
Resveratrol Alleviates the Inhibitory Effect of Tunicamycin-Induced Endoplasmic Reticulum Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes


Abstract
:1. Introduction
2. Results
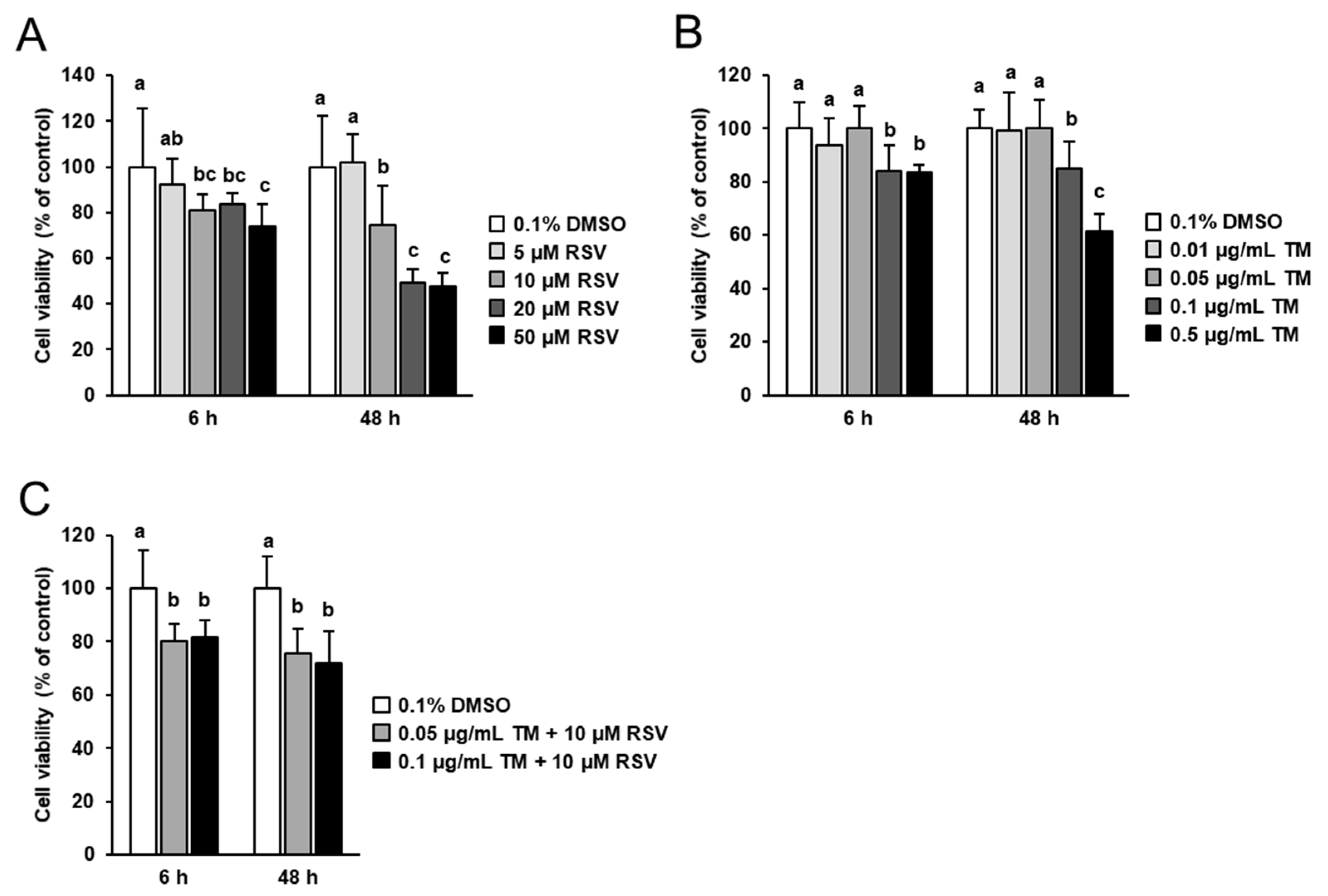
2.1. Effect of RSV and TM on FRTL-5 Cell Viability
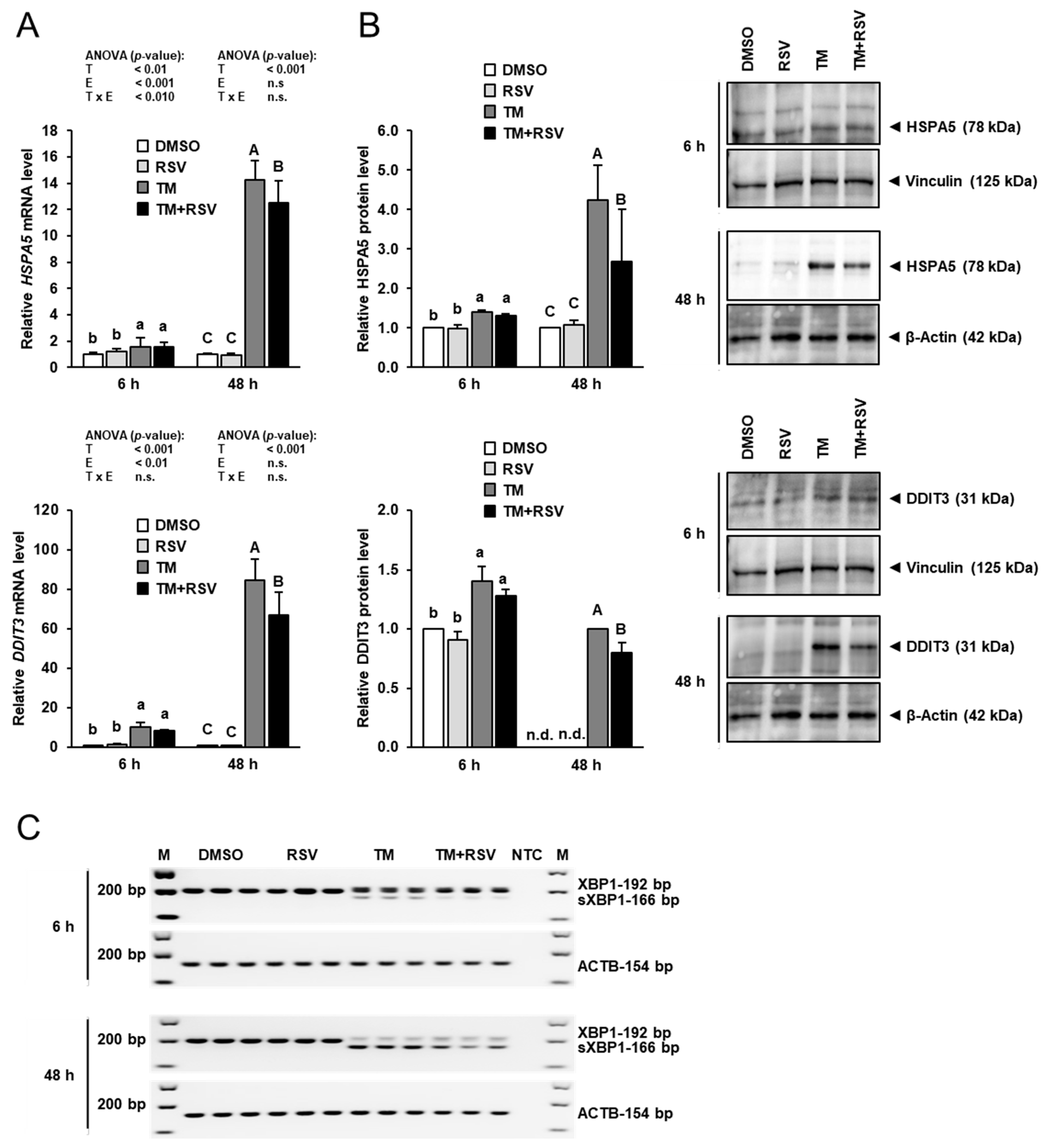
2.2. Effect of RSV on TM-Induced ER Stress in FRTL-5 Cells
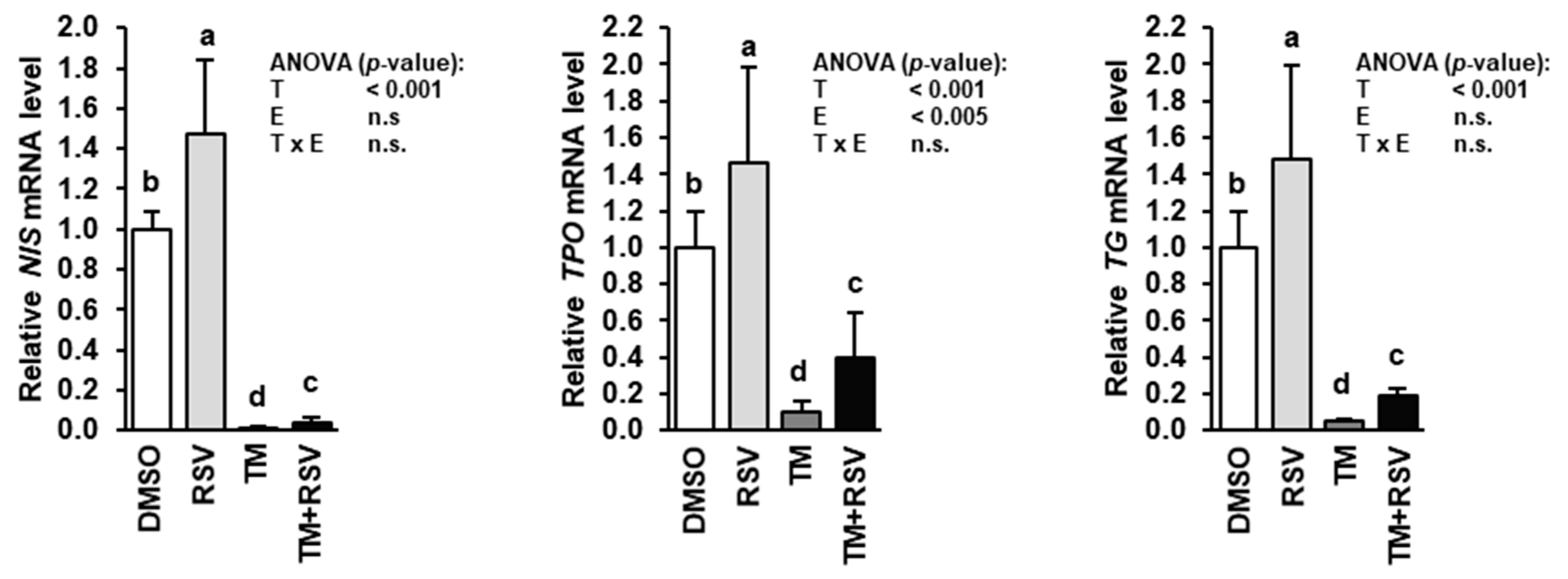
2.3. RSV Alleviates the Inhibitory Effect of TM-Induced ER Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes
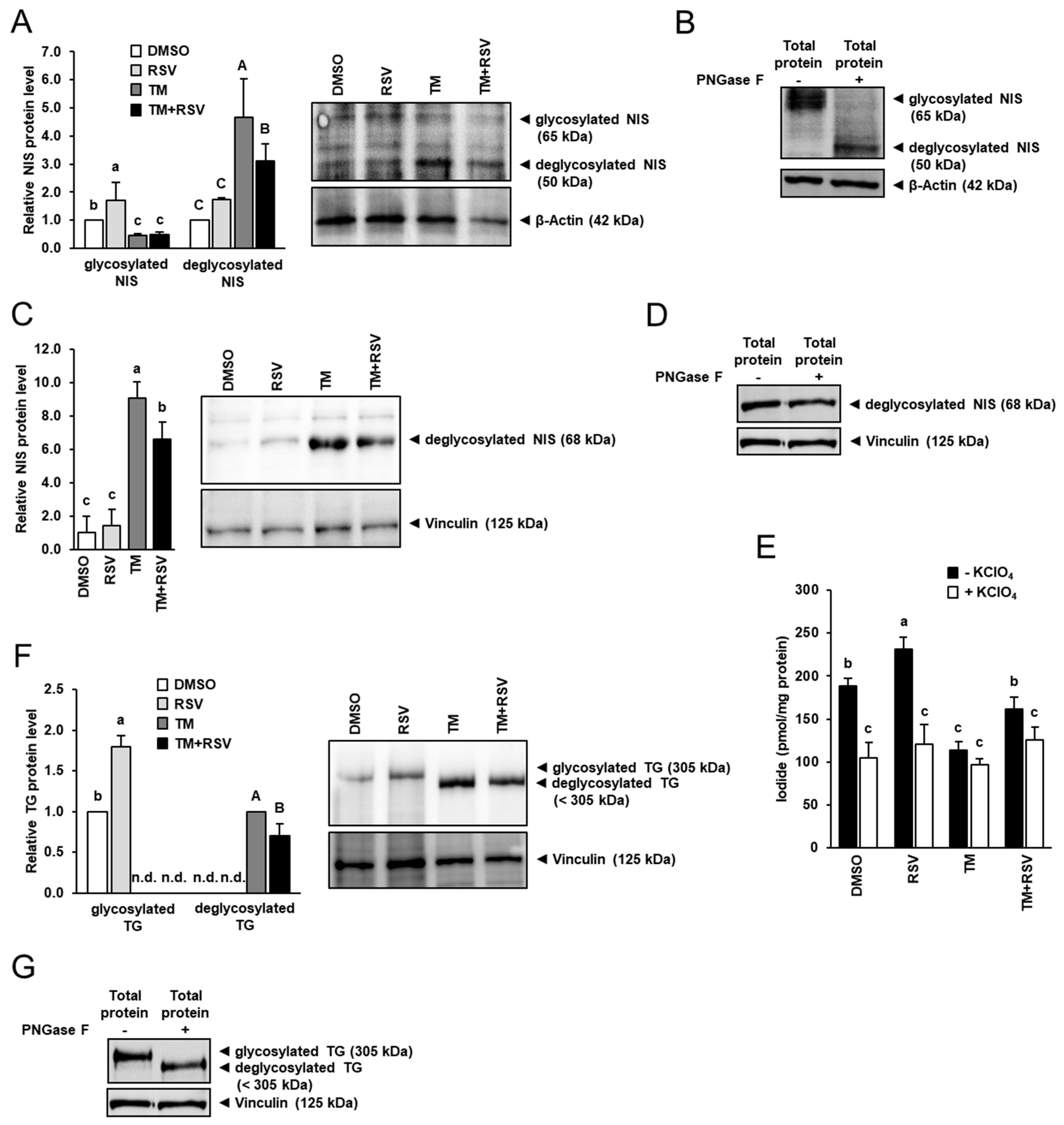
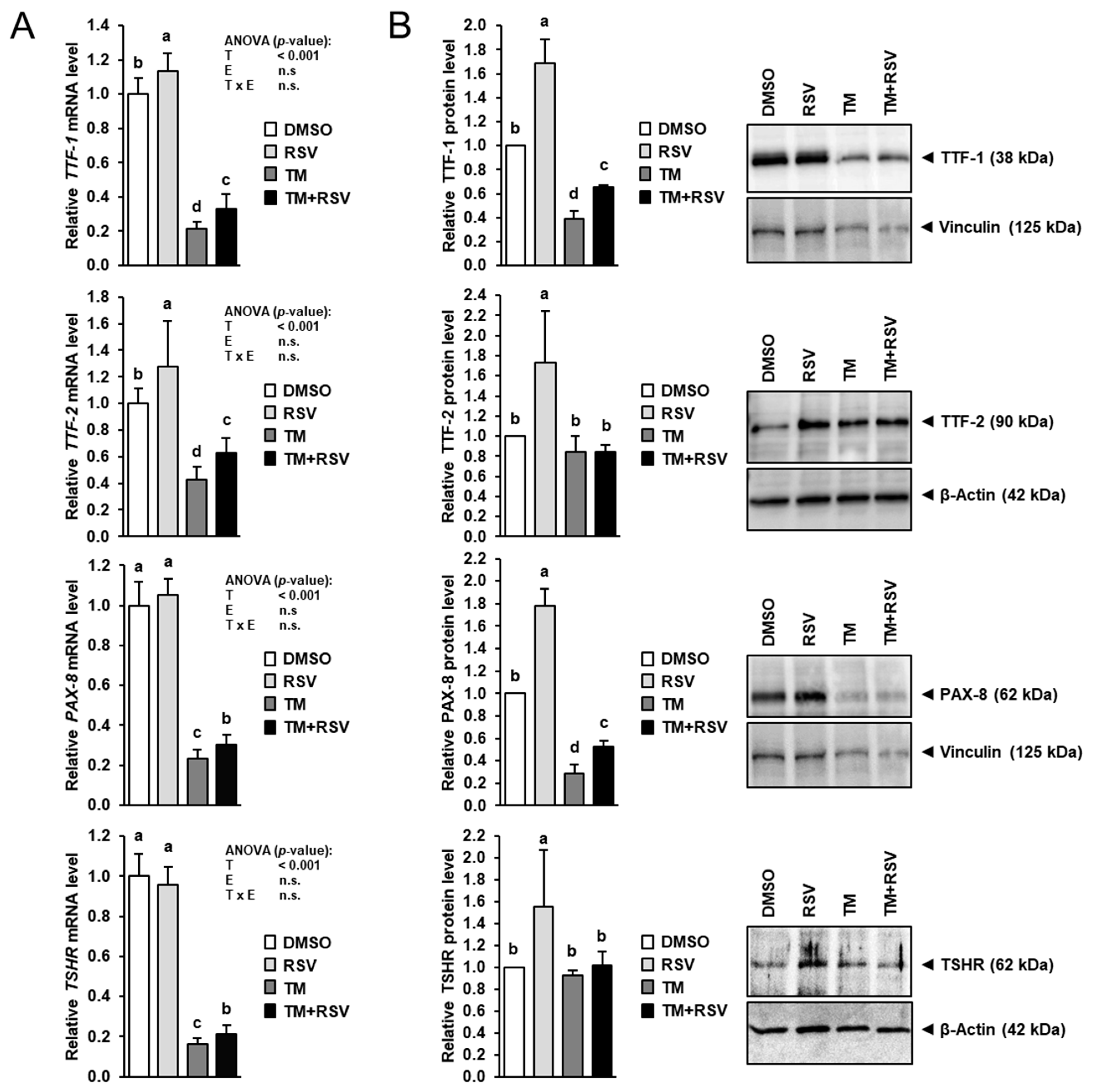
2.4. RSV Alleviates the Inhibitory Effect of TM-Induced ER Stress on Expression of Key Regulators of Thyroid Hormone Synthesis in FRTL-5 Thyrocytes
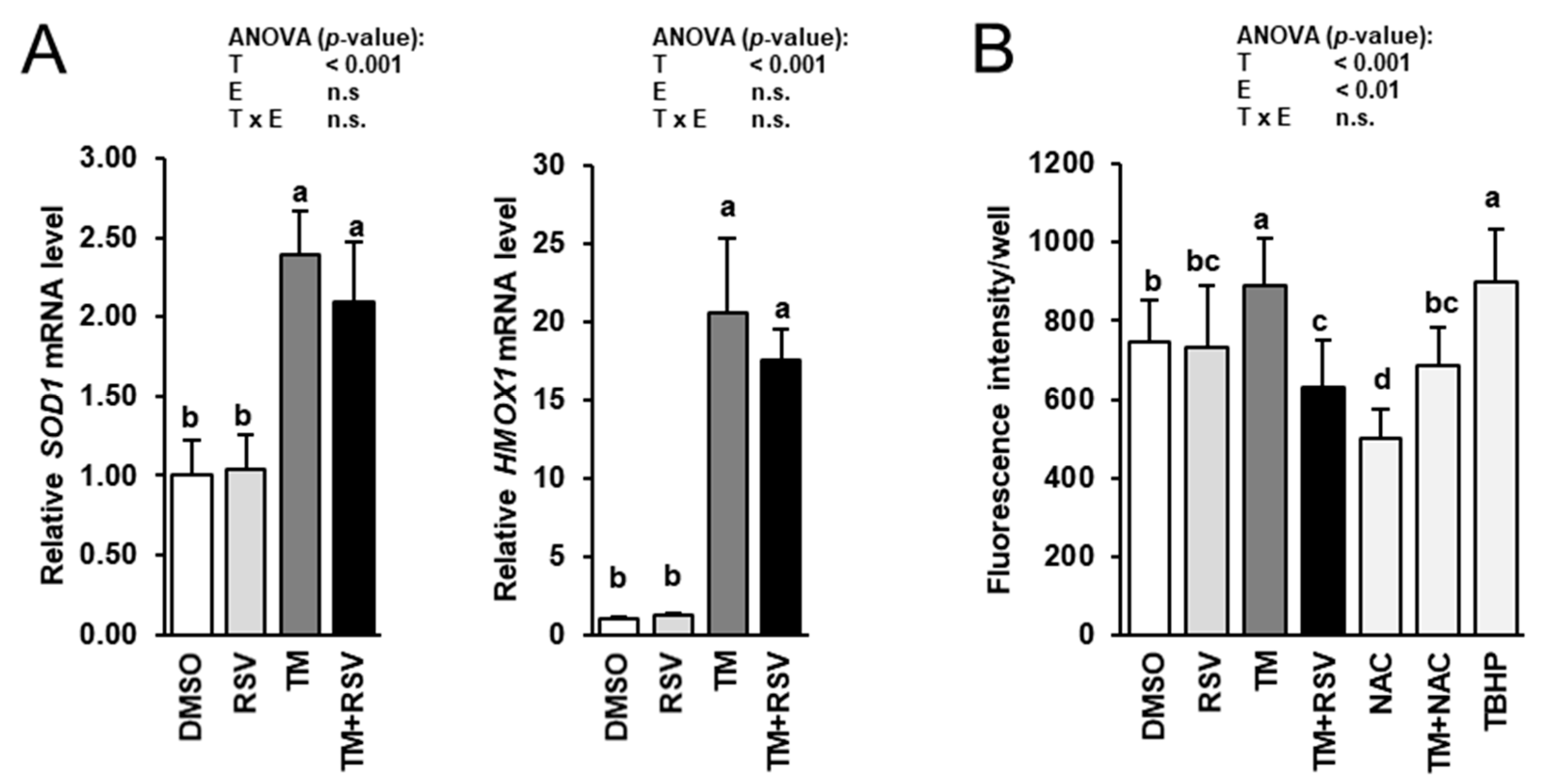
2.5. RSV Does Not Modulate TM-Induced Induction of Cytoprotective Genes but Decreases ROS Production in FRTL-5 Thyrocytes
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability Assay
4.3. RNA Isolation and qPCR Analysis
4.4. Protein Separation and Immunoblotting
4.5. Analysis of NIS and TG Deglycosylation
4.6. Nonradioactive Iodide Uptake Assay
4.7. Intracellular ROS Production Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oakes, S.A. Endoplasmic reticulum proteostasis: A key checkpoint in cancer. Am. J. Physiol. Cell Physiol. 2017, 312, C93–C102. [Google Scholar] [CrossRef] [Green Version]
- Tu, B.P.; Weissman, J.S. Oxidative protein folding in eukaryotes: Mechanisms and consequences. J. Cell Biol. 2004, 164, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Kaufman, R.J. Signaling the unfolded protein response from the endoplasmic reticulum. J. Biol. Chem. 2004, 279, 25935–25938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.H.; Iwakoshi, N.N.; Glimcher, L.H. XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Mol. Cell Biol. 2003, 23, 7448–7459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, A.; Vito, P.; Mauro, C.; Pacifico, F.; Ulianich, L.; Consiglio, E.; Formisano, S.; Di Jeso, B. Endoplasmic reticulum stress causes thyroglobulin retention in this organelle and triggers activation of nuclear factor-kappa B via tumor necrosis factor receptor-associated factor 2. Endocrinology 2002, 143, 2169–2177. [Google Scholar] [CrossRef]
- Baryshev, M.; Sargsyan, E.; Wallin, G.; Lejnieks, A.; Furudate, S.; Hishinuma, A.; Mkrtchian, S. Unfolded protein response is involved in the pathology of human congenital hypothyroid goiter and rat non-goitrous congenital hypothyroidism. J. Mol. Endocrinol. 2004, 32, 903–920. [Google Scholar] [CrossRef] [Green Version]
- Kim, P.S.; Kwon, O.Y.; Arvan, P. An endoplasmic reticulum storage disease causing congenital goiter with hypothyroidism. J. Cell Biol. 1996, 133, 517–527. [Google Scholar] [CrossRef]
- Medeiros-Neto, G.; Kim, P.S.; Yoo, S.E.; Vono, J.; Targovnik, H.M.; Camargo, R.; Hossain, S.A.; Arvan, P. Congenital hypothyroid goiter with deficient thyroglobulin. Identification of an endoplasmic reticulum storage disease with induction of molecular chaperones. J. Clin. Investig. 1996, 98, 2838–2844. [Google Scholar] [CrossRef]
- Gaide Chevronnay, H.P.; Janssens, V.; Van Der Smissen, P.; Liao, X.H.; Abid, Y.; Nevo, N.; Antignac, C.; Refetoff, S.; Cherqui, S.; Pierreux, C.E.; et al. A mouse model suggests two mechanisms for thyroid alterations in infantile cystinosis: Decreased thyroglobulin synthesis due to endoplasmic reticulum stress/unfolded protein response and impaired lysosomal processing. Endocrinology 2015, 156, 2349–2364. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, A.; Inabnet, W.B., 3rd; Owen, R.; Farenholtz, K.E.; Tomer, Y. Endoplasmic reticulum stress as a novel mechanism in amiodarone-induced destructive thyroiditis. J. Clin. Endocrinol. Metab. 2015, 100, E1–E10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Virgilio, M.; Kitzmüller, C.; Schwaiger, E.; Klein, M.; Kreibich, G.; Ivessa, N.E. Degradation of a short-lived glycoprotein from the lumen of the endoplasmic reticulum: The role of N-linked glycans and the unfolded protein response. Mol. Biol. Cell 1999, 10, 4059–4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, G.; Ringseis, R.; Eder, K. Endoplasmic reticulum stress inhibits expression of genes involved in thyroid hormone synthesis and their key transcriptional regulators in FRTL-5 thyrocytes. PLoS ONE 2017, 12, e0187561. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, A.J.; Lonigro, R.; Civitareale, D.; Ghibelli, L.; Di Lauro, R. The tissue-specific expression of the thyroglobulin gene requires interaction between thyroid-specific and ubiquitous factors. Eur. J. Biochem. 1990, 193, 311–318. [Google Scholar] [CrossRef]
- Endo, T.; Kaneshige, M.; Nakazato, M.; Ohmori, M.; Harii, N.; Onaya, T. Thyroid transcription factor-1 activates the promoter activity of rat thyroid Na+/I- symporter gene. Mol. Endocrinol. 1997, 11, 1747–1755. [Google Scholar]
- Esposito, C.; Miccadei, S.; Saiardi, A.; Civitareale, D. PAX 8 activates the enhancer of the human thyroperoxidase gene. Biochem. J. 1998, 331, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Kambe, F.; Nomura, Y.; Okamoto, T.; Seo, H. Redox regulation of thyroid-transcription factors, Pax-8 and TTF-1, is involved in their increased DNA-binding activities by thyrotropin in rat thyroid FRTL-5 cells. Mol. Endocrinol. 1996, 10, 801–812. [Google Scholar]
- Damante, G.; Tell, G.; Di Lauro, R. A unique combination of transcription factors controls differentiation of thyroid cells. Prog. Nucleic Acid. Res. Mol. Biol. 2001, 66, 307–356. [Google Scholar]
- Li, C.; Wang, L.; Huang, K.; Zheng, L. Endoplasmic reticulum stress in retinal vascular degeneration: Protective role of resveratrol. Invest. Ophthalmol. Vis. Sci. 2012, 53, 3241–3249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Hong, S.W.; Kwon, H.; Park, S.E.; Rhee, E.J.; Park, C.Y.; Oh, K.W.; Park, S.W.; Lee, W.Y. Resveratrol, an activator of SIRT1, improves ER stress by increasing clusterin expression in HepG2 cells. Cell Stress Chaperones 2019, 24, 825–833. [Google Scholar] [CrossRef]
- Pan, P.T.; Lin, H.Y.; Chuang, C.W.; Wang, P.K.; Wan, H.C.; Lee, M.C.; Kao, M.C. Resveratrol alleviates nuclear factor-κB-mediated neuroinflammation in vasculitic peripheral neuropathy induced by ischaemia-reperfusion via suppressing endoplasmic reticulum stress. Clin. Exp. Pharmacol. Physiol. 2019, 46, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, Y.; Shu, L.; Song, G.; Ma, H. Resveratrol reduces liver endoplasmic reticulum stress and improves insulin sensitivity in vivo and in vitro. Drug Des. Devel. Ther. 2019, 13, 1473–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Dong, X.J.; Ding, M.R.; You, C.Y.; Lin, X.; Wang, Y.; Wu, M.J.; Xu, G.F.; Wang, G.D. Resveratrol decreases high glucose-induced apoptosis in renal tubular cells via suppressing endoplasmic reticulum stress. Mol. Med. Rep. 2020, 22, 4367–4375. [Google Scholar] [PubMed]
- Shati, A.A. Resveratrol protects against cadmium chloride-induced hippocampal neurotoxicity by inhibiting ER stress and GAAD 153 and activating sirtuin 1/AMPK/Akt. Environ. Toxicol. 2019, 34, 1340–1353. [Google Scholar] [CrossRef]
- Sebai, H.; Hovsépian, S.; Ristorcelli, E.; Aouani, E.; Lombardo, D.; Fayet, G. Resveratrol increases iodide trapping in the rat thyroid cell line FRTL-5. Thyroid 2010, 20, 195–203. [Google Scholar] [CrossRef]
- Yu, X.M.; Jaskula-Sztul, R.; Ahmed, K.; Harrison, A.D.; Kunnimalaiyaan, M.; Chen, H. Resveratrol induces differentiation markers expression in anaplastic thyroid carcinoma via activation of Notch1 signaling and suppresses cell growth. Mol. Cancer Ther. 2013, 12, 1276–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, C.; Bucci, I.; Di Santo, S.; Rossi, C.; Grassadonia, A.; Mariotti, M.; Piantelli, M.; Monaco, F.; Napolitano, G. Resveratrol inhibits sodium/iodide symporter gene expression and function in rat thyroid cells. PLoS ONE 2014, 9, e107936. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, C.; Iezzi, M.; Ciolli, L.; Hysi, A.; Bucci, I.; Di Santo, S.; Rossi, C.; Zucchelli, M.; Napolitano, G. Resveratrol has anti-thyroid effects both in vitro and in vivo. Food Chem. Toxicol. 2017, 107, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Paik, M.J.; Kim, Y.J.; Lee, G.; Lee, Y.S.; Choi, H.D.; Kim, B.C.; Pack, J.K.; Kim, N.; Ahn, Y.H. Effects of whole-body exposure to 915 MHz RFID on secretory functions of the thyroid system in rats. Bioelectromagnetics 2013, 34, 521–529. [Google Scholar] [CrossRef]
- Maliković, J.; Feyissa, D.D.; Kalaba, P.; Marouf, B.S.; Höger, H.; Hartmann, M.F.; Wudy, S.A.; Schuler, G.; Lubec, G.; Aradska, J.; et al. Age and cognitive status dependent differences in blood steroid and thyroid hormone concentrations in intact male rats. Behav. Brain Funct. 2019, 15, 10. [Google Scholar] [CrossRef]
- Davies, T.F.; Platzer, M. hCG-induced TSH receptor activation and growth acceleration in FRTL-5 thyroid cells. Endocrinology 1986, 118, 2149–2151. [Google Scholar] [CrossRef] [PubMed]
- Olden, K.; Pratt, R.M.; Yamada, K.M. Role of carbohydrates in protein secretion and turnover: Effects of tunicamycin on the major cell surface glycoprotein of chick embryo fibroblasts. Cell 1978, 13, 461–473. [Google Scholar] [CrossRef]
- Olden, K.; Pratt, R.M.; Jaworski, C.; Yamada, K.M. Evidence for role of glycoprotein carbohydrates in membrane transport: Specific inhibition by tunicamycin. Proc. Natl. Acad. Sci. USA 1979, 76, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Chung, T.; Youn, H.; Yeom, C.J.; Kang, K.W.; Chung, J.K. Glycosylation of Sodium/Iodide Symporter (NIS) Regulates Its Membrane Translocation and Radioiodine Uptake. PLoS ONE 2015, 10, e0142984. [Google Scholar] [CrossRef] [Green Version]
- Levy, O.; De la Vieja, A.; Ginter, C.S.; Riedel, C.; Dai, G.; Carrasco, N. N-linked glycosylation of the thyroid Na+/I− symporter (NIS). Implications for its secondary structure model. J. Biol. Chem. 1998, 273, 22657–22663. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.Q.; Fan, Z.Q.; Tang, Y.F.; Ke, Z.J. The resveratrol attenuates ethanol-induced hepatocyte apoptosis via inhibiting ER-related caspase-12 activation and PDE activity in vitro. Alcohol. Clin. Exp. Res. 2014, 38, 683–693. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Miao, H.; Zhang, K.; Wolfson, A.; Pennathur, S.; Pipe, S.W.; Kaufman, R.J. Antioxidants reduce endoplasmic reticulum stress and improve protein secretion. Proc. Natl. Acad. Sci. USA 2008, 105, 18525–18530. [Google Scholar] [CrossRef] [Green Version]
- Holthoff, J.H.; Woodling, K.A.; Doerge, D.R.; Burns, S.T.; Hinson, J.A.; Mayeux, P.R. Resveratrol, a dietary polyphenolic phytoalexin, is a functional scavenger of peroxynitrite. Biochem. Pharmacol. 2010, 80, 1260–1265. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.S.; Xia, C.; Jiang, B.H.; Stinefelt, B.; Klandorf, H.; Harris, G.K.; Shi, X. Resveratrol scavenges reactive oxygen species and effects radical-induced cellular responses. Biochem. Biophys. Res. Commun. 2003, 309, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, S.; Holme, A.L. Resveratrol: Its biologic targets and functional activity. Antioxid. Redox Signal. 2009, 11, 2851–2897. [Google Scholar] [CrossRef] [PubMed]
- Palsamy, P.; Subramanian, S. Resveratrol protects diabetic kidney by attenuating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling. Biochim. Biophys. Acta 2011, 1812, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, L.; Zhou, L.; Lei, Y.; Zhang, Y.; Huang, C. Redox signaling and unfolded protein response coordinate cell fate decisions under ER stress. Redox Biol. 2019, 25, 101047. [Google Scholar] [CrossRef]
- Guarente, L.; Franklin, H. Epstein Lecture: Sirtuins, aging, and medicine. N. Engl. J. Med. 2011, 364, 2235–2244. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, Q.; Wang, M.; Liang, M.; Yang, X.; Xu, X.; Zou, H.; Qiu, J. Activation of Sirt1 by resveratrol inhibits TNF-alpha induced inflammation in fibroblasts. PLoS ONE 2011, 6, e27081. [Google Scholar] [CrossRef]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, T.; Gu, J.; Chen, W.; Chang, Q. Resveratrol inhibits high-glucose-induced inflammatory “metabolic memory” in human retinal vascular endothelial cells through SIRT1-dependent signaling. Can. J. Physiol. Pharm. 2019, 97, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Pang, Y.; Wei, W.; Shao, A.; Deng, C.; Li, X.; Chang, H.; Hu, P.; Liu, X.; Zhang, X. Resveratrol protects retinal ganglion cell axons through regulation of the SIRT1-JNK pathway. Exp. Eye Res. 2020, 200, 108249. [Google Scholar] [CrossRef]
- Böttner, M.; Christoffel, J.; Rimoldi, G.; Wuttke, W. Effects of long-term treatment with resveratrol and subcutaneous and oral estradiol administration on the pituitary-thyroid-axis. Exp. Clin. Endocrinol. Diabetes 2006, 114, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva, W.S.; Harney, J.W.; Kim, B.W.; Li, J.; Bianco, S.D.; Crescenzi, A.; Christoffolete, M.A.; Huang, S.A.; Bianco, A.C. The small polyphenolic molecule kaempferol increases cellular energy expenditure and thyroid hormone activation. Diabetes 2007, 56, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Ringseis, R.; Rauer, C.; Rothe, S.; Gessner, D.K.; Schütz, L.M.; Luci, S.; Wen, G.; Eder, K. Sterol regulatory element-binding proteins are regulators of the NIS gene in thyroid cells. Mol. Endocrinol. 2013, 27, 781–800. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.; Ringseis, R.; Koc, A.; Lukas, I.; Kluge, H.; Eder, K. Supplementation with l-carnitine downregulates genes of the ubiquitin proteasome system in the skeletal muscle and liver of piglets. Animal 2012, 6, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiappisi, E.; Ringseis, R.; Eder, K.; Gessner, D.K. Effect of endoplasmic reticulum stress on metabolic and stress signaling and kidney-specific functions in Madin-Darby bovine kidney cells. J. Dairy Sci. 2017, 100, 6689–6706. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.; Eder, K.; Ringseis, R. 1,25-hydroxyvitamin D3 decreases endoplasmic reticulum stress-induced inflammatory response in mammary epithelial cells. PLoS ONE 2020, 15, e0228945. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waltz, F.; Pillette, L.; Ambroise, Y. A nonradioactive iodide uptake assay for sodium iodide symporter function. Anal. Biochem. 2010, 396, 91–95. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (Forward, Reverse) | Product Size (bp) | NCBI GenBank |
|---|---|---|---|
| Reference Genes | |||
| ACTB | GACCTCTATGCCAACACAGT CACCAATCCACACAGAGTAC | 154 | NM_031144 |
| RPL13 | CTTAAATTGGCCACGCAGCT CTTCTCAACGTCTTGCTCTG | 198 | XR_086310 |
| TOP1 | GAAGAACGCTATCCAGAAGG GCTTTGGGACTCAGCTTCAT | 137 | NM_022615 |
| YWHAZ | GACGGAAGGTGCTGAGAAA GCAGCAACCTCAGCCAAGT | 198 | NM_013011 |
| Target Genes | |||
| DDIT3 | ACAAGCACCTCCCAAAGCCC TGCTCCTTCTCCTTCATGCGC | 155 | NM_001109986 |
| HMOX1 | AGCATGTCCCAGGATTTGTC TCACCAGCTTAAAGCCTTCC | 130 | NM_012580 |
| HSAP5 | TCAGCCCACCGTAACAATCAAGG TCCTCAGCAAACTTCTCGGCG | 282 | NM_013083 |
| NIS | GCTGTGGCATTGTCATGTTC TGAGGTCTTCCACAGTCACA | 219 | NM_052983 |
| PAX-8 | CCTTACTCAACAGTACCCTGG AGCTAGAACTGGAGAGCTCTG | 162 | NM_031141 |
| SOD1 | TATGGTGGTCCACGAGAAAC AATCACACCACAAGCCAAGC | 100 | NM_017050 |
| TG | GTTCCTACGTGTACTAGTGAG CATACTGGAGTTGGAGAGCAG | 196 | NM_030988 |
| TPO | CAGGTGTTGAGAAGCAGTTG CTTTGAAAGCTGTAGCCAGG | 255 | NM_019353 |
| TSHR | CCAGAAGCTTGACTTACATAG CATGTAAGGGTTGTCTGTGAT | 161 | NM_012888 |
| TTF-1 | GCATGAATATGAGCGGCATGG ACTTCTGCTGCTTGAAGCGTC | 153 | NM_013093 |
| TTF-2 | GAAGTGGCAGAACAGCATCC AGCTGCCGCTCTCGAACATG | 139 | NM_138909 |
| XBP1 | GACACGCTTGGGGATGAATGC AGAGGCAACAGCGTCAGAATCC | 192/166 | NM_001004210 (XBP1) NM_001271731 (sXBP1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, G.; Eder, K.; Ringseis, R. Resveratrol Alleviates the Inhibitory Effect of Tunicamycin-Induced Endoplasmic Reticulum Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes. Int. J. Mol. Sci. 2021, 22, 4373. https://doi.org/10.3390/ijms22094373
Wen G, Eder K, Ringseis R. Resveratrol Alleviates the Inhibitory Effect of Tunicamycin-Induced Endoplasmic Reticulum Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes. International Journal of Molecular Sciences. 2021; 22(9):4373. https://doi.org/10.3390/ijms22094373
Chicago/Turabian StyleWen, Gaiping, Klaus Eder, and Robert Ringseis. 2021. "Resveratrol Alleviates the Inhibitory Effect of Tunicamycin-Induced Endoplasmic Reticulum Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes" International Journal of Molecular Sciences 22, no. 9: 4373. https://doi.org/10.3390/ijms22094373
APA StyleWen, G., Eder, K., & Ringseis, R. (2021). Resveratrol Alleviates the Inhibitory Effect of Tunicamycin-Induced Endoplasmic Reticulum Stress on Expression of Genes Involved in Thyroid Hormone Synthesis in FRTL-5 Thyrocytes. International Journal of Molecular Sciences, 22(9), 4373. https://doi.org/10.3390/ijms22094373