
Confocal Blood Flow Videomicroscopy of Thrombus Formation over Human Arteries and Local Targeting of P2X7

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
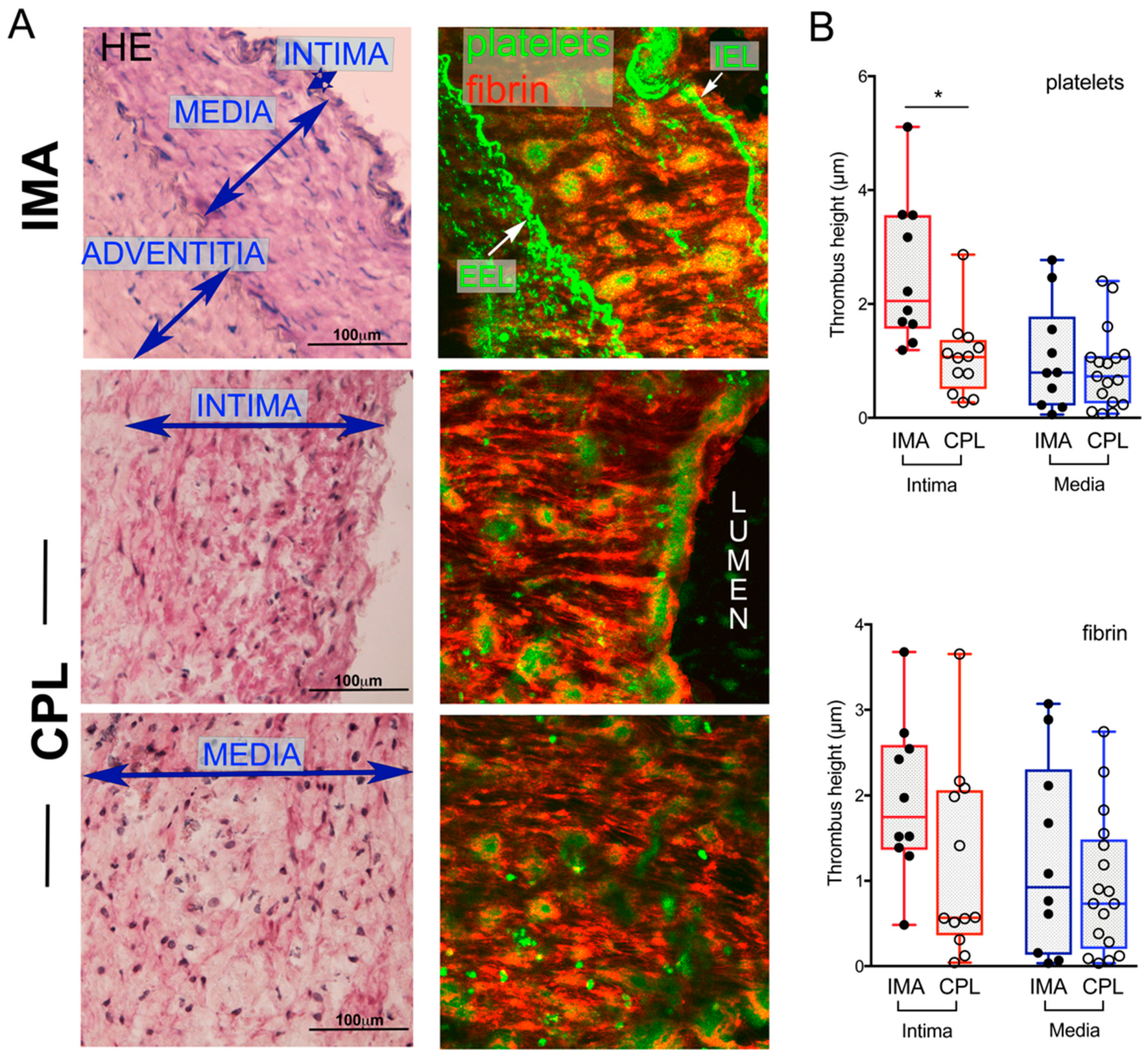
2.1. Thrombogenic Response of IMA and CPL Sections
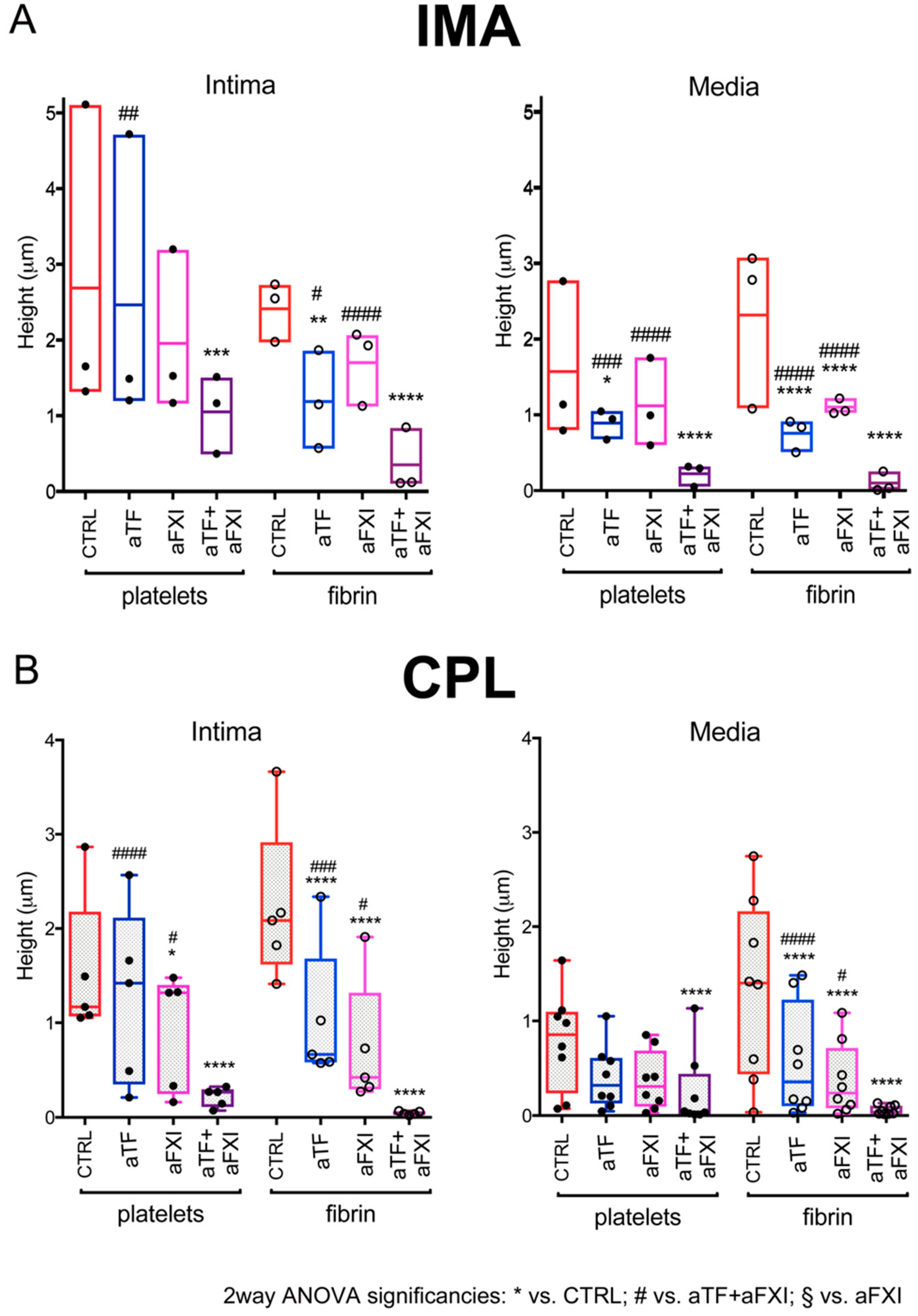
2.2. Effects of Selective Blockade of Coagulation Pathways on Thrombogenesis over IMA and CPL
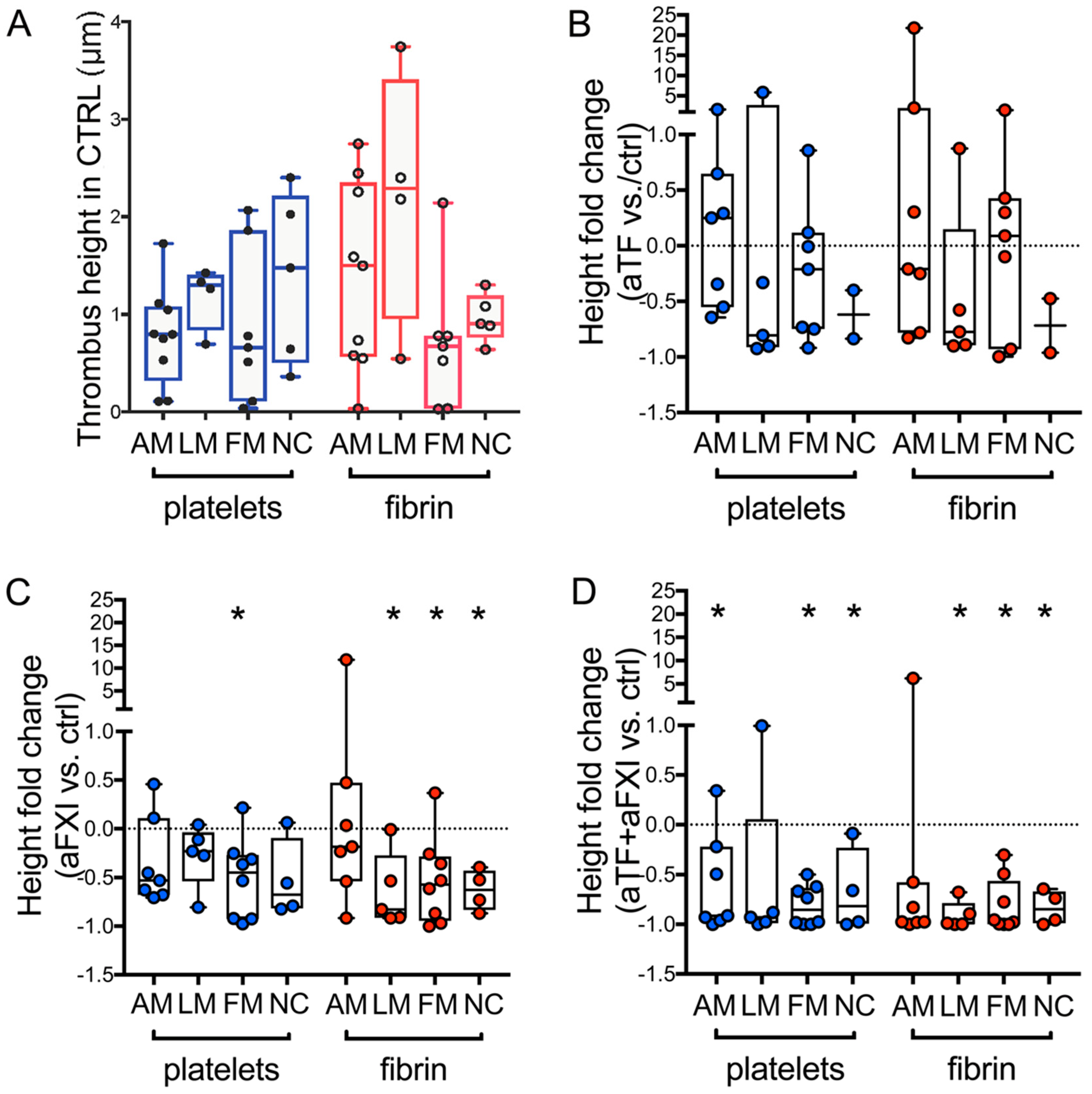
2.3. Sub Analysis of CPL Media with Morphological Alterations: Effects of Selective Blockade of Coagulation Pathways on Thrombogenicity
2.4. Sub Analysis of CPL Intima with Distinct Histological Features: Effects of Selective Blockade of Coagulation Pathways on Thrombogenicity
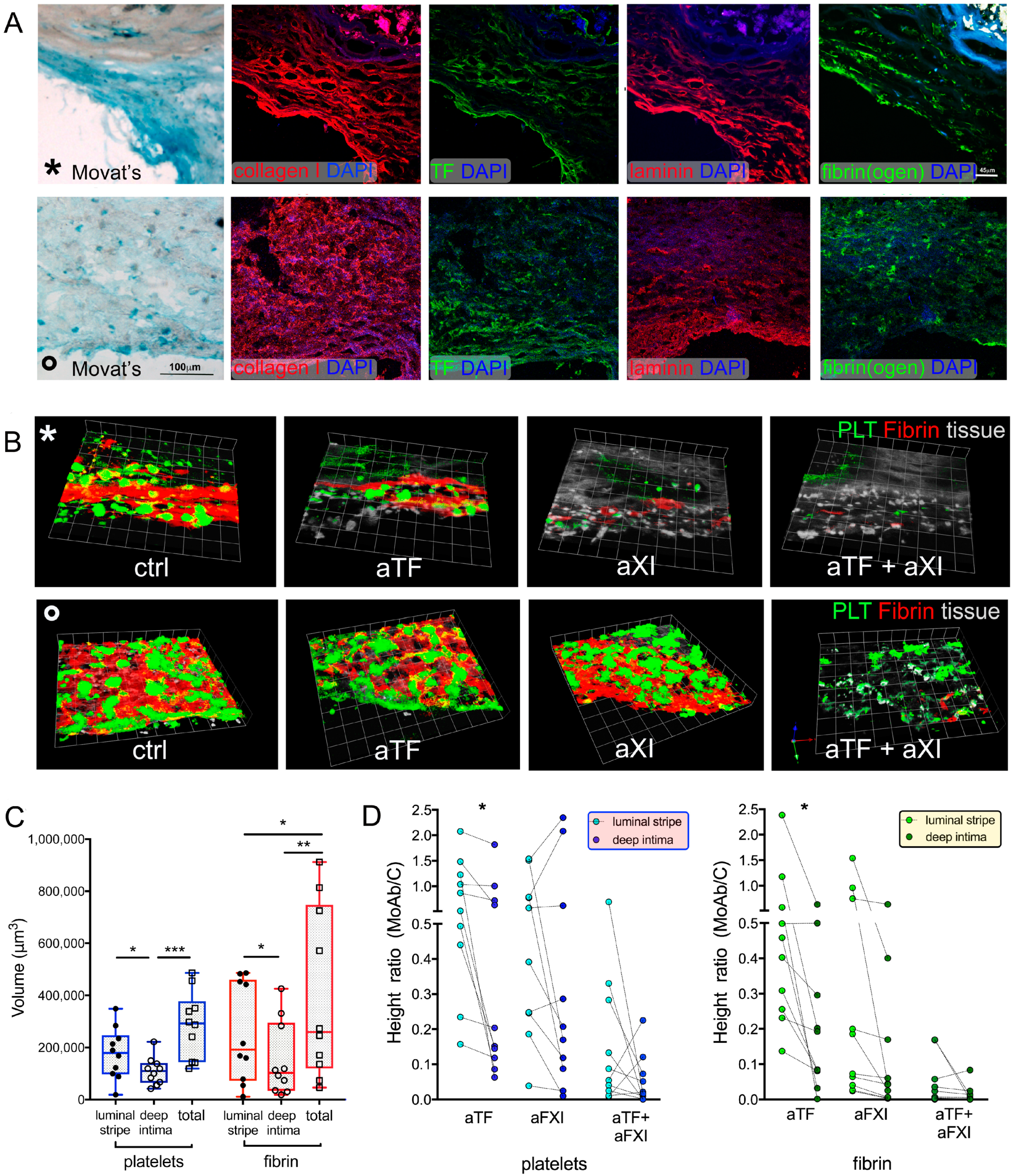
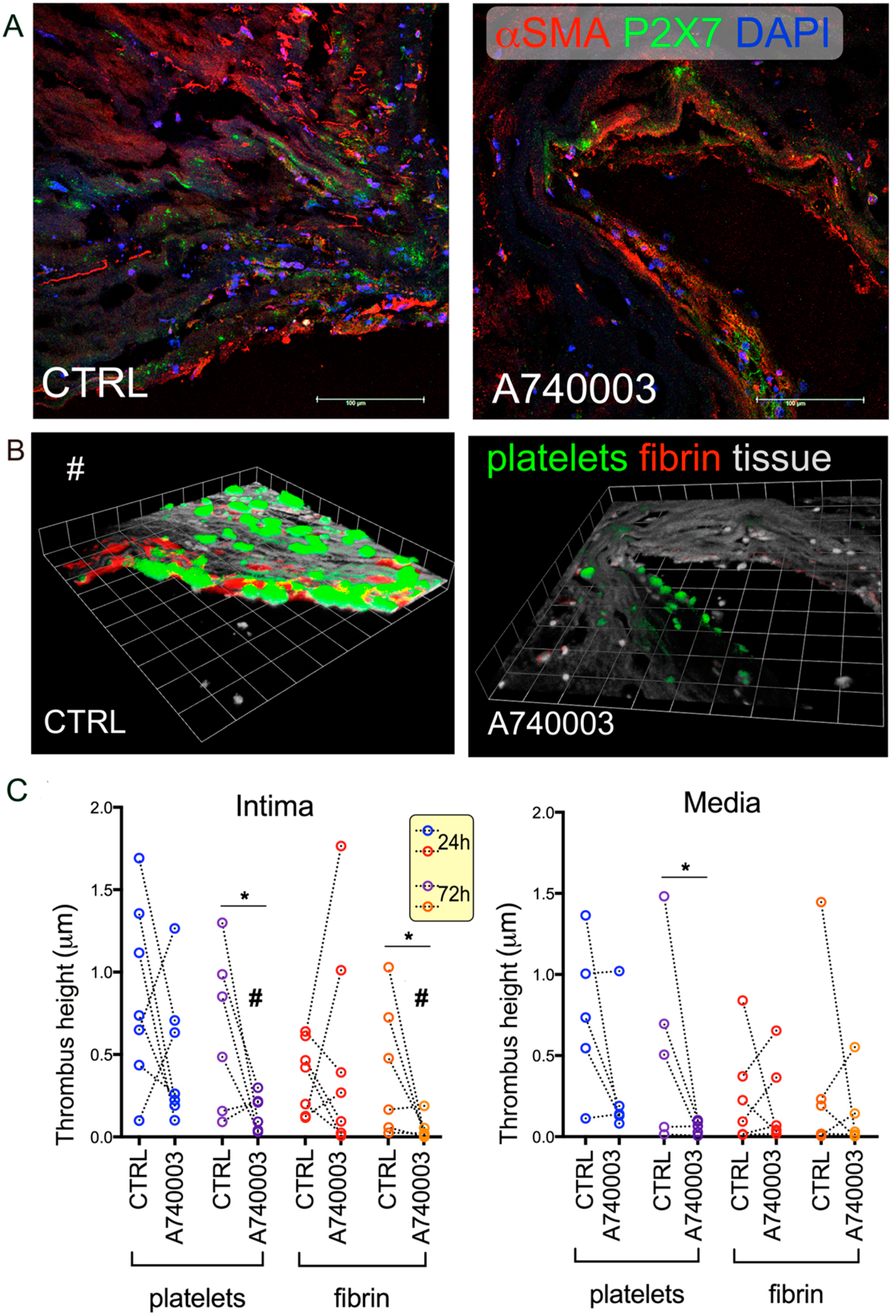
2.5. Effect of Vascular Tissue Pre-Treatment with P2X7 Antagonist on Thrombogenesis
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. CPL and IMA Processing
4.3. Histology and Immunofluorescence
4.4. Confocal Blood Flow Videomicroscopy Experiments
4.5. Statistical Analysis
5. Conclusions
6. Study Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badimon, L.; Vilahur, G. Thrombosis formation on atherosclerotic lesions and plaque rupture. J. Intern. Med. 2014, 276, 618–632. [Google Scholar] [CrossRef]
- Bentzon, J.F.; Otsuka, F.; Virmani, R.; Falk, E. Mechanisms of plaque formation and rupture. Circ. Res. 2014, 114, 1852–1866. [Google Scholar] [CrossRef]
- Gallone, G.; Baldetti, L.; Pagnesi, M.; Latib, A.; Colombo, A.; Libby, P.; Giannini, F. Medical Therapy for Long-Term Prevention of Atherothrombosis Following an Acute Coronary Syndrome: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2018, 72, 2886–2903. [Google Scholar] [CrossRef]
- Kubo, T.; Maehara, A.; Mintz, G.S.; Doi, H.; Tsujita, K.; Choi, S.Y.; Katoh, O.; Nasu, K.; Koenig, A.; Pieper, M.; et al. The dynamic nature of coronary artery lesion morphology assessed by serial virtual histology intravascular ultrasound tissue characterization. J. Am. Coll. Cardiol. 2010, 55, 1590–1597. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, A.C.; Truijman, M.T.; Hussain, B.; Zadi, T.; Saiedie, G.; de Rotte, A.A.; Liem, M.I.; van der Steen, A.F.; Daemen, M.J.; Koudstaal, P.J.; et al. Intraplaque Hemorrhage and the Plaque Surface in Carotid Atherosclerosis: The Plaque At RISK Study (PARISK). AJNR Am. J. Neuroradiol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, K.S.; Gotlieb, A.I. The role of shear stress in the pathogenesis of atherosclerosis. Lab. Investig. 2005, 85, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depairon, M.; Chessex, S.; Sudre, P.; Rodondi, N.; Doser, N.; Chave, J.P.; Riesen, W.; Nicod, P.; Darioli, R.; Telenti, A.; et al. Premature atherosclerosis in HIV-infected individuals--focus on protease inhibitor therapy. AIDS 2001, 15, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Harloff, A. Carotid plaque hemodynamics. Interv. Neurol. 2012, 1, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Range, H.; Labreuche, J.; Louedec, L.; Rondeau, P.; Planesse, C.; Sebbag, U.; Bourdon, E.; Michel, J.B.; Bouchard, P.; Meilhac, O. Periodontal bacteria in human carotid atherothrombosis as a potential trigger for neutrophil activation. Atherosclerosis 2014, 236, 448–455. [Google Scholar] [CrossRef]
- Levi, M.; van der Poll, T.; Buller, H.R. Bidirectional relation between inflammation and coagulation. Circulation 2004, 109, 2698–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmeier, W.; Hynes, R.O. Extracellular matrix proteins in hemostasis and thrombosis. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Cosemans, J.M.; Kuijpers, M.J.; Lecut, C.; Loubele, S.T.; Heeneman, S.; Jandrot-Perrus, M.; Heemskerk, J.W. Contribution of platelet glycoprotein VI to the thrombogenic effect of collagens in fibrous atherosclerotic lesions. Atherosclerosis 2005, 181, 19–27. [Google Scholar] [CrossRef]
- Crawley, J.; Lupu, F.; Westmuckett, A.D.; Severs, N.J.; Kakkar, V.V.; Lupu, C. Expression, localization, and activity of tissue factor pathway inhibitor in normal and atherosclerotic human vessels. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1362–1373. [Google Scholar] [CrossRef]
- Razuvaev, A.; Ekstrand, J.; Folkersen, L.; Agardh, H.; Markus, D.; Swedenborg, J.; Hansson, G.K.; Gabrielsen, A.; Paulsson-Berne, G.; Roy, J.; et al. Correlations between clinical variables and gene-expression profiles in carotid plaque instability. Eur. J. Vasc. Endovasc. Surg. 2011, 42, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Reininger, A.J.; Bernlochner, I.; Penz, S.M.; Ravanat, C.; Smethurst, P.; Farndale, R.W.; Gachet, C.; Brandl, R.; Siess, W. A 2-step mechanism of arterial thrombus formation induced by human atherosclerotic plaques. J. Am. Coll. Cardiol. 2010, 55, 1147–1158. [Google Scholar] [CrossRef] [Green Version]
- Sayed, S.; Cockerill, G.W.; Torsney, E.; Poston, R.; Thompson, M.M.; Loftus, I.M. Elevated Tissue Expression of Thrombomodulatory Factors Correlates with Acute Symptomatic Carotid Plaque Phenotype. Eur. J. Vasc. Endovasc. Surg. 2009, 38, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsumi, K.; Mackman, N. Tissue Factor and Atherothrombosis. J. Atheroscler. Thromb. 2015, 22, 543–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toschi, V.; Gallo, R.; Lettino, M.; Fallon, J.T.; Gertz, S.D.; Fernandez-Ortiz, A.; Chesebro, J.H.; Badimon, L.; Nemerson, Y.; Fuster, V.; et al. Tissue Factor Modulates the Thrombogenicity of Human Atherosclerotic Plaques. Circulation 1997, 95, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.B. Fibrinogen, fibrin and the arterial wall. Eur. Heart J. 1995, 16 (Suppl. A), 11–14; discussion 14–15. [Google Scholar] [CrossRef]
- Chen, Y.; Li, G.; Liu, M.L. Microvesicles as Emerging Biomarkers and Therapeutic Targets in Cardiometabolic Diseases. Genom. Proteom. Bioinform. 2018, 16, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Cesselli, D.; Parisse, P.; Aleksova, A.; Veneziano, C.; Cervellin, C.; Zanello, A.; Beltrami, A.P. Extracellular Vesicles: How Drug and Pathology Interfere With Their Biogenesis and Function. Front. Physiol. 2018, 9, 1394. [Google Scholar] [CrossRef] [PubMed]
- Van Kruchten, R.; Cosemans, J.M.; Heemskerk, J.W. Measurement of whole blood thrombus formation using parallel-plate flow chambers—A practical guide. Platelets 2012, 23, 229–242. [Google Scholar] [CrossRef]
- Ruggeri, Z.M. Platelet adhesion under flow. Microcirculation 2009, 16, 58–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, L.H.; Feghhi, S.; Taparia, N.; Smith, A.O.; Karchin, A.; Lim, E.; John, A.S.; Wang, X.; Rue, T.; White, N.J.; et al. Contractile forces in platelet aggregates under microfluidic shear gradients reflect platelet inhibition and bleeding risk. Nat. Commun. 2019, 10, 1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Neelamegham, S. Application of microfluidic devices in studies of thrombosis and hemostasis. Platelets 2017, 28, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Yang, J.; Huang, J.; Long, Z.; Ruan, Z.; Xiao, B.; Xi, X. Effects of different shear rates on the attachment and detachment of platelet thrombi. Mol. Med. Rep. 2016, 13, 2447–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackman, N. New targets for atherothrombosis. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1607–1608. [Google Scholar] [CrossRef] [Green Version]
- Penz, S.; Reininger, A.J.; Brandl, R.; Goyal, P.; Rabie, T.; Bernlochner, I.; Rother, E.; Goetz, C.; Engelmann, B.; Smethurst, P.A.; et al. Human atheromatous plaques stimulate thrombus formation by activating platelet glycoprotein VI. FASEB J. 2005, 19, 898–909. [Google Scholar] [CrossRef] [Green Version]
- Foglieni, C.; Maisano, F.; Dreas, L.; Giazzon, A.; Ruotolo, G.; Ferrero, E.; Li Volsi, L.; Coli, S.; Sinagra, G.; Zingone, B.; et al. Mild inflammatory activation of mammary arteries in patients with acute coronary syndromes. Am. J. Physiol. Heart. Circ. Physiol. 2008, 294, H2831–H2837. [Google Scholar] [CrossRef]
- Otsuka, F.; Yahagi, K.; Sakakura, K.; Virmani, R. Why is the mammary artery so special and what protects it from atherosclerosis? Ann. Cardiothorac. Surg. 2013, 2, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Piscopiello, M.; Sessa, M.; Anzalone, N.; Castellano, R.; Maisano, F.; Ferrero, E.; Chiesa, R.; Alfieri, O.; Comi, G.; Ferrero, M.E.; et al. P2X7 receptor is expressed in human vessels and might play a role in atherosclerosis. Int. J. Cardiol. 2013, 168, 2863–2866. [Google Scholar] [CrossRef] [PubMed]
- Furlan-Freguia, C.; Marchese, P.; Gruber, A.; Ruggeri, Z.M.; Ruf, W. P2X7 receptor signaling contributes to tissue factor-dependent thrombosis in mice. J. Clin. Investig. 2011, 121, 2932–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, Y.; Xin, G.; Ji, B.H.; Ji, C.J.; Wei, Z.L.; Zhang, B.L.; Zhang, J.H.; Yu, K.; Zhang, X.Y.; Li, S.Y.; et al. Entecavir as a P2X7R antagonist ameliorates platelet activation and thrombus formation. J. Pharmacol. Sci. 2020, 144, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Olie, R.H.; van der Meijden, P.E.J.; Spronk, H.M.H.; Ten Cate, H. Antithrombotic Therapy: Prevention and Treatment of Atherosclerosis and Atherothrombosis. Handb. Exp. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Honore, P.; Donnelly-Roberts, D.; Namovic, M.T.; Hsieh, G.; Zhu, C.Z.; Mikusa, J.P.; Hernandez, G.; Zhong, C.; Gauvin, D.M.; Chandran, P.; et al. A-740003 [N-(1-{[(cyanoimino)(5-quinolinylamino) methyl]amino}-2,2-dimethylpropyl)-2-(3,4-dimethoxyphenyl)acetamide], a novel and selective P2X7 receptor antagonist, dose-dependently reduces neuropathic pain in the rat. J. Pharmacol. Exp. Ther. 2006, 319, 1376–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shindo, S.; Fujii, K.; Shirakawa, M.; Uchida, K.; Enomoto, Y.; Iwama, T.; Kawasaki, M.; Ando, Y.; Yoshimura, S. Morphologic Features of Carotid Plaque Rupture Assessed by Optical Coherence Tomography. AJNR Am. J. Neuroradiol. 2015, 36, 2140–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, M.; Mantione, M.E.; Baccellieri, D.; Ferrara, D.; Castellano, R.; Chiesa, R.; Alfieri, O.; Foglieni, C. P2X7 receptor antagonism modulates IL-1beta and MMP9 in human atherosclerotic vessels. Sci. Rep. 2017, 7, 4872. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Yamamoto, S.; Funayama, H.; Mitsuhashi, T.; Momomura, S.-I. Acute occlusion of the left internal mammary artery graft in the late postoperative period. J. Cardiol. Cases 2014, 10, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Yong, A.; Groenestein, P.; Brieger, D.; Lowe, H.; Kritharides, L. Late thrombotic occlusion of a left internal mammary artery graft causing ST-elevation myocardial infarction. Int. J. Cardiol. 2010, 142, e42–e44. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef]
- Kearney, K.J.; Butler, J.; Posada, O.M.; Wilson, C.; Heal, S.; Ali, M.; Hardy, L.; Ahnström, J.; Gailani, D.; Foster, R.; et al. Kallikrein directly interacts with and activates Factor IX, resulting in thrombin generation and fibrin formation independent of Factor XI. Proc. Nat. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Puy, C.; Tucker, E.I.; Matafonov, A.; Cheng, Q.; Zientek, K.D.; Gailani, D.; Gruber, A.; McCarty, O.J. Activated factor XI increases the procoagulant activity of the extrinsic pathway by inactivating tissue factor pathway inhibitor. Blood 2015, 125, 1488–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, N.C.; Eikelboom, J.W.; Weitz, J.I. Evolving Treatments for Arterial and Venous Thrombosis: Role of the Direct Oral Anticoagulants. Circ. Res. 2016, 118, 1409–1424. [Google Scholar] [CrossRef] [Green Version]
- Gradoli, J.; Vidal, V.; Brady, A.J.; Facila, L. Anticoagulation in Patients with Ischaemic Heart Disease and Peripheral Arterial Disease: Clinical Implications of COMPASS Study. Eur. Cardiol. 2018, 13, 115–118. [Google Scholar] [CrossRef]
- Grover, S.P.; Mackman, N. Intrinsic Pathway of Coagulation and Thrombosis. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Vojacek, J.F. Should We Replace the Terms Intrinsic and Extrinsic Coagulation Pathways With Tissue Factor Pathway? Clin. Appl. Thromb. Hemost. 2017, 23, 922–927. [Google Scholar] [CrossRef]
- Badimon, J.J.; Lettino, M.; Toschi, V.; Fuster, V.; Berrozpe, M.; Chesebro, J.H.; Badimon, L. Local inhibition of tissue factor reduces the thrombogenicity of disrupted human atherosclerotic plaques: Effects of tissue factor pathway inhibitor on plaque thrombogenicity under flow conditions. Circulation 1999, 99, 1780–1787. [Google Scholar] [CrossRef] [Green Version]
- Arulkumaran, N.; Unwin, R.J.; Tam, F.W.K. A potential therapeutic role for P2X7 receptor (P2X7R) antagonists in the treatment of inflammatory diseases. Expert Opin. Inv. Drug 2011, 20, 897–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantione, M.E.; Lombardi, M.; Baccellieri, D.; Ferrara, D.; Castellano, R.; Chiesa, R.; Alfieri, O.; Foglieni, C. IL-1beta/MMP9 activation in primary human vascular smooth muscle-like cells: Exploring the role of TNFalpha and P2X7. Int. J. Cardiol. 2019, 278, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Virmani, R.; Kolodgie, F.D.; Burke, A.P.; Farb, A.; Schwartz, S.M. Lessons from sudden coronary death: A comprehensive morphological classification scheme for atherosclerotic lesions. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1262–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genkel, V.V.; Salashenko, A.O.; Shamaeva, T.N.; Sumerkina, V.A.; Shaposhnik, I.I. Association between Carotid Wall Shear Rate and Arterial Stiffness in Patients with Hypertension and Atherosclerosis of Peripheral Arteries. Int. J. Vasc. Med. 2018, 2018, 6486234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffel, J.; Luscher, T.F.; Tanner, F.C. Tissue factor in cardiovascular diseases: Molecular mechanisms and clinical implications. Circulation 2006, 113, 722–731. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | IMA | CPL | ||
|---|---|---|---|---|
| Intima | Media | Intima | Media | |
| TF (tissue factor) | +cells of the luminal lining; −/traces intima | +/traces | ++cells of the luminal lining and several intima cells * | ++ ‡ |
| Fibrin(ogen) | +cells of the luminal lining; −intima | − | ++ not continuous along the luminal lining; +/−intima † | +/++cells *, † |
| vWF (von Willebrand factor) | ++ not continuous along the luminal lining; intima | − | ++ not continuous along the luminal lining; −intima | traces/– |
| αSMA (alpha smooth muscle actin) | +intima | +++ | +/++ † | +/++ †, § |
| SM22 (smooth muscle protein 22) | +several intima cells | + | ++scattered cells | + † |
| SMMHC (smooth muscle myosin heavy chain) | +intima | ++ | +rare cells | ++scattered cells § |
| CD68 | − | ++rare cells | −; +several cells † | ++rare cells § |
| FSP1 (fibroblast-specific protein-1) | + not continuous along the luminal lining and/or several intima cells | +rare cells | +several intima cells | ++rare cells § |
| Vimentin | +++ not continuous along the luminal lining and in the intima | + | + | ++ § |
| Laminin | +++luminal lining;+intima | ++ | ++luminal lining; +/++intima † | +/− |
| Collagen type I | + | +/traces | +/++ | +/++ † |
| P2X7 | traces | −/traces | + † | +/traces/− † |
| CPL Media Subtypes | ||||
|---|---|---|---|---|
| AM | LM | FM | Necrotic Core | |
| TF (tissue factor) | ++ ‡; +/traces other cells * | ++/− *, † | −; ++ ‡ | ++/− *, † |
| Fibrin(ogen) | ++/− † | ++ | traces/++ *, † | traces/– |
| vWF (von Willebrand factor) | − | − | − | − |
| αSMA (alpha smooth muscle actin) | + † | + rare cells | − | − |
| SM22 (smooth muscle protein 22) | ++ scattered /rare cells | + rare cells | ++rare cells | − |
| SMMHC (smoot muscle myosin heavy chain) | ++ scattered /rare cells | + rare cells | +rare cells | − (+ rare cells) † |
| CD68 | + rare cells | ++ scattered cells | ++rare cells | − |
| FSP1 (fibroblast-specific protein-1) | traces | +/traces | – | traces/− |
| Vimentin | +/++ † | traces/– | – | − |
| Laminin | +/− † | − | − | − |
| Collagen type I | +/++ † | +/traces † | traces/– | traces/− |
| P2X7 | traces/− | +/traces | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchese, P.; Lombardi, M.; Mantione, M.E.; Baccellieri, D.; Ferrara, D.; Chiesa, R.; Alfieri, O.; Foglieni, C. Confocal Blood Flow Videomicroscopy of Thrombus Formation over Human Arteries and Local Targeting of P2X7. Int. J. Mol. Sci. 2021, 22, 4066. https://doi.org/10.3390/ijms22084066
Marchese P, Lombardi M, Mantione ME, Baccellieri D, Ferrara D, Chiesa R, Alfieri O, Foglieni C. Confocal Blood Flow Videomicroscopy of Thrombus Formation over Human Arteries and Local Targeting of P2X7. International Journal of Molecular Sciences. 2021; 22(8):4066. https://doi.org/10.3390/ijms22084066
Chicago/Turabian StyleMarchese, Patrizia, Maria Lombardi, Maria Elena Mantione, Domenico Baccellieri, David Ferrara, Roberto Chiesa, Ottavio Alfieri, and Chiara Foglieni. 2021. "Confocal Blood Flow Videomicroscopy of Thrombus Formation over Human Arteries and Local Targeting of P2X7" International Journal of Molecular Sciences 22, no. 8: 4066. https://doi.org/10.3390/ijms22084066
APA StyleMarchese, P., Lombardi, M., Mantione, M. E., Baccellieri, D., Ferrara, D., Chiesa, R., Alfieri, O., & Foglieni, C. (2021). Confocal Blood Flow Videomicroscopy of Thrombus Formation over Human Arteries and Local Targeting of P2X7. International Journal of Molecular Sciences, 22(8), 4066. https://doi.org/10.3390/ijms22084066