
Differential Response of Grapevine to Infection with ‘Candidatus Phytoplasma solani’ in Early and Late Growing Season through Complex Regulation of mRNA and Small RNA Transcriptomes

 , , , ,
, , , ,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. mRNAs and sRNAs Show Differential Temporal Involvement in Grapevine Infected with ‘Ca. P. solani’
2.2. Gene Set Enrichment Analysis Confirms High Transcriptional Activity in the Early Growing Season
2.3. Genes and miRNAs Not Associated with Phytoplasma Diseases Contribute to Resolving the Sanitary Status of Grapevines
2.4. The Most Differentially Expressed Genes and sRNAs Are Associated with Different Aspects of Biotic Stress Signaling
2.4.1. Important Involvement of Genes Associated with the Cell Wall in Bois Noir Pathogenesis
2.4.2. Hormonal Balance Is Disturbed Already in Pre-Symptomatic Phase of Phytoplasma Infection
2.5. Known Pathways with the Involvement of Novel Genes
2.6. Temporal Network Modeling Reveals New Cross-Talk between Pathways Involved in Grapevines Infected with ‘Ca. P. solani’
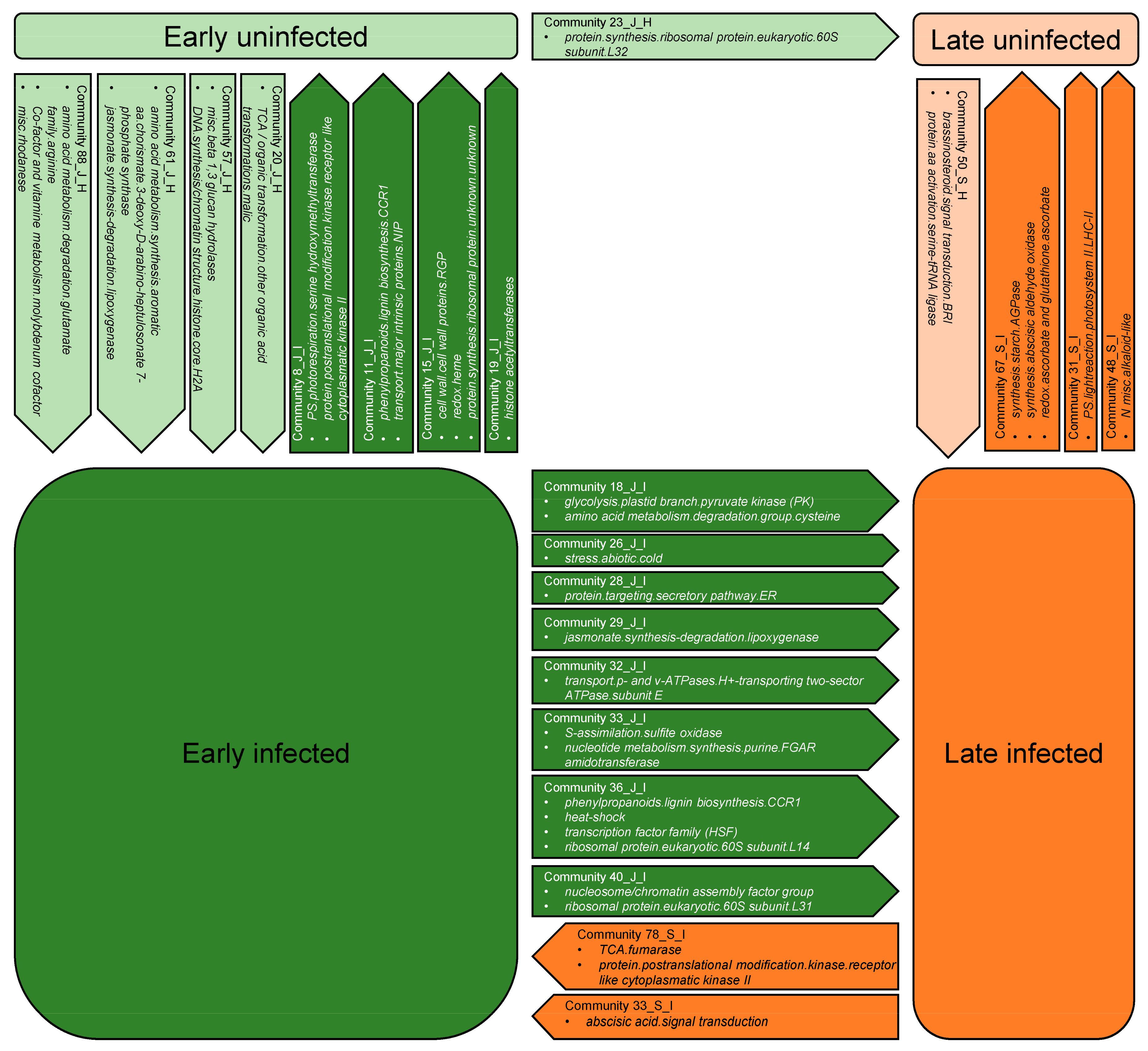
2.6.1. Disintegrated Communities in Infected Grapevines
2.6.2. Early Growing Season Communities with a High Dissipation Index in Uninfected Grapevines
2.6.3. Early versus Late Growing Season Communities in Infected Grapevines
2.6.4. Exploring mRNA-mRNA-miRNA Interaction Networks
3. Conclusions
4. Materials and Methods
4.1. Plant Material
4.2. Detection of Phytoplasma
4.3. RNA Extraction and Sequencing
4.4. mRNA Data Analysis
4.5. sRNA Data Analysis
4.6. sRNA Target Prediction
4.7. Differential Expression Analysis and Visualization of mRNA and sRNA Expression
4.8. Network Community Analyses
4.9. Targeted Grapevine Gene Expression Analysis by qPCR
4.10. Enzymatic Activities
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Starý, M.; Válová, P.; Šafářová, D.; Lauterer, P.; Ackermann, P.; Navrátil, M. Survey and molecular detection of Bois noir in vineyards of the Czech Republic. Hortic. Sci. 2013, 40, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Quaglino, F.; Zhao, Y.; Casati, P.; Bulgari, D.; Bianco, P.A.; Wei, W.; Davis, R.E. ‘Candidatus Phytoplasma solani’, a novel taxon associated with stolbur- and bois noir-related diseases of plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2879–2894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermastia, M. Interactions between grapevines and grapevine yellows phytoplasmas BN and FD. In Grapevine Yellows Diseases and Their Phytoplasma Agents; Dermastia, M., Bertaccini, A., Constable, F., Mehle, N., Eds.; Springer: Cham, Switzerland, 2017; pp. 47–67. [Google Scholar]
- Albertazzi, G.; Milc, J.; Caffagni, A.; Francia, E.; Roncaglia, E.; Ferrari, F.; Tagliafico, E.; Stefani, E.; Pecchioni, N. Gene expression in grapevine cultivars in response to Bois Noir phytoplasma infection. Plant Sci. 2009, 176, 792–804. [Google Scholar] [CrossRef]
- Bertamini, M.; Nedunchezhian, N. Decline of photosynthetic pigments, ribulose-1,5-bisphosphate carboxylase and soluble protein contents, nitrate reductase and photosynthetic activities, and changes in tylakoid membrane protein pattern in canopy shade grapevine (Vitis vinifera L.). Photosynthetica 2001, 39, 529–537. [Google Scholar] [CrossRef]
- Bertamini, M.; Nedunchezhian, N. Effects of phytoplasma [stolbur-subgroup (Bois noir-BN)] on photosynthetic pigments, saccharides, ribulose 1,5-bisphosphate carboxylase, nitrate and nitrite reductases, and photosynthetic activities in field-grown grapevine (Vitis vinifera L.) cv. Photosynthetica 2001, 39, 119–122. [Google Scholar] [CrossRef]
- Bertamini, M.; Nedunchezhian, N.; Tomasi, F.; Grando, M. Phytoplasma [Stolbur-subgroup (Bois Noir-BN)] infection inhibits photosynthetic pigments, ribulose-1,5-bisphosphate carboxylase and photosynthetic activities in field grown grapevine (Vitis vinifera L. cv. Chardonnay) leaves. Physiol. Mol. Plant Pathol. 2002, 61, 357–366. [Google Scholar] [CrossRef]
- Hren, M.; Nikolić, P.; Rotter, A.; Blejec, A.; Terrier, N.; Ravnikar, M.; Dermastia, M.; Gruden, K. “Bois noir” phytoplasma induces significant reprogramming of the leaf transcriptome in the field grown grapevine. BMC Genom. 2009, 10, 460. [Google Scholar] [CrossRef] [Green Version]
- Lepka, P.; Stitt, M.; Moll, E.; Seemmüller, E. Effect of phytoplasmal infection on concentration and translocation of carbohydrates and amino acids in periwinkle and tobacco. Physiol. Mol. Plant Pathol. 1999, 55, 59–68. [Google Scholar] [CrossRef]
- Guthrie, J.N.; Walsh, K.B.; Scott, P.T.; Rasmussen, T.S.; Group, P.S.; Industries, P.; Centre, R.S.; Road, M.; Guthrie, J.N.; Walsh, K.B.; et al. The phytopathology of Australian papaya dieback: A proposed role for the phytoplasma. Physiol. Mol. Plant Pathol. 2001, 58, 23–30. [Google Scholar] [CrossRef]
- Maust, B.E.; Espadas, F.; Talavera, C.; Aguilar, M.; Santamaría, J.M.; Oropeza, C. Changes in carbohydrate metabolism in coconut palms infected with the lethal yellowing phytoplasma. Phytopathology 2003, 93, 976–981. [Google Scholar] [CrossRef]
- Gai, Y.P.; Han, X.J.; Li, Y.Q.; Yuan, C.Z.; Mo, Y.Y.; Guo, F.Y.; Liu, Q.X.; Ji, X.L. Metabolomic analysis reveals the potential metabolites and pathogenesis involved in mulberry yellow dwarf disease. Plant Cell Environ. 2014, 37, 1474–1490. [Google Scholar] [CrossRef] [PubMed]
- Prezelj, N. Molecular Interactions between Phytoplasmal Causal Agents of Grapevine Yellows Disease and Grapevine (Vitis vinifera L.); University of Ljubljana: Ljubljana, Slovenia, 2014. [Google Scholar]
- Prezelj, N.; Covington, E.; Roitsch, T.; Gruden, K.; Fragner, L.; Weckwerth, W.; Chersicola, M.; Vodopivec, M.; Dermastia, M. Metabolic consequences of infection of grapevine (Vitis vinifera L.) cv. “Modra frankinja” with flavescence dorée phytoplasma. Front. Plant Sci. 2016, 7, 711. [Google Scholar] [CrossRef] [PubMed]
- Luge, T.; Kube, M.; Freiwald, A.; Meierhofer, D.; Seemüller, E.; Sauer, S. Transcriptomics assisted proteomic analysis of Nicotiana occidentalis infected by Candidatus Phytoplasma mali strain AT. Proteomics 2014, 14, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Santi, S.; De Marco, F.; Polizzotto, R.; Grisan, S.; Musetti, R. Recovery from stolbur disease in grapevine involves changes in sugar transport and metabolism. Front. Plant Sci. 2013, 4, 171. [Google Scholar] [CrossRef] [Green Version]
- Prezelj, N.; Fragener, L.; Weckwerth, W.; Dermastia, M. Metabolome of grapevine leaf vein-enriched tissue infected with ‘Candidatus Phytoplasma solani’. Mitt. Klost. Rebe Wein Obs. Früchteverwertung 2016, 66, 74–78. [Google Scholar]
- Santi, S.; Grisan, S.; Pierasco, A.; De Marco, F.; Musetti, R. Laser microdissection of grapevine leaf phloem infected by stolbur reveals site-specific gene responses associated to sucrose transport and metabolism. Plant Cell Environ. 2013, 36, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Covington Dunn, E.; Roitsch, T.; Dermastia, M.; Covington, E.D.; Roitsch, T.; Dermastia, M. Determination of the activity signature of key carbohydrate metabolism enzymes in phenolic-rich grapevine tissues. Acta Chim. Slov. 2016, 63, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Rotter, A.; Nikolić, P.; Turnšek, N.; Kogovšek, P.; Blejec, A.; Gruden, K.; Dermastia, M. Statistical modeling of long-term grapevine response to ‘Candidatus Phytoplasma solani’ infection in the field. Eur. J. Plant Pathol. 2018, 150, 653–668. [Google Scholar] [CrossRef] [Green Version]
- Hren, M.; Ravnikar, M.; Brzin, J.; Ermacora, P.; Carraro, L.; Bianco, P.A.; Casati, P.; Borgo, M.; Angelini, E.; Rotter, A.; et al. Induced expression of sucrose synthase and alcohol dehydrogenase I genes in phytoplasma-infected grapevine plants grown in the field. Plant Pathol. 2009, 58, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Dermastia, M.; Nikolic, P.; Chersicola, M.; Gruden, K. Transcriptional profiling in infected and recovered grapevine plant responses to ’Candidatus Phytoplasma solani’. Phytopathogenic Mollicutes 2015, 5, S123. [Google Scholar] [CrossRef]
- Rusjan, D.; Mikulic-Petkovsek, M. Phenolic responses in 1-year-old canes of V itis vinifera cv. Chardonnay induced by grapevine yellows (Bois noir). Aust. J. Grape Wine Res. 2015, 21, 123–134. [Google Scholar] [CrossRef]
- Rusjan, D.; Halbwirth, H.; Stich, K.; Mikulič-Petkovšek, M.; Veberič, R. Biochemical response of grapevine variety “Chardonnay” (Vitis vinifera L.) to infection with grapevine yellows (Bois noir). Eur. J. Plant Pathol. 2012, 134, 231–237. [Google Scholar] [CrossRef]
- Rusjan, D.; Veberič, R.; Mikulič-Petkovšek, M. The response of phenolic compounds in grapes of the variety “Chardonnay” (Vitis vinifera L.) to the infection by phytoplasma Bois noir. Eur. J. Plant Pathol. 2012, 133, 965–974. [Google Scholar] [CrossRef]
- Landi, L.; Romanazzi, G. Seasonal variation of defense-related gene expression in leaves from bois noir affected and recovered grapevines. J. Agric. Food Chem. 2011, 59, 6628–6637. [Google Scholar] [CrossRef]
- Dermastia, M. Plant hormones in phytoplasma infected plants. Front. Plant Sci. 2019, 10, 477. [Google Scholar] [CrossRef] [Green Version]
- Cora, D.; Re, A.; Caselle, M.; Bussolino, F. MicroRNA-mediated regulatory circuits: Outlook and perspectives. Phys. Biol. 2017, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megraw, M.; Cumbie, J.S.; Ivanchenko, M.G.; Filichkin, S.A. Small genetic circuits and microRNAs: Big players in polymerase II transcriptional control in plants. Plant Cell 2015, 28, 286–303. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.L.V.; Chekanova, J.A. Small RNAs: Essential regulators of gene expression and defenses against environmental stresses in plants. Wiley Interdiscip. Rev. RNA 2016, 7, 356–381. [Google Scholar] [CrossRef]
- Belli Kullan, J.; Lopes Paim Pinto, D.; Bertolini, E.; Fasoli, M.; Zenoni, S.; Tornielli, G.B.; Pezzotti, M.; Meyers, B.C.; Farina, L.; Pè, M.E.; et al. miRVine: A microRNA expression atlas of grapevine based on small RNA sequencing. BMC Genom. 2015, 16, 393. [Google Scholar] [CrossRef] [Green Version]
- Ehya, F.; Monavarfeshani, A.; Mohseni Fard, E.; Karimi Farsad, L.; Khayam Nekouei, M.; Mardi, M.; Salekdeh, G.H. Phytoplasma-responsive microRNAs modulate hormonal, nutritional, and stress signalling pathways in Mexican lime trees. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Shao, F.; Zhang, Q.; Liu, H.; Lu, S.; Qiu, D. Genome-wide identification and analysis of microRNAs involved in witches’-broom phytoplasma response in Ziziphus jujuba. PLoS ONE 2016, 11, e0166099. [Google Scholar] [CrossRef]
- Gai, Y.-P.; Zhao, H.-N.; Zhao, Y.-N.; Zhu, B.-S.; Yuan, S.-S.; Li, S.; Guo, F.-Y.; Ji, X.-L. MiRNA-seq-based profiles of miRNAs in mulberry phloem sap provide insight into the pathogenic mechanisms of mulberry yellow dwarf disease. Sci. Rep. 2018, 8, 812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, G.; Niu, S.; Zhao, Z.; Deng, M.; Xu, E.; Wang, Y.; Yang, L. Identification of microRNAs and their targets in Paulownia fortunei plants free from phytoplasma pathogen after methyl methane sulfonate treatment. Biochimie 2016, 127, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Pagliarani, C.; Abbà, S.; Boccacci, P.; Birello, G.; Rossi, M.; Palmano, S.; Marzachì, C.; Perrone, I.; Gambino, G. miRVIT: A novel miRNA database and its application to uncover Vitis responses to favescence dorée infection. Front. Plant Sci. 2018, 9, 1034. [Google Scholar] [CrossRef] [PubMed]
- Snyman, M.C.; Solofoharivelo, M.-C.; Souza-Richards, R.; Stephan, D.; Murray, S.; Burger, J.T. The use of high-throughput small RNA sequencing reveals differentially expressed microRNAs in response to aster yellows phytoplasma-infection in Vitis vinifera cv. ‘Chardonnay’. PLoS ONE 2017, 12, e0182629. [Google Scholar] [CrossRef] [PubMed]
- Emamjomeh, A.; Saboori Robat, E.; Zahiri, J.; Solouki, M.; Khosravi, P. Gene co-expression network reconstruction: A review on computational methods for inferring functional information from plant-based expression data. Plant Biotechnol. Rep. 2017, 11, 71–86. [Google Scholar] [CrossRef]
- Škrlj, B.; Pompe Novak, M.; Brader, G.; Anžič, B.; Ramšak, Ž.; Gruden, K.; Kralj, J.; Lavrač, N.; Roitsch, T.; Dermastia, M. New cross-talks between pathways involved in grapevine infection with ‘Candidatus Phytoplasma solani’ revealed by temporal network modelling. Plants 2021, in press. [Google Scholar]
- Aryan, A.; Brader, G.; Mörtel, J.; Pastar, M.; Riedle-Bauer, M. An abundant ‘Candidatus Phytoplasma solani’ tuf b strain is associated with grapevine, stinging nettle and Hyalesthes obsoletus. Eur. J. Plant Pathol. Eur. Found. Plant Pathol. 2014, 140, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Murolo, S.; Garbarino, M.; Mancini, V.; Romanazzi, G. Spatial pattern of Bois noir: Case study of a delicate balance between disease progression and recovery. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Riedle-Bauer, M.; Hanak, K.; Regner, F.; Tiefenbrunner, W. Influence of pruning measures on recovery of Bois Noir-infected grapevines. J. Phytopathol. 2010, 158, 628–632. [Google Scholar] [CrossRef]
- Dermastia, M.; Kube, M.; Šeruga-Musić, M. Transcriptomic and proteomic studies of phytoplasma-infected plants. In Phytoplasmas: Plant Pathogenic Bacteria—III; Springer: Singapore, 2019; pp. 35–55. [Google Scholar]
- Williams, M.; Cheng, Y.Y.; Blenkiron, C.; Reid, G. Exploring Mechanisms of MicroRNA Downregulation in Cancer. MicroRNA 2017, 6, 2–16. [Google Scholar] [CrossRef]
- Zombardo, A.; Crosatti, C.; Bagnaresi, P.; Bassolino, L.; Reshef, N.; Puccioni, S.; Faccioli, P.; Tafuri, A.; Delledonne, M.; Fait, A.; et al. Transcriptomic and biochemical investigations support the role of rootstock-scion interaction in grapevine berry quality. BMC Genom. 2020, 21, 468. [Google Scholar] [CrossRef]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Krüger, P.; Selbig, J.; Müller, L.A.; Rhee, S.Y.; Stitt, M. MAPMAN: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef] [PubMed]
- Ballicora, M.A.; Iglesias, A.A.; Preiss, J. ADP-Glucose Pyrophosphorylase: A Regulatory Enzyme for Plant Starch Synthesis. Photosynth. Res. 2004, 79, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Mardi, M.; Karimi Farsad, L.; Gharechahi, J.; Salekdeh, G.H. In-depth transcriptome sequencing of Mexican lime trees infected with “Candidatus Phytoplasma aurantifolia”. PLoS ONE 2015, 10, e0130425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamurthy, P.; Muthusamy, M.; Kim, J.A.; Jeong, M.J.; Lee, S.I. Brassica rapa expansin-like B1 gene (BrEXLB1) regulate growth and development in transgenic Arabidopsis and elicits response to abiotic stresses. J. Plant Biochem. Biotechnol. 2019, 28, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Paolacci, A.R.; Catarcione, G.; Ederli, L.; Zadra, C.; Pasqualini, S.; Badiani, M.; Musetti, R.; Santi, S.; Ciaffi, M. Jasmonate-mediated defence responses, unlike salicylate-mediated responses, are involved in the recovery of grapevine from bois noir disease. BMC Plant Biol. 2017, 17, 118. [Google Scholar] [CrossRef] [Green Version]
- Sugio, A.; Kingdom, H.N.; MacLean, A.M.; Grieve, V.M.; Hogenhout, S.A. Phytoplasma protein effector SAP11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1254–E1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Music, M.S.; Samarzija, I.; Hogenhout, S.A.; Haryono, M.; Cho, S.T.; Kuo, C.H. The genome of ‘Candidatus Phytoplasma solani’ strain SA-1 is highly dynamic and prone to adopting foreign sequences. Syst. Appl. Microbiol. 2019, 42, 117–127. [Google Scholar] [CrossRef]
- Ren, H.; Gray, W.M. SAUR Proteins as Effectors of Hormonal and Environmental Signals in Plant Growth. Mol. Plant 2015, 8, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, A.; Oshima, K.; Kakizawa, S.; Ishii, Y.; Ozeki, J.; Hashimoto, M. A unique virulence factor for proliferation and dwarfism in plants identified from a phytopathogenic bacterium. Proc. Natl. Acad. Sci. USA 2009, 106, 6416–6421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minato, N.; Himeno, M.; Hoshi, A.; Maejima, K.; Komatsu, K.; Takebayashi, Y.; Kasahara, H.; Yusa, A.; Yamaji, Y.; Oshima, K.; et al. The phytoplasmal virulence factor TENGU causes plant sterility by downregulating of the jasmonic acid and auxin pathways. Sci. Rep. 2014, 4, 7399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes Ferreira, M.D.; Araújo Castro, J.; Santana Silva, R.J.; Micheli, F. HVA22 from citrus: A small gene family whose some members are involved in plant response to abiotic stress. Plant Physiol. Biochem. 2019, 142, 395–404. [Google Scholar] [CrossRef]
- Monavarfeshani, A.; Mirzaei, M.; Sarhadi, E.; Amirkhani, A.; Khayam Nekouei, M.; Haynes, P.A.; Mardi, M.; Salekdeh, G.H. Shotgun proteomic analysis of the Mexican lime tree infected with “Candidatus Phytoplasma aurantifolia”. J. Proteome Res. 2013, 12, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Musetti, R.; Paolacci, A.; Ciaffi, M.; Tanzarella, O.A.; Polizzotto, R.; Tubaro, F.; Mizzau, M.; Ermacora, P.; Badiani, M.; Osler, R. Phloem cytochemical modification and gene expression following the recovery of apple plants from apple proliferation disease. Phytopathology 2010, 100, 390–399. [Google Scholar] [CrossRef] [Green Version]
- Margaria, P.; Abbà, S.; Palmano, S. Novel aspects of grapevine response to phytoplasma infection investigated by a proteomic and phospho-proteomic approach with data integration into functional networks. BMC Genom. 2013, 14, 38. [Google Scholar] [CrossRef] [Green Version]
- Margaria, P.; Palmano, S. Response of the Vitis vinifera L. cv. “Nebbiolo” proteome to Flavescence dorée phytoplasma infection. Proteomics 2011, 11, 212–224. [Google Scholar] [CrossRef]
- Musetti, R.; Marabottini, R.; Badiani, M.; Martini, M.; Sanit, L.; Sanita di Toppi, L.; Borselli, S.; Borgo, M.; Osler, R. On the role of H 2 O 2 in the recovery of grapevine (Vitis vinifera cv. Prosecco ) from Flavescence dorée disease. Funct. Plant Biol. 2007, 34, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Dermastia, M.; Bertaccini, A.; Constable, F.; Mehle, N. Grapevine Yellows Diseases and Their Phytoplasma Agents: Biology and Detection; Springer: Cham, Switzerland, 2017; ISBN 9783319506487. [Google Scholar]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase properties, physiological functions, and evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fan, G.; Wang, Z.; Gu, Z. Phytoplasma-induced changes in the acetylome and succinylome of paulownia tomentosa provide evidence for involvement of acetylated proteins in witches’ broom disease. Mol. Cell. Proteom. 2019, 18, 1210–1226. [Google Scholar] [CrossRef] [PubMed]
- Musetti, R.; Buxa, S.V.; De Marco, F.; Loschi, A.; Polizzotto, R.; Kogel, K.-H.; van Bel, A.J.E. Phytoplasma-triggered Ca(2+) influx is involved in sieve-tube blockage. Mol. Plant. Microbe. Interact. 2013, 26, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takanashi, K.; Shitan, N.; Yazaki, K. The multidrug and toxic compound extrusion (MATE) family in plants. Plant Biotechnol. 2014, 31, 417–430. [Google Scholar] [CrossRef] [Green Version]
- Kusakizako, T.; Miyauchi, H.; Ishitani, R.; Nureki, O. Structural biology of the multidrug and toxic compound extrusion superfamily transporters. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183154. [Google Scholar] [CrossRef]
- Sun, X.; Gilroy, E.M.; Chini, A.; Nurmberg, P.L.; Hein, I.; Lacomme, C.; Birch, P.R.J.; Hussain, A.; Yun, B.-W.; Loake, G.J. ADS1 encodes a MATE-transporter that negatively regulates plant disease resistance. New Phytol. 2011, 192, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, H.; Pan, Y.; Yu, Y.; Luan, S.; Li, L. A DTX/MATE-type transporter facilitates abscisic acid efflux and modulates ABA sensitivity and drought tolerance in Arabidopsis. Mol. Plant 2014, 7, 1522–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Du, X. Identification, characterization and expression analysis of calmodulin and calmodulin-like proteins in Solanum pennellii. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Du, L.; Wang, H.; Yang, T. Recent advances in calcium/calmodulin-mediated signaling with an emphasis on plant-microbe interactions. Plant Physiol. 2013, 163, 531–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshiba, T.; Saito, E.; Ono, N.; Yamamoto, N.; Satô, M. Purification and properties of flavin- and molybdenum-containing aldehyde oxidase from coleoptiles of maize. Plant Physiol. 1996, 110, 781–789. [Google Scholar] [CrossRef]
- Mendel, R.-R.; Müller, A.J. A Common Genetic Determinant of Xanthine Dehydrogenase and Nitrate Reductase in Nicotiana tabacum. Biochem. Physiol. Pflanz. 1976, 170, 538–541. [Google Scholar] [CrossRef]
- Watanabe, S.; Sato, M.; Sawada, Y.; Tanaka, M.; Matsui, A.; Kanno, Y.; Hirai, M.Y.; Seki, M.; Sakamoto, A.; Seo, M. Arabidopsis molybdenum cofactor sulfurase ABA3 contributes to anthocyanin accumulation and oxidative stress tolerance in ABA-dependent and independent ways. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Mehle, N.; Dermastia, M.; Brus, R.; Jurc, D. First report of ‘Candidatus Phytoplasma ulmi’ in Ulmus minor and Ulmus glabra in Slovenia. Plant Dis. 2017, 101. [Google Scholar] [CrossRef]
- Le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant Sci. 2016, 7, 735. [Google Scholar] [CrossRef] [Green Version]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Ludwig-Müller, J.; Chung, G.; Ahmad, M.Q.; Yang, S.H.; Lee, S.I. Comparative genomic and transcriptomic analyses of Family-1 UDP glycosyltransferase in three Brassica species and Arabidopsis indicates stress-responsive regulation. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, X.; Chang, X.; Sun, M.; Zhang, Y.; Li, W.; Li, Y. Overexpression of germin-like protein GmGLP10 enhances resistance to Sclerotinia sclerotiorum in transgenic tobacco. Biochem. Biophys. Res. Commun. 2018, 497, 160–166. [Google Scholar] [CrossRef]
- Liu, N.; Shang, W.; Li, C.; Jia, L.; Wang, X.; Xing, G.; Zheng, W.M. Evolution of the SPX gene family in plants and its role in the response mechanism to phosphorus stress. Open Biol. 2018, 8, 170231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.-T.; Li, M.-Y.; Cheng, K.-T.; Tan, C.M.; Su, L.-W.; Lin, W.-Y.; Shih, H.-T.; Chiou, T.-J.; Yang, J.-Y. Transgenic plants that express the phytoplasma effector SAP11 show altered phosphate starvation and defense responses. Plant Physiol. 2014, 164, 1456–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef] [Green Version]
- Ouellette, A.J.; Hsieh, M.M.; Nosek, M.T.; Cano-Gauci, D.F.; Huttner, K.M.; Buick, R.N.; Selsted, M.E. Mouse Paneth cell defensins: Primary structures and antibacterial activities of numerous cryptdin isoforms. Infect. Immun. 1994, 62, 5040–5047. [Google Scholar] [CrossRef] [Green Version]
- Ouellette, A.J.; Satchell, D.P.; Hsieh, M.M.; Hagen, S.J.; Selsted, M.E. Characterization of luminal Paneth cell α-defensins in mouse small intestine: Attenuated antimicrobial activities of peptides with truncated amino termini. J. Biol. Chem. 2000, 275, 33969–33973. [Google Scholar] [CrossRef] [Green Version]
- Ayabe, T.; Satchell, D.P.; Wilson, C.L.; Parks, W.C.; Selsted, M.E.; Ouellette, A.J. Secretion of microbicidal α-defensins by intestinal Paneth cells in response to bacteria. Nat. Immunol. 2000, 1, 113–118. [Google Scholar] [CrossRef]
- Carra, A.; Gambino, G.; Schubert, A. A cetyltrimethylammonium bromide-based method to extract low-molecular-weight RNA from polysaccharide-rich plant tissues. Anal. Biochem. 2007, 360, 318–320. [Google Scholar] [CrossRef]
- R Core Team. R A Language and Environment for Statistical Computing. Scientific Research Publishing. 2017. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=2144573 (accessed on 21 January 2021).
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.; Agrawal, S.; Birney, E.; Bruford, E.A.; Bujnicki, J.M.; Cochrane, G.; Cole, J.R.; Dinger, M.E.; Enright, A.J.; Gardner, P.P.; et al. RNAcentral: A vision for an international database of RNA sequences. RNA 2011, 17, 1941–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, S.; Axtell, M.J. Identification and annotation of small RNA genes using ShortStack. Methods 2014, 67, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Sun, Y. MiR-PREFeR: An accurate, fast and easy-to-use plant miRNA prediction tool using small RNA-Seq data. Bioinformatics 2014, 30, 2837–2839. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef]
- De Oliveira, L.F.V.; Christoff, A.P.; Margis, R. isomiRID: A framework to identify microRNA isoforms. Bioinformatics 2013, 29, 2521–2523. [Google Scholar] [CrossRef] [Green Version]
- Gebert, D.; Hewel, C.; Rosenkranz, D. Unitas: The universal tool for annotation of small RNAs. BMC Genom. 2017, 18, 644. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. PsRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Ramšak, Ž.; Baebler, Š.; Rotter, A.; Korbar, M.; Mozetič, I.; Usadel, B.; Gruden, K. GoMapMan: Integration, consolidation and visualization of plant gene annotations within the MapMan ontology. Nucleic Acids Res. 2014, 42, D1167–D1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosvall, M.; Axelsson, D.; Bergstrom, C.T. The map equation. Eur. Phys. J. Spec. Top. 2009, 178, 13–23. [Google Scholar] [CrossRef]
- Hagberg, A.; Schult, D.; Swart, P. Exploring network structure, dynamics, and function using networkX. In Proceedings of the 7th Python in Science Conference, Pasadena, CA, USA, 19–24 August 2008; pp. 11–15. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Baebler, Š.; Svalina, M.; Petek, M.; Stare, K.; Rotter, A.; Pompe-Novak, M.; Gruden, K. QuantGenius: Implementation of a decision support system for qPCR-based gene quantification. BMC Bioinform. 2017, 18, 276. [Google Scholar] [CrossRef] [PubMed]
- Jammer, A.; Gasperl, A.; Luschin-Ebengreuth, N.; Heyneke, E.; Chu, H.; Cantero-Navarro, E.; Großkinsky, D.K.; Albacete, A.A.; Stabentheiner, E.; Franzaring, J.; et al. Simple and robust determination of the activity signature of key carbohydrate metabolism enzymes for physiological phenotyping in model and crop plants. J. Exp. Bot. 2015, 66, 5531–5542. [Google Scholar] [CrossRef] [Green Version]
- Anžič, B. Transformation of Periwinkle (Catharanthus roseus) for Studying Infection with ’Candidatus Phytoplasma Solani’ and Enzymatic Activities in Infectedon of Madagascar Plants, University of Ljubljana. Available online: https://repozitorij.uni-lj.si/IzpisGradiva.php?id=111641&lang=slv (accessed on 29 February 2021).
- Fimognari, L.; Dölker, R.; Kaselyte, G.; Jensen, C.N.G.; Akhtar, S.S.; Großkinsky, D.K.; Roitsch, T. Simple semi-high throughput determination of activity signatures of key antioxidant enzymes for physiological phenotyping. Plant Methods 2020, 16, 42. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Description | Log2 FC | |||
|---|---|---|---|---|---|
| E-I : E-U | L-I : L-U | L-U : E-U | L-I : E-I | ||
| Early/Late | |||||
| Vitvi02g01406 | Thaumatin family | 6.62 | 3.27 | 5.52 | 2.18 |
| Vitvi06g01696 | Metallothionein | 1.86 | 2.88 | 1.14 | 2.16 |
| Vitvi18g00740 | Granulin repeat cysteine protease family protein | 0.93 | 1.19 | 1.72 | 1.98 |
| Vitvi19g00434 | Ubiquitin family protein | 0.93 | 0.31 | 2.10 | 1.48 |
| Vitvi05g01756 | Pathogenesis-related protein 10 | 1.50 | −0.27 | 1.91 | 0.13 |
| Vitvi02g00605 | Chloroplast β-amylase | 0.56 | −0.40 | 5.71 | 4.75 |
| Vitvi06g01697 | Metallothionein | 0.55 | 1.02 | −0.43 | 0.05 |
| Vitvi02g01341 | Cellulose synthase-like G3 | 0.73 | 1.13 | 4.34 | 4.75 |
| Vitvi03g00327 | Cold circadian rhythm and RNA binding 2 | 0.01 | 0.28 | 1.81 | 2.07 |
| Vitvi07g01690 | Cysteine proteinase1 | 1.04 | 0.96 | 2.05 | 1.98 |
| Uninfected/Infected | |||||
| Vitvi19g01871 | Metallothionein 3 | −1.68 | −0.47 | 0.26 | 1.48 |
| Vitvi02g00605 | Chloroplast β-amylase | 0.56 | −0.40 | 5.71 | 4.75 |
| Vitvi08g01245 | Rubisco activase | −1.14 | −1.80 | 2.25 | 1.59 |
| Vitvi01g00714 | Galactinol synthase 4 | −0.55 | 0.07 | 0.87 | 1.49 |
| Vitvi19g00549 | GDP-L-galactose phosphorylase vitamin C defective 5 | −1.13 | −1.51 | 0.54 | 0.16 |
| Vitvi17g00038 | CLPC homolog 1 | −0.24 | −1.05 | 2.01 | 1.19 |
| Vitvi05g00563 | Early light-induced protein 1, chloroplastic-related | 0.24 | −0.73 | 2.00 | 1.03 |
| Vitvi17g00320 | Ribulose bisphosphate carboxylase (small chain) family protein | −1.14 | −1.67 | −0.28 | −0.81 |
| Vitvi19g00434 | Ubiquitin family protein | 0.93 | 0.31 | 2.10 | 1.48 |
| Vitvi06g00513 | Rubisco activase | −1.48 | −1.84 | −0.35 | −0.71 |
| miRNA ID | Log2 FC | |||
|---|---|---|---|---|
| E-I : E-U | L-I : L-U | L-U : E-U | L-I : E-I | |
| Early/Late | ||||
| vvi-miR166c-h | −1.24 | −1.18 | 0.37 | 0.43 |
| vvi-miR162 | −1.05 | −0.77 | −0.05 | 0.24 |
| vvi-miR3623.5 | 0.05 | 2.33 | 0.65 | 2.92 |
| vvi-miR3624-3p | 0.94 | 0.83 | 1.08 | 0.96 |
| vvi-miR3623.4 | −0.20 | −0.67 | 0.83 | 0.35 |
| vvi-miR159c | −0.34 | −0.06 | −0.26 | 0.02 |
| vvi-miR162.3 | −0.84 | −0.50 | −0.01 | 0.33 |
| vvi-miR3623-5p | 0.50 | 2.47 | 0.50 | 2.47 |
| vvi-miR159c.1 | −0.36 | −0.07 | −0.16 | 0.12 |
| vvi-miR3634.3 | −1.09 | −1.84 | −0.14 | −0.89 |
| Uninfected/Infected | ||||
| vvi-miR3624-3p | 0.94 | 0.83 | 1.08 | 0.96 |
| vvi-miR3623.5 | 0.05 | 2.33 | 0.65 | 2.92 |
| vvi-miR3623-5p | 0.50 | 2.47 | 0.50 | 2.47 |
| vvi-miR166c-h | −1.24 | −1.18 | 0.37 | 0.43 |
| vvi-miR156g.1 | −0.42 | −0.18 | 1.74 | 1.97 |
| vvi-miR482 | −0.05 | 1.33 | −0.05 | 1.34 |
| vvi-miR398b,c | −0.54 | −0.43 | 1.68 | 1.79 |
| vvi-miR166d.2 | −0.63 | −1.67 | 2.08 | 1.04 |
| vvi-miR482.4 | 0.98 | 0.31 | 1.00 | 0.32 |
| vvi-miR168.5 | 1.05 | 0.58 | 1.01 | 0.55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dermastia, M.; Škrlj, B.; Strah, R.; Anžič, B.; Tomaž, Š.; Križnik, M.; Schönhuber, C.; Riedle-Bauer, M.; Ramšak, Ž.; Petek, M.; et al. Differential Response of Grapevine to Infection with ‘Candidatus Phytoplasma solani’ in Early and Late Growing Season through Complex Regulation of mRNA and Small RNA Transcriptomes. Int. J. Mol. Sci. 2021, 22, 3531. https://doi.org/10.3390/ijms22073531
Dermastia M, Škrlj B, Strah R, Anžič B, Tomaž Š, Križnik M, Schönhuber C, Riedle-Bauer M, Ramšak Ž, Petek M, et al. Differential Response of Grapevine to Infection with ‘Candidatus Phytoplasma solani’ in Early and Late Growing Season through Complex Regulation of mRNA and Small RNA Transcriptomes. International Journal of Molecular Sciences. 2021; 22(7):3531. https://doi.org/10.3390/ijms22073531
Chicago/Turabian StyleDermastia, Marina, Blaž Škrlj, Rebeka Strah, Barbara Anžič, Špela Tomaž, Maja Križnik, Christina Schönhuber, Monika Riedle-Bauer, Živa Ramšak, Marko Petek, and et al. 2021. "Differential Response of Grapevine to Infection with ‘Candidatus Phytoplasma solani’ in Early and Late Growing Season through Complex Regulation of mRNA and Small RNA Transcriptomes" International Journal of Molecular Sciences 22, no. 7: 3531. https://doi.org/10.3390/ijms22073531
APA StyleDermastia, M., Škrlj, B., Strah, R., Anžič, B., Tomaž, Š., Križnik, M., Schönhuber, C., Riedle-Bauer, M., Ramšak, Ž., Petek, M., Kladnik, A., Lavrač, N., Gruden, K., Roitsch, T., Brader, G., & Pompe-Novak, M. (2021). Differential Response of Grapevine to Infection with ‘Candidatus Phytoplasma solani’ in Early and Late Growing Season through Complex Regulation of mRNA and Small RNA Transcriptomes. International Journal of Molecular Sciences, 22(7), 3531. https://doi.org/10.3390/ijms22073531