
The Relevance of Physico-Chemical Properties and Protein Corona for Evaluation of Nanoparticles Immunotoxicity—In Vitro Correlation Analysis on THP-1 Macrophages

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Characterization
2.2. Impurities
2.3. NP Corona Protein Composition
2.4. Viability
2.5. Nanoparticle Internalization
2.6. Reactive Oxygen Species
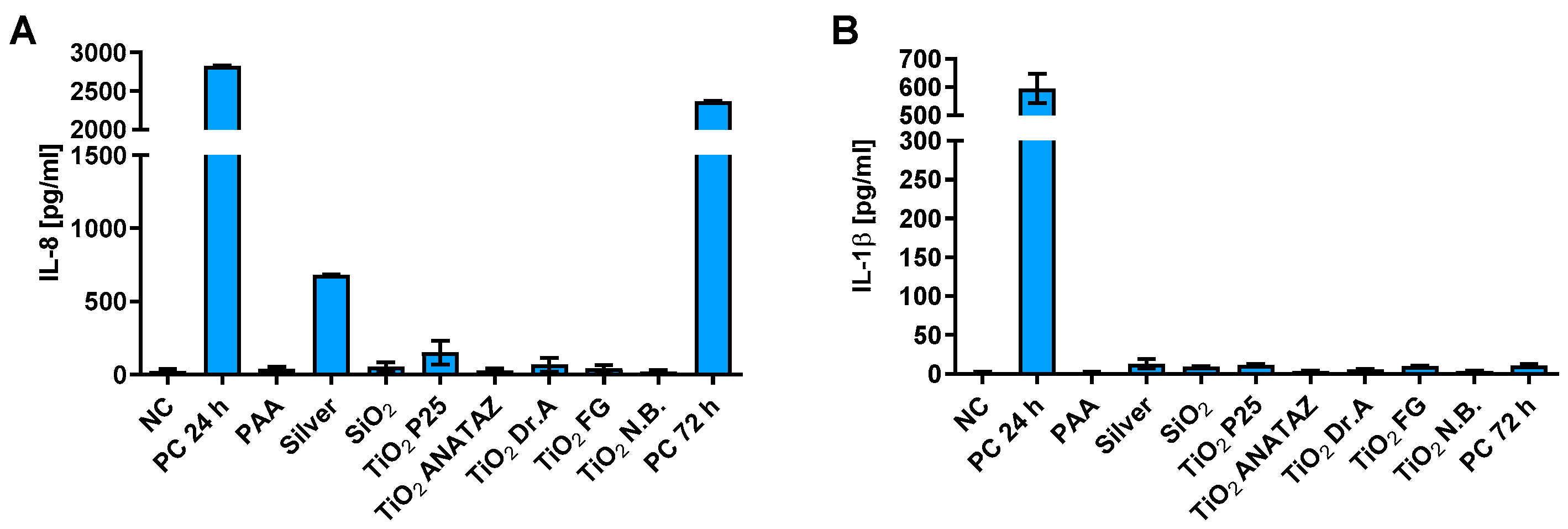
2.7. Cytokine Secretion
2.8. IL-1β Secretion and NLRP3 Inflammasome Activation
2.9. Resolution of Cytokine Secretion
3. Discussion
4. Materials and Methods
4.1. Nanoparticles
4.2. Nanoparticle Characterization
4.3. ICP-MS Analysis of Ions Leaching from NPs
4.4. NP Corona Preparation
4.5. Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Mass Spectrometry (MS)
4.6. Cell Culture
4.7. Viability
4.8. ROS Assay
4.9. Transmission Electron Microscopy
4.10. ELISA
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaudhry, Q.; Scotter, M.; Blackburn, J.; Ross, B.; Boxall, A.; Castle, L.; Aitken, R.; Watkins, R. Applications and implications of nanotechnologies for the food sector. Food Addit. Contam. Part A 2008, 25, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Lorenzetti, M.; Drame, A.; Vidmar, J.; Ščančar, J.; Filipič, M. Diversity of TiO2 nanopowders’ characteristics relevant to toxicity testing. J. Nanopart. Res. 2016, 18, 130. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources Added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Re-evaluation of silicon dioxide (E 551) as a food additive. EFSA J. 2018, 16, e05088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR). Risk Assessment of Products of Nanotechnologies; Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR): Brussels, Belgium, 2009. [Google Scholar]
- Novak, S.; Drobne, D.; Menard, A. Prolonged feeding of terrestrial isopod (Porcellio scaber, Isopoda, Crustacea) on TiO2 nanoparicles. Absence of toxic effect. ZooKeys 2012, 176, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Carregal-Romero, S.; Casula, M.F.; Gutiérrez, L.; Morales, M.P.; Böhm, I.B.; Heverhagen, J.T.; Prosperi, D.; Parak, W.J. Biological applications of magnetic nanoparticles. Chem. Soc. Rev. 2012, 41, 4306–4334. [Google Scholar] [CrossRef]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [Green Version]
- Bachler, G.; von Goetz, N.; Hungerbuhler, K. Using physiologically based pharmacokinetic (PBPK) modeling for dietary risk assessment of titanium dioxide (TiO2) nanoparticles. Nanotoxicology 2015, 9, 373–380. [Google Scholar] [CrossRef]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; von Goetz, N. Titanium Dioxide Nanoparticles in Food and Personal Care Products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Re-evaluation of titanium dioxide (E 171) as a food additive. EFSA J. 2016, 14, e04545. [Google Scholar] [CrossRef]
- Heller, A.; Jarvis, K.; Coffman, S.S. Association of Type 2 Diabetes with Submicron Titanium Dioxide Crystals in the Pancreas. Chem. Res. Toxicol. 2018, 31, 506–509. [Google Scholar] [CrossRef] [PubMed]
- Milosevic, A.; Romeo, D.; Wick, P. Understanding Nanomaterial Biotransformation: An Unmet Challenge to Achieving Predictive Nanotoxicology. Small 2020, 16, 1907650. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-J.; Park, K. Oxidative stress and pro-inflammatory responses induced by silica nanoparticles in vivo and in vitro. Toxicol. Lett. 2009, 184, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Hunter, A.C.; Dadswell, C.M.; Savay, S.; Alving, C.R.; Szebeni, J. Causative factors behind poloxamer 188 (Pluronic F68, FlocorTM)-induced complement activation in human sera: A protective role against poloxamer-mediated complement activation by elevated serum lipoprotein levels. Biochim. Biophys. Acta BBA-Mol. Basis Dis. 2004, 1689, 103–113. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Andersen, A.J.; Ahmadvand, D.; Wibroe, P.P.; Andresen, T.L.; Hunter, A.C. Material properties in complement activation. Adv. Drug Deliv. Rev. 2011, 63, 1000–1007. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Andersen, A.; Hashemi, S.; Lettiero, B.; Ahmadvand, D.; Hunter, A.; Andresen, T.L.; Hamad, I.; Szebeni, J. Complement activation cascade triggered by PEG–PL engineered nanomedicines and carbon nanotubes: The challenges ahead. J. Control. Release 2010, 146, 175–181. [Google Scholar] [CrossRef]
- Hsiao, J.-K.; Chu, H.-H.; Wang, Y.-H.; Lai, C.-W.; Chou, P.-T.; Hsieh, S.-T.; Wang, J.-L.; Liu, H.-M. Macrophage physiological function after superparamagnetic iron oxide labeling. NMR Biomed. 2008, 21, 820–829. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, L.; Bai, R.; Zhang, T.; Chen, C. Silver nanoparticles impede phorbol myristate acetate-induced monocyte–macrophage differentiation and autophagy. Nanoscale 2015, 7, 16100–16109. [Google Scholar] [CrossRef]
- Giovanni, M.; Yue, J.; Zhang, L.; Xie, J.; Ong, C.N.; Leong, D.T. Pro-inflammatory responses of RAW264.7 macrophages when treated with ultralow concentrations of silver, titanium dioxide, and zinc oxide nanoparticles. J. Hazard. Mater. 2015, 297, 146–152. [Google Scholar] [CrossRef]
- Kodali, V.; Littke, M.H.; Tilton, S.C.; Teeguarden, J.; Shi, L.; Frevert, C.W.; Wang, W.; Pounds, J.; Thrall, B.D. Dysregulation of Macrophage Activation Profiles by Engineered Nanoparticles. ACS Nano 2013, 7, 6997–7010. [Google Scholar] [CrossRef]
- Shin, S.-H.; Ye, M.-K.; Kim, H.-S.; Kang, H.-S. The effects of nano-silver on the proliferation and cytokine expression by peripheral blood mononuclear cells. Int. Immunopharmacol. 2007, 7, 1813–1818. [Google Scholar] [CrossRef]
- Mrakovcic, M.; Meindl, C.; Roblegg, E.; Fröhlich, E. Reaction of monocytes to polystyrene and silica nanoparticles in short-term and long-term exposures. Toxicol. Res. 2014, 3, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Banda, N.K.; Mehta, G.; Chao, Y.; Wang, G.; Inturi, S.; Fossati-Jimack, L.; Botto, M.; Wu, L.; Moghimi, S.M.; Simberg, D. Mechanisms of complement activation by dextran-coated superparamagnetic iron oxide (SPIO) nanoworms in mouse versus human serum. Part. Fibre Toxicol. 2014, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.H.; Van Der Ven, L.T.; Sleijffers, A.; Park, M.; Jansen, E.H.; Van Loveren, H.; Vandebriel, R. Systemic and immunotoxicity of silver nanoparticles in an intravenous 28 days repeated dose toxicity study in rats. Biomaterials 2013, 34, 8333–8343. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.-H.; Jang, J.; Kim, S.; Kang, T.; Lee, K.; Choi, I.-H. The effects of sub-lethal concentrations of silver nanoparticles on inflammatory and stress genes in human macrophages using cDNA microarray analysis. Biomaterials 2012, 33, 4690–4699. [Google Scholar] [CrossRef]
- Kermanizadeh, A.; Chauché, C.; Balharry, D.; Brown, D.M.; Kanase, N.; Boczkowski, J.; Lanone, S.; Stone, V. The role of Kupffer cells in the hepatic response to silver nanoparticles. Nanotoxicology 2013, 8, 149–154. [Google Scholar] [CrossRef]
- Andersen, A.J.; Windschiegl, B.; Ilbasmis-Tamer, S.; Degim, I.T.; Hunter, A.C.; Andresen, T.L.; Moghimi, S.M. Complement activation by PEG-functionalized multi-walled carbon nanotubes is independent of PEG molecular mass and surface density. Nanomed. Nanotechnol. Biol. Med. 2013, 9, 469–473. [Google Scholar] [CrossRef]
- Elsabahy, M.; Wooley, K.L. Cytokines as biomarkers of nanoparticle immunotoxicity. Chem. Soc. Rev. 2013, 42, 5552–5576. [Google Scholar] [CrossRef] [Green Version]
- Peeters, P.M.; Eurlings, I.M.J.; Perkins, T.N.; Wouters, E.F.; Schins, R.P.F.; Borm, P.J.A.; Drommer, W.; Reynaert, N.L.; Albrecht, C. Silica-induced NLRP3 inflammasome activation in vitro and in rat lungs. Part. Fibre Toxicol. 2014, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-G.; Lee, P.-H.; Lee, S.-H.; Park, M.-K.; Jang, A.-S. Effect of TiO2Nanoparticles on Inflammasome-Mediated Airway Inflammation and Responsiveness. Allergy Asthma Immunol. Res. 2017, 9, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Gómez, D.M.; Urcuqui-Inchima, S.; Hernandez, J.C. Silica nanoparticles induce NLRP3 inflammasome activation in human primary immune cells. Innate Immun. 2017, 23, 697–708. [Google Scholar] [CrossRef]
- Sun, B.; Wang, X.; Ji, Z.; Li, R.; Xia, T. NLRP3 Inflammasome Activation Induced by Engineered Nanomaterials. Small 2012, 9, 1595–1607. [Google Scholar] [CrossRef]
- Mishra, A.R.; Zheng, J.; Tang, X.; Goering, P.L. Silver Nanoparticle-Induced Autophagic-Lysosomal Disruption and NLRP3-Inflammasome Activation in HepG2 Cells Is Size-Dependent. Toxicol. Sci. 2016, 150, 473–487. [Google Scholar] [CrossRef]
- Borm, P.J.A.; Tran, L.; Donaldson, K. The carcinogenic action of crystalline silica: A review of the evidence supporting secondary inflammation-driven genotoxicity as a principal mechanism. Crit. Rev. Toxicol. 2011, 41, 756–770. [Google Scholar] [CrossRef]
- Shirasuna, K.; Karasawa, T.; Takahashi, M. Exogenous nanoparticles and endogenous crystalline molecules as danger signals for the NLRP3 inflammasomes. J. Cell. Physiol. 2018, 234, 5436–5450. [Google Scholar] [CrossRef]
- Mech, A.; Wohlleben, W.; Ghanem, A.; Hodoroaba, V.; Weigel, S.; Babick, F.; Brüngel, R.; Friedrich, C.M.; Rasmussen, K.; Rauscher, H. Nano or Not Nano? A Structured Approach for Identifying Nanomaterials According to the European Commission’s Definition. Small 2020, 16, e2002228. [Google Scholar] [CrossRef]
- Crist, R.; Grossman, J.H.; Patri, A.K.; Stern, S.T.; Dobrovolskaia, M.A.; Adiseshaiah, P.P.; Clogston, J.; McNeil, S.E. Common pitfalls in nanotechnology: Lessons learned from NCI’s Nanotechnology Characterization Laboratory. Integr. Biol. Quant. Biosci. Nano Macro 2012, 5, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Dobrovolskaia, M.A.; Germolec, D.R.; Weaver, J.L. Evaluation of nanoparticle immunotoxicity. Nat. Nanotechnol. 2009, 4, 411–414. [Google Scholar] [CrossRef]
- OECD-WPMN. Report of the Workshop on Risk Assessment of Manufactured Nanomaterials in a Regulatory Context; No. 21-ENV/JM/MONO(2010)10; OECD. Available online: https://www.oecd.org/science/nanosafety/publications-series-safety-manufactured-nanomaterials.htm (accessed on 12 April 2022).
- Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR). Opinion on the Appropriateness of Existing Methodologies to Assess the Potential Risks Associated with Engineered and Adventitious Products of Nanotechnologies; Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR): Brussels, Belgium, 2005. [Google Scholar]
- Oberdörster, G. Safety assessment for nanotechnology and nanomedicine: Concepts of nanotoxicology. J. Intern. Med. 2010, 267, 89–105. [Google Scholar] [CrossRef]
- Oberdörster, G.; Oberdörster, E.; Oberdörster, J. Nanotoxicology: An Emerging Discipline Evolving from Studies of Ultrafine Particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef]
- Dobrovolskaia, M.A.; Shurin, M.R.; Shvedova, A.A. Current understanding of interactions between nanoparticles and the immune system. Toxicol. Appl. Pharmacol. 2016, 299, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Gioacchino, M.; Petrarca, C.; Lazzarin, F.; Di Giampaolo, L.; Sabbioni, E.; Boscolo, P.; Mariani-Costantini, R.; Bernardini, G. Immunotoxicity of nanoparticles. Int. J. Immunopathol. Pharmacol. 2011, 24, 65S–71S. [Google Scholar] [PubMed]
- Dobrovolskaia, M.A.; McNeil, S.E. Understanding the correlation between in vitro and in vivo immunotoxicity tests for nanomedicines. J. Control. Release Off. J. Control. Release Soc. 2013, 172, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Gajewicz, A. How to judge whether QSAR/read-across predictions can be trusted: A novel approach for establishing a model’s applicability domain. Environ. Sci. Nano 2017, 5, 408–421. [Google Scholar] [CrossRef]
- Mikolajczyk, A.; Gajewicz, A.; Mulkiewicz, E.; Rasulev, B.; Marchelek, M.; Diak, M.; Hirano, S.; Zaleska-Medynska, A.; Puzyn, T. Nano-QSAR modeling for ecosafe design of heterogeneous TiO2-based nano-photocatalysts. Environ. Sci. Nano 2018, 5, 1150–1160. [Google Scholar] [CrossRef]
- Huang, Y.; Li, X.; Xu, S.; Zheng, H.; Zhang, L.; Chen, J.; Hong, H.; Kusko, R.; Li, R. Quantitative Structure–Activity Relationship Models for Predicting Inflammatory Potential of Metal Oxide Nanoparticles. Environ. Health Perspect. 2020, 128, 067010. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risks to public health related to the presence of chromium in food and drinking water. EFSA J. 2014, 12, 3595. [Google Scholar]
- Pavlin, M.; Bregar, V.B.; Lojk, J.; Šuštar, V.; Veranic, P. Visualization of internalization of functionalized cobalt ferrite nanoparticles and their intracellular fate. Int. J. Nanomed. 2013, 8, 919–931. [Google Scholar] [CrossRef] [Green Version]
- Pavlin, M.; Lojk, J.; Bregar, V.B.; Rajh, M.; Mis, K.; Kreft, M.E.; Pirkmajer, S.; Veranic, P. Cell type-specific response to high intracellular loading of polyacrylic acid-coated magnetic nanoparticles. Int. J. Nanomed. 2015, 10, 1449–1462. [Google Scholar] [CrossRef] [Green Version]
- Strojan, K.; Leonardi, A.; Bregar, V.B.; Križaj, I.; Svete, J.; Pavlin, M. Dispersion of Nanoparticles in Different Media Importantly Determines the Composition of Their Protein Corona. PLoS ONE 2017, 12, e0169552. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-F.; Liu, Z.-G.; Shen, W.; Gurunathan, S. Silver Nanoparticles: Synthesis, Characterization, Properties, Applications, and Therapeutic Approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef] [PubMed]
- Dorier, M.; Brun, E.; Veronesi, G.; Barreau, F.; Pernet-Gallay, K.; Desvergne, C.; Rabilloud, T.; Carapito, C.; Herlin-Boime, N.; Carrière, M. Impact of anatase and rutile titanium dioxide nanoparticles on uptake carriers and efflux pumps in Caco-2 gut epithelial cells. Nanoscale 2015, 7, 7352–7360. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Manawan, M.; Feng, T.; Qian, R.; Zhao, T.; Zhou, G.; Kong, F.; Wang, Q.; Dai, S.; Pan, J.H. Anatase and rutile in evonik aeroxide P25: Heterojunctioned or individual nanoparticles? Catal. Today 2018, 300, 12–17. [Google Scholar] [CrossRef]
- Nicosia, A.; Vento, F.; Di Mari, G.M.; D’Urso, L.; Mineo, P. TiO2-Based Nanocomposites Thin Film Having Boosted Photocatalytic Activity for Xenobiotics Water Pollution Remediation. Nanomaterials 2021, 11, 400. [Google Scholar] [CrossRef] [PubMed]
- Dréno, B.; Alexis, A.; Chuberre, B.; Marinovich, M. Safety of titanium dioxide nanoparticles in cosmetics. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 34–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlin, M.; Bregar, V.B. Stability of nanoparticle suspensions in different biologically relevant media. Dig. J. Nanomater. Biostruct. 2012, 7, 1389–1400. [Google Scholar]
- Li, Y.; Fujita, M.; Boraschi, D. Endotoxin Contamination in Nanomaterials Leads to the Misinterpretation of Immunosafety Results. Front. Immunol. 2017, 8, 472. [Google Scholar] [CrossRef] [PubMed]
- Oostingh, G.J.; Casals, E.; Italiani, P.; Colognato, R.; Stritzinger, R.; Ponti, J.; Pfaller, T.; Kohl, Y.; Ooms, D.; Favilli, F.; et al. Problems and challenges in the development and validation of human cell-based assays to determine nanoparticle-induced immunomodulatory effects. Part. Fibre Toxicol. 2011, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Vittorio, O.; Raffa, V.; Cuschieri, A. Influence of purity and surface oxidation on cytotoxicity of multiwalled carbon nanotubes with human neuroblastoma cells. Nanomed. Nanotechnol. Biol. Med. 2009, 5, 424–431. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Semin. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef]
- Dostert, C.; Pétrilli, V.; Van Bruggen, R.; Steele, C.; Mossman, B.T.; Tschopp, J. Innate Immune Activation Through Nalp3 Inflammasome Sensing of Asbestos and Silica. Science 2008, 320, 674–677. [Google Scholar] [CrossRef] [Green Version]
- Halle, A.; Hornung, V.; Petzold, G.C.; Stewart, C.R.; Monks, B.G.; Reinheckel, T.; Fitzgerald, K.A.; Latz, E.; Moore, K.J.; Golenbock, D.T. The NALP3 inflammasome is involved in the innate immune response to amyloid-beta. Nat. Immunol. 2008, 9, 857–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelblanco, M.; Lugrin, J.; Ehirchiou, D.; Nasi, S.; Ishii, I.; So, A.; Martinon, F.; Busso, N. Hydrogen sulfide inhibits NLRP3 inflammasome activation and reduces cytokine production both in vitro and in a mouse model of inflammation. J. Biol. Chem. 2018, 293, 2546–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedé, S.; Blázquez, A.B.; Chiang, D.; Tordesillas, L.; Berin, M.C. The rise of food allergy: Environmental factors and emerging treatments. eBioMedicine 2016, 7, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun. Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannahan, J.H.; Brown, J.M. Engineered nanomaterial exposure and the risk of allergic disease. Curr. Opin. Allergy Clin. Immunol. 2014, 14, 95–99. [Google Scholar] [CrossRef]
- Lee, Y.K.; Choi, E.-J.; Webster, T.J.; Kim, S.-H.; Khang, D. Effect of the protein corona on nanoparticles for modulating cytotoxicity and immunotoxicity. Int. J. Nanomed. 2014, 10, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Szebeni, J. Complement activation-related pseudoallergy: A stress reaction in blood triggered by nanomedicines and biologicals. Mol. Immunol. 2014, 61, 163–173. [Google Scholar] [CrossRef]
- Szebeni, J. Mechanism of nanoparticle-induced hypersensitivity in pigs: Complement or not complement? Drug Discov. Today 2018, 23, 487–492. [Google Scholar] [CrossRef]
- Strojan, K.; Lojk, J.; Bregar, V.B.; Veranič, P.; Pavlin, M. Glutathione reduces cytotoxicity of polyethyleneimine coated magnetic nanoparticles in CHO cells. Toxicol. Vitro Int. J. Publ. Assoc. BIBRA 2017, 41, 12–20. [Google Scholar] [CrossRef]
- Treuel, L.; Docter, D.; Maskos, M.; Stauber, R.H. Protein corona—From molecular adsorption to physiological complexity. Beilstein J. Nanotechnol. 2015, 6, 857–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on Lead in Food: Lead in Food. EFSA J. 2010, 8, 1570. [Google Scholar]
- Cedervall, T.; Lynch, I.; Lindman, S.; Berggard, T.; Thulin, E.; Nilsson, H.; Dawson, K.A.; Linse, S. Understanding the nanoparticle-protein corona using methods to quantify exchange rates and affinities of proteins for nanoparticles. Proc. Natl. Acad. Sci. USA 2007, 104, 2050–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Wang, G.; Griffin, J.I.; Brenneman, B.; Banda, N.K.; Holers, N.K.B.V.M.; Backos, D.S.; Wu, L.; Moghimi, S.M.; Simberg, F.C.G.W.J.I.G.B.B.D. Complement proteins bind to nanoparticle protein corona and undergo dynamic exchange in vivo. Nat. Nanotechnol. 2016, 12, 387–393. [Google Scholar] [CrossRef]
- Aggarwal, P.; Hall, J.B.; McLeland, C.B.; Dobrovolskaia, M.A.; McNeil, S.E. Nanoparticle interaction with plasma proteins as it relates to particle biodistribution, biocompatibility and therapeutic efficacy. Adv. Drug Deliv. Rev. 2009, 61, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Hajipour, M.J.; Laurent, S.; Aghaie, A.; Rezaee, F.; Mahmoudi, M. Personalized protein coronas: A “key” factor at the nanobiointerface. Biomater. Sci. 2014, 2, 1210. [Google Scholar] [CrossRef]
- Escamilla-Rivera, V.; Uribe-Ramírez, M.; González-Pozos, S.; Lozano, O.; Lucas, S.; De Vizcaya-Ruiz, A. Protein corona acts as a protective shield against Fe3O4-PEG inflammation and ROS-induced toxicity in human macrophages. Toxicol. Lett. 2015, 240, 172–184. [Google Scholar] [CrossRef]
- Lin, P.-C.; Lin, S.; Wang, P.C.; Sridhar, R. Techniques for physicochemical characterization of nanomaterials. Biotechnol. Adv. 2013, 32, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Tenzer, S.; Docter, D.; Rosfa, S.; Wlodarski, A.; Kuharev, J.; Rekik, A.; Knauer, S.K.; Bantz, C.; Nawroth, T.; Bier, C.; et al. Nanoparticle Size Is a Critical Physicochemical Determinant of the Human Blood Plasma Corona: A Comprehensive Quantitative Proteomic Analysis. ACS Nano 2011, 5, 7155–7167. [Google Scholar] [CrossRef]
- Winzen, S.; Schoettler, S.; Baier, G.; Rosenauer, C.; Mailaender, V.; Landfester, K.; Mohr, K. Complementary analysis of the hard and soft protein corona: Sample preparation critically effects corona composition. Nanoscale 2015, 7, 2992–3001. [Google Scholar] [CrossRef] [Green Version]
- Lundqvist, M.; Stigler, J.; Cedervall, T.; Berggård, T.; Flanagan, M.B.; Lynch, I.; Elia, G.; Dawson, K.A. The Evolution of the Protein Corona around Nanoparticles: A Test Study. ACS Nano 2011, 5, 7503–7509. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.; Hunter, A.; Andresen, T. Factors Controlling Nanoparticle Pharmacokinetics: An Integrated Analysis and Perspective. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 481–503. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Krüger, K.; Klempt, M. Titanium dioxide nanoparticles induced activation of extracellular signal-regulated kinases signaling pathway in human colonic epithelial Caco-2 cells is mediated by epidermal growth factor receptor. Front. Nanosci. Nanotechnol. 2016, 2, 58–63. [Google Scholar] [CrossRef]
- Yazdi, A.S.; Guarda, G.; Riteau, N.; Drexler, S.K.; Tardivel, A.; Couillin, I.; Tschopp, J. Nanoparticles activate the NLR pyrin domain containing 3 (Nlrp3) inflammasome and cause pulmonary inflammation through release of IL-1alpha and IL-1beta. Proc. Natl. Acad. Sci. USA 2010, 107, 19449–19454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornung, V.; Bauernfeind, F.; Halle, A.; Samstad, E.O.; Kono, H.; Rock, K.L.; Fitzgerald, K.A.; Latz, E. Silica crystals and aluminum salts mediate NALP-3 inflammasome activation via phagosomal destabilization. Nat. Immunol. 2008, 9, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A. Pre-clinical immunotoxicity studies of nanotechnology-formulated drugs: Challenges, considerations and strategy. J. Control. Release Off. J. Control. Release Soc. 2015, 220, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Maher, B.A.; Ahmed, I.A.M.; Karloukovski, V.; MacLaren, D.A.; Foulds, P.G.; Allsop, D.; Mann, D.M.A.; Torres-Jardón, R.; Calderon-Garciduenas, L. Magnetite pollution nanoparticles in the human brain. Proc. Natl. Acad. Sci. USA 2016, 113, 10797–10801. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.; McLeland, C.B.; Potter, T.M.; Stern, S.T.; Adiseshaiah, P.P. Assessing NLRP3 Inflammasome Activation by Nanoparticles. In Characterization of Nanoparticles Intended for Drug Delivery; McNeil, S.E., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 135–147. [Google Scholar] [CrossRef]
- Rajamäki, K.; Lappalainen, J.; Öörni, K.; Välimäki, E.; Matikainen, S.; Kovanen, P.T.; Eklund, K.K. Cholesterol Crystals Activate the NLRP3 Inflammasome in Human Macrophages: A Novel Link between Cholesterol Metabolism and Inflammation. PLoS ONE 2010, 5, e11765. [Google Scholar] [CrossRef] [Green Version]
- Akinc, A.; Thomas, M.; Klibanov, A.M.; Langer, R. Exploring polyethylenimine-mediated DNA transfection and the proton sponge hypothesis. J. Gene Med. 2004, 7, 657–663. [Google Scholar] [CrossRef]
- van de Veerdonk, F.L.; Smeekens, S.P.; Joosten, L.A.B.; Kullberg, B.J.; Dinarello, C.A.; van der Meer, J.W.M.; Netea, M.G. Reactive oxygen species–independent activation of the IL-1β inflammasome in cells from patients with chronic granulomatous disease. Proc. Natl. Acad. Sci. USA 2010, 107, 3030–3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walkey, C.D.; Olsen, J.B.; Song, F.; Liu, R.; Guo, H.; Olsen, D.W.H.; Cohen, Y.; Emili, A.; Chan, W.C.W. Protein Corona Fingerprinting Predicts the Cellular Interaction of Gold and Silver Nanoparticles. ACS Nano 2014, 8, 2439–2455. [Google Scholar] [CrossRef] [PubMed]
- Neagu, M.; Piperigkou, Z.; Karamanou, K.; Engin, A.B.; Docea, A.O.; Constantin, C.; Negrei, C.; Nikitovic, D.; Tsatsakis, A. Protein bio-corona: Critical issue in immune nanotoxicology. Arch. Toxicol. 2016, 91, 1031–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrovolskaia, M.A.; Neun, B.W.; Man, S.; Ye, X.; Hansen, M.; Patri, A.K.; Crist, R.M.; McNeil, S.E. Protein corona composition does not accurately predict hematocompatibility of colloidal gold nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 1453–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf-Grosse, S.; Rokstad, A.M.; Ali, S.; Lambris, J.D.; Mollnes, T.E.; Nilsen, A.M.; Stenvik, J. Iron oxide nanoparticles induce cytokine secretion in a complement-dependent manner in a human whole blood model. Int. J. Nanomed. 2017, 12, 3927–3940. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.J.; Liang, M.; Toth, I.; Monteiro, M.J.; Minchin, R.F. Molecular Interaction of Poly(acrylic acid) Gold Nanoparticles with Human Fibrinogen. ACS Nano 2012, 6, 8962–8969. [Google Scholar] [CrossRef]
- Deng, Z.J.; Liang, M.; Monteiro, M.; Toth, I.; Minchin, R.F. Nanoparticle-induced unfolding of fibrinogen promotes Mac-1 receptor activation and inflammation. Nat. Nanotechnol. 2010, 6, 39–44. [Google Scholar] [CrossRef]
- Miller, M.R.; Poland, C.A. Nanotoxicology: The Need for a Human Touch? Small 2020, 16, 2001516. [Google Scholar] [CrossRef]
- Spurgeon, D.J.; Lahive, E.; Schultz, C.L. Nanomaterial Transformations in the Environment: Effects of Changing Exposure Forms on Bioaccumulation and Toxicity. Small 2020, 16, e2000618. [Google Scholar] [CrossRef]
- Shin, S.W.; Song, I.H.; Um, S.H. Role of Physicochemical Properties in Nanoparticle Toxicity. Nanomaterials 2015, 5, 1351–1365. [Google Scholar] [CrossRef] [Green Version]
- Skočaj, M.; Bizjak, M.; Strojan, K.; Lojk, J.; Kreft, M.E.; Miš, K.; Pirkmajer, S.; Bregar, V.B.; Veranič, P.; Pavlin, M. Proposing Urothelial and Muscle In Vitro Cell Models as a Novel Approach for Assessment of Long-Term Toxicity of Nanoparticles. Int. J. Mol. Sci. 2020, 21, 7545. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Biass, D.; Kordiš, D.; Stöcklin, R.; Favreau, P.; Križaj, I. Conus consors Snail Venom Proteomics Proposes Functions, Pathways, and Novel Families Involved in Its Venomic System. J. Proteome Res. 2012, 11, 5046–5058. [Google Scholar] [CrossRef] [PubMed]
- Bantscheff, M.; Schirle, M.; Sweetman, G.; Rick, J.; Kuster, B. Quantitative mass spectrometry in proteomics: A critical review. Anal. Bioanal. Chem. 2007, 389, 1017–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid-Burgk, J.L.; Gaidt, M.M.; Schmidt, T.; Ebert, T.S.; Bartok, E.; Hornung, V. Caspase-4 mediates non-canonical activation of the NLRP3 inflammasome in human myeloid cells. Eur. J. Immunol. 2015, 45, 2911–2917. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter\NP Type | PAA | Silver | SiO2 | TiO2 P25 | TiO2 ANAT | TiO2 DrA | TiO2 FG | TiO2 N.B. | |
|---|---|---|---|---|---|---|---|---|---|
| PCP | Zeta potentialRPMI [mV] | −14 ± 0.2 | −7.0 ± 0.3 | −7.3 ± 0.6 | −9.3 ± 0.9 | −7.4 ± 1.0 | −8.1 ± 0.1 | −8.2 ± 1.7 | −10.3 ± 1.03 |
| Z-averageRPMI [nm] | 199 ± 6 | 48 ± 2.6 | 871 ± 547 a | 589 ± 131 | 1040± 375 | 2182 ± 166 | 1552 ± 884 | 321 ± 95 | |
| Impurities | Endotoxin | NO | NO | NO | NO | NO | NO | NO | NO |
| Detected impurities | Mn, Co, Ti, Zn, Mg | Ag, Fe, Ba, Mn | Al, Ti, Fe, Zn, Pb | Mg, Al, Si, Ti, Mn, Fe, Co, Zn, Sr, Sb, As | Zn, Mg | Zn, Mg, Fe, Mn, V | Mg, Al, Ti, Mn, Fe, Ni, Zn, Ge, As, Sb, Ba, Pb | Zn, Mg, Fe, | |
| Protein Corona | HSP90A | NO | NO | NO | YES | NO | YES | NO | NO |
| FINC | NO | NO | NO | NO | YES | YES | YES | NO | |
| CFAH | NO | NO | NO | YES | NO | NO | NO | YES | |
| Toxicity mechanisms | IC90 [µg mL−1] | 13 | 2.5 | 2.9 | 12 | 4.2 | 9.1 | 103 | 15 |
| Membrane damage | NO | YES | YES | YES | NO | YES | NO | NO | |
| Internalization | YES | YES | YES | YES | YES | YES | YES | YES | |
| ROS | o | o | + | + | o | + | o | + | |
| Cytokine response | TNF-α | o | o | o | o | o | o | o | o |
| IL-6 | o | o | o | o | o | o | o | o | |
| IL-8 | o | + | o | + | + | + | + | o | |
| IL-1β | o | o | + | + | + | + | + | o | |
| Dhyd H2O | Dhyd RPMI | Z-aver H2O | Z-aver RPMI | Zeta H2O | Zeta RPMI | PDI H2O | PDI RPMI | IC90 | Mem dam. | ROS | IL-8 | IL-1β | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dhyd H2O | 1 | 0.72 | 0.55 | 0.58 | 0.44 | 0.11 | 0 | −0.3 | −0.2 | −0.39 | 0.12 | 0.01 | 0.58 |
| DhydRPMI | 0.72 | 1 | 0.4 | 0.93 | 0.06 | 0.31 | −0.08 | −0.2 | 0.17 | −0.28 | 0.05 | 0.43 | 0.75 |
| Z-av H2O | 0.55 | 0.4 | 1 | 0.56 | 0.31 | 0.37 | 0.77 | 0.27 | −0.19 | −0.34 | 0.44 | −0.06 | 0.5 |
| Z-avRPMI | 0.58 | 0.93 | 0.56 | 1 | 0.07 | 0.45 | 0.19 | 0.01 | 0.35 | −0.38 | 0.17 | 0.34 | 0.71 |
| Zeta H2O | 0.44 | 0.06 | 0.31 | 0.07 | 1 | 0.23 | 0.26 | −0.31 | −0.26 | −0.22 | 0.6 | 0.08 | 0.42 |
| ZetaRPMI | 0.11 | 0.31 | 0.37 | 0.45 | 0.23 | 1 | 0.17 | −0.12 | 0.17 | 0.43 | 0 | 0.37 | 0.26 |
| PDI H2O | 0 | −0.08 | 0.77 | 0.19 | 0.26 | 0.17 | 1 | 0.67 | −0.18 | −0.4 | 0.52 | −0.27 | 0.16 |
| PDI med | −0.3 | −0.2 | 0.27 | 0.01 | −0.31 | −0.12 | 0.67 | 1 | −0.08 | −0.38 | −0.15 | −0.56 | −0.37 |
| IC90 | −0.2 | 0.17 | −0.19 | 0.35 | −0.26 | 0.17 | −0.18 | −0.08 | 1 | −0.22 | −0.07 | 0.03 | −0.05 |
| Mem.dam | −0.39 | −0.28 | −0.34 | −0.38 | −0.22 | 0.43 | −0.4 | −0.38 | −0.22 | 1 | −0.3 | 0.45 | −0.2 |
| ROS | 0.12 | 0.05 | 0.44 | 0.17 | 0.6 | 0 | 0.52 | −0.15 | −0.07 | −0.3 | 1 | 0.36 | 0.66 |
| IL-8 | 0.01 | 0.43 | −0.06 | 0.34 | 0.08 | 0.37 | −0.27 | −0.56 | 0.03 | 0.45 | 0.36 | 1 | 0.69 |
| IL-1b | 0.58 | 0.75 | 0.5 | 0.71 | 0.42 | 0.26 | 0.16 | −0.37 | −0.05 | −0.2 | 0.66 | 0.69 | 1 |
| Dhyd H2O | Dhyd RPMI | Z-aver H2O | Z-aver RPMI | PDI H2O | PDI RPMI | Zeta H2O | Zeta RPMI | IC90 | Mem dam | ROS | IL-8 | IL-1β | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PROTab | −0.09 | 0.27 | 0.14 | 0.29 | 0.01 | −0.31 | −0.22 | −0.02 | 0.21 | 0.11 | 0.53 | 0.71 | 0.6 |
| A1AT | 0.59 | 0.75 | 0.47 | 0.6 | −0.06 | −0.32 | −0.11 | 0.17 | −0.21 | 0.08 | 0.16 | 0.57 | 0.74 |
| ALBU | −0.05 | 0.4 | 0.03 | 0.41 | −0.14 | −0.43 | −0.17 | 0.09 | 0.4 | 0.1 | 0.48 | 0.79 | 0.65 |
| ANT3 | 0.21 | 0.39 | −0.03 | 0.17 | −0.34 | −0.66 | 0.15 | 0.18 | −0.33 | 0.48 | 0.31 | 0.9 | 0.68 |
| APOA1 | −0.3 | −0.4 | 0.47 | −0.22 | 0.68 | 0.57 | −0.33 | −0.14 | −0.23 | −0.02 | 0.19 | −0.32 | −0.21 |
| APOE | −0.16 | −0.49 | −0.14 | −0.5 | 0.09 | 0.27 | −0.24 | −0.89 | −0.06 | −0.47 | 0.03 | −0.63 | −0.43 |
| CFAH | −0.26 | −0.19 | −0.32 | −0.24 | −0.1 | −0.43 | 0.51 | −0.08 | −0.09 | 0.14 | 0.65 | 0.57 | 0.36 |
| FETUA | −0.61 | −0.44 | −0.48 | −0.38 | −0.36 | −0.23 | −0.56 | −0.2 | 0.56 | 0.31 | −0.12 | 0.06 | −0.39 |
| FINC | 0.13 | 0.32 | 0.07 | 0.41 | 0.19 | 0.48 | 0.17 | 0.34 | 0.1 | −0.31 | −0.29 | −0.22 | −0.04 |
| HBA | 0.13 | 0.24 | 0.72 | 0.01 | 0.87 | 0.46 | 0.06 | 0.26 | −0.21 | −0.01 | 0.45 | −0.13 | 0.07 |
| HBBF | 0.51 | 0.59 | 0.75 | 0.57 | 0.38 | 0.01 | −0.04 | 0.16 | −0.3 | −0.08 | 0.37 | 0.38 | 0.71 |
| HS90A | 0.03 | 0.15 | −0.15 | 0.04 | −0.15 | −0.55 | 0.51 | −0.02 | −0.14 | 0.09 | 0.68 | 0.75 | 0.65 |
| IPSP | −0.35 | −0.3 | −0.43 | −0.43 | −0.51 | −0.26 | −0.4 | 0.3 | −0.21 | 0.93 | −0.61 | 0.19 | −0.44 |
| TETN | −0.37 | −0.25 | −0.45 | −0.25 | −0.19 | 0.27 | −0.59 | −0.8 | 0.38 | −0.42 | −0.2 | −0.38 | −0.4 |
| THRB | −0.46 | −0.51 | −0.45 | −0.63 | −0.41 | −0.37 | −0.15 | 0.22 | −0.33 | 0.95 | −0.28 | 0.29 | −0.35 |
| TSP1 | 0.67 | 0.82 | 0.32 | 0.64 | −0.17 | −0.48 | 0.17 | 0.11 | −0.16 | −0.06 | 0.32 | 0.68 | 0.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlin, M.; Lojk, J.; Strojan, K.; Hafner-Bratkovič, I.; Jerala, R.; Leonardi, A.; Križaj, I.; Drnovšek, N.; Novak, S.; Veranič, P.; et al. The Relevance of Physico-Chemical Properties and Protein Corona for Evaluation of Nanoparticles Immunotoxicity—In Vitro Correlation Analysis on THP-1 Macrophages. Int. J. Mol. Sci. 2022, 23, 6197. https://doi.org/10.3390/ijms23116197
Pavlin M, Lojk J, Strojan K, Hafner-Bratkovič I, Jerala R, Leonardi A, Križaj I, Drnovšek N, Novak S, Veranič P, et al. The Relevance of Physico-Chemical Properties and Protein Corona for Evaluation of Nanoparticles Immunotoxicity—In Vitro Correlation Analysis on THP-1 Macrophages. International Journal of Molecular Sciences. 2022; 23(11):6197. https://doi.org/10.3390/ijms23116197
Chicago/Turabian StylePavlin, Mojca, Jasna Lojk, Klemen Strojan, Iva Hafner-Bratkovič, Roman Jerala, Adrijana Leonardi, Igor Križaj, Nataša Drnovšek, Saša Novak, Peter Veranič, and et al. 2022. "The Relevance of Physico-Chemical Properties and Protein Corona for Evaluation of Nanoparticles Immunotoxicity—In Vitro Correlation Analysis on THP-1 Macrophages" International Journal of Molecular Sciences 23, no. 11: 6197. https://doi.org/10.3390/ijms23116197
APA StylePavlin, M., Lojk, J., Strojan, K., Hafner-Bratkovič, I., Jerala, R., Leonardi, A., Križaj, I., Drnovšek, N., Novak, S., Veranič, P., & Bregar, V. B. (2022). The Relevance of Physico-Chemical Properties and Protein Corona for Evaluation of Nanoparticles Immunotoxicity—In Vitro Correlation Analysis on THP-1 Macrophages. International Journal of Molecular Sciences, 23(11), 6197. https://doi.org/10.3390/ijms23116197