
Overexpression of miR-210-3p Impairs Extravillous Trophoblast Functions Associated with Uterine Spiral Artery Remodeling

 ,
,
Abstract
:1. Introduction:
2. Results
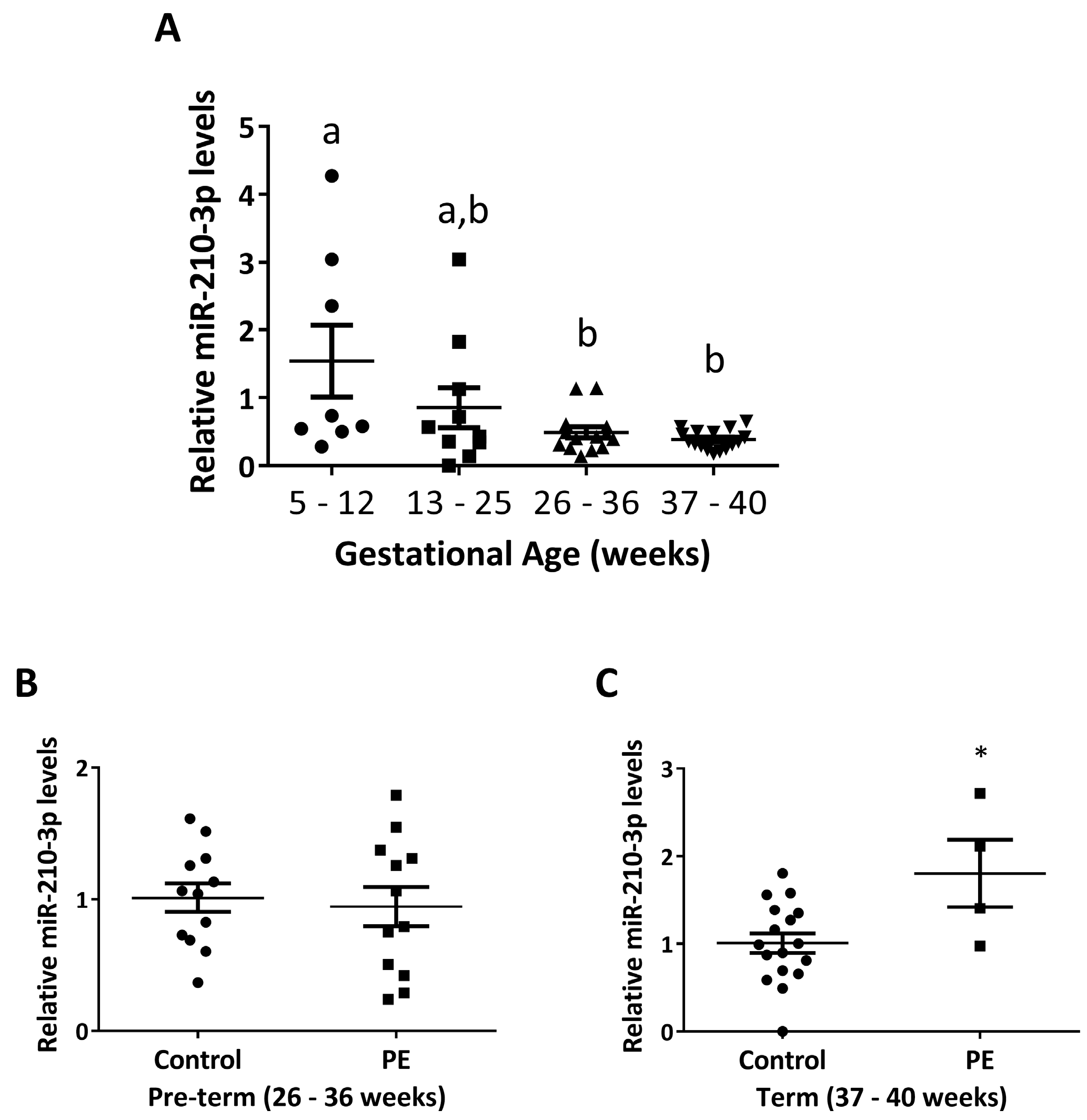
2.1. miR-210-3p Level Decreases with Increasing Gestation in Healthy Pregnancies and Is Upregulated in PE
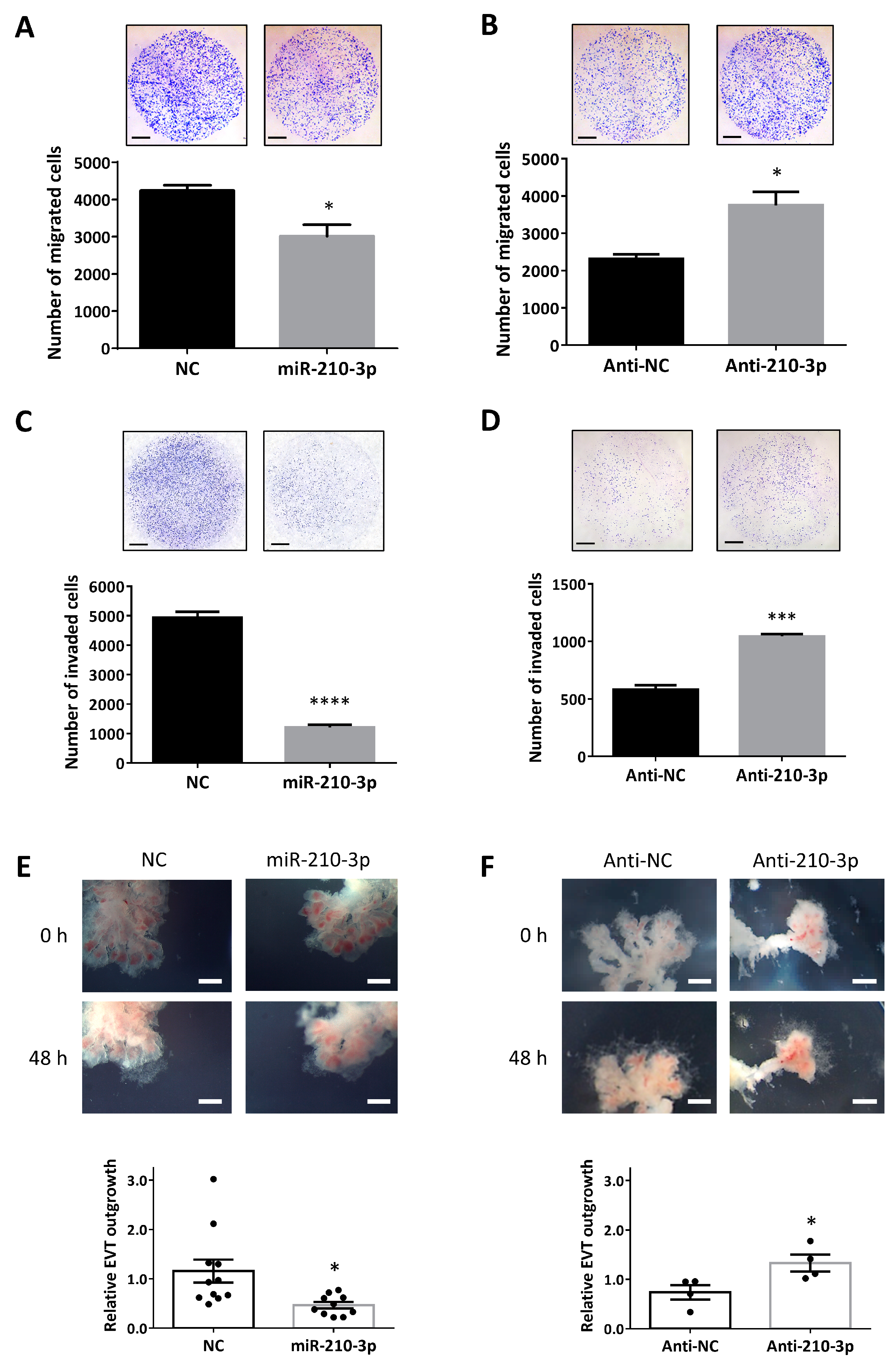
2.2. miR-210-3p Downregulates Migration and Invasion of HTR8/SVneo Cells and EVT Outgrowth in First Trimester Placental Explants
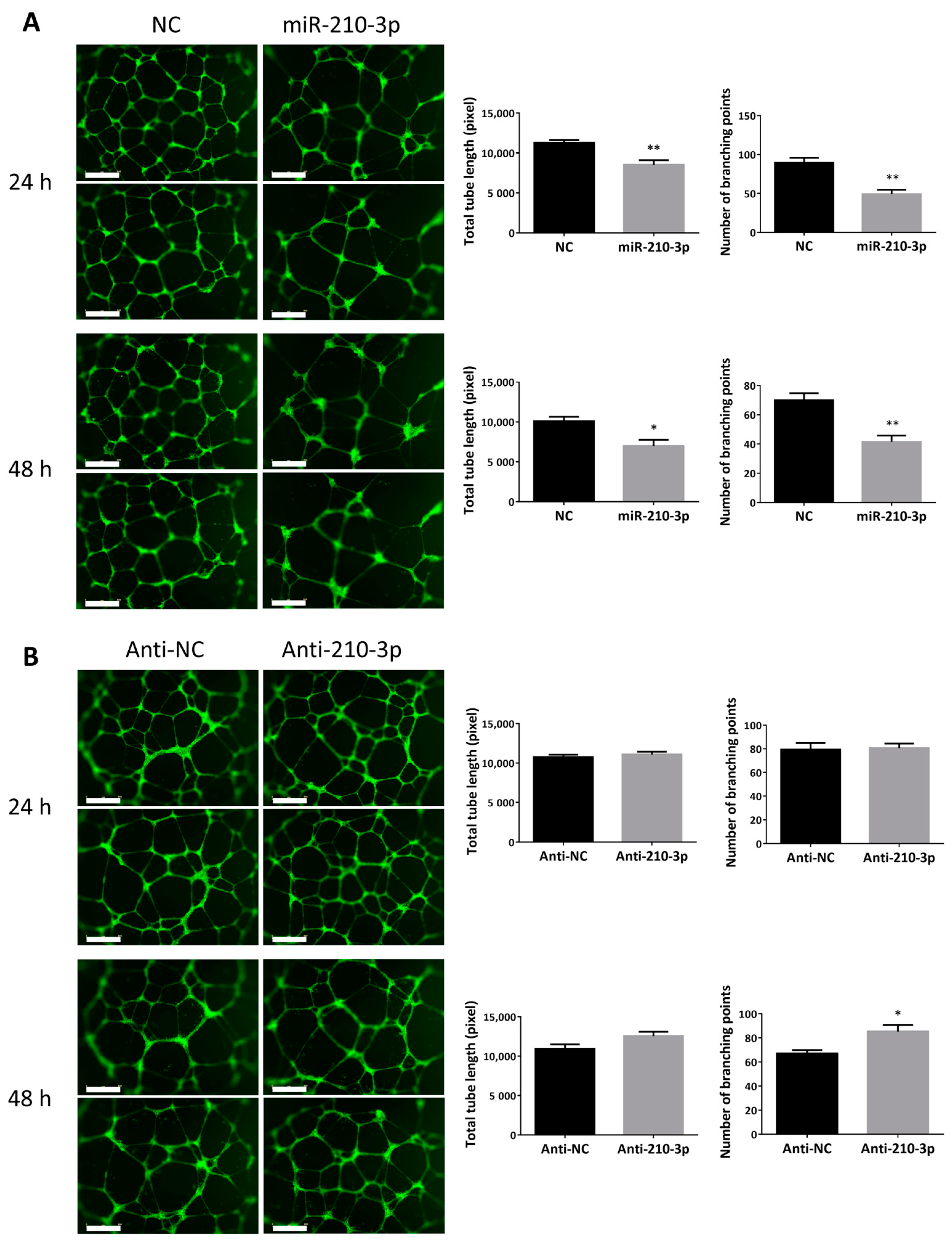
2.3. miR-210-3p Reduced the Ability of HTR8/SVneo to Form Endothelial-Like Networks
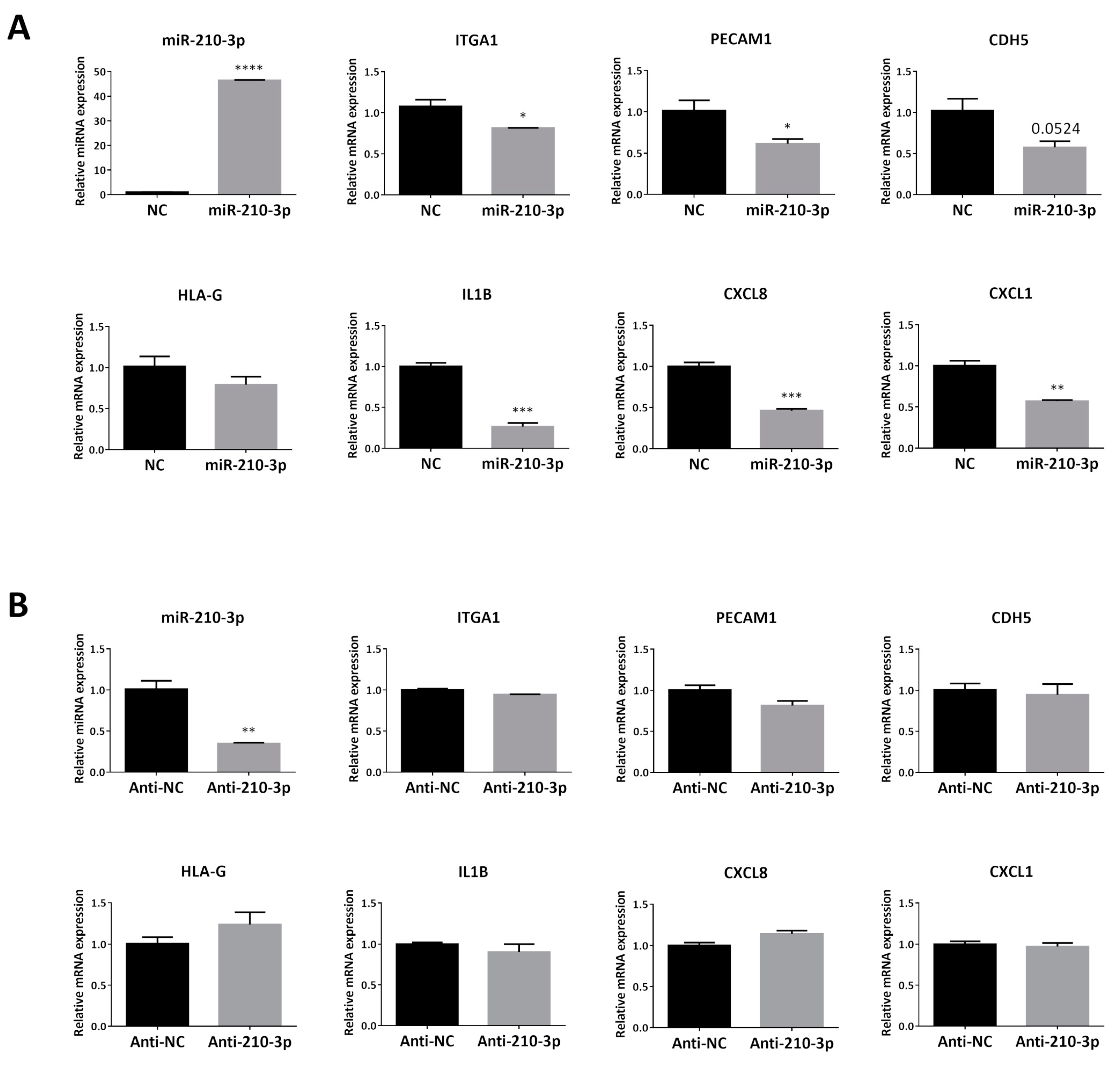
2.4. Overexpressing miR-210-3p in HTR8/SVneo Decreased the mRNA Levels of Invasion and enEVT Differentiation Markers Especially IL1B, CXCL8 and CXCL1
2.5. Upregulation of Endogenous miR-210-3p by Low Oxygen Tension Led to a Decrease in IL1B, CXCL8 and CXCL1 at 1% O2
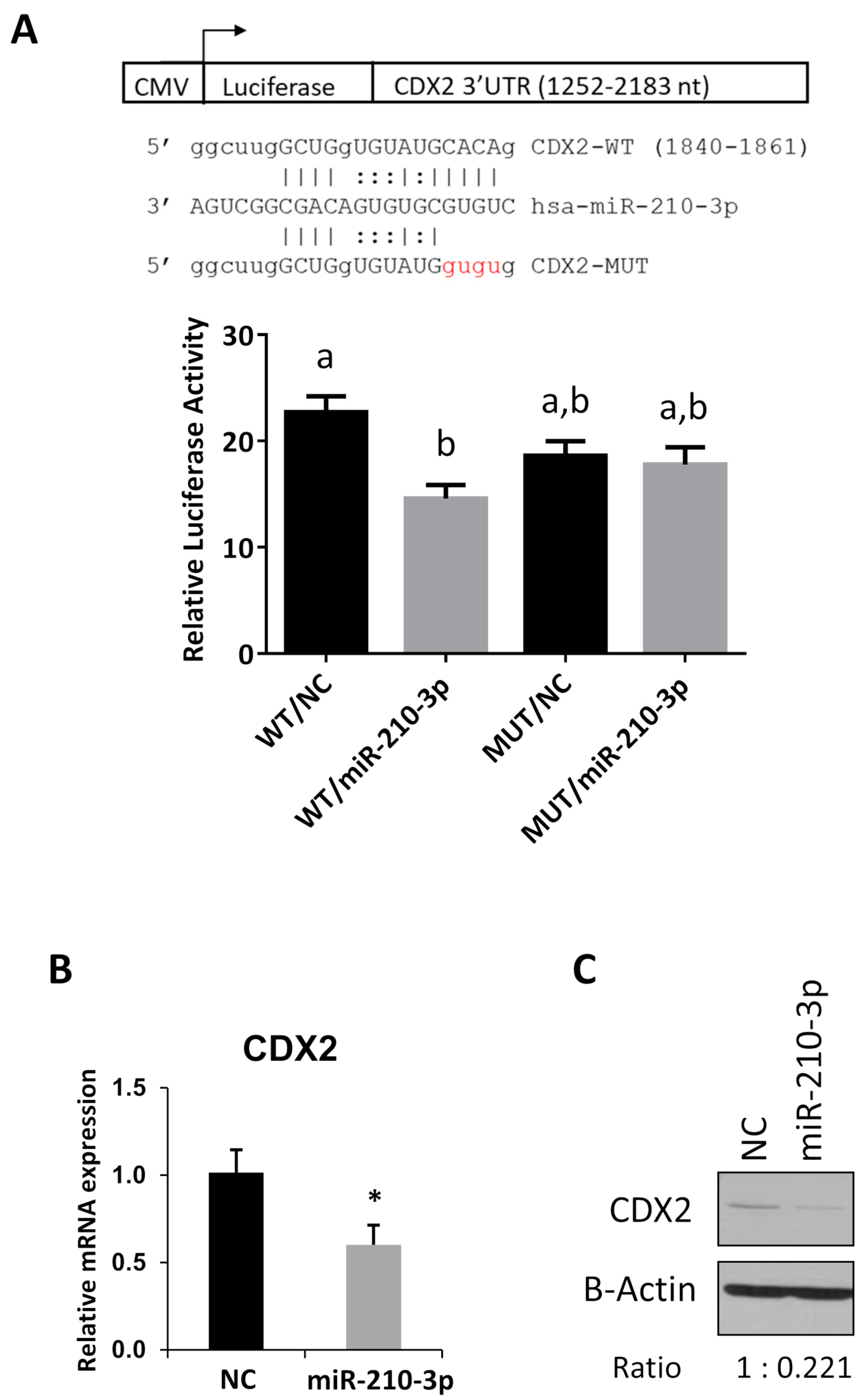
2.6. CDX2 Is a Novel Target of miR-210-3p
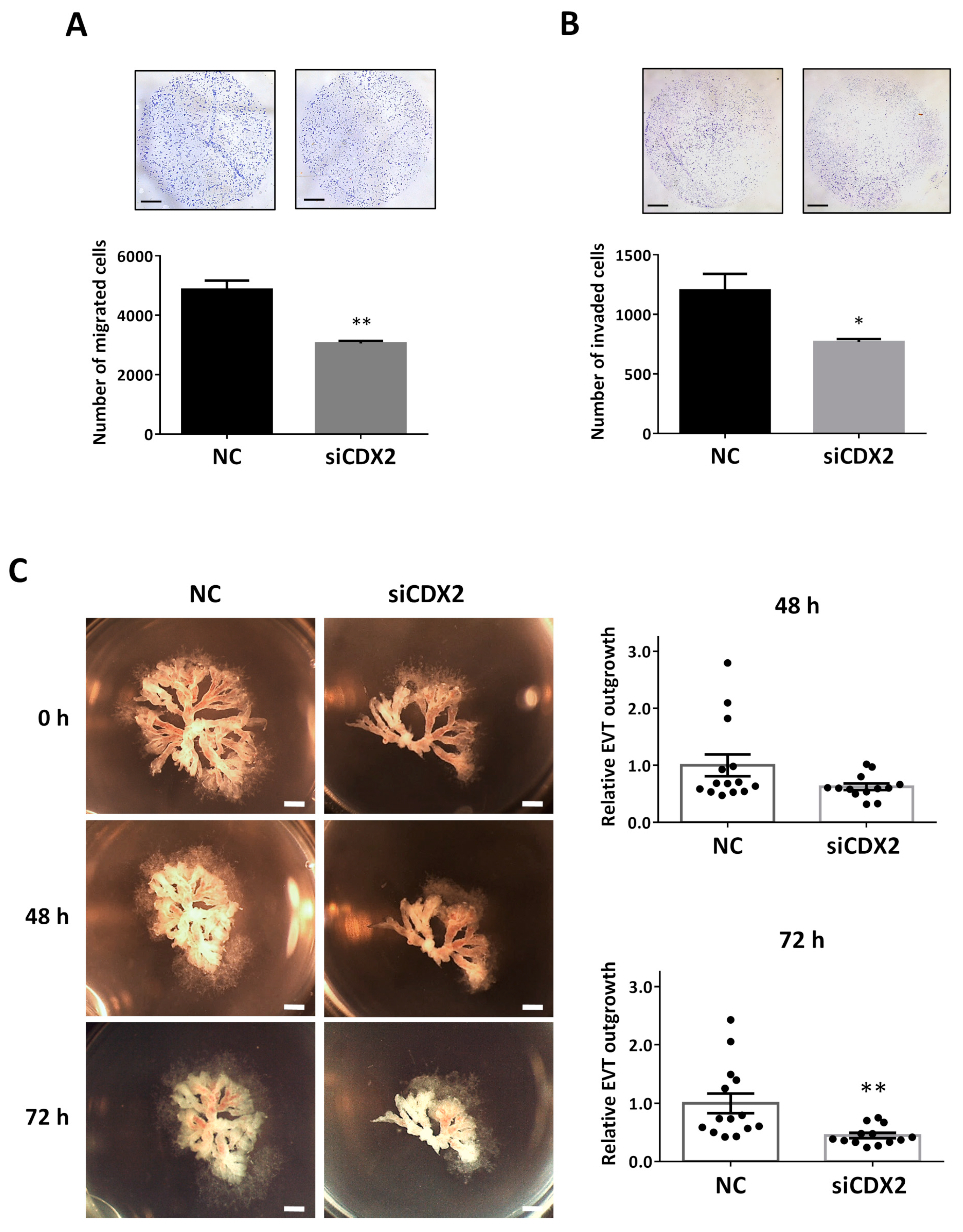
2.7. CDX2 Knockdown Mimics the Effect of miR-210-3p Overexpression and Led to Decreased HTR8/SVneo Migration and Invasion and Decreased EVT Outgrowth in First Trimester Placental Explants
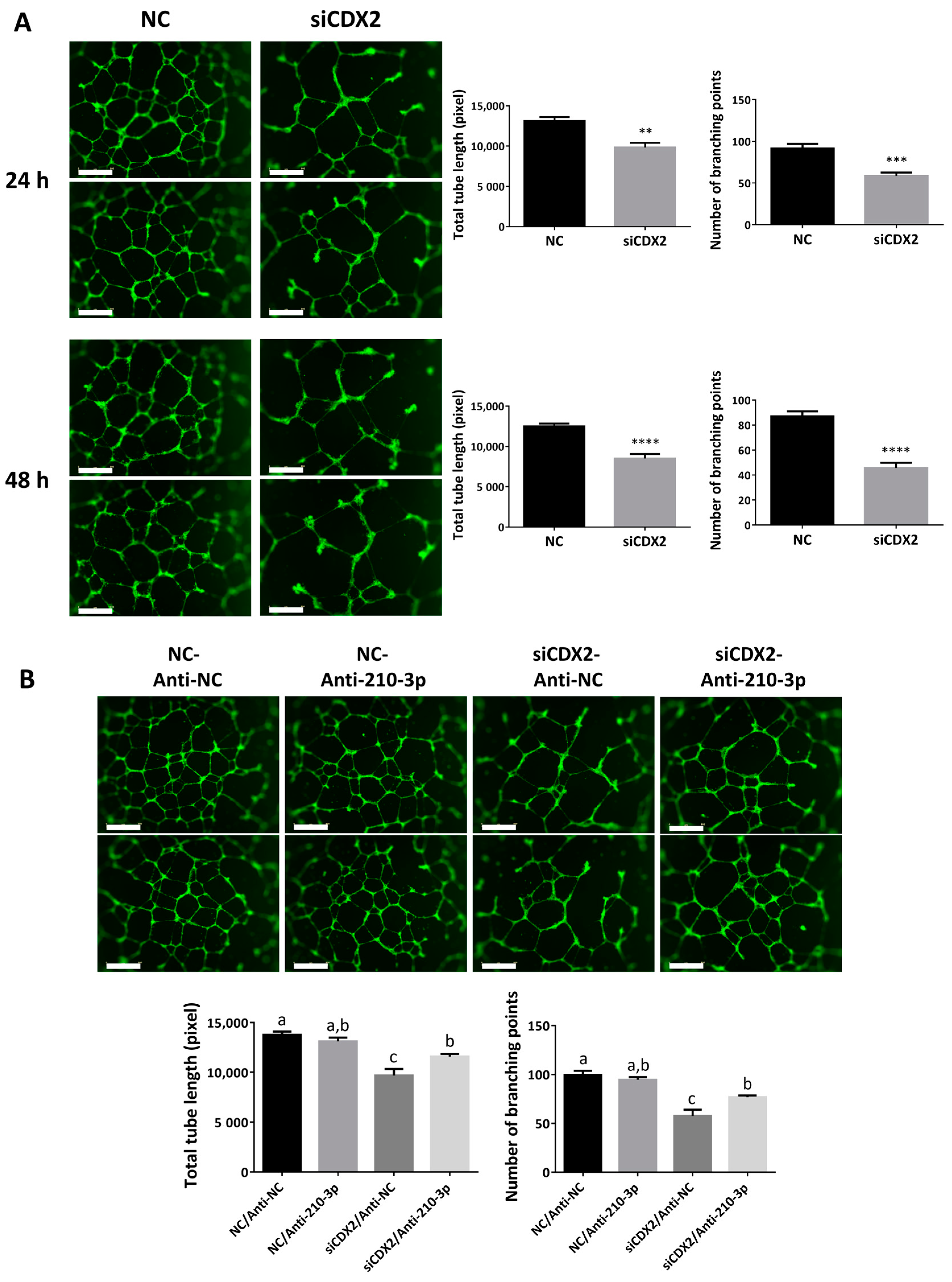
2.8. CDX2 Downregulation Reduced the Ability of HTR8/SVneo to Form Endothelial-Like Networks
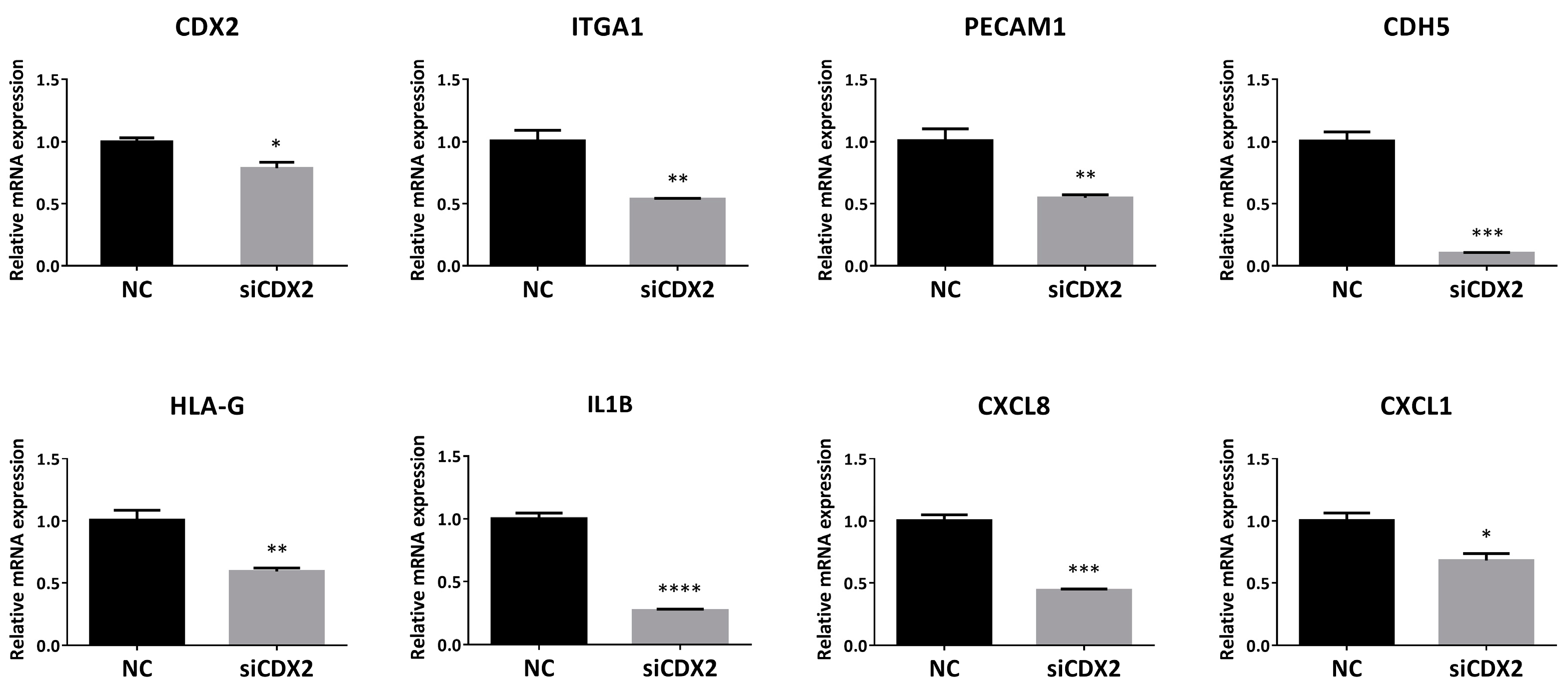
2.9. CDX2 Knockdown in HTR8/SVneo Decreased the Levels of EVT and enEVT Differentiation Markers, As Well As IL1B, CXCL8 and CXCL1
3. Discussion
4. Material and Methods
4.1. Clinical Samples and Tissue Collection
4.2. Cell Lines and Cell Culture
4.3. Transient Transfections and Treatments
4.4. RNA Extraction, Reverse Transcription and Quantitative Real Time PCR (qRT-PCR)
4.5. Transwell Migration and Invasion Assay
4.6. First-Trimester Human Placenta Explant Culture
4.7. Tube Formation Assay
4.8. Protein Extraction and Immunoblot Analysis
4.9. Luciferase Reporter Assay
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PE | Preeclampsia |
| EVT | Extravillous trophoblasts |
| enEVT | Endovascular trophoblasts |
| U6 | Small nuclear RNA U6 |
| U48 | Small nucleolar RNA U48 |
| ITGA1 | Integrin subunit alpha 1 |
| PECAM1 | Platelet and endothelial cell adhesion molecule 1/CD31 |
| CDH5 | Cadherin-5/Vascular endothelial cadherin/CD144 |
| HLA-G | Major histocompatibility complex, Class I, G |
| IL1B | Interleukin 1 beta |
| CXCL8 | C-X-C motif chemokine ligand 8/Interleukin 8 |
| CXCL1 | C-X-C motif chemokine ligand 1 |
| CDX2 | Caudal type homeobox 2 |
| CYC1 | Cytochrome C1 |
References
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.; Brkić, J.; Liu, M.; Fu, G.; Peng, C.; Wang, Y.-L. Placental trophoblast cell differentiation: Physiological regulation and pathological relevance to preeclampsia. Mol. Asp. Med. 2013, 34, 981–1023. [Google Scholar] [CrossRef]
- Burton, G.J.; Cindrova-Davies, T.; Yung, H.W.; Jauniaux, E. Hypoxia and Reproductive Health: Oxygen and development of the human placenta. Reproduction 2021, 161, F53–F65. [Google Scholar] [CrossRef]
- Anton, L.; Olarerin-George, A.O.; Schwartz, N.; Srinivas, S.; Bastek, J.; Hogenesch, J.B.; Elovitz, M.A. miR-210 inhibits trophoblast invasion and is a serum biomarker for preeclampsia. Am. J. Pathol. 2013, 183, 1437–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, G.; Brkic, J.; Hayder, H.; Peng, C. MicroRNAs in Human Placental Development and Pregnancy Complications. Int. J. Mol. Sci. 2013, 14, 5519–5544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.; Yang, Q.; Dai, Y.; Guo, W.; Du, H.; Song, L.; Peng, X. Oncogenic miR-210-3p promotes prostate cancer cell EMT and bone metastasis via NF-κB signaling pathway. Mol. Cancer 2017, 16, 117. [Google Scholar] [CrossRef] [Green Version]
- Dang, K.; Myers, K.A. The role of hypoxia-induced miR-210 in cancer progression. Int. J. Mol. Sci. 2015, 16, 6353–6372. [Google Scholar] [CrossRef]
- Devlin, C.; Greco, S.; Martelli, F.; Ivan, M. miR-210: More than a silent player in hypoxia. IUBMB Life 2011, 63, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Dasgupta, C.; Li, Y.; Huang, L.; Zhang, L. MicroRNA-210 Downregulates ISCU and Induces Mitochondrial Dysfunction and Neuronal Death in Neonatal Hypoxic-Ischemic Brain Injury. Mol. Neurobiol. 2019, 56, 5608–5625. [Google Scholar] [CrossRef]
- Chan, Y.C.; Banerjee, J.; Choi, S.Y.; Sen, C.K. miR-210: The master hypoxamir. Microcirculation 2012, 19, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhou, Y.; Zhang, Z. MiR-210: An important player in the pathogenesis of preeclampsia? J. Cell. Mol. Med. 2012, 16, 943–944. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fei, M.; Xue, G.; Zhou, Q.; Jia, Y.; Li, L.; Xin, H.; Sun, S. Elevated levels of hypoxia-inducible microRNA-210 in pre-eclampsia: New insights into molecular mechanisms for the disease. J. Cell. Mol. Med. 2012, 16, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Long, A.; Jiang, L.; Cai, L.; Xie, L.I.; Gu, J.; Chen, X.; Tan, L. Quantification of preeclampsia-related microRNAs in maternal serum. Biomed. Rep. 2015, 3, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Muralimanoharan, S.; Maloyan, A.; Mele, J.; Guo, C.; Myatt, L.G.; Myatt, L. MIR-210 modulates mitochondrial respiration in placenta with preeclampsia. Placenta 2012, 33, 816–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Q.; Furong, W.; Baosheng, L. Multiple functions of hypoxia-regulated miR-210 in cancer. J. Exp. Clin. Cancer Res. 2014, 33, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Le, Q.T.; Giaccia, A.J. MiR-210--micromanager of the hypoxia pathway. Trends Mol. Med. 2010, 16, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.C.; Romero, R.; Kim, J.S.; Tarca, A.L.; Montenegro, D.; Pineles, B.L.; Kim, E.; Lee, J.; Kim, S.Y.; Draghici, S.; et al. miR-210 targets iron-sulfur cluster scaffold homologue in human trophoblast cell lines: Siderosis of interstitial trophoblasts as a novel pathology of preterm preeclampsia and small-for-gestational-age pregnancies. Am. J. Pathol. 2011, 179, 590–602. [Google Scholar] [CrossRef]
- Luo, R.; Wang, Y.; Xu, P.; Cao, G.; Zhao, Y.; Shao, X.; Li, Y.-X.; Chang, C.; Peng, C.; Wang, Y.-L. Hypoxia-inducible miR-210 contributes to preeclampsia via targeting thrombospondin type I domain containing 7A. Sci. Rep. 2016, 6, 19588. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Shao, X.; Xu, P.; Liu, Y.; Wang, Y.; Zhao, Y.; Liu, M.; Ji, L.; Li, Y.-X.; Chang, C.; et al. MicroRNA-210 Contributes to Preeclampsia by Downregulating Potassium Channel Modulatory Factor 1. Hypertension 2014, 64, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Huang, X.; He, Z.; Xiong, Y.; Fang, Q. miRNA-210-3p regulates trophoblast proliferation and invasiveness through fibroblast growth factor 1 in selective intrauterine growth restriction. J. Cell. Mol. Med. 2019, 23, 4422–4433. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, Y.; Luo, R.; Bian, X.; Wang, Y.; Shao, X.; Li, Y.X.; Liu, M.; Wang, Y.L. A positive feedback self-regulatory loop between miR-210 and HIF-1α mediated by CPEB2 is involved in trophoblast syncytialization: Implication of trophoblast malfunction in preeclampsia. Biol. Reprod. 2020, 102, 560–570. [Google Scholar]
- Kunath, T.; Strumpf, D.; Rossant, J. Early Trophoblast Determination and Stem Cell Maintenance in the Mouse—A Review. Placenta 2004, 25, S32–S38. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Bai, H.; Konno, T.; Ideta, A.; Aoyagi, Y.; Godkin, J.D.; Imakawa, K. Function of a Transcription Factor CDX2 Beyond Its Trophectoderm Lineage Specification. Endocrinology 2010, 151, 5873–5881. [Google Scholar] [CrossRef] [PubMed]
- Strumpf, D.; Mao, C.-A.; Yamanaka, Y.; Ralston, A.; Chawengsaksophak, K.; Beck, F.; Rossant, J. Cdx2 is required for correct cell fate specification and differentiation of trophectoderm in the mouse blastocyst. Development 2005, 132, 2093–2102. [Google Scholar] [CrossRef] [Green Version]
- Jia, R.Z.; Ding, G.C.; Gu, C.M.; Huang, T.; Rui, C.; Wang, Y.X.; Lu, Q. CDX2 Enhances HTR-8/SVneo Trophoblast Cell Invasion by Altering the Expression of Matrix Metalloproteinases. Cell. Physiol. Biochem. 2014, 34, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Na, Q.; Huang, L.; Song, G.; Jin, F.; Li, Y.; Hou, Y.; Kang, D.; Qiao, C. YAP Is Decreased in Preeclampsia and Regulates Invasion and Apoptosis of HTR-8/SVneo. Reprod. Sci. 2018, 25, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, R.H.; Dunk, C.E.; Lye, S.J.; Aplin, J.D.; Harris, L.K.; Jones, R.L. Extravillous Trophoblast and Endothelial Cell Crosstalk Mediates Leukocyte Infiltration to the Early Remodeling Decidual Spiral Arteriole Wall. J. Immunol. 2017, 198, 4115–4128. [Google Scholar] [CrossRef]
- Ramhorst, R.; Grasso, E.; Paparini, D.; Hauk, V.; Gallino, L.; Calo, G.; Vota, D.; Pérez Leirós, C. Decoding the chemokine network that links leukocytes with decidual cells and the trophoblast during early implantation. Cell Adhes. Migr. 2016, 10, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Tessier, D.R.; Yockell-Lelièvre, J.; Gruslin, A. Uterine Spiral Artery Remodeling: The Role of Uterine Natural Killer Cells and Extravillous Trophoblasts in Normal and High-Risk Human Pregnancies. Am. J. Reprod. Immunol. 2015, 74, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hauguel-de Mouzon, S.; Guerre-Millo, M. The Placenta Cytokine Network and Inflammatory Signals. Placenta 2006, 27, 794–798. [Google Scholar] [CrossRef]
- Bowen, J.M.; Chamley, L.; Keelan, J.A.; Mitchell, M.D. Cytokines of the Placenta and Extra-placental Membranes: Roles and Regulation During Human Pregnancy and Parturition. Placenta 2002, 23, 257–273. [Google Scholar] [CrossRef]
- Bowen, J.M.; Chamley, L.; Mitchell, M.D.; Keelan, J.A. Cytokines of the Placenta and Extra-placental Membranes: Biosynthesis, Secretion and Roles in Establishment of Pregnancy in Women. Placenta 2002, 23, 239–256. [Google Scholar] [CrossRef]
- Prutsch, N.; Fock, V.; Haslinger, P.; Haider, S.; Fiala, C.; Pollheimer, J.; Knöfler, M. The role of interleukin-1β in human trophoblast motility. Placenta 2012, 33, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Librach, C.L.; Feigenbaum, S.L.; Bass, K.E.; Cui, T.Y.; Verastas, N.; Sadovsky, Y.; Quigley, J.P.; French, D.L.; Fisher, S.J. Interleukin-1 beta regulates human cytotrophoblast metalloproteinase activity and invasion in vitro. J. Biol. Chem. 1994, 269, 17125–17131. [Google Scholar] [CrossRef]
- Schraufstatter, I.U.; Chung, J.; Burger, M. IL-8 activates endothelial cell CXCR1 and CXCR2 through Rho and Rac signaling pathways. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001, 280, L1094–L1103. [Google Scholar] [CrossRef] [Green Version]
- Ramhorst, R.; Fraccaroli, L.; Aldo, P.; Alvero, A.B.; Cardenas, I.; Leirós, C.P.; Mor, G. Modulation and Recruitment of Inducible Regulatory T Cells by First Trimester Trophoblast Cells. Am. J. Reprod. Immunol. 2012, 67, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Baston-Buest, D.M.; Altergot-Ahmad, O.; Pour, S.J.; Krüssel, J.-S.; Markert, U.R.; Fehm, T.N.; Bielfeld, A.P. Syndecan-1 Acts as an Important Regulator of CXCL1 Expression and Cellular Interaction of Human Endometrial Stromal and Trophoblast Cells. Mediat. Inflamm. 2017, 2017, 8379256. [Google Scholar] [CrossRef]
- Jovanović, M.; Stefanoska, I.; Radojcić, L.; Vićovac, L. Interleukin-8 (CXCL8) stimulates trophoblast cell migration and invasion by increasing levels of matrix metalloproteinase (MMP)2 and MMP9 and integrins alpha5 and beta1. Reproduction 2010, 139, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Ain, R. MicroRNA regulation of murine trophoblast stem cell self-renewal and differentiation. Life Sci. Alliance 2020, 3, e202000674. [Google Scholar] [CrossRef] [PubMed]
- Brkić, J.; Dunk, C.; O’Brien, J.; Fu, G.; Nadeem, L.; Wang, Y.-L.; Rosman, D.; Salem, M.; Shynlova, O.; Yougbaré, I.; et al. MicroRNA-218-5p Promotes Endovascular Trophoblast Differentiation and Spiral Artery Remodeling. Mol. Ther. 2018, 26, 2189–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, U.; Ye, G.; Salem, M.; Peng, C. MicroRNA-378a-5p Targets Cyclin G2 to Inhibit Fusion and Differentiation in BeWo Cells. Biol. Reprod. 2014, 91, 76. [Google Scholar] [CrossRef] [Green Version]
- Bulla, R.; Villa, A.; Bossi, F.; Cassetti, A.; Radillo, O.; Spessotto, P.; De Seta, F.; Guaschino, S.; Tedesco, F. VE-cadherin is a critical molecule for trophoblast-endothelial cell interaction in decidual spiral arteries. Exp. Cell Res. 2005, 303, 101–113. [Google Scholar] [CrossRef]
- Matsumura, T.; Wolff, K.; Petzelbauer, P. Endothelial Cell Tube Formation Depends on Cadherin 5 and CD31 Interactions with Filamentous Actin. J. Immunol. 1997, 158, 3408–3416. [Google Scholar] [PubMed]
- Sato, Y. Endovascular trophoblast and spiral artery remodeling. Mol. Cell. Endocrinol. 2020, 503, 110699. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, E.D.; Pepe, G.J. Regulation of Uterine Spiral Artery Remodeling: A Review. Reprod. Sci. 2020, 27, 1932–1942. [Google Scholar] [CrossRef] [PubMed]
- Hess, A.P.; Hamilton, A.E.; Talbi, S.; Dosiou, C.; Nyegaard, M.; Nayak, N.; Genbecev-Krtolica, O.; Mavrogianis, P.; Ferrer, K.; Kruessel, J.; et al. Decidual stromal cell response to paracrine signals from the trophoblast: Amplification of immune and angiogenic modulators. Biol. Reprod. 2007, 76, 102–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naruse, K.; Innes, B.A.; Bulmer, J.N.; Robson, S.C.; Searle, R.F.; Lash, G.E. Secretion of cytokines by villous cytotrophoblast and extravillous trophoblast in the first trimester of human pregnancy. J. Reprod. Immunol. 2010, 86, 148–150. [Google Scholar] [CrossRef]
- Red-Horse, K.; Drake, P.M.; Gunn, M.D.; Fisher, S.J. Chemokine Ligand and Receptor Expression in the Pregnant Uterus: Reciprocal Patterns in Complementary Cell Subsets Suggest Functional Roles. Am. J. Pathol. 2001, 159, 2199–2213. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, K.; Gong, Y.G.; Khoo, S.K.; Leach, R. Roles of CDX2 and EOMES in human induced trophoblast progenitor cells. Biochem. Biophys. Res. Commun. 2013, 431, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Malysheva, S.V.; Wunderlich, S.; Haase, A.; Göhring, G.; Martin, U.; Merkert, S. Generation of a human CDX2 knock-in reporter iPSC line (MHHi007-A-1) to model human trophoblast differentiation. Stem Cell Res. 2018, 30, 117–121. [Google Scholar] [CrossRef]
- Tolkunova, E.; Cavaleri, F.; Eckardt, S.; Reinbold, R.; Christenson, L.K.; Schöler, H.R.; Tomilin, A. The Caudal-Related Protein Cdx2 Promotes Trophoblast Differentiation of Mouse Embryonic Stem Cells. Stem Cells 2006, 24, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Ma, L.; Wang, Z.; Yu, Y.; Lye, S.J.; Shan, Y.; Wei, Y. Deciphering a distinct regulatory network of TEAD4, CDX2 and GATA3 in humans for trophoblast transition from embryonic stem cells. Biochim. Biophys. Acta (Bba) Mol. Cell Res. 2020, 1867, 118736. [Google Scholar] [CrossRef] [PubMed]
- Hemberger, M.; Udayashankar, R.; Tesar, P.; Moore, H.; Burton, G.J. ELF5-enforced transcriptional networks define an epigenetically regulated trophoblast stem cell compartment in the human placenta. Hum. Mol. Genet. 2010, 19, 2456–2467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.J.; Cindrova-Davies, T.; Turco, M.Y. Review: Histotrophic nutrition and the placental-endometrial dialogue during human early pregnancy. Placenta 2020, 102, 21–26. [Google Scholar] [CrossRef]
- Schiffmacher, A.T.; Keefer, C.L. CDX2 regulates multiple trophoblast genes in bovine trophectoderm CT-1 cells. Mol. Reprod. Dev. 2013, 80, 826–839. [Google Scholar] [CrossRef]
- Scott, I.C.; Anson-Cartwright, L.; Riley, P.; Reda, D.; Cross, J.C. The HAND1 Basic Helix-Loop-Helix Transcription Factor Regulates Trophoblast Differentiation via Multiple Mechanisms. Mol. Cell. Biol. 2000, 20, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Kanda, H.; Tanaka, S.; Takamatsu, N.; Shiba, T.; Ito, M. Sox15 enhances trophoblast giant cell differentiation induced by Hand1 in mouse placenta. Differentiation 2006, 74, 212–221. [Google Scholar] [CrossRef]
- Kolahi, K.S.; Valent, A.M.; Thornburg, K.L. Cytotrophoblast, Not Syncytiotrophoblast, Dominates Glycolysis and Oxidative Phosphorylation in Human Term Placenta. Sci. Rep. 2017, 7, 42941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Li, Y.; Zhang, H.; Huang, P.; Luthra, R. Hypoxia-regulated microRNA-210 modulates mitochondrial function and decreases ISCU and COX10 expression. Oncogene 2010, 29, 4362–4368. [Google Scholar] [CrossRef] [Green Version]
- Huppertz, B. The Critical Role of Abnormal Trophoblast Development in the Etiology of Preeclampsia. Curr. Pharm. Biotechnol. 2018, 19, 771–780. [Google Scholar] [CrossRef]
- Enquobahrie, D.A.; Hensley, M.; Qiu, C.; Abetew, D.F.; Hevner, K.; Tadesse, M.G.; Williams, M.A. Candidate Gene and MicroRNA Expression in Fetal Membranes and Preterm Delivery Risk. Reprod. Sci. 2016, 23, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Allport, V.; Sykes, A.; Lindstrom, T.; Slater, D.; Bennett, P. The effects of labour and of interleukin 1 beta upon the expression of nuclear factor kappa B related proteins in human amnion. Mol. Hum. Reprod. 2003, 9, 213–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allport, V.C.; Pieber, D.; Slater, D.M.; Newton, R.; White, J.O.; Bennett, P.R. Human labour is associated with nuclear factor-kappaB activity which mediates cyclo-oxygenase-2 expression and is involved with the ‘functional progesterone withdrawal’. Mol. Hum. Reprod. 2001, 7, 581–586. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; MacIntyre, D.A.; Lee, Y.S.; Khanjani, S.; Terzidou, V.; Teoh, T.G.; Bennett, P.R. Nuclear Factor Kappa B Activation Occurs in the Amnion Prior to Labour Onset and Modulates the Expression of Numerous Labour Associated Genes. PLoS ONE 2012, 7, e34707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappas, M.; Rice, G.E. The Role and Regulation of the Nuclear Factor Kappa B Signalling Pathway in Human Labour. Placenta 2007, 28, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Lindström, T.M.; Bennett, P.R. The role of nuclear factor kappa B in human labour. Reproduction 2005, 130, 569–581. [Google Scholar] [CrossRef]
- Muralimanoharan, S.; Guo, C.; Myatt, L.; Maloyan, A. Sexual dimorphism in miR-210 expression and mitochondrial dysfunction in the placenta with maternal obesity. Int. J. Obes. 2015, 39, 1274–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejad, R.M.A.; Saeidi, K.; Gharbi, S.; Salari, Z.; Saleh-Gohari, N. Quantification of circulating miR-517c-3p and miR-210-3p levels in preeclampsia. Pregnancy Hypertens. 2019, 16, 75–78. [Google Scholar] [CrossRef]
- Xu, P.; Zhao, Y.; Liu, M.; Wang, Y.; Wang, H.; Li, Y.-X.; Zhu, X.; Yao, Y.; Wang, H.; Qiao, J.; et al. Variations of MicroRNAs in Human Placentas and Plasma From Preeclamptic Pregnancy. Hypertension 2014, 63, 1276–1284. [Google Scholar] [CrossRef]
- Munaut, C.; Tebache, L.; Blacher, S.; Noël, A.; Nisolle, M.; Chantraine, F. Dysregulated circulating miRNAs in preeclampsia. Biomed. Rep. 2016, 5, 686–692. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.-S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human Villous Trophoblasts Express and Secrete Placenta-Specific MicroRNAs into Maternal Circulation via Exosomes1. Biol. Reprod. 2009, 81, 717–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donker, R.B.; Mouillet, J.F.; Chu, T.; Hubel, C.A.; Stolz, D.B.; Morelli, A.E.; Sadovsky, Y. The expression profile of C19MC microRNAs in primary human trophoblast cells and exosomes. Mol. Hum. Reprod. 2012, 18, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brkić, J.; Dunk, C.; Shan, Y.; O’Brien, J.A.; Lye, P.; Qayyum, S.; Yang, P.; Matthews, S.G.; Lye, S.J.; Peng, C. Differential Role of Smad2 and Smad3 in the Acquisition of an Endovascular Trophoblast-Like Phenotype and Preeclampsia. Front. Endocrinol. 2020, 11, 436. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Peng, C. Automated Quantification and Analysis of Cell Counting Procedures Using ImageJ Plugins. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, L.; Munir, S.; Fu, G.; Dunk, C.; Baczyk, D.; Caniggia, I.; Lye, S.; Peng, C. Nodal signals through activin receptor-like kinase 7 to inhibit trophoblast migration and invasion: Implication in the pathogenesis of preeclampsia. Am. J. Pathol. 2011, 178, 1177–1189. [Google Scholar] [CrossRef]
- Caniggia, I.; Taylor, C.V.; Ritchie, J.W.K.; Lye, S.J.; Letarte, M. Endoglin regulates trophoblast differentiation along the invasive Pathway in human placental villous explants. Endocrinology 1997, 138, 4977–4988. [Google Scholar] [CrossRef]
- Fu, G.; Ye, G.; Nadeem, L.; Ji, L.; Manchanda, T.; Wang, Y.; Zhao, Y.; Qiao, J.; Wang, Y.-L.; Lye, S.; et al. MicroRNA-376c Impairs Transforming Growth Factor-β and Nodal Signaling to Promote Trophoblast Cell Proliferation and Invasion. Hypertension 2013, 61, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Meijering, E.; Jacob, M.; Sarria, J.C.; Steiner, P.; Hirling, H.; Unser, M. Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytom. A 2004, 58, 167–176. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Term Control | Pre-Term PE | p Value | Term Control | Term PE | p Value | |
|---|---|---|---|---|---|---|
| Gestational age (weeks) | 29.83 ± 0.51 | 30.27 ± 0.36 | 0.4824 | 38.32 ± 0.14 | 37.25 ± 0.25 | 0.0030 |
| Maternal age (years) | 31.95 ± 1.15 | 30.8 ± 1.85 | 0.6087 | 33.36 ± 0.73 | 34.8 ± 2.21 | 0.4383 |
| Systolic blood pressure (mmHg) | 120.56 ± 2.9 | 165.13 ± 2.93 | <0.0001 | 118.88 ± 2.11 | 166.8 ± 6.05 | <0.0001 |
| Diastolic blood pressure (mmHg) | 73.83 ± 2.27 | 106.67 ± 1.9 | <0.0001 | 77.04 ± 1.79 | 101.2 ± 1.46 | <0.0001 |
| Proteinuria (g/24 h) | None | 3.46 ± 0.13 | - | None | 2.41 ± 0.25 | - |
| % Laboring | 100% | 35.7% | 0.0004 | 11.8% | 75% | 0.0276 |
| Oligomer Name | Sequence: 5′ → 3′ |
|---|---|
| NC (non-targeting control) GenePharma (Shanghai, China) | Sense: UUCUCCGAACGUGUCACGUTT Anti-sense: ACGUGACACGUUCGGAGAATT |
| hsa-miR-210-3p GenePharma (Shanghai, China) | Sense: CUGUGCGUGUGACAGCGGCUGATT Anti-sense: UCAGCCGCUGUCACACGCACAGTT |
| siCDX2 GenePharma (Shanghai, China) | Sense: CCAGGACGAAAGACAAAUATT Anti-sense: UAUUUGUCUUUCGUCCUGGTT |
| Anti-NC RiboBio (Guangzhou, China) | RiboBio™ miRNA Inhibitor, Negative Control |
| Anti-hsa-miR-210-3p RiboBio (Guangzhou, China) | RiboBio™ anti- hsa-miR-210-3p Inhibitor |
| Primer Name | Sequence: 5′ → 3′ |
|---|---|
| miR-210-3p | FP: GTGACAGCGGCTGAA RP: NCode Universal primer |
| U48 | FP: CCCAGGTAACTCTGAGTGTGTC RP: NCode Universal primer |
| U6 | FP: CGCAAGGATGACACGCAATT RP: NCode Universal primer |
| ITGA1 | FP: GCTGGCTCCTCACTGTTGTT RP: CACCTCTCCCAACTGGACAC |
| PECAM1 | FP: ATTGCAGTGGTTATCATCGGAGTG RP: CTCGTTGTTGGAGTTCAGAAGTGG |
| CDH5 | FP: GCCAGTTCTTCCGAGTCACA RP: TTTCCTGTGGGGGTTCCAGT |
| HLA-G | FP: CTGACCCTGACCGAGACCTGG RP: GTCGCAGCCAATCATCCACTGGA |
| IL1B | FP: AATCTGTACCTGTCCTGCGTGTT RP: TGGGTAATTTTTGGGATCTACACTCT |
| CXCL8 | FP: CAGAGACAGCAGAGCACACA RP: GGCAAAACTGCACCTTCACA |
| CXCL1 | FP: CAGGGAATTCACCCCAAGAACA RP: GGATGCAGGATTGAGGCAAGC |
| CDX2 | FP: CGCTTCTGGGCTGCTGCAAAC RP: CGACTGTAGTGAAACTCCTTCTCC |
| CYC1 | FP: CAGATAGCCAAGGATGTGTG RP: CATCATCAACATCTTGAGCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayder, H.; Fu, G.; Nadeem, L.; O’Brien, J.A.; Lye, S.J.; Peng, C. Overexpression of miR-210-3p Impairs Extravillous Trophoblast Functions Associated with Uterine Spiral Artery Remodeling. Int. J. Mol. Sci. 2021, 22, 3961. https://doi.org/10.3390/ijms22083961
Hayder H, Fu G, Nadeem L, O’Brien JA, Lye SJ, Peng C. Overexpression of miR-210-3p Impairs Extravillous Trophoblast Functions Associated with Uterine Spiral Artery Remodeling. International Journal of Molecular Sciences. 2021; 22(8):3961. https://doi.org/10.3390/ijms22083961
Chicago/Turabian StyleHayder, Heyam, Guodong Fu, Lubna Nadeem, Jacob A. O’Brien, Stephen J. Lye, and Chun Peng. 2021. "Overexpression of miR-210-3p Impairs Extravillous Trophoblast Functions Associated with Uterine Spiral Artery Remodeling" International Journal of Molecular Sciences 22, no. 8: 3961. https://doi.org/10.3390/ijms22083961
APA StyleHayder, H., Fu, G., Nadeem, L., O’Brien, J. A., Lye, S. J., & Peng, C. (2021). Overexpression of miR-210-3p Impairs Extravillous Trophoblast Functions Associated with Uterine Spiral Artery Remodeling. International Journal of Molecular Sciences, 22(8), 3961. https://doi.org/10.3390/ijms22083961