
Fluoxetine Treatment Decreases Cardiac Vagal Input and Alters the Serotonergic Modulation of the Parasympathetic Outflow in Diabetic Rats

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Systemic Haemodynamic Parameters
2.2. Alterations in the HR by Electrical Vagal Stimulation or Exogenous ACh
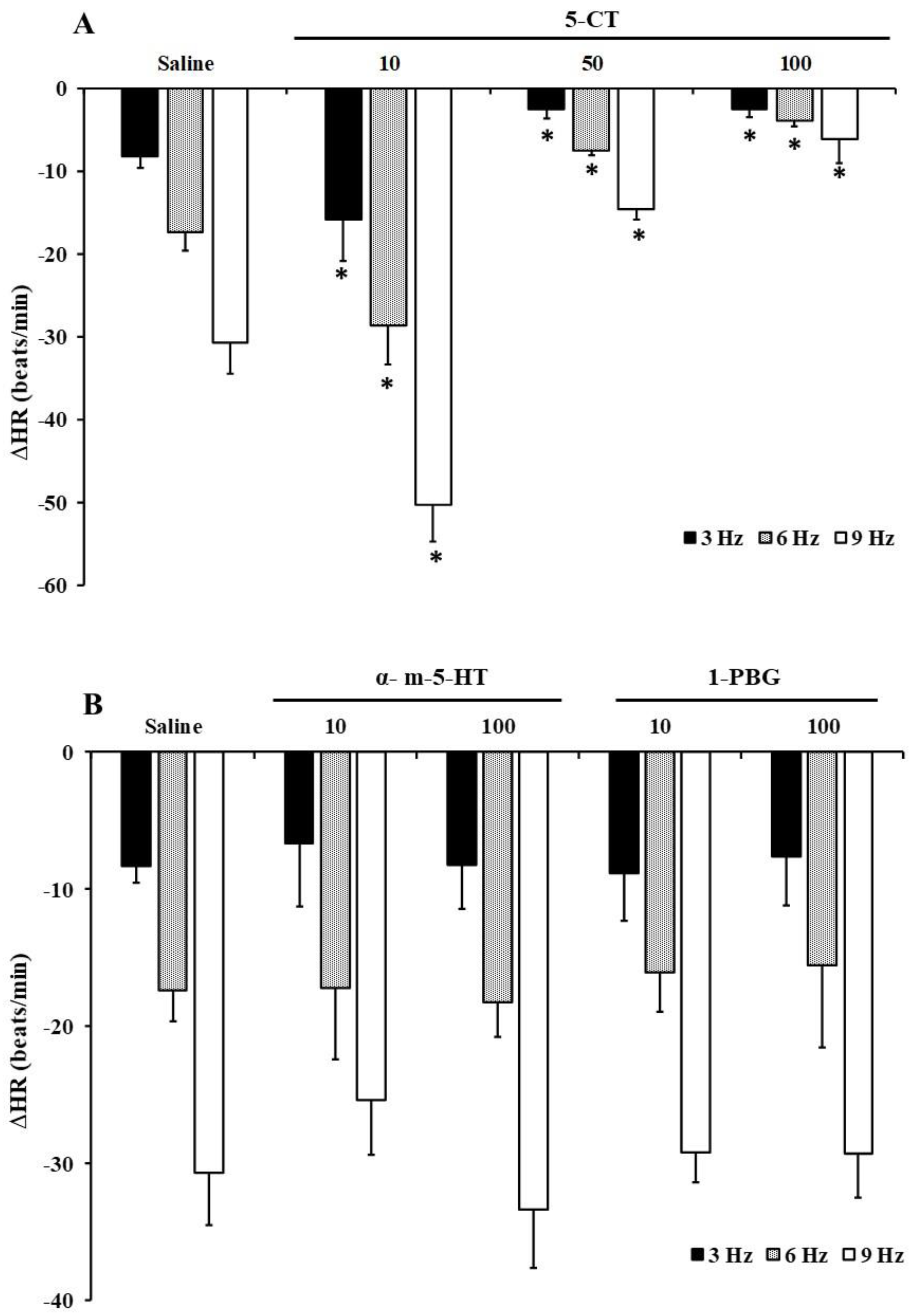
2.3. Effect of 5-HT on Electrically-Induced Bradycardia in FT Diabetic Rats
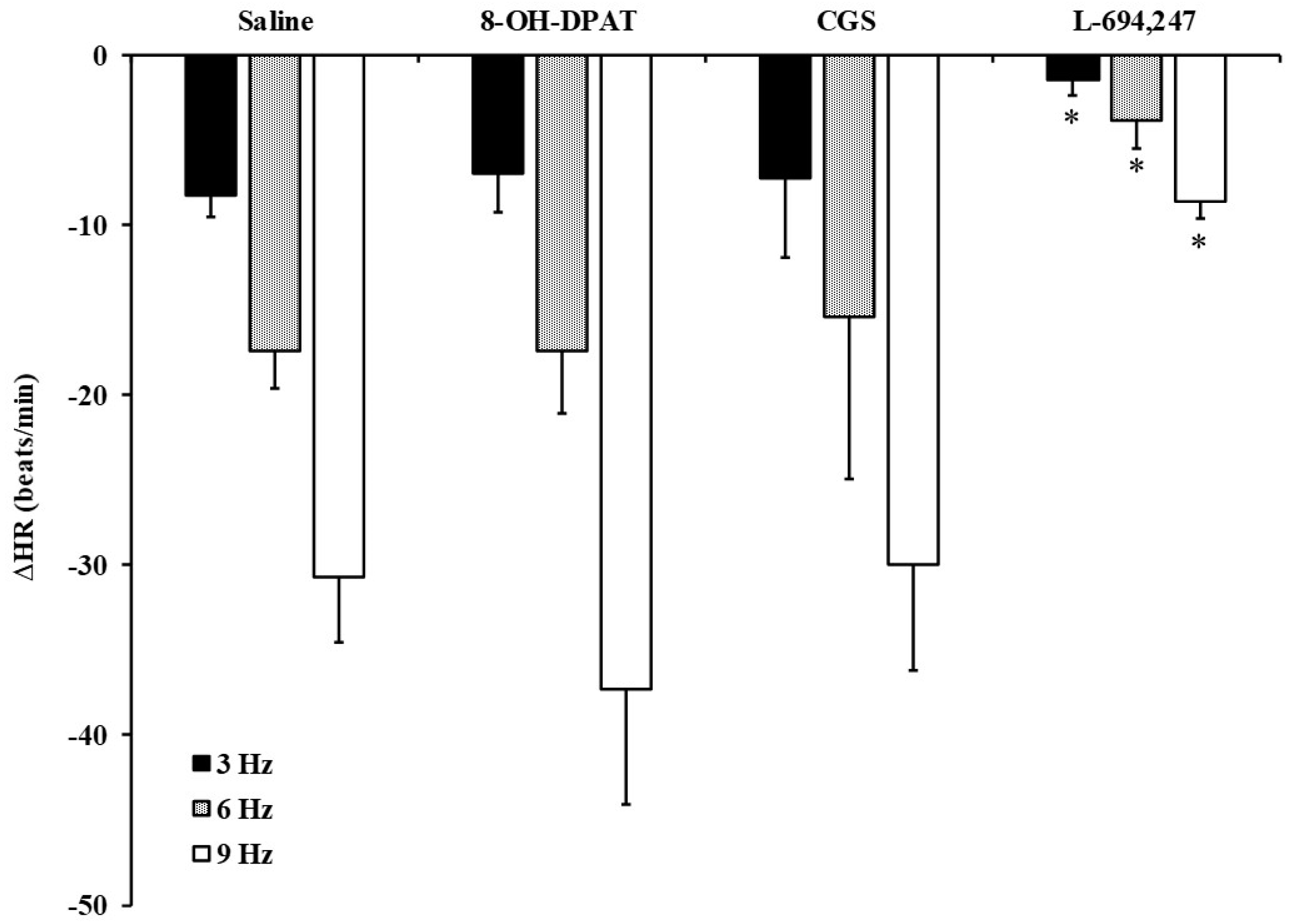
2.4. Role of 5-HT Receptor Serotonergic Agonists on the Vagally Induced Bradycardia in FT Diabetic Rats
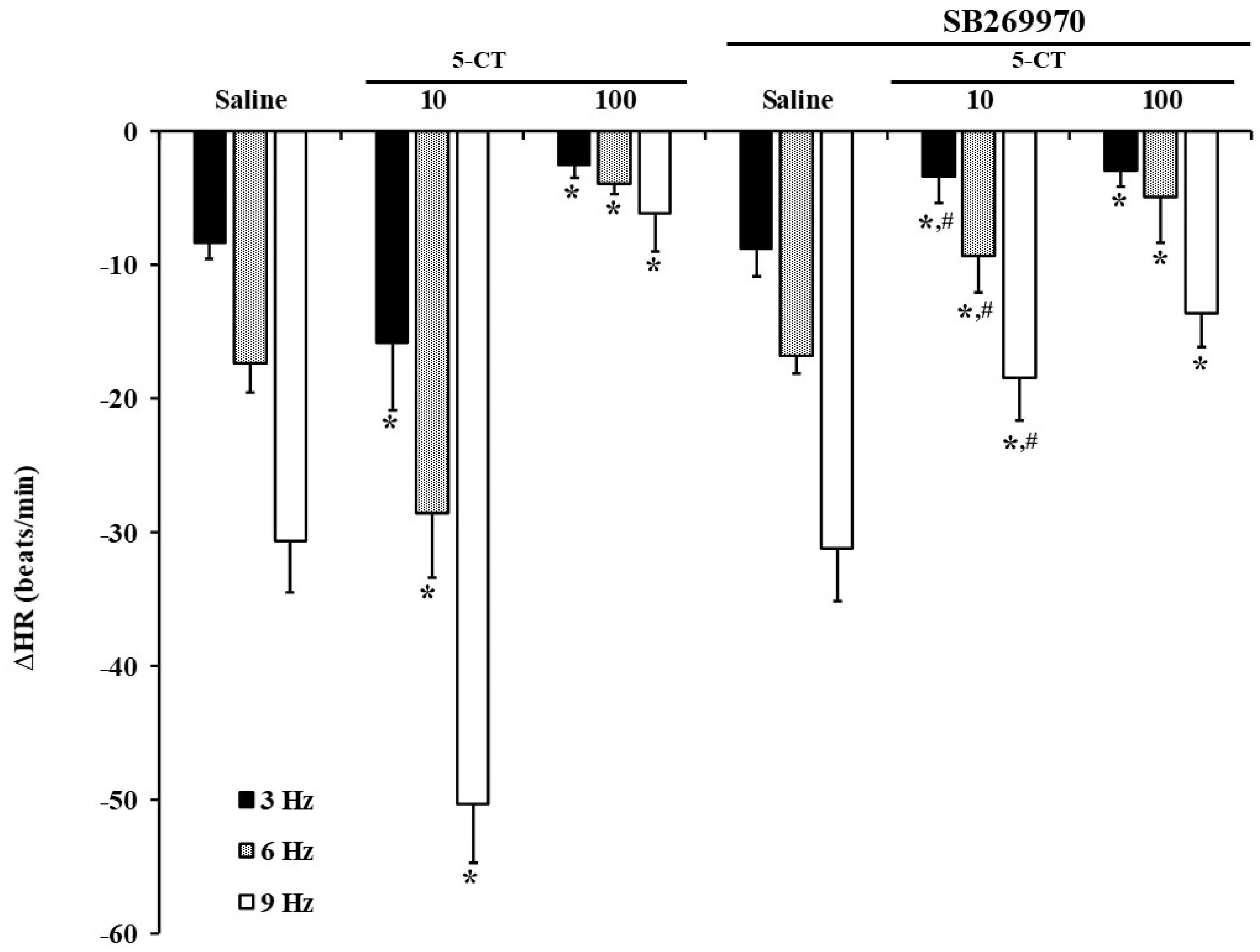
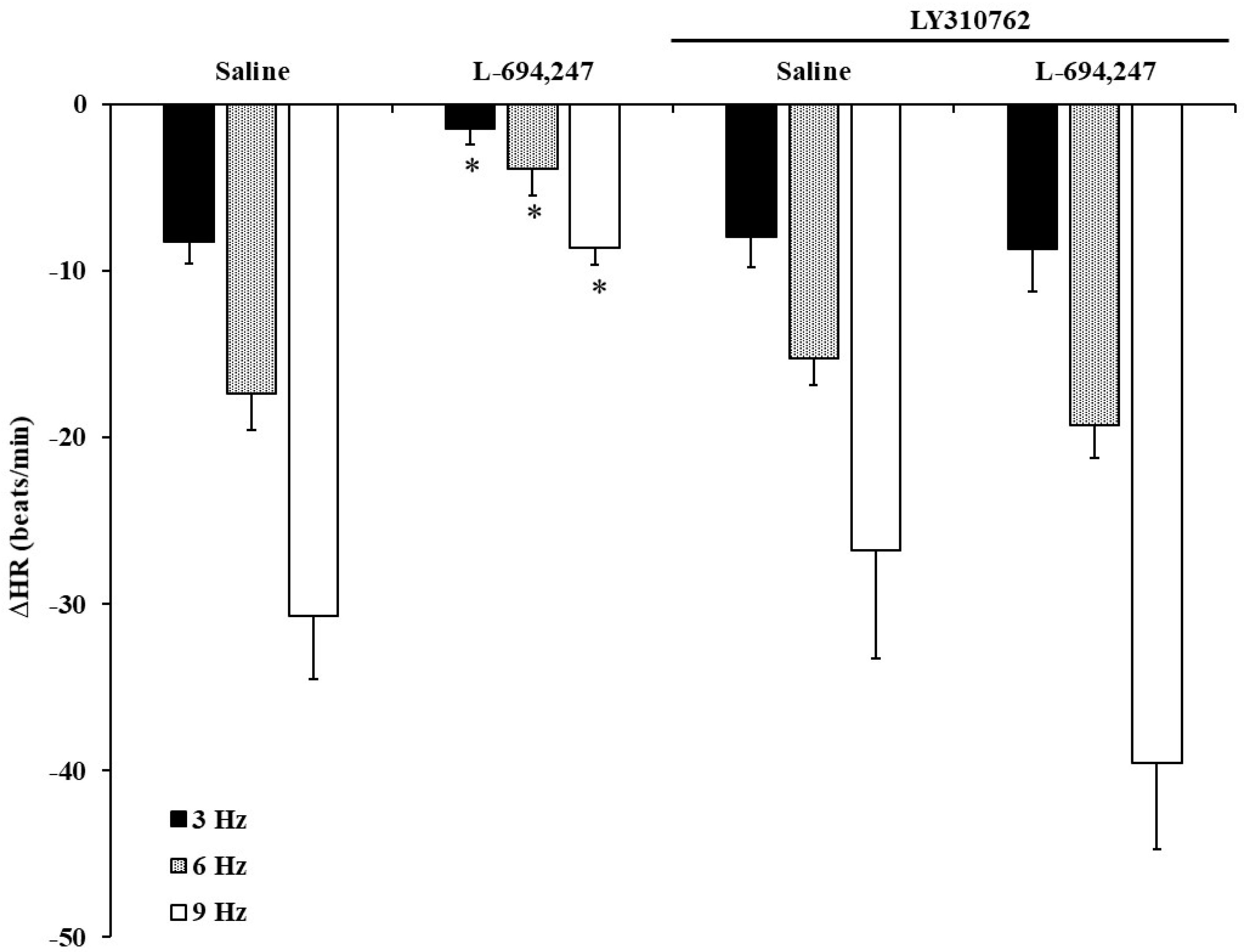
2.5. Influence of Saline, SB269970 or LY310762 on the 5-CT or L-694,247 Effect on the Electrical-Induced Bradycardic Responses in FT Diabetic Rats
2.6. 5-CT Action on the Bradycardia Originated by Administration of Exogenous ACh (i.v.)
3. Discussion
4. Materials and Methods
4.1. Drugs Used
4.2. General Methods
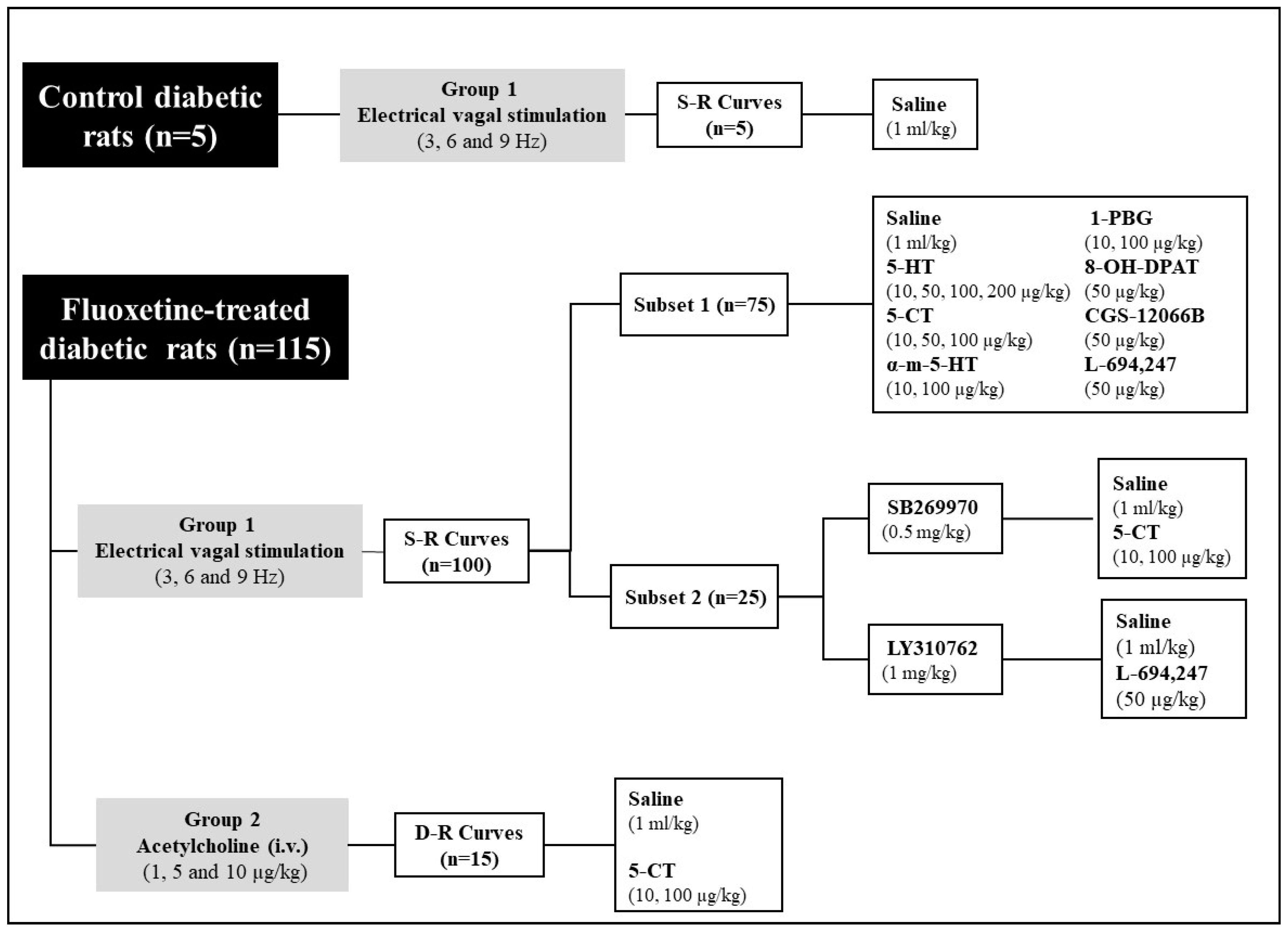
4.3. Experimental Design
4.3.1. Protocol I: Vagus Nerve Electrical Stimulation
4.3.2. Protocol II: Intravenous Administration of Exogenous ACh
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steenblock, C.; Schwarz, P.E.H.; Perakakis, N.; Brajshori, N.; Beqiri, P.; Bornstein, S.R. The interface of COVID-19, diabetes, and depression. Discov. Ment. Health 2022, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Herrera, P.A.; Campos-Romero, S.; Szabo, W.; Martinez, P.; Guajardo, V.; Rojas, G. Understanding the relationship between depression and chronic diseases such as diabetes and hypertension: A grounded theory study. Int. J. Environ. Res. Public Health 2021, 18, 12130. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.I.; de Groot, M.; Golden, S.H. Diabetes and depression. Curr. Diabetes Rep. 2014, 14, 491. [Google Scholar] [CrossRef] [Green Version]
- Maser, R.E.; Mitchell, B.D.; Vinik, A.I.; Freeman, R. The association between cardiovascular autonomic neuropathy and mortality in individuals with diabetes: A meta-analysis. Diabetes Care 2003, 26, 1895–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinik, A.I.; Ziegler, D. Diabetic cardiovascular autonomic neuropathy. Circulation 2007, 115, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Duque, A.; Mediano, M.F.F.; De Lorenzo, A.; Rodrigues Jr, L.F. Cardiovascular autonomic neuropathy in diabetes: Pathophysiology, clinical assessment and implications. World J. Diabetes 2021, 12, 855–867. [Google Scholar] [CrossRef]
- Park, H.J.; Zhang, Y.; Du, C.; Welzig, C.M.; Madias, C.; Aronovitz, M.J.; Georgescu, S.P.; Naggar, I.; Wang, B.; Kim, Y.B.; et al. Role of SREBP-1 in the development of parasympathetic dysfunction in the hearts of type 1 diabetic Akita mice. Circ. Res. 2009, 105, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Krishnaswamy, P.S.; Egom, E.E.; Moghtadaei, M.; Jansen, H.J.; Azer, J.; Bogachev, O.; Mackasey, M.; Robbins, C.; Rose, R.A. Altered parasympathetic nervous system regulation of the sinoatrial node in Akita diabetic mice. J. Mol. Cell. Cardiol. 2015, 82, 125–135. [Google Scholar] [CrossRef]
- Balcıoğlu, A.S.; Müderrisoğlu, H. Diabetes and cardiac autonomic neuropathy: Clinical manifestations, cardiovascular consequences, diagnosis and treatment. World J. Diabetes 2015, 6, 80–91. [Google Scholar] [CrossRef]
- Castillo-Quan, J.I.; Barrera-Buenfil, D.J.; Pérez-Osorio, J.M.; Alvarez-Cervera, F.J. Depression and diabetes: From epidemiology to neurobiology. Rev. Neurol. 2010, 51, 347–359. [Google Scholar]
- Rossi, A.; Barraco, A.; Donda, P. Fluoxetine: A review on evidence based medicine. Ann. Gen. Hosp. Psychiatry 2004, 3, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, D.A.; Dawood, T.; Lambert, E.A.; Esler, M.D.; Haikerwal, D.; Brenchley, C.; Socratous, F.; Kaye, D.M.; Schlaich, M.P.; Hickie, I.; et al. Sympathetic activity in major depressive disorder: Identifying those at increased cardiac risk? J. Hypertens. 2007, 25, 2117–2124. [Google Scholar] [CrossRef] [PubMed]
- Dawood, T.; Lambert, E.A.; Barton, D.A.; Laude, D.; Elghozi, J.L.; Esler, M.D.; Haikerwal, D.; Kaye, D.M.; Hotchkin, E.J.; Lambert, G.W. Specific serotonin reuptake inhibition in major depressive disorder adversely affects novel markers of cardiac risk. Hypertens. Res. 2007, 30, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, C.A.; Rodrigues, F.L.; Ruginsk, S.G.; Zanotto, C.Z.; Rodrigues, J.A.; Duarte, D.A.; Costa-Neto, C.M.; Resstel, L.B.; Carneiro, F.S.; Tostes, R.C. Chronic treatment with fluoxetine modulates vascular adrenergic responses by inhibition of pre- and post-synaptic mechanisms. Eur. J. Pharmacol. 2017, 800, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, V.J.; Ertl, A.C.; Tate, D.B.; Davis, S.N. Effects of the selective serotonin reuptake inhibitor fluoxetine on counterregulatory responses to hypoglycemia in individuals with type 1 diabetes. Diabetes 2008, 57, 3315–3322. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Lopez, A.; Centurión, D.; Vázquez, E.; Arulmani, U.; Saxena, P.R.; Villalón, C.M. Pharmacological profile of the 5-HT-induced inhibition of cardioaccelerator sympathetic outflow in pithed rats: Correlation with 5-HT1 and putative 5-ht5A/5B receptors. Br. J. Pharmacol. 2003, 140, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-López, A.; Centurión, D.; Vázquez, E.; Arulmani, U.; Saxena, P.R.; Villalón, C.M. Further characterization of the 5-HT1 receptors mediating cardiac sympatho-inhibition in pithed rats: Pharmacological correlation with the 5-HT1B and 5-HT1D subtypes. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2004, 369, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Morán, A.; Velasco, C.; Martín, M.L.; San Román, L. Pharmacological characterization of 5-HT receptors in parasympathetic innervation of rat heart. Eur. J. Pharmacol. 1994, 252, 161–166. [Google Scholar] [CrossRef]
- García, M.; Morán, A.; Martín, M.L.; Ortiz de Urbina, A.V.; San Román, L. Diabetes-induced changes in 5-hydroxytryptamine modulation of vagally-induced bradycardia in rat heart. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1199–1206. [Google Scholar] [CrossRef]
- Restrepo, B.; Martín, M.L.; San Román, L.; Morán, A. Peripheral 5-HT1A and 5-HT7 serotonergic receptors modulate parasympathetic neurotransmission in long-term diabetic rats. Exp. Diabetes Res. 2010, 2010, 686734. [Google Scholar] [CrossRef] [Green Version]
- García-Pedraza, J.Á.; García, M.; Martín, M.L.; Eleno, N.; Morán, A. Chronic Sarpogrelate Treatment Reveals 5-HT7 Receptor in the Serotonergic Inhibition of the Rat Vagal Bradycardia. J. Cardiovasc. Pharmacol. 2017, 69, 13–22. [Google Scholar] [CrossRef] [PubMed]
- García-Pedraza, J.Á.; Hernández-Abreu, O.; García, M.; Morán, A.; Villalón, C.M. Chronic 5-HT(2) receptor blockade unmasks the role of 5-HT(1F) receptors in the inhibition of rat cardioaccelerator sympathetic outflow. Can. J. Physiol. Pharmacol. 2018, 96, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Pedraza, J.Á.; Hernández-Abreu, O.; Morán, A.; Carretero, J.; García-Domingo, M.; Villalón, C.M. Role of peripheral 5-HT(5A) receptors in 5-HT-induced cardiac sympatho-inhibition in type 1 diabetic rats. Sci. Rep. 2020, 10, 19358. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Abreu, O.I.; García-Pedraza, J.Á.; Rivera-Mancilla, E.; Villanueva-Castillo, B.; Morán, A.; García-Domingo, M.; Manrique-Maldonado, G.; Altamirano-Espinoza, A.H.; Villalón, C.M. Blockade of 5-HT(2) receptors with sarpogrelate uncovers 5-HT(7) receptors inhibiting the tachycardic sympathetic drive in pithed rats. Clin. Exp. Pharmacol. Physiol. 2020, 47, 403–411. [Google Scholar] [CrossRef]
- López, C.; Gómez-Roso, M.; García-Pedraza, J.Á.; Martín, M.L.; Morán, A.; García-Domingo, M. Fluoxetine oral treatment discloses 5-HT(1D) receptor as vagoinhibitor of the cardiac cholinergic neurotransmission in rat. Can. J. Physiol. Pharmacol. 2019, 97, 90–98. [Google Scholar] [CrossRef]
- Vinik, A.I.; Maser, R.E.; Mitchell, B.D.; Freeman, R. Diabetic autonomic neuropathy. Diabetes Care 2003, 26, 1553–1579. [Google Scholar] [CrossRef] [Green Version]
- García-Pedraza, J.Á.; Fernández-González, J.F.; López, C.; Martín, M.L.; Alarcón-Torrecillas, C.; Rodríguez-Barbero, A.; Morán, A.; García-Domingo, M. Oral fluoxetine treatment changes serotonergic sympatho-regulation in experimental type 1 diabetes. Life Sci. 2022, 293, 120335. [Google Scholar] [CrossRef]
- Mondol, D.; Islam, M.N.; Biswas, S.; Jodder, P.; Sana, S.; Saleh, M.A.; Islam, M.R. Investigation of the synergistic effect of glimepiride and rosuvastatin on alloxan-induced diabetic rat. J. Diabetes Metab. Disord. 2020, 19, 1415–1422. [Google Scholar] [CrossRef]
- Hong, L.Z.; Huang, K.F.; Hung, S.W.; Kuo, L.T. Chronic fluoxetine treatment enhances sympathetic activities associated with abnormality of baroreflex function in conscious normal rats. Eur. J. Pharmacol. 2017, 811, 164–170. [Google Scholar] [CrossRef]
- García-Pedraza, J.Á.; López, C.; Fernández-González, J.F.; Martín, M.L.; Morán, A.; García-Domingo, M. Vascular sympathetic neurotransmission and its serotonergic regulation are modified by chronic fluoxetine treatment. J. Pharmacol. Sci. 2021, 147, 48–57. [Google Scholar] [CrossRef]
- Able, J.A.; Liu, Y.; Jandacek, R.; Rider, T.; Tso, P.; McNamara, R.K. Omega-3 fatty acid deficient male rats exhibit abnormal behavioral activation in the forced swim test following chronic fluoxetine treatment: Association with altered 5-HT1A and alpha2A adrenergic receptor expression. J. Psychiatr. Res. 2014, 50, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Amsterdam, J.D.; Fawcett, J.; Quitkin, F.M.; Reimherr, F.W.; Rosenbaum, J.F.; Michelson, D.; Hornig-Rohan, M.; Beasley, C.M. Fluoxetine and norfluoxetine plasma concentrations in major depression: A multicenter study. Am. J. Phsychiatry 1997, 154, 963–969. [Google Scholar] [CrossRef] [Green Version]
- Moosa, M.Y.; Panz, V.R.; Jeenah, F.Y.; Joffe, B.I. African women with depression: The effect of imipramine and fluoxetine on body mass index and leptin secretion. J. Clin. Psychopharmacol. 2003, 23, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Ghaeli, P.; Shahsavand, E.; Mesbahi, M.; Kamkar, M.Z.; Sadeghi, M.; Dashti-Khavidaki, S. Comparing the effects of 8-week treatment with fluoxetine and imipramine on fasting blood glucose of patients with major depressive disorder. J. Clin. Psychopharmacol. 2004, 24, 386–388. [Google Scholar] [CrossRef] [PubMed]
- Pop-Busui, R. Cardiac autonomic neuropathy in diabetes: A clinical perspective. Diabetes Care 2010, 33, 434–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, C.; Jessen, N.; Brock, B.; Jakobsen, P.E.; Hansen, T.K.; Rantanen, J.M.; Riahi, S.; Dimitrova, Y.K.; Dons-Jensen, A.; Aziz, Q.; et al. Cardiac vagal tone, a non-invasive measure of parasympathetic tone, is a clinically relevant tool in Type 1 diabetes mellitus. Diabet. Med. 2017, 34, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Monti, J.M.; Jantos, H. The role of serotonin 5-HT7 receptor in regulating sleep and wakefulness. Rev. Neurosci. 2014, 25, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Boehm, S.; Kubista, H. Fine tuning of sympathetic transmitter release via ionotropic and metabotropic presynaptic receptors. Pharmacol. Rev. 2002, 54, 43–99. [Google Scholar] [CrossRef]
- Soslau, G. Cardiovascular serotonergic system: Evolution, receptors, transporter, and function. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2022, 337, 115–127. [Google Scholar] [CrossRef]
- Vanhoutte, P.M.; Zhao, Y.; Xu, A.; Leung, S.W. Thirty Years of Saying NO: Sources, Fate, Actions, and Misfortunes of the Endothelium-Derived Vasodilator Mediator. Circ. Res. 2016, 119, 375–396. [Google Scholar] [CrossRef] [Green Version]
- Mathur, C.; Gupta, D. QTc prolongation in diabetes mellitus-An indicator of cardiac autonomic neuropathy. J. Indian Acad. Clin. Med. 2006, 7, 130–132. [Google Scholar]
- Gorodeski, E.Z.; Ishwaran, H.; Blackstone, E.H.; Lauer, M.S. Quantitative electrocardiographic measures and long-term mortality in exercise test patients with clinically normal resting electrocardiograms. Am. Heart J. 2009, 158, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, C.R.; Salles, G.F.; Deccache, W. QTc interval prolongation is a predictor of future strokes in patients with type 2 diabetes mellitus. Stroke 2003, 34, 2187–2194. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.K.; Gall, M.A.; Major-Pedersen, A.; Sato, A.; Rossing, P.; Breum, L.; Pietersen, A.; Kastrup, J.; Parving, H.H. QTc interval length and QT dispersion as predictors of mortality in patients with non-insulin-dependent diabetes. Scand. J. Clin. Lab. Investig. 2000, 60, 323–332. [Google Scholar] [CrossRef]
- Chu, A.; Wadhwa, R. Selective Serotonin Reuptake Inhibitors. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5-HT Receptor Agonists (μg/kg) | ΔMBP (mm Hg) | ΔHR (beats/min) |
|---|---|---|
| 5-HT (10) | 39.2 ± 2.6 * | 7.3 ± 2.4 |
| 5-HT (100) | 76.1 ± 4.3 * | 6.7 ± 1.2 |
| 5-HT (200) | 89.5 ± 1.2 * | 2.1 ± 0.6 |
| 5-CT (10) | −2.1 ± 0.6 | 3.4 ± 1.7 |
| 5-CT (100) | −6.5 ± 1.7 | 4.6 ± 1.5 |
| α- m-5-HT (10) | 42.2 ± 7.1 * | 3.7 ± 0.8 |
| α- m-5-HT (100) | 86.4 ± 5.6 * | 5.4 ± 2.1 |
| 1-PBG (10) | 3.2 ± 0.9 | 3.2 ± 2.4 |
| 1-PBG (100) | 6.1 ± 1.7 | 9.3 ± 4.9 |
| 5-HT (μg/kg) | Vagal Stimulation Frequencies | ||
|---|---|---|---|
| 3 Hz | 6 Hz | 9 Hz | |
| Saline | −8.3 ± 1.3 | −17.4 ± 2.2 | −30.7 ± 3.8 |
| 10 | −16.0 ± 2.0 * | −30.6 ± 6.2 * | −60.2 ± 7.5 * |
| 50 | −10.8 ± 8.5 | −23.6 ± 8.2 | −47.4 ± 8.8 * |
| 100 | −5.1 ± 1.2 * | −9.3 ± 1.7 * | −20.6 ± 3.2 * |
| 200 | −3.3 ± 1.1 * | −9.4 ± 3.1 * | −13.8 ± 1.0 * |
| ΔHR (beats/min) | |||
| ACh (μg/kg) | Saline (ml/kg) | 5-CT (μg/kg) | |
|---|---|---|---|
| 1 | 10 | 100 | |
| 1 | −38.2 ± 3.3 | −41.6 ± 5.7 | −42.2 ± 2.1 |
| 5 | −47.9 ± 6.1 | −50.1 ± 5.4 | −56.2 ± 4.1 |
| 10 | −72.2 ± 4.7 | −69.8 ± 6.3 | −68.1 ± 3.9 |
| ΔHR (beats/min) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Domingo, M.; García-Pedraza, J.Á.; Fernández-González, J.F.; López, C.; Martín, M.L.; Morán, A. Fluoxetine Treatment Decreases Cardiac Vagal Input and Alters the Serotonergic Modulation of the Parasympathetic Outflow in Diabetic Rats. Int. J. Mol. Sci. 2022, 23, 5736. https://doi.org/10.3390/ijms23105736
García-Domingo M, García-Pedraza JÁ, Fernández-González JF, López C, Martín ML, Morán A. Fluoxetine Treatment Decreases Cardiac Vagal Input and Alters the Serotonergic Modulation of the Parasympathetic Outflow in Diabetic Rats. International Journal of Molecular Sciences. 2022; 23(10):5736. https://doi.org/10.3390/ijms23105736
Chicago/Turabian StyleGarcía-Domingo, Mónica, José Ángel García-Pedraza, Juan Francisco Fernández-González, Cristina López, María Luisa Martín, and Asunción Morán. 2022. "Fluoxetine Treatment Decreases Cardiac Vagal Input and Alters the Serotonergic Modulation of the Parasympathetic Outflow in Diabetic Rats" International Journal of Molecular Sciences 23, no. 10: 5736. https://doi.org/10.3390/ijms23105736
APA StyleGarcía-Domingo, M., García-Pedraza, J. Á., Fernández-González, J. F., López, C., Martín, M. L., & Morán, A. (2022). Fluoxetine Treatment Decreases Cardiac Vagal Input and Alters the Serotonergic Modulation of the Parasympathetic Outflow in Diabetic Rats. International Journal of Molecular Sciences, 23(10), 5736. https://doi.org/10.3390/ijms23105736