
Effects of Edible Insect Tenebrio molitor Larva Fermentation Extract as a Substitute Protein on Hepatosteatogenesis and Proteomic Changes in Obese Mice Induced by High-Fat Diet



Abstract
:1. Introduction
2. Results
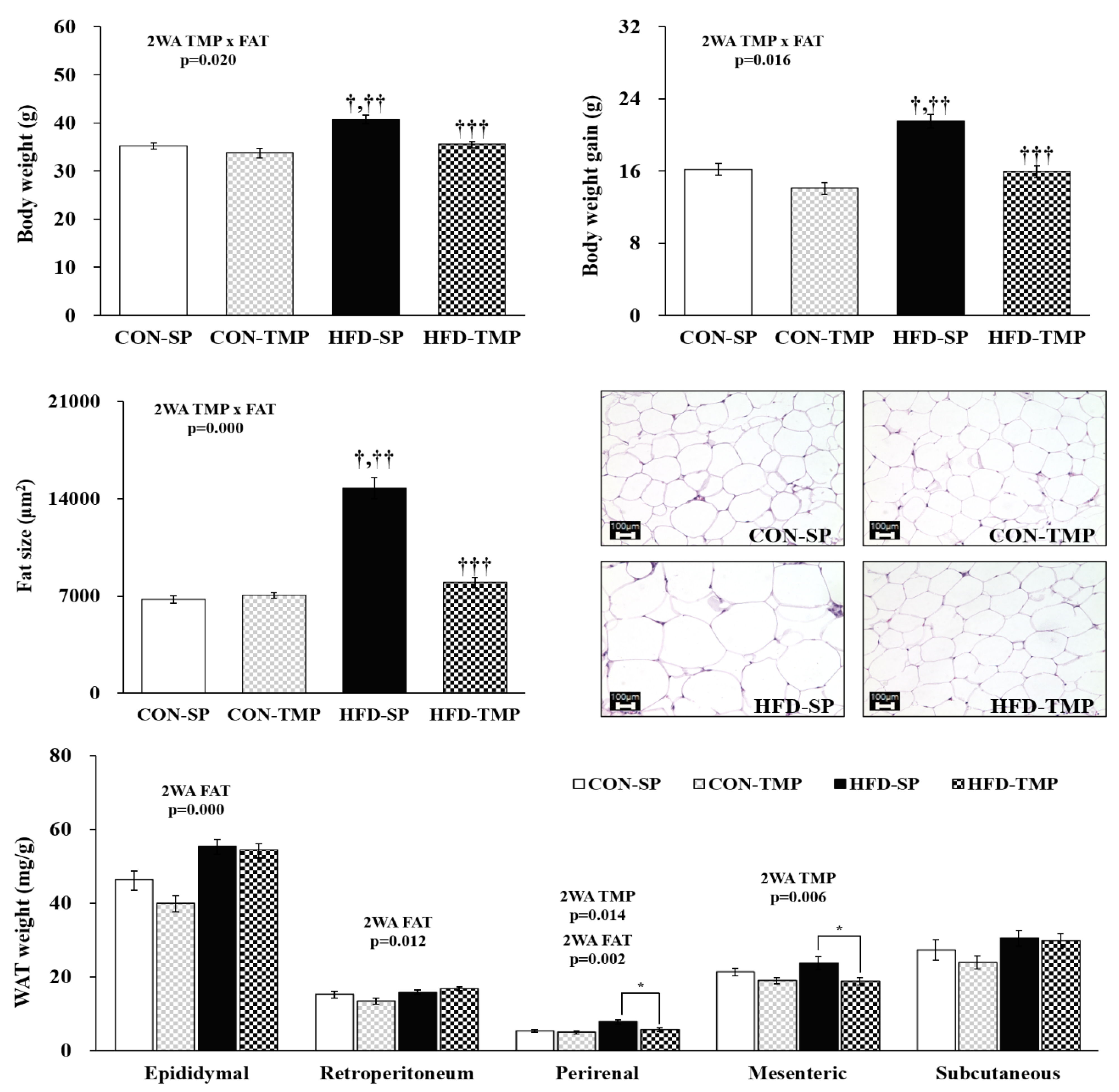
2.1. TMP Reduced the Body Weight and Fat Size
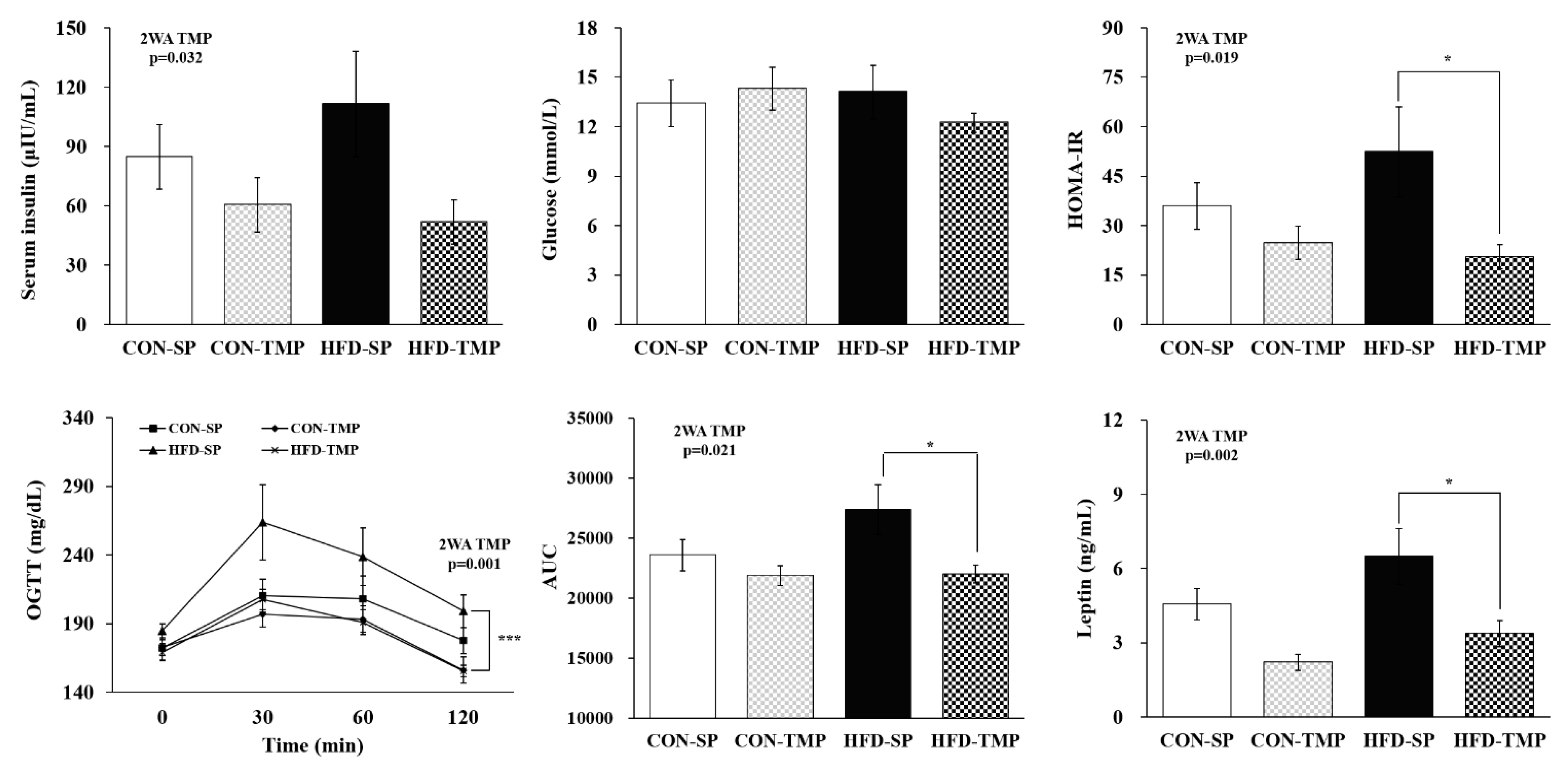
2.2. TMP Affected the Insulin Resistance Induced by High-Fat Diet
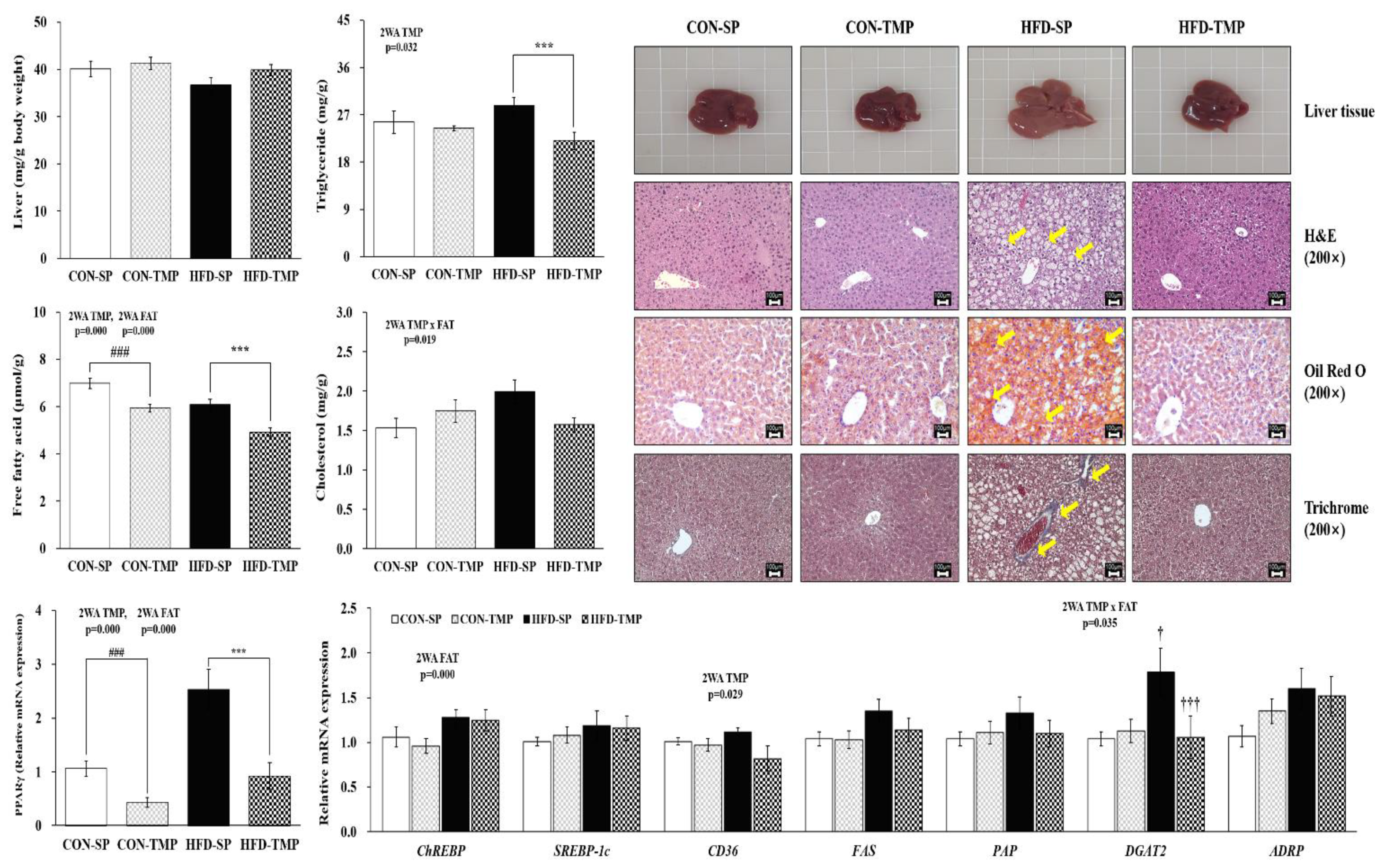
2.3. TMP Attenuated Hepatic Steatosis Induced by High-Fat Diet
2.4. TMP Modified Hepatic Changes of Proteomic Profiles
2.5. TMP Has a Similar Effect on the Serum Lipid Contents and Inflammation Markers to SP
3. Discussion
4. Materials and Methods
4.1. TMP Preparation
4.2. Animal Experiments
4.3. Oral Glucose Tolerance Test
4.4. Serum Parameters
4.5. Lipid Profile Evaluation
4.6. Histological Analysis
4.7. RNA Isolation and Quantitative Real-Time PCR Analysis
4.8. Protein Sample Preparation
4.9. Two-Dimensional Polyacrylamide Gel Electrophoresis (2-DE)
4.10. Peptide Mass Fingerprinting (PMF)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anstee, Q.M.; Reeves, H.L.; Kotsiliti, E.; Govaere, O.; Heikenwalder, M. From NASH to HCC: Current concepts and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Berg, E.H.V.D.; Amini, M.; Schreuder, T.C.M.A.; Dullaart, R.P.F.; Faber, K.N.; Alizadeh, B.Z.; Blokzijl, H. Prevalence and determinants of non-alcoholic fatty liver disease in lifelines: A large Dutch population cohort. PLoS ONE 2017, 12, e0171502. [Google Scholar] [CrossRef]
- Ong, J.P.; Elariny, H.; Collantes, R.; Younoszai, A.; Chandhoke, V.; Reines, H.D.; Goodman, Z.; Younossi, Z.M. Predictors of nonalcoholic steatohepatitis and advanced fibrosis in morbidly obese patients. Obes. Surg. 2005, 15, 310–315. [Google Scholar] [CrossRef]
- Eguchi, Y.; Nafld, J.; Hyogo, H.; Ono, M.; Mizuta, T.; Ono, N.; Fujimoto, K.; Chayama, K.; Saibara, T. Prevalence and associated metabolic factors of nonalcoholic fatty liver disease in the general population from 2009 to 2010 in Japan: A multicenter large retrospective study. J. Gastroenterol. 2012, 47, 586–595. [Google Scholar] [CrossRef]
- Chang, Y.; Jung, H.-S.; Cho, J.; Zhang, Y.; Yun, K.E.; Lazo, M.; Pastor-Barriuso, R.; Ahn, J.; Kim, C.-W.; Rampal, S.; et al. Metabolically healthy obesity and the development of nonalcoholic fatty liver disease. Am. J. Gastroenterol. 2016, 111, 1133–1140. [Google Scholar] [CrossRef]
- Pang, Q.; Zhang, J.-Y.; Song, S.-D.; Qu, K.; Xu, X.-S.; Liu, S.-S.; Liu, C. Central obesity and nonalcoholic fatty liver disease risk after adjusting for body mass index. World J. Gastroenterol. 2015, 21, 1650–1662. [Google Scholar] [CrossRef] [PubMed]
- Van Thielen, L.; Vermuyten, S.; Storms, B.; Rumpold, B.; Van Campenhout, L. Consumer acceptance of foods containing edible insects in Belgium two years after their introduction to the market. J. Insects Food Feed. 2019, 5, 35–44. [Google Scholar] [CrossRef]
- Im, A.-R.; Yang, W.-K.; Park, Y.-C.; Kim, S.H.; Chae, S. Hepatoprotective effects of insect extracts in an animal model of nonalcoholic fatty liver disease. Nutrients 2018, 10, 735. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Han, T.; Kim, Y. Mealworm (Tenebrio molitor Larvae) as an alternative protein source for monogastric animal: A review. Animals 2020, 10, 2068. [Google Scholar] [CrossRef]
- Gessner, D.K.; Schwarz, A.; Meyer, S.; Wen, G.; Most, E.; Zorn, H.; Ringseis, R.; Eder, K. Insect meal as alternative protein source exerts pronounced lipid-lowering effects in hyperlipidemic obese Zucker rats. J. Nutr. 2019, 149, 566–577. [Google Scholar] [CrossRef]
- Sim, S.; Jang, S.; Cho, Y.; Ahn, H. Effect of Tenebrio molitor larvae fermented with Saccharomyces cerevisiae M1 (KACC 93023) on non-alcoholic fatty liver-induced rats. J. Life Sci. 2020, 30, 434–442. [Google Scholar]
- Choi, R.-Y.; Ham, J.R.; Ryu, H.-S.; Lee, S.S.; Miguel, M.A.; Paik, M.-J.; Ji, M.; Park, K.-W.; Kang, K.-Y.; Lee, H.-I.; et al. Defatted Tenebrio molitor larva fermentation extract modifies steatosis, inflammation and intestinal microflora in chronic alcohol-fed rats. Nutrients 2020, 12, 1426. [Google Scholar] [CrossRef]
- Shang, X.; Scott, D.; Hodge, A.M.; English, D.R.; Giles, G.G.; Ebeling, P.R.; Sanders, K.M. Dietary protein intake and risk of type 2 diabetes: Results from the Melbourne collaborative cohort study and a meta-analysis of prospective studies. Am. J. Clin. Nutr. 2016, 104, 1352–1365. [Google Scholar] [CrossRef]
- Navas-Carretero, S.; San-Cristobal, R.; Livingstone, K.M.; Celis-Morales, C.; Marsaux, C.F.; Macready, A.L.; Fallaize, R.; O’Donovan, C.B.; Forster, H.; Woolhead, C.; et al. Higher vegetable protein consumption, assessed by an isoenergetic macronutrient exchange model, is associated with a lower presence of overweight and obesity in the web-based Food4me European study. Int. J. Food Sci. Nutr. 2019, 70, 240–253. [Google Scholar] [CrossRef] [Green Version]
- De Chiara, F.; Checcllo, C.U.; Azcón, J.R. High protein diet and metabolic plasticity in non-alcoholic fatty liver disease: Myths and truths. Nutrients 2019, 11, 2985. [Google Scholar] [CrossRef] [Green Version]
- Soltis, A.R.; Kennedy, N.J.; Xin, X.; Zhou, F.; Ficarro, S.B.; Yap, Y.S.; Matthews, B.J.; Lauffenburger, D.A.; White, F.M.; Marto, J.A. Hepatic dysfunction caused by consumption of a high-fat diet. Cell Rep. 2017, 21, 3317–3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, S.; Mulligan, K.; Schwarz, J.M. Dietary carbohydrates and fatty liver disease: De novo lipogenesis. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Softic, S.; Gupta, M.K.; Wang, G.-X.; Fujisaka, S.; O’Neill, B.T.; Rao, T.N.; Willoughby, J.; Harbison, C.; Fitzgerald, K.; Ilkayeva, O.; et al. Divergent effects of glucose and fructose on hepatic lipogenesis and insulin signaling. J. Clin. Investig. 2017, 127, 4059–4074. [Google Scholar] [CrossRef] [Green Version]
- Andrikopoulos, S.; Proietto, J. The biochemical basis of increased hepatic glucose production in a mouse model of type 2 (non-insulin-dependent) diabetes mellitus. Diabetologia 1995, 38, 1389–1396. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Andrikopoulos, S.; Filippis, C.; Thorburn, A.W.; Khan, D.; Proietto, J. Mechanism of fat-induced hepatic gluconeogenesis: Effect of metformin. Am. J. Physiol. Metab. 2001, 281, E275–E282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, J.-C.; Converset, V.; Nolan, A.; Schmid, G.; Wang, S.; Heller, M.; Sennitt, M.V.; Hochstrasser, D.F.; Cawthorne, M.A. Effect of rosiglitazone on the differential expression of diabetes-associated proteins in pancreatic islets of C57Bl/6 lep/lep mice. Mol. Cell. Proteom. 2002, 1, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Thornalley, P.J. Glyoxalase in Diabetes, Obesity and Related Disorders, Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2011; pp. 309–317. [Google Scholar]
- Xu, C.; Zhang, X.; Yu, C.; Lu, G.; Chen, S.; Xu, L.; Ding, W.; Shi, Q.; Li, Y. Proteomic analysis of hepatic ischemia/reperfusion injury and ischemic preconditioning in mice revealed the protective role of ATP5β. Proteomics 2009, 9, 409–419. [Google Scholar] [CrossRef]
- Ogawa, T.; Kimoto, M.; Sasaoka, K. Occurrence of a new enzyme catalyzing the direct conversion of NG,NG-dimethyl-L-arginine to L-citrulline in rats. Biochem. Biophys. Res. Commun. 1987, 148, 671–677. [Google Scholar] [CrossRef]
- Hui, X.; Zhu, W.; Wang, Y.; Lam, K.S.L.; Zhang, J.; Wu, D.; Kraegen, E.W.; Li, Y.; Xu, A. Major urinary protein-1 increases energy expenditure and improves glucose intolerance through enhancing mitochondrial function in skeletal muscle of diabetic mice. J. Biol. Chem. 2009, 284, 14050–14057. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Jiang, L.; Rui, L. Identification of MUP1 as a regulator for glucose and lipid metabolism in mice. J. Biol. Chem. 2009, 284, 11152–11159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- De Chiara, F.; Heebøll, S.; Marrone, G.; Montoliu, C.; Hamilton-Dutoit, S.; Ferrandez, A.; Andreola, F.; Rombouts, K.; Grønbæk, H.; Felipo, V.; et al. Urea cycle dysregulation in non-alcoholic fatty liver disease. J. Hepatol. 2018, 69, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, P.L.; Vilstrup, H.; Rigbolt, K.; Suppli, M.P.; Sørensen, M.; Heebøll, S.; Veidal, S.S.; Knop, F.K.; Thomsen, K.L. Non-alcoholic fatty liver disease alters expression of genes governing hepatic nitrogen conversion. Liver Int. 2019, 39, 2094–2101. [Google Scholar] [CrossRef]
- De Chiara, F.; Thomsen, K.L.; Habtesion, A.; Jones, H.; Davies, N.; Gracia-Sancho, J.; Manicardi, N.; Hall, A.; Andreola, F.; Paish, H.L.; et al. Ammonia scavenging prevents progression of fibrosis in experimental nonalcoholic fatty liver disease. Hepatology 2020, 71, 874–892. [Google Scholar] [CrossRef]
- Anderson, N.L.; Esquer-Blasco, R.; Hofmann, J.-P.; Meheus, L.; Raymackers, J.; Steiner, S.; Witzmann, F.; Anderson, N.G.; Anderson, N.L.; Anderson, N.G. An updated two-dimensional gel database of rat liver proteins useful in gene regulation and drug effect studies. Electrophoresis 1995, 16, 1977–1981. [Google Scholar] [CrossRef]
- Santos, C.X.; Tanaka, L.Y.; Wosniak, J., Jr.; Laurindo, F.R. Mechanisms and implications of reactive oxygen species generation during the unfolded protein response: Roles of endoplasmic reticulum oxidoreductases, mitochondrial electron transport, and NADPH oxidase. Antioxid. Redox Signal. 2009, 11, 2409–2427. [Google Scholar] [CrossRef]
- Shimizu, Y.; Hendershot, L.M. Oxidative folding: Cellular strategies for dealing with the resultant equimolar production of reactive oxygen species. Antioxid. Redox Signal. 2009, 11, 2317–2331. [Google Scholar] [CrossRef]
- Hanada, S.; Harada, M.; Kumemura, H.; Omary, M.B.; Koga, H.; Kawaguchi, T.; Taniguchi, E.; Yoshida, T.; Hisamoto, T.; Yanagimoto, C.; et al. Oxidative stress induces the endoplasmic reticulum stress and facilitates inclusion formation in cultured cells. J. Hepatol. 2007, 47, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Piao, J.-H.; Nakajima, A.; Sakon-Komazawa, S.; Kojima, Y.; Mori, K.; Yagita, H.; Okumura, K.; Harding, H.; Nakano, H. Tumor necrosis factor α (TNFα) induces the unfolded protein response (UPR) in a reactive oxygen species (ROS)-dependent fashion, and the UPR counteracts ROS accumulation by TNFα. J. Biol. Chem. 2005, 280, 33917–33925. [Google Scholar] [CrossRef] [Green Version]
- Yokouchi, M.; Hiramatsu, N.; Hayakawa, K.; Okamura, M.; Du, S.; Kasai, A.; Takano, Y.; Shitamura, A.; Shimada, T.; Yao, J.; et al. Involvement of selective reactive oxygen species upstream of proapoptotic branches of unfolded protein response. J. Biol. Chem. 2008, 283, 4252–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, J.; Kaneda, S.; Kuwabara, K.; Ogawa, S.; Kobayashi, T.; Matsumoto, M.; Yura, T.; Yanagi, H. Cloning and expression of cDNA encoding the human 150 kDa oxygen-regulated protein, ORP150. Biochem. Biophys. Res. Commun. 1997, 230, 94–99. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ogawa, S.; Yura, T.; Yanagi, H. Abundant expression of 150-kDa oxygen-regulated protein in mouse pancreatic beta cells is correlated with insulin secretion. Biochem. Biophys. Res. Commun. 2000, 267, 831–837. [Google Scholar] [CrossRef]
- Ozawa, K.; Miyazaki, M.; Matsuhisa, M.; Takano, K.; Nakatani, Y.; Hatazaki, M.; Tamatani, T.; Yamagata, K.; Miyagawa, J.-I.; Kitao, Y.; et al. The endoplasmic reticulum chaperone improves insulin resistance in type 2 diabetes. Diabetes 2005, 54, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Sanson, M.; Augé, N.; Vindis, C.; Muller, C.; Bando, Y.; Thiers, J.; Marachet, M.; Zarkovic, K.; Sawa, Y.; Salvayre, R. Oxidized low-density lipoproteins trigger endoplasmic reticulum stress in vascular cells: Prevention by oxygen-regulated protein 150 expression. Circ. Res. 2009, 104, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, Z.; Li, D.; Wang, D.; Wang, X.; Feng, X.; Xia, M. Involvement of oxygen-regulated protein 150 in AMP-activated protein kinase-mediated alleviation of lipid-induced endoplasmic reticulum stress. J. Biol. Chem. 2011, 286, 11119–11131. [Google Scholar] [CrossRef] [Green Version]
- Jang, I.; Pottekat, A.; Poothong, J.; Yong, J.; Lagunas-Acosta, J.; Charbono, A.; Chen, Z.; Scheuner, D.L.; Liu, M.; Itkin-Ansari, P.; et al. PDIA1/P4HB is required for efficient proinsulin maturation and ß cell health in response to diet induced obesity. eLife 2019, 8, 44528. [Google Scholar] [CrossRef]
- Lian, J.; Wei, E.; Groenendyk, J.; Das, S.K.; Hermansson, M.; Li, L.; Watts, R.; Thiesen, A.; Oudit, G.Y.; Michalak, M.; et al. Ces3/TGH deficiency attenuates steatohepatitis. Sci. Rep. 2016, 6, 25747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raabe, M.; Véniant, M.M.; Sullivan, M.A.; Zlot, C.H.; Björkegren, J.; Nielsen, L.B.; Wong, J.S.; Hamilton, R.L.; Young, S.G. Analysis of the role of microsomal triglyceride transfer protein in the liver of tissue-specific knockout mice. J. Clin. Investig. 1999, 103, 1287–1298. [Google Scholar] [CrossRef] [Green Version]
- Wei, E.; Ben Ali, Y.; Lyon, J.; Wang, H.; Nelson, R.; Dolinsky, V.W.; Dyck, J.R.; Mitchell, G.; Korbutt, G.S.; Lehner, R. Loss of TGH/Ces3 in mice decreases blood lipids, improves glucose tolerance, and increases energy expenditure. Cell Metab. 2010, 11, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Lian, J.; Wei, E.; Wang, S.P.; Quiroga, A.D.; Li, L.; Di Pardo, A.; Van Der Veen, J.; Sipione, S.; Mitchell, G.A.; Lehner, R. Liver specific inactivation of carboxylesterase 3/triacylglycerol hydrolase decreases blood lipids without causing severe steatosis in mice. Hepatology 2012, 56, 2154–2162. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bahk, Y.Y.; Kim, S.A.; Kim, J.-S.; Euh, H.-J.; Bai, G.-H.; Cho, S.-N.; Kim, Y.S. Antigens secreted from Mycobacterium tuberculosis: Identification by proteomics approach and test for diagnostic marker. Proteomics 2004, 4, 3299–3307. [Google Scholar] [CrossRef]
- Lee, Y.; Hwang, Y.-H.; Kim, K.-J.; Park, A.-K.; Paik, M.-J.; Kim, S.H.; Lee, S.U.; Yee, S.-T.; Son, Y.-J. Proteomic and transcriptomic analysis of lung tissue in OVA-challenged mice. Arch. Pharmacal Res. 2018, 41, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.; Gharahdaghi, F.; Mische, S.M. Routine identification of proteins from sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels or polyvinyl difluoride membranes using matrix assisted laser desorption/ionization-time of flight-mass spectrometry (MALDI-TOF-MS). Electrophoresis 1998, 19, 1036–1045. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Symbol | Protein Description | Accession | Fold Change Protein Ratio | Protein Sequence Coverage (%) b | MW | pI | Score c | ||
|---|---|---|---|---|---|---|---|---|---|---|
| B/A | C/B | p-Value of C/B a | ||||||||
| 1 | ALDOB | Fructose-bisphosphate aldolase B | NP_659152.1 | 1.9 | 0.05 | 0.306 | 45 | 24 | 8.52 | 148 |
| 2 | ADH | Alcohol dehydrogenase 1 | NP_031435.1 | 2.38 | 0.17 | 0.335 | 32 | 39 | 8.44 | 114 |
| 3 | KHK | Ketohexokinase | NP_032465.2 | 1.57 | 0.62 | 0.019 | 63 | 30 | 5.81 | 158 |
| 4 | FBP1 | Fructose-1,6-bisphosphatase 1 | NP_062268.1 | 1.6 | 0.59 | 0.057 | 61 | 46 | 6.15 | 242 |
| 5 | PC | Pyruvate carboxylase, mitochondrial | NP_000911.2 | 1.69 | 0.61 | 0.404 | 46 | 62 | 6.25 | 447 |
| 6 | GLO1 | Lactoylglutathione lyase (=glyoxalase 1) | NP_079650.3 | 1.44 | 0.43 | 0.029 | 72 | 38 | 5.24 | 182 |
| 7 | hsc71 | Heat shock cognate 71 kDa | NP_112442.2 | 3.23 | 0.39 | 0.311 | 44 | 46 | 5.37 | 224 |
| 8 | HSPA9 | Stress-70 protein | NP_034611.2 | 2.73 | 0.26 | 0.277 | 37 | 72 | 5.81 | 274 |
| 9 | BHMT | Betaine—homocysteine S-methyltransferase 1 | NP_057877.1 | 2.78 | 0.07 | 0.351 | 57 | 56 | 8.01 | 160 |
| 10 | PHB | Prohibitin | NP_032857.1 | 1.82 | 0.22 | 0.138 | 53 | 20 | 5.57 | 181 |
| 11 | MAT1A | S-adenosylmethionine synthase isoform type-1 | NP_598414.1 | 1.6 | 0.47 | 0.089 | 48 | 53 | 5.51 | 183 |
| 12 | BCKDHA | 2-oxoisovalerate dehydrogenase subunit alpha | NP_031559.3 | 1.91 | 0.63 | 0.117 | 48 | 45 | 8.15 | 160 |
| 13 | hutU | Urocanate hydratase | NP_659189.2 | 1.47 | 0.62 | 0.275 | 41 | 44 | 7.27 | 290 |
| 14 | HMGCS2 | Hydroxymethylglutaryl-CoA synthase, mitochondrial | NP_032282.2 | 1.65 | 0.1 | 0.281 | 37 | 42 | 8.65 | 120 |
| 15 | ATP5H | ATP synthase subunit d, mitochondrial | NP_082138.1 | 1.69 | 0.68 | 0.035 | 65 | 49 | 5.52 | 108 |
| 16 | SOD | Superoxide dismutase | NP_035564.1 | 1.93 | 0.42 | 0.022 | 42 | 10 | 6.02 | 108 |
| 17 | DDAH1 | N(G),N(G)-dimethylarginine dimethylaminohydrolase 1 | NP_081269.1 | 1.57 | 0.18 | 0.022 | 35 | 25 | 5.64 | 88 |
| 18 | GSTM2 | Glutathione S-transferase Mu 7 | NP_080948.2 | 1.79 | 0.6 | 0.542 | 49 | 56 | 6.34 | 146 |
| 19 | ALDH1A1 | Retinal dehydrogenase 1 | NP_038495.2 | 1.46 | 0.21 | 0.299 | 37 | 63 | 7.92 | 123 |
| 20 | IMMT | MICOS complex subunit MIC60 | NP_001240617.1 | 1.78 | 0.56 | 0.631 | 33 | 44 | 7.65 | 135 |
| No. | Symbol | Protein Description | Accession | Fold Change Protein Ratio | Protein Sequence Coverage (%) b | MW | pI | Score c | ||
|---|---|---|---|---|---|---|---|---|---|---|
| B/A | C/B | p-Value of C/B a | ||||||||
| 21 | ACO1 | Cytoplasmic aconitate hydratase | NP_031412.2 | 0.49 | 2.03 | 0.141 | 48 | 56 | 7.23 | 311 |
| 22 | DLD | Dihydrolipoyl dehydrogenase, mitochondrial | NP_031887.2 | 0.43 | 5.62 | 0.027 | 52 | 45 | 7.99 | 172 |
| 23 | Mup1 | Major urinary protein 1 | NP_001334083.1 | 0.49 | 18.3 | 0.034 | 66 | 34 | 4.96 | 130 |
| 24 | CPS1 | Carbamoyl-phosphate synthase ammonia, mitochondrial | NP_001074278.1 | 0.29 | 7.26 | 0.019 | 28 | 51 | 6.48 | 249 |
| 25 | Ces3b | Carboxylesterase 3B | NP_001152887.1 | 0.34 | 3.49 | 0.000 | 20 | 28 | 5.65 | 84 |
| 26 | Ces3a | Isoform 2 of carboxylesterase 3A | NP_001158153.1 | 0.46 | 2.73 | 0.069 | 34 | 35 | 5.43 | 123 |
| 27 | PDI | Protein disulfide-isomerase | NP_035162.1 | 0.62 | 2.47 | 0.043 | 50 | 42 | 4.77 | 223 |
| 28 | HSPA5 | Endoplasmic reticulum chaperone BiP | NP_001156906.1 | 0.68 | 1.31 | 0.611 | 41 | 53 | 5.07 | 268 |
| 29 | HYOU1 | Hypoxia up-regulated protein 1 | NP_067370.3 | 0.29 | 7.26 | 0.019 | 22 | 51 | 5.12 | 78 |
| 30 | PNP | Purine nucleoside phosphorylase | NP_038660.1 | 0.67 | 1.41 | 0.321 | 41 | 31 | 5.78 | 99 |
| 31 | NME1 | Nucleoside diphosphate kinase A | NP_032730.1 | 0.59 | 1.58 | 0.107 | 73 | 33 | 6.84 | 131 |
| 32 | GSTP1 | Glutathione S-transferase P 1 | NP_038569.1 | 0.54 | 1.6 | 0.236 | 62 | 30 | 7.68 | 164 |
| 33 | DDT | D-dopachrome decarboxylase | NP_034157.1 | 0.29 | 1.71 | 0.429 | 85 | 35 | 6.09 | 107 |
| 34 | Tf | Serotransferrin | NP_598738.1 | 0.61 | 1.43 | 0.217 | 33 | 36 | 6.94 | 156 |
| 35 | FGA | Isoform 2 of fibrinogen alpha chain | NP_001104518.1 | 0.62 | 2.95 | 0.272 | 33 | 47 | 7.16 | 122 |
| 36 | ALDH6A1 | Methylmalonate-semialdehyde dehydrogenase (acylating), mitochondrial | NP_598803.1 | 0.61 | 3.79 | 0.217 | 25 | 57 | 8.29 | 67 |
| 37 | ALDH12A1 | Delta-1-pyrroline-5-carboxylate dehydrogenase, mitochondrial | NP_780647.3 | 0.62 | 2.95 | 0.128 | 26 | 47 | 8.45 | 80 |
| CON-SP | CON-TMP | HFD-SP | HFD-TMP | 2WA 1 | |
|---|---|---|---|---|---|
| Serum markers | |||||
| AST (U/L) | 49.55 ± 3.33 | 39.33 ± 1.56 | 53.00 ± 5.29 | 42.77 ± 2.67 | F 2 (p = 0.008) |
| ALT (U/L) | 22.36 ± 1.95 | 17.22 ± 3.48 | 33.82 ± 5.57 | 27.80 ± 6.22 | F (p = 0.018) |
| TG (mg/dL) | 97.01 ± 9.79 | 74.84 ± 8.85 | 78.15 ± 7.15 | 70.38 ± 6.51 | F (p = 0.000) |
| FFA (μmol/L) | 0.64 ± 0.04 | 0.55 ± 0.04 | 0.52 ± 0.04 | 0.52 ± 0.04 | |
| TC (mg/dL) | 120.01 ± 5.40 | 112.91 ± 5.51 | 149.41 ± 8.67 | 153.84 ± 6.58 | |
| HDL-C (mg/dL) | 84.64 ± 4.72 | 87.52 ± 4.56 | 92.47 ± 5.32 | 97.03 ± 5.64 | |
| HTR (%) | 70.40 ± 2.59 | 77.53 ± 1.87 | 62.19 ± 2.17 | 63.34 ± 3.25 | F (p = 0.000) |
| AI | 0.44 ± 0.05 | 0.30 ± 0.03 | 0.63 ± 0.05 | 0.62 ± 0.08 | F (p = 0.000) |
| TNF-α (pg/mL) | 9.84 ± 0.71 | 9.49 ± 0.80 | 10.68 ± 0.91 | 10.86 ± 0.85 | |
| IL-6 (pg/mL) | 18.07 ± 4.67 | 11.84 ± 1.21 | 17.63 ± 2.69 | 16.51 ± 2.54 | |
| Adiponectin (μg/mL) | 3.63 ± 0.18 | 3.55 ± 0.16 | 3.60 ± 0.14 | 3.36 ± 0.18 | |
| Fecal lipid contents | |||||
| TG (mg/g) | 67.94 ± 5.66 | 390.60 ± 15.67 † | 333.32 ± 15.26 † | 416.33 ± 73.37 † | TxF 3 (p = 0.000) |
| FFA (μmol/g) | 61.16 ± 4.63 | 64.83 ± 4.18 | 155.48 ± 7.75 | 172.52 ± 17.41 | F (p = 0.000) |
| Cholesterol (mg/g) | 66.39 ± 3.07 | 48.69 ± 1.54 † | 51.22 ± 2.49 † | 60.23 ± 3.50 | TxF (p = 0.013) |
| Ingredients (g/kg Diet) | CON-SP | CON-TMP | HFD-SP | HFD-TMP |
|---|---|---|---|---|
| Soy protein | 200 | - | 200 | - |
| TMP 1 | - | 200 | - | 200 |
| DL-methionine | 3 | 3 | 3 | 3 |
| Choline bitartrate | 2 | 2 | 2 | 2 |
| Corn starch | 500 | 500 | 340 | 340 |
| Sucrose | 150 | 150 | 150 | 150 |
| Cellulose | 50 | 50 | 50 | 50 |
| Corn oil | 50 | 50 | 30 | 30 |
| Lard | 0 | 0 | 180 | 180 |
| Mineral mixture | 35 | 35 | 35 | 35 |
| Vitamin mixture | 10 | 10 | 10 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ham, J.R.; Choi, R.-Y.; Lee, Y.; Lee, M.-K. Effects of Edible Insect Tenebrio molitor Larva Fermentation Extract as a Substitute Protein on Hepatosteatogenesis and Proteomic Changes in Obese Mice Induced by High-Fat Diet. Int. J. Mol. Sci. 2021, 22, 3615. https://doi.org/10.3390/ijms22073615
Ham JR, Choi R-Y, Lee Y, Lee M-K. Effects of Edible Insect Tenebrio molitor Larva Fermentation Extract as a Substitute Protein on Hepatosteatogenesis and Proteomic Changes in Obese Mice Induced by High-Fat Diet. International Journal of Molecular Sciences. 2021; 22(7):3615. https://doi.org/10.3390/ijms22073615
Chicago/Turabian StyleHam, Ju Ri, Ra-Yeong Choi, Yongjin Lee, and Mi-Kyung Lee. 2021. "Effects of Edible Insect Tenebrio molitor Larva Fermentation Extract as a Substitute Protein on Hepatosteatogenesis and Proteomic Changes in Obese Mice Induced by High-Fat Diet" International Journal of Molecular Sciences 22, no. 7: 3615. https://doi.org/10.3390/ijms22073615
APA StyleHam, J. R., Choi, R. -Y., Lee, Y., & Lee, M. -K. (2021). Effects of Edible Insect Tenebrio molitor Larva Fermentation Extract as a Substitute Protein on Hepatosteatogenesis and Proteomic Changes in Obese Mice Induced by High-Fat Diet. International Journal of Molecular Sciences, 22(7), 3615. https://doi.org/10.3390/ijms22073615