,  , ,
, , 
Abstract
:1. Introduction
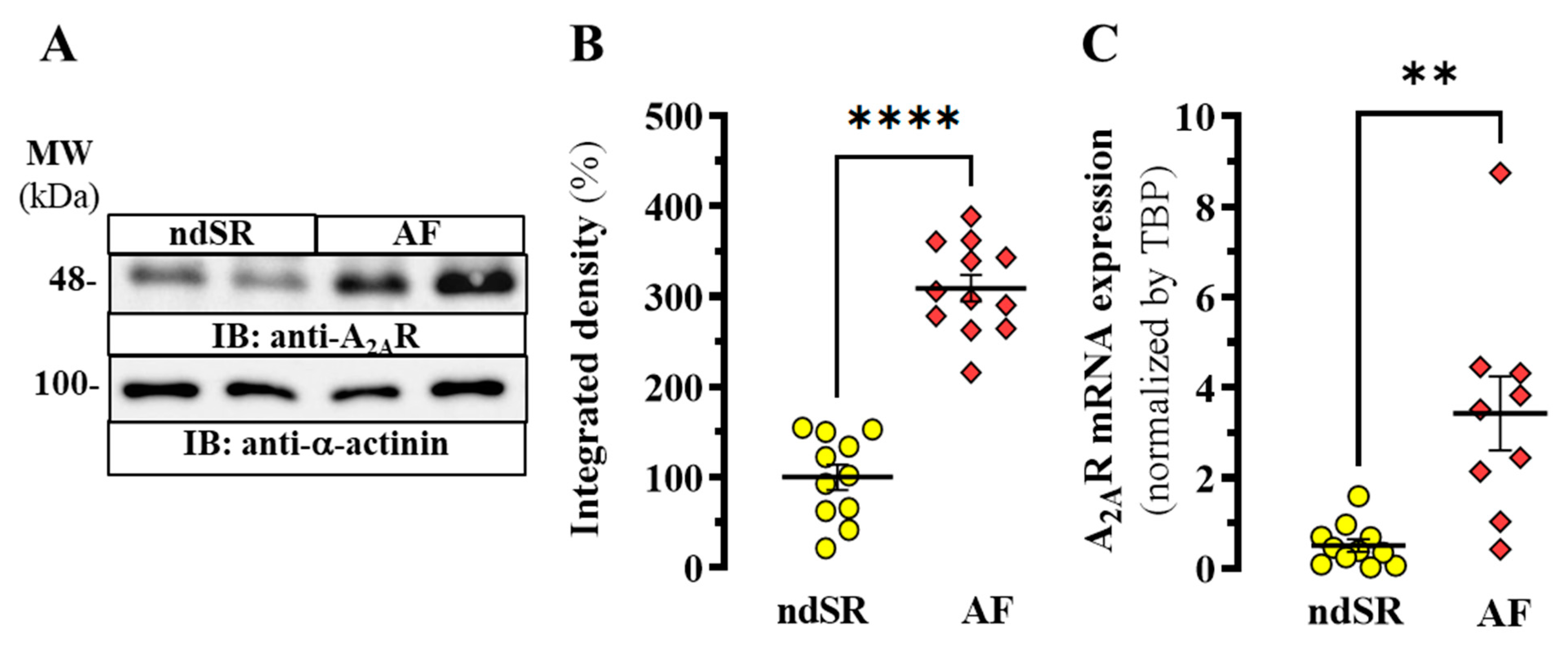
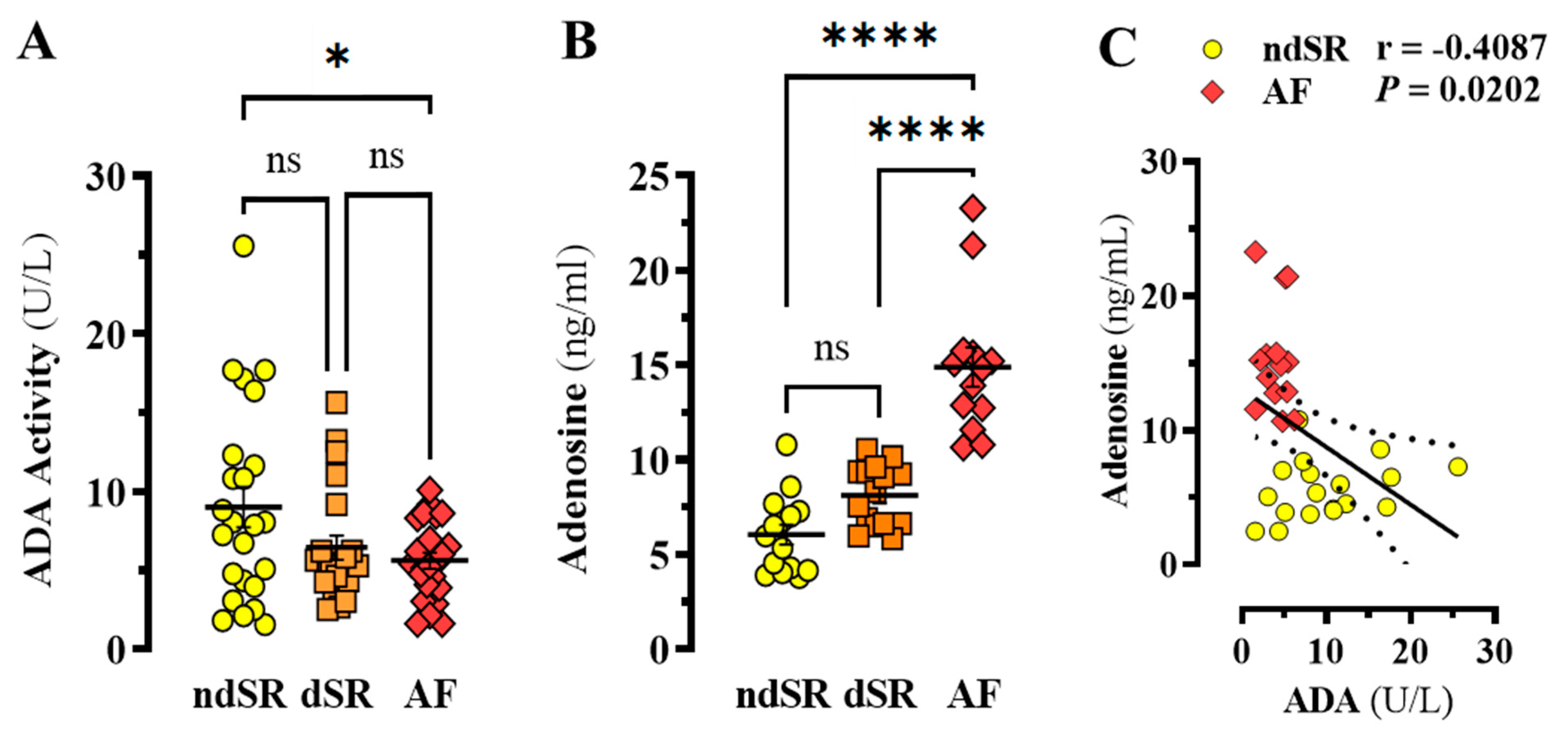
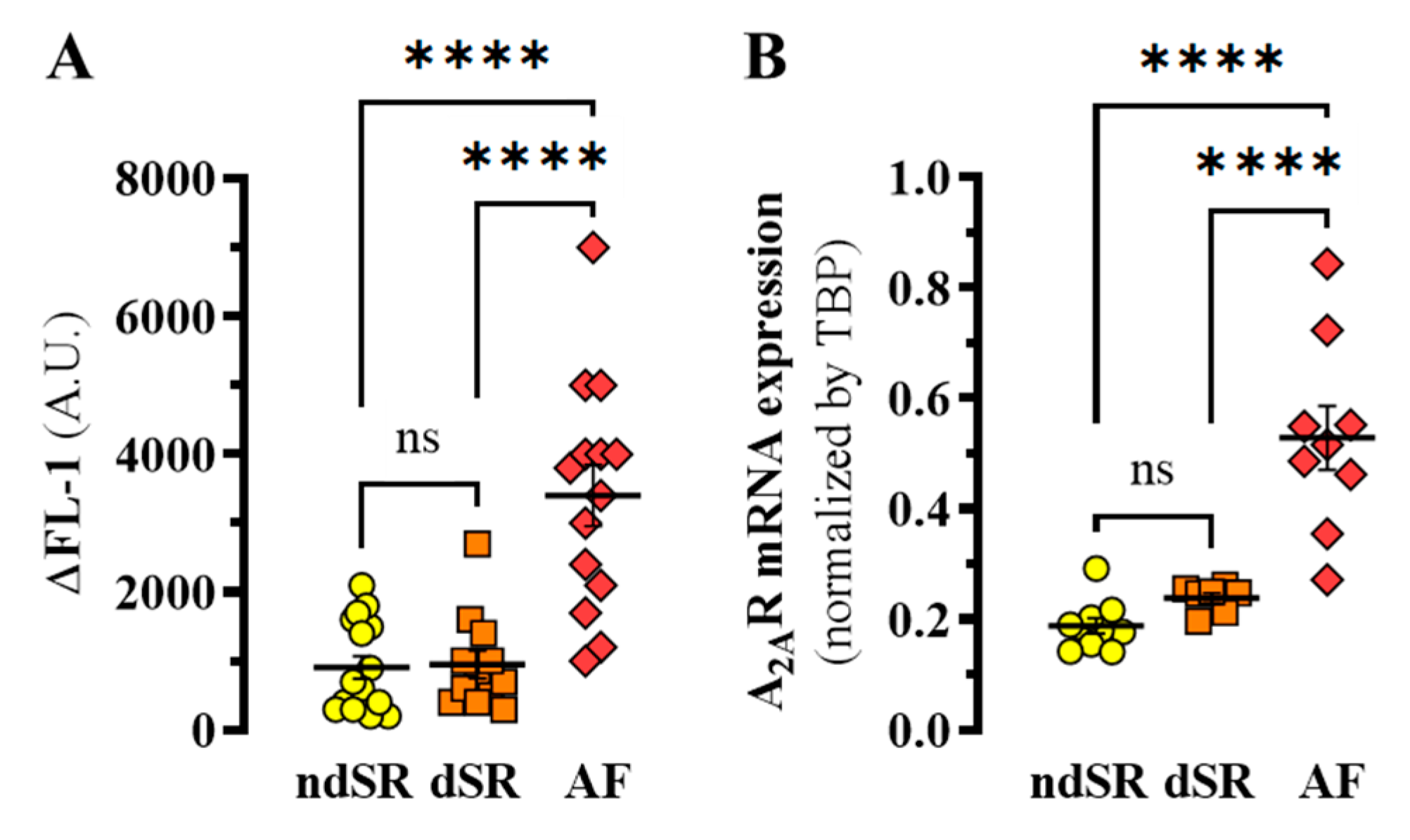
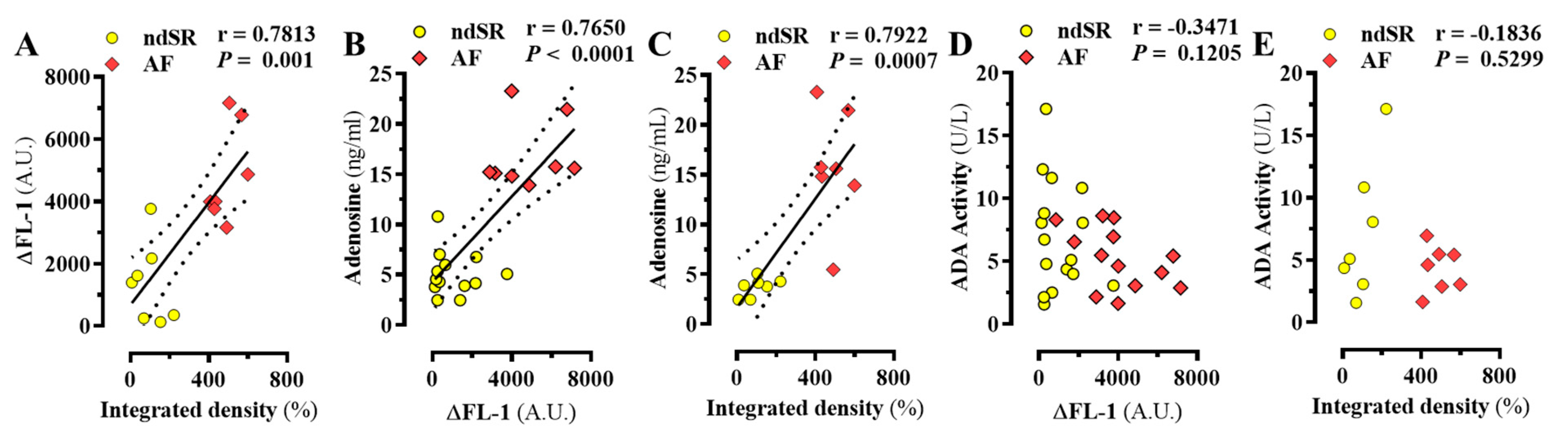
2. Results
Adenosinergic System in AF
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Human Samples (Subject Demographics)
4.3. Membrane Preparation
4.4. Gel Electrophoresis and Immunoblotting
4.5. RT-qPCR
4.6. Sample Preparation and HPLC-MS/MS Determination of Ribonucleosides
4.7. Adenosine Deaminase Activity Determination
4.8. Cell Culture
4.9. Flow Cytometry and Confocal Imaging
4.10. Data and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoni-Berisso, M.; Lercari, F.; Carazza, T.; Domenicucci, S. Epidemiology of atrial fbrillation: European perspective. Clin. Epidemiol. 2014, 6, 213–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravens, U. Antiarrhythmic therapy in atrial fibrillation. Pharmacol. Ther. 2010, 128, 129–145. [Google Scholar] [CrossRef]
- Staerk, L.; Sherer, J.A.; Ko, D.; Benjamin, E.J.; Helm, R.H. Atrial Fibrillation: Epidemiology, Pathophysiology, and Clinical Outcomes. Circ. Res. 2017, 120, 1501–1517. [Google Scholar] [CrossRef] [Green Version]
- Harada, M.; Van Wagoner, D.R.; Nattel, S. Role of inflammation in Atrial fibrillation pathophysiology and management. Circ. J. 2015, 79, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korantzopoulos, P.; Letsas, K.P.; Tse, G.; Fragakis, N.; Goudis, C.A.; Liu, T. Inflammation and atrial fibrillation: A comprehensive review. J. Arrhythmia 2018, 34, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Berkovitch, A.; Younis, A.; Grossman, Y.; Segev, S.; Kivity, S.; Sidi, Y.; Beinart, R.; Goldenberg, I.; Maor, E. Relation of Neutrophil to Lymphocyte Ratio to Risk of Incident Atrial Fibrillation. Am. J. Cardiol. 2019, 123, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Gessi, S.; Varani, K.; Merighi, S.; Fogli, E.; Sacchetto, V.; Benini, A.; Leung, E.; Mac-Lennan, S.; Borea, P.A. Adenosine and lymphocyte regulation. Purinergic Signal. 2007, 3, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Newby, A.C.; Worku, Y.; Holmquist, C.A. Adenosine formation. Evidence for a direct biochemical link with energy metabolism. Adv. Myocardiol. 1985, 6, 273–284. [Google Scholar]
- Haskó, G.; Linden, J.; Cronstein, B.; Pacher, P. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. Nature 2014, 509, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltzschig, H.K.; Faigle, M.; Knapp, S.; Karhausen, J.; Ibla, J.; Rosenberger, P.; Odegard, K.C.; Laussen, P.C.; Thompson, L.F.; Colgan, S.P. Endothelial catabolism of extracellular adenosine during hypoxia: The role of surface adenosine deaminase and CD26. Blood 2006, 108, 1602–1610. [Google Scholar] [CrossRef]
- Grenz, A.; Homann, D.; Eltzschig, H.K. Extracellular adenosine: A safety signal that dampens hypoxia-induced inflammation during ischemia. Antioxid. Redox Signal. 2011, 15, 2221–2234. [Google Scholar] [CrossRef] [Green Version]
- Sumi, Y.; Woehrle, T.; Chen, Y.; Yao, Y.; Li, A.; Junger, W.G. Adrenergic receptor activation involves ATP release and feedback through purinergic receptors. Am. J. Physiol. Cell Physiol. 2010, 299, C1118–C1126. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Csepe, T.A.; Hansen, B.J.; Sul, L.V.; Kalyanasundaram, A.; Zakharkin, S.O.; Zhao, J.; Guha, A.; Van Wagoner, D.R.; Kilic, A.; et al. Adenosine-induced atrial fibrillation. Circulation 2016, 134, 486–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, T.; Takasugi, N.; Naruse, G.; Okura, H. Adenosine-induced atrial fibrillation arising from arrhythmogenic right atrial appendage. J. Cardiovasc. Electrophysiol. 2020, 31, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Hove-Madsen, L.; Prat-Vidal, C.; Llach, A.; Ciruela, F.; Casadó, V.; Lluis, C.; Bayes-Genis, A.; Cinca, J.; Franco, R. Adenosine A2A receptors are expressed in human atrial myocytes and modulate spontaneous sarcoplasmic reticulum calcium release. Cardiovasc. Res. 2006, 72, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llach, A.; Molina, C.E.; Prat-Vidal, C.; Fernandes, J.; Casad, V.; Ciruela, F.; Llus, C.; Franco, R.; Cinca, J.; Hove-Madsen, L. Abnormal calcium handling in atrial fibrillation is linked to up-regulation of adenosine A2A receptors. Eur. Heart J. 2011, 32, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Boknik, P.; Drzewiecki, K.; Eskandar, J.; Gergs, U.; Hofmann, B.; Treede, H.; Grote-Wessels, S.; Fabritz, L.; Kirchhof, P.; Fortmüller, L.; et al. Evidence for arrhythmogenic effects of A2A-adenosine receptors. Front. Pharmacol. 2019, 10, 1051. [Google Scholar] [CrossRef]
- Soattin, L.; Lubberding, A.F.; Bentzen, B.H.; Christ, T.; Jespersen, T. Inhibition of Adenosine Pathway Alters Atrial Electrophysiology and Prevents Atrial Fibrillation. Front. Physiol. 2020, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- Guieu, R.; Deharo, J.-C.; Maille, B.; Crotti, L.; Torresani, E.; Brignole, M.; Parati, G. Adenosine and the Cardiovascular System: The Good and the Bad. J. Clin. Med. 2020, 9, 1366. [Google Scholar] [CrossRef]
- Duroux, R.; Ciancetta, A.; Mannes, P.; Yu, J.; Boyapati, S.; Gizewski, E.; Yous, S.; Ciruela, F.; Auchampach, J.A.; Gao, Z.-G.; et al. Bitopic fluorescent antagonists of the A2A adenosine receptor based on pyrazolo[4,3-: E] [1,2,4]triazolo[1,5- c] pyrimidin-5-amine functionalized congeners. Medchemcomm 2017, 8, 1659–1667. [Google Scholar] [CrossRef] [Green Version]
- Boknik, P.; Eskandar, J.; Hofmann, B.; Zimmermann, N.; Neumann, J.; Gergs, U. Role of cardiac A2A receptors under normal and pathophysiological conditions. Front. Pharmacol. 2021, 11, 627838. [Google Scholar] [CrossRef]
- Gessi, S.; Varani, K.; Merighi, S.; Ongini, E.; Borea, P.A. A(2A) adenosine receptors in human peripheral blood cells. Br. J. Pharmacol. 2000, 129, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Bortoluzzi, A.; Vincenzi, F.; Govoni, M.; Padovan, M.; Ravani, A.; Borea, P.A.; Varani, K. A2A adenosine receptor upregulation correlates with disease activity in patients with systemic lupus erythematosus. Arthritis Res. Ther. 2016, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vincenzi, F.; Corciulo, C.; Targa, M.; Casetta, I.; Gentile, M.; Granieri, E.; Borea, P.A.; Popoli, P.; Varani, K. A2A adenosine receptors are up-regulated in lymphocytes from amyotrophic lateral sclerosis patients. Amyotroph. Lateral Scler. Front. Degener. 2013, 14, 406–413. [Google Scholar] [CrossRef]
- Vincenzi, F.; Corciulo, C.; Targa, M.; Merighi, S.; Gessi, S.; Casetta, I.; Gentile, M.; Granieri, E.; Borea, P.A.; Varani, K. Multiple sclerosis lymphocytes upregulate A2A adenosine receptors that are antiinflammatory when stimulated. Eur. J. Immunol. 2013, 43, 2206–2216. [Google Scholar] [CrossRef]
- Vincenzi, F.; Padovan, M.; Targa, M.; Corciulo, C.; Giacuzzo, S.; Merighi, S.; Gessi, S.; Govoni, M.; Borea, P.A.; Varani, K. A2A Adenosine Receptors Are Differentially Modulated by Pharmacological Treatments in Rheumatoid Arthritis Patients and Their Stimulation Ameliorates Adjuvant-Induced Arthritis in Rats. PLoS ONE 2013, 8, e54195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gariboldi, V.; Vairo, D.; Guieu, R.; Marlingue, M.; Ravis, E.; Lagier, D.; Mari, A.; Thery, E.; Collart, F.; Gaudry, M.; et al. Expressions of adenosine A2A receptors in coronary arteries and peripheral blood mononuclear cells are correlated in coronary artery disease patients. Int. J. Cardiol. 2017, 230, 427–431. [Google Scholar] [CrossRef]
- Varani, K.; Laghi-Pasini, F.; Camurri, A.; Capecchi, P.L.; Maccherini, M.; Diciolla, F.; Ceccatelli, L.; Lazzerini, P.E.; Ulouglu, C.; Cattabeni, F.; et al. Changes of peripheral A2A adenosine receptors in chronic heart failure and cardiac transplantation. FASEB J. 2003, 17, 280–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodadadi, I.; Vahedi, M.S.; Abdi, M.; Daneshkhah, N.; Rahbari, R.; Menbari, S.; Ahmadi, D.; Ahmadi, A.; Lahoorpour, F.; Hakhamaneshi, M.S.; et al. Evaluation of adenosine deaminase (ADA) isoenzymes activity and tumor necrosis factor-α (TNFα) concentration in chronic heart failure. EXCLI J. 2014, 13, 58–66. [Google Scholar] [PubMed]
- Molina, C.E.; Llach, A.; Herraiz-Martínez, A.; Tarifa, C.; Barriga, M.; Wiegerinck, R.F.; Fernandes, J.; Cabello, N.; Vallmitjana, A.; Benitéz, R.; et al. Prevention of adenosine A2A receptor activation diminishes beat-to-beat alternation in human atrial myocytes. Basic Res. Cardiol. 2016, 111, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biaggioni, I. Adenosine Receptors and Autonomic Regulation. In Primer on the Autonomic Nervous System; Academic Press: Cambridge, MA, USA, 2012; pp. 95–97. [Google Scholar]
- Lanznaster, D.; Massari, C.M.; Marková, V.; Šimková, T.; Duroux, R.; Jacobson, K.A.; Fernández-Dueñas, V.; Tasca, C.I.; Ciruela, F. Adenosine A1-A2A Receptor-Receptor Interaction: Contribution to Guanosine-Mediated Effects. Cells 2019, 8, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Dueñas, V.; Gómez-Soler, M.; Valle-León, M.; Watanabe, M.; Ferrer, I.; Ciruela, F. Revealing Adenosine A2A-Dopamine D2 Receptor Heteromers in Parkinson’s Disease Post-Mortem Brain through a New AlphaScreen-Based Assay. Int. J. Mol. Sci. 2019, 20, 3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morató, X.; Gonçalves, F.Q.; Lopes, J.P.; Jauregui, O.; Soler, C.; Fernández-Dueñas, V.; Cunha, R.A.; Ciruela, F. Chronic adenosine A 2A receptor blockade induces locomotor sensitization and potentiates striatal LTD IN GPR37-deficient mice. J. Neurochem. 2019, 148, 796–809. [Google Scholar] [CrossRef]
- Maille, B.; Marlinge, M.; Vairo, D.; Mottola, G.; Koutbi, L.; Deharo, P.; Gastaldi, M.; Gaudry, M.; Guiol, C.; Bottone, S.; et al. Adenosine plasma level in patients with paroxysmal or persistent atrial fibrillation and normal heart during ablation procedure and/or cardioversion. Purinergic Signal. 2019, 15, 45–52. [Google Scholar] [CrossRef]
- Fernández-Dueñas, V.; Taura, J.J.; Cottet, M.; Gómez-Soler, M.; López-Cano, M.; Ledent, C.; Watanabe, M.; Trinquet, E.; Pin, J.-P.; Luján, R.; et al. Untangling dopamine-adenosine receptor-receptor assembly in experimental parkinsonism in rats. Dis. Model. Mech. 2015, 8, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Dueñas, V.; Gomez-Soler, M.; Morato, X.; Nunez, F.; Das, A.; Kumar, T.S.; Jauma, S.; Jacobson, K.A.; Ciruela, F. Dopamine D2 receptor-mediated modulation of adenosine A2A receptor agonist binding within the A2AR/D2R oligomer framework. Neurochem. Int. 2013, 63, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Motulsky, H.J.; Brown, R.E. Detecting outliers when fitting data with nonlinear regression—A new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ndSR | dSR | AF | |
|---|---|---|---|
| Number of patients | 28 | 24 | 27 |
| Weight (mean±SD) (Kg) | 83.18 ± 9.86 | 71.5 ± 10.5 | 75.5 ± 11.386 |
| Height (mean±SD) (cm) | 169.4 ± 9.8 | 160.8 ± 8.7 | 163.8 ± 8.9 |
| Age (mean±SD) | 63.1 ± 9.7 | 73.5 ± 8.4 | 72.9 ± 9.4 |
| Sex (male/female) | 23/5 | 17/7 | 14/13 |
| Body Surface (mean ± SD) (m2) | 1.93 ± 0.14 | 1.75 ± 0.15 | 1.81 ± 0.15 |
| LA diameter index (mean ± SD) | 1.99 ± 0.17 | 2.6 ± 0.2 | 2.65 ± 0.55 |
| LV diameter index (mean ± SD) | 2.7 ± 0.5 | 2.83 ± 0.54 | 2.82 ± 0.42 |
| Tabaquism (y/n/ex) | 9/12/7 | 3/18/3 | 6/18/3 |
| Enolism (y/n/ex) | 0/28/0 | 1/23/0 | 1/26/0 |
| Hipertension | 25 | 22 | 20 |
| Diabetes | 11 | 12 | 9 |
| Dislipemia | 15 | 18 | 15 |
| Drugs | |||
| IECAs | 16 | 12 | 12 |
| Beta-bloquers | 12 | 9 | 16 |
| β1 | 3 | 2 | 8 |
| β2 | 0 | 0 | 1 |
| α | 0 | 0 | 0 |
| RyR-inhibitors | 0 | 0 | 0 |
| ARA-II | 5 | 4 | 2 |
| Ca2+ antagonist | 6 | 3 | 4 |
| Sintrom | 3 | 1 | 16 |
| AAS | 17 | 15 | 6 |
| Statins | 17 | 18 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godoy-Marín, H.; Duroux, R.; Jacobson, K.A.; Soler, C.; Colino-Lage, H.; Jiménez-Sábado, V.; Montiel, J.; Hove-Madsen, L.; Ciruela, F. Adenosine A2A Receptors Are Upregulated in Peripheral Blood Mononuclear Cells from Atrial Fibrillation Patients. Int. J. Mol. Sci. 2021, 22, 3467. https://doi.org/10.3390/ijms22073467
Godoy-Marín H, Duroux R, Jacobson KA, Soler C, Colino-Lage H, Jiménez-Sábado V, Montiel J, Hove-Madsen L, Ciruela F. Adenosine A2A Receptors Are Upregulated in Peripheral Blood Mononuclear Cells from Atrial Fibrillation Patients. International Journal of Molecular Sciences. 2021; 22(7):3467. https://doi.org/10.3390/ijms22073467
Chicago/Turabian StyleGodoy-Marín, Héctor, Romain Duroux, Kenneth A. Jacobson, Concepció Soler, Hildegard Colino-Lage, Veronica Jiménez-Sábado, José Montiel, Leif Hove-Madsen, and Francisco Ciruela. 2021. "Adenosine A2A Receptors Are Upregulated in Peripheral Blood Mononuclear Cells from Atrial Fibrillation Patients" International Journal of Molecular Sciences 22, no. 7: 3467. https://doi.org/10.3390/ijms22073467
APA StyleGodoy-Marín, H., Duroux, R., Jacobson, K. A., Soler, C., Colino-Lage, H., Jiménez-Sábado, V., Montiel, J., Hove-Madsen, L., & Ciruela, F. (2021). Adenosine A2A Receptors Are Upregulated in Peripheral Blood Mononuclear Cells from Atrial Fibrillation Patients. International Journal of Molecular Sciences, 22(7), 3467. https://doi.org/10.3390/ijms22073467