
Nerve Growth Factor (NGF) as Partaker in the Modulation of UV-Response in Cultured Human Conjunctival Fibroblasts

 ,
, 
Abstract
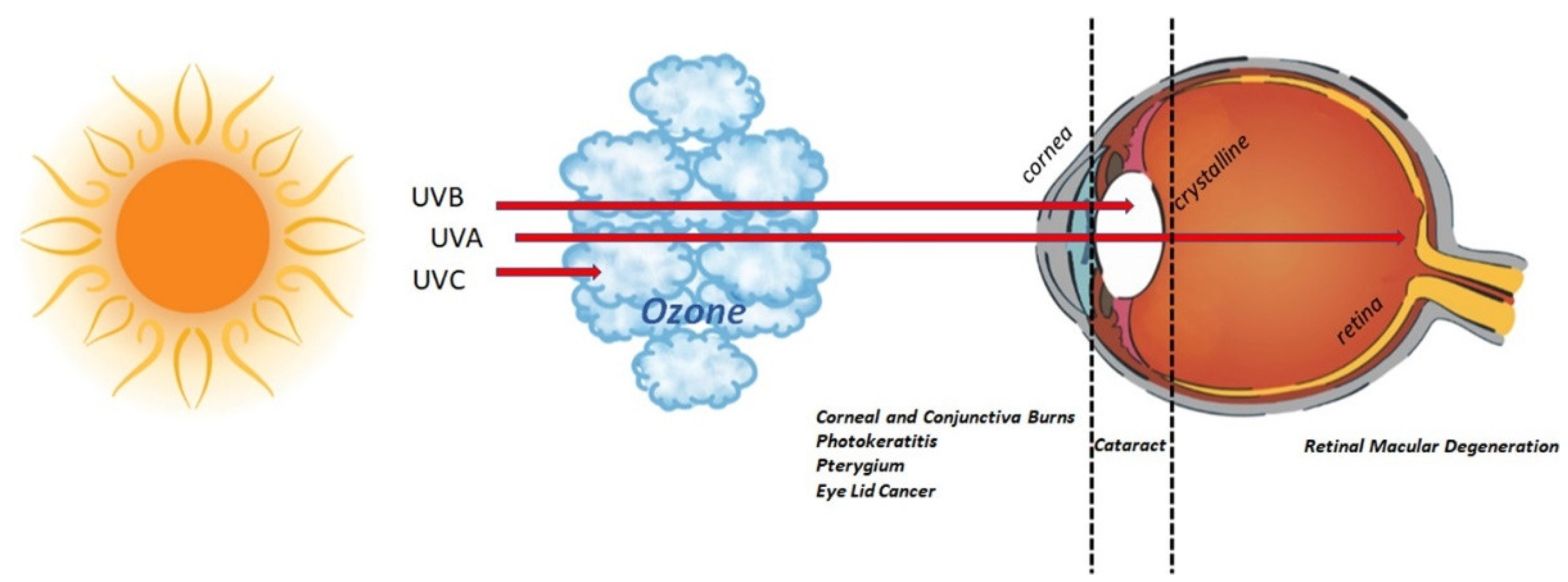
:1. Introduction
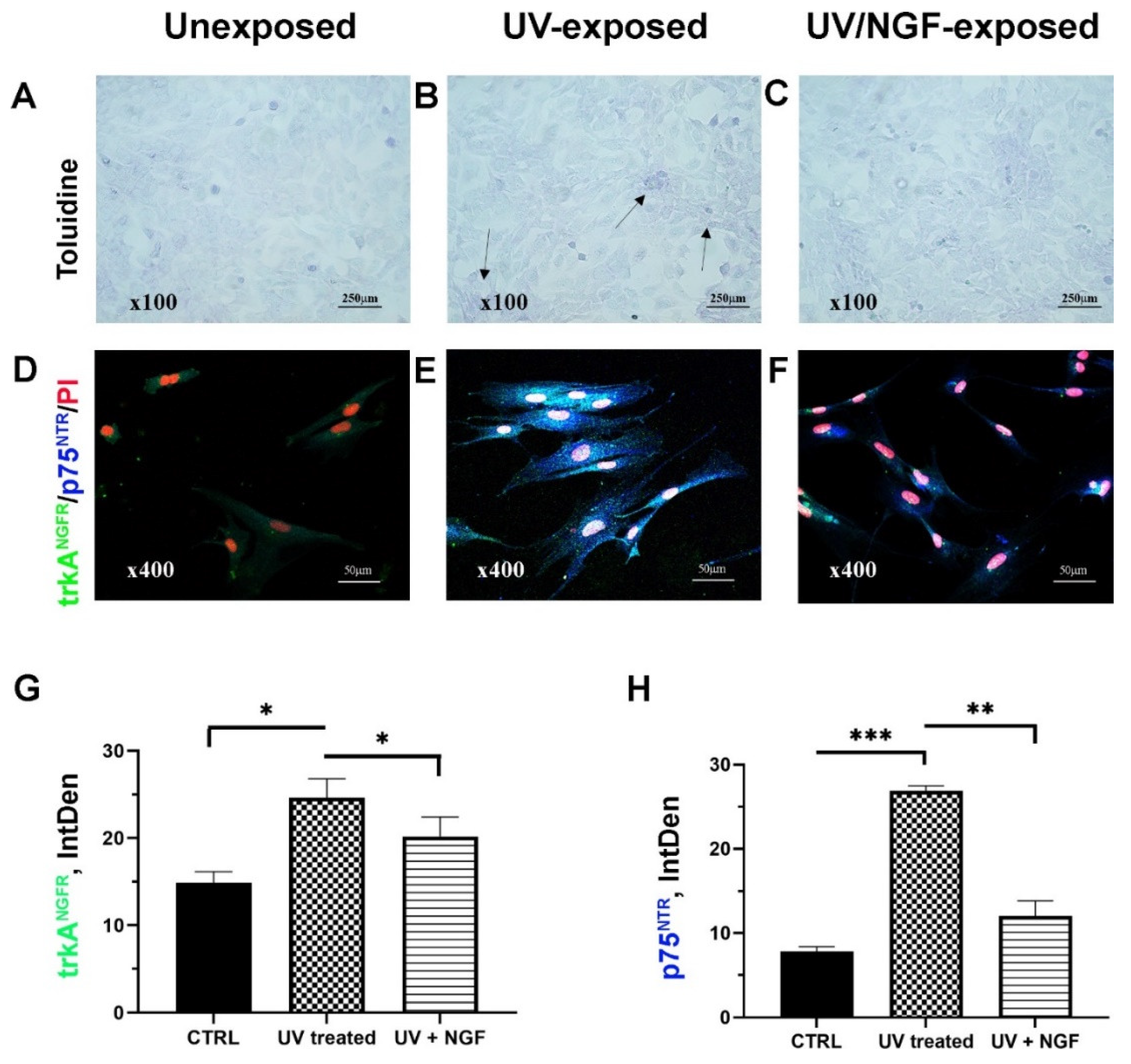
2. Results
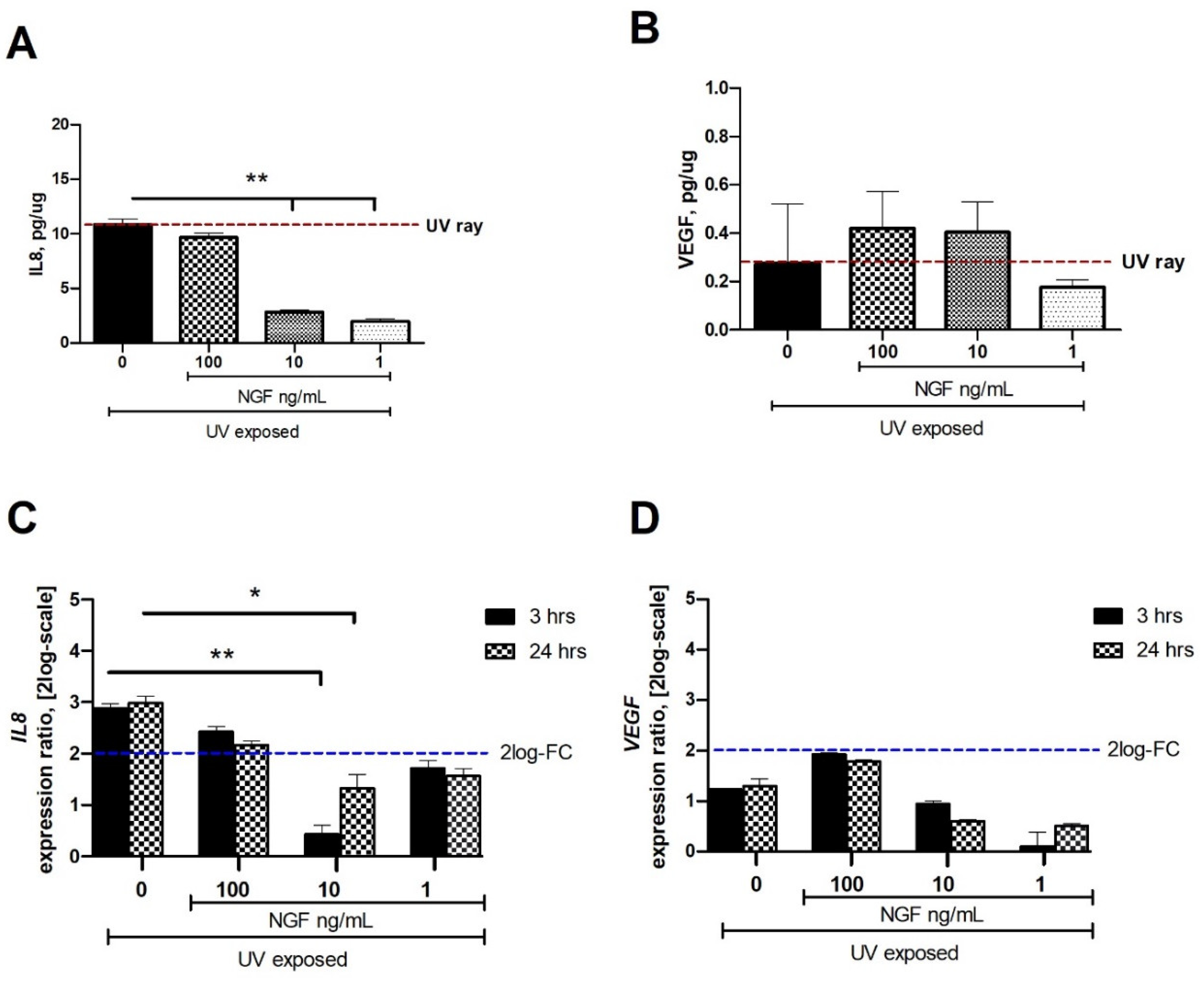
2.1. Changes in IL8 and VEGF Levels in the Conditioned Media from UV and UV/NGF Exposures
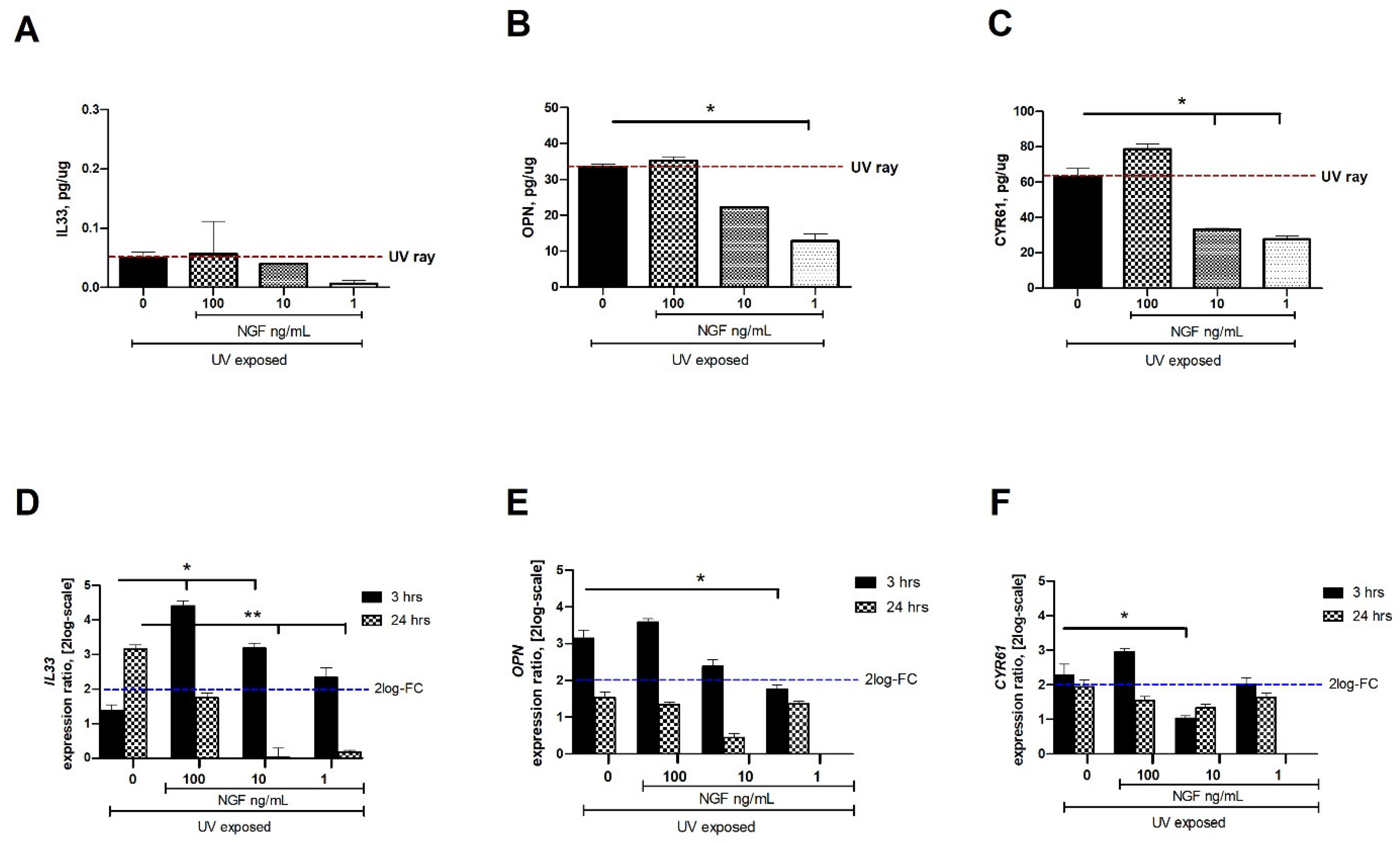
2.2. Changes in IL33, OPN, and CYR61 Matrix-Associated Proteins upon Exposures
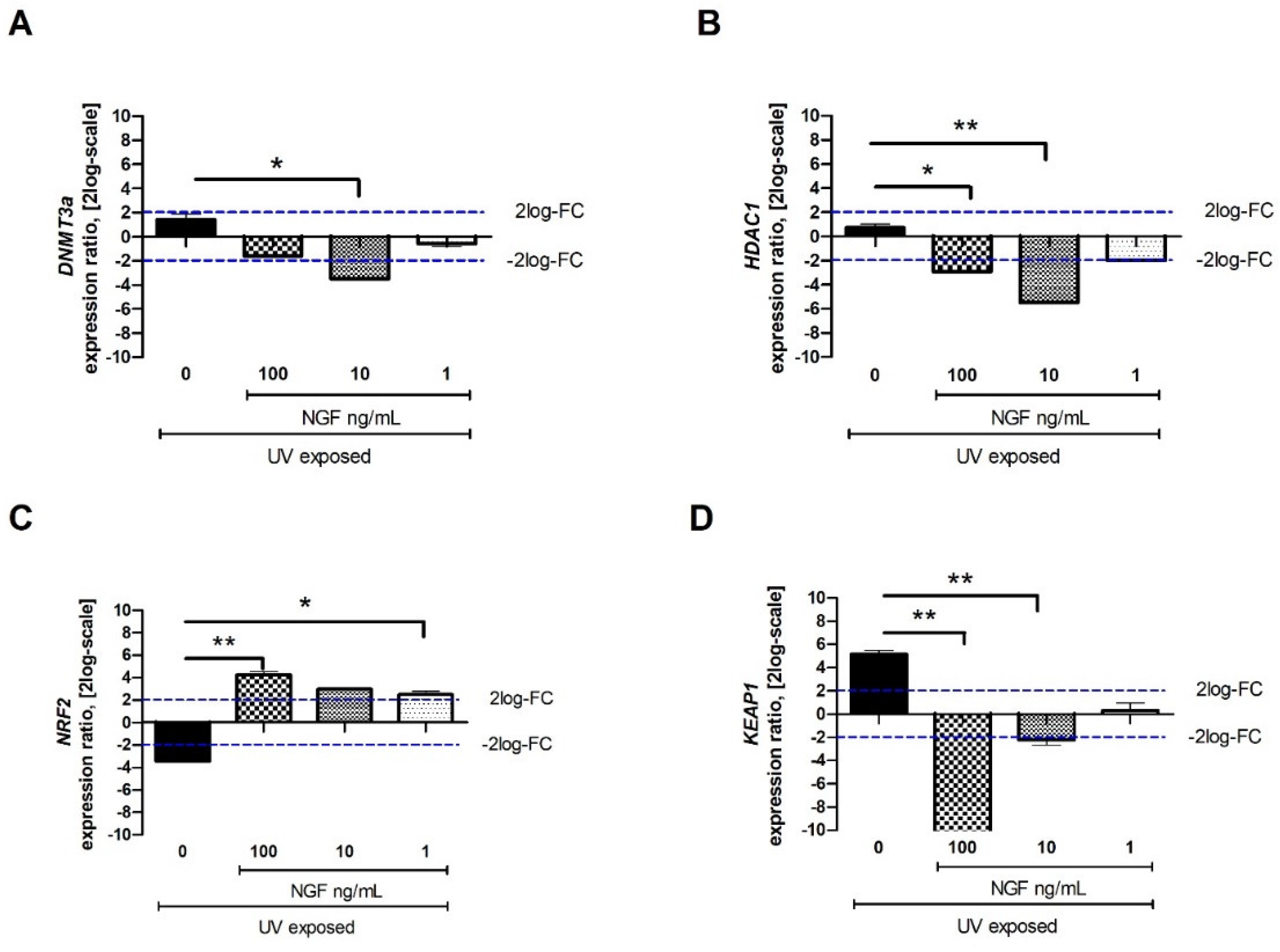
2.3. DNMT3a, HDAC1, NRF2, and KEAP1 Transcript Expression upon Exposures
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Analytical Reagents
4.2. UV Experiments and Subgroups
4.3. Conditioned Media and Monolayer Preparation
4.4. Light and Confocal Microscopy
4.4.1. Toluidine Blue Staining
4.4.2. Immunofluorescence
4.5. Protein Analysis
4.5.1. Ella Microfluidics Platform
4.5.2. ELISA Analysis
4.6. Molecular Analysis: Two-Step Real Time PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolffsohn, J.S.; Dhallu, S.; Aujla, M.; Laughton, D.; Tempany, K.; Powell, D.; Gifford, K.; Gifford, P.; Wan, K.; Cho, P.; et al. International multi-centre study of potential benefits of ultraviolet radiation protection using contact lenses. Contact Lens Anterior Eye J. Br. Contact Lens Assoc. 2022, 101593, in press. [Google Scholar] [CrossRef]
- Busanello, A.; Santucci, D.; Bonini, S.; Micera, A. Review: Environmental impact on ocular surface disorders: Possible epigenetic mechanism modulation and potential biomarkers. Ocul. Surf. 2017, 15, 680–687. [Google Scholar] [CrossRef]
- Rolando, M.; Cantera, E.; Mencucci, R.; Rubino, P.; Aragona, P. The correct diagnosis and therapeutic management of tear dysfunction: Recommendations of the P.I.C.A.S.S.O. board. Int. Ophthalmol. 2018, 38, 875–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.; Katiyar, S.K. Crosstalk Among UV-Induced Inflammatory Mediators, DNA Damage and Epigenetic Regulators Facilitates Suppression of the Immune System. Photochem. Photobiol. 2017, 93, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Hollmann, G.; Linden, R.; Giangrande, A.; Allodi, S. Increased p53 and decreased p21 accompany apoptosis induced by ultraviolet radiation in the nervous system of a crustacean. Aquat. Toxicol. 2016, 173, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Ishida, H.; Kiyokawa, E.; Singh, D.P.; Sasaki, H.; Kubo, E. Relative gene expression analysis of human pterygium tissues and UV radiation-evoked gene expression patterns in corneal and conjunctival cells. Exp. Eye Res. 2020, 199, 108194. [Google Scholar] [CrossRef]
- Kennedy, M.; Kim, K.H.; Harten, B.; Brown, J.; Planck, S.; Meshul, C.; Edelhauser, H.; Rosenbaum, J.T.; Armstrong, C.A.; Ansel, J.C. Ultraviolet irradiation induces the production of multiple cytokines by human corneal cells. Investig. Ophthalmol. Vis. Sci. 1997, 38, 2483–2491. [Google Scholar]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Boxman, I.L.A.; Ruwhof, C.; Boerman, O.C.; Löwik, C.W.G.M.; Ponec, M. Role of fibroblasts in the regulation of proinflammatory interleukin IL-1, IL-6 and IL-8 levels induced by keratinocyte-derived IL-1. Arch. Dermatol. Res. 1996, 288, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Rocco, M.L.; Balzamino, B.O.; Passeri, P.P.; Micera, A.; Aloe, L. Effect of Purified Murine NGF on Isolated Photoreceptors of a Rodent Developing Retinitis Pigmentosa. PLoS ONE 2015, 10, e0124810. [Google Scholar] [CrossRef] [Green Version]
- Rocco, M.L.; Balzamino, B.O.; Aloe, L.; Micera, A. NGF protects corneal, retinal, and cutaneous tissues/cells from phototoxic effect of UV exposure. Graefe’s Arch. Clin. Exp. 2018, 256, 729–738. [Google Scholar] [CrossRef]
- Byrne, S.N.; Beaugie, C.; O’Sullivan, C.; Leighton, S.; Halliday, G.M. The Immune-Modulating Cytokine and Endogenous Alarmin Interleukin-33 Is Upregulated in Skin Exposed to Inflammatory UVB Radiation. Am. J. Pathol. 2011, 179, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Kim, Y.K.; Eun, H.C.; Cho, K.H.; Chung, J.H. All-Trans Retinoic Acid Antagonizes UV-Induced VEGF Production and Angiogenesis via the Inhibition of ERK Activation in Human Skin Keratinocytes. J. Investig. Dermatol. 2006, 126, 2697–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, E.Y.-C.; Liu, P.-K.; Wen, Y.-T.; Quinn, P.M.J.; Levi, S.R.; Wang, N.-K.; Tsai, R.-K. Role of Oxidative Stress in Ocular Diseases Associated with Retinal Ganglion Cells Degeneration. Antioxidants 2021, 10, 1948. [Google Scholar] [CrossRef]
- Hibbert, S.A.; Watson, R.E.; Gibbs, N.K.; Costello, P.; Baldock, C.; Weiss, A.S.; Griffiths, C.E.; Sherratt, M.J. A potential role for endogenous proteins as sacrificial sunscreens and antioxidants in human tissues. Redox Biol. 2015, 5, 101–113. [Google Scholar] [CrossRef]
- Chang, P.-L.; Hsieh, Y.-H.; Wang, C.-C.; Juliana, M.; Tsuruta, Y.; Timares, L.; Elmets, C.; Ho, K.-J. Osteopontin facilitates ultraviolet B-induced squamous cell carcinoma development. J. Dermatol. Sci. 2014, 75, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, T.; Qin, Z.; Xu, Y.; He, T.; Kang, S.; Voorhees, J.J.; Fisher, G.J. Ultraviolet Irradiation Induces CYR61/CCN1, a Mediator of Collagen Homeostasis, through Activation of Transcription Factor AP-1 in Human Skin Fibroblasts. J. Investig. Dermatol. 2010, 130, 1697–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedittis, G.; Ciccacci, C.; Latini, A.; Novelli, L.; Novelli, G.; Borgiani, P. Emerging Role of microRNAs and Long Non-Coding RNAs in Sjögren’s Syndrome. Genes 2021, 12, 903. [Google Scholar] [CrossRef]
- Li, D.; Nie, J.; Han, Y.; Ni, L. Epigenetic Mechanism and Therapeutic Implications of Atrial Fibrillation. Front. Cardiovasc. Med. 2022, 8, 763824. [Google Scholar] [CrossRef]
- Campos, A.C.; Molognoni, F.; Melo, F.H.; Galdieri, L.C.; Carneiro, C.R.; D’Almeida, V.; Correa, M.; Jasiulionis, M.G. Oxidative Stress Modulates DNA Methylation during Melanocyte Anchorage Blockade Associated with Malignant Transformation. Neoplasia 2007, 9, 1111–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhou, P.; Lu, Y.; Luo, Y. Progresses of DNA methylation in common ocular tumor. [Zhonghua yan ke za zhi]. Chin. J. Ophthalmol. 2015, 51, 950–954. [Google Scholar]
- Zhu, Q.; Battu, A.; Ray, A.; Wani, G.; Qian, J.; He, J.; Wang, Q.-E.; Wani, A.A. Damaged DNA-binding protein down-regulates epigenetic mark H3K56Ac through histone deacetylase 1 and 2. Mutat. Res. Mol. Mech. Mutagen. 2015, 776, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Micera, A.; Puxeddu, I.; Aloe, L.; Levi-Schaffer, F. New insights on the involvement of Nerve Growth Factor in allergic inflammation and fibrosis. Cytokine Growth Factor Rev. 2003, 14, 369–374. [Google Scholar] [CrossRef]
- Micera, A.; Puxeddu, I.; Balzamino, B.O.; Bonini, S.; Levi-Schaffer, F. Chronic Nerve Growth Factor Exposure Increases Apoptosis in a Model of In Vitro Induced Conjunctival Myofibroblasts. PLoS ONE 2012, 7, e47316. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Balzamino, B.O.; Stigliano, E.; Biamonte, F.; Urbani, A.; Micera, A. Nerve Growth Factor (NGF) modulates in vitro induced myofibroblasts by highlighting a differential protein signature. Sci. Rep. 2021, 11, 1672. [Google Scholar] [CrossRef]
- Esposito, G.; Balzamino, B.O.; Bruno, L.; Cacciamani, A.; Micera, A. NGF in Inflammatory and Neurodegenerative Diseases of the Eye: New Findings Supporting Neuroprotection and Proper Tissue Remodeling in Vitreoretinal Disorders. Adv. Exp. Med. Biol. 2021, 1331, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Micera, A.; Stampachiacchiere, B.; Di Zazzo, A.; Sgrulletta, R.; Cortes, M.; Normando, E.M.; Lambiase, A.; Bonini, S. NGF Modulates trkANGFR/p75NTR in αSMA-Expressing Conjunctival Fibroblasts from Human Ocular Cicatricial Pemphigoid (OCP). PLoS ONE 2015, 10, e0142737. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.V.; Mappes, T.; Schaupp, P.; Lappe, C.; Wahl, S. Ultraviolet radiation oxidative stress affects eye health. J. Biophotonics 2018, 11, e201700377. [Google Scholar] [CrossRef] [Green Version]
- Söderberg, P.G.; Michael, R.; Merriam, J.C. Maximum acceptable dose of ultraviolet radiation: A safety limit for cataract. Acta Ophthalmol. Scand. 2003, 81, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Grimes, D.R. Ultraviolet radiation therapy and UVR dose models. Med. Phys. 2015, 42, 440–455. [Google Scholar] [CrossRef]
- Yam, J.C.S.; Kwok, A.K.H. Ultraviolet light and ocular diseases. Int. Ophthalmol. 2014, 34, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Artigas, J.M.; Felipe, A.; Navea, A.; Artigas, C.; Menezo, J. Ultraviolet radiation at Mediterranean latitudes and protection efficacy of intraocular lenses. J. Fr. D’ophtalmologie 2013, 36, 23–28. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, F.; Fan, D.; Jiang, Q.; Xue, Z.; Zhang, J.; Yu, X.; Li, K.; Qu, J.; Su, J. EyeDiseases: An integrated resource for dedicating to genetic variants, gene expression and epigenetic factors of human eye diseases. NAR Genom. Bioinform. 2021, 3, lqab050. [Google Scholar] [CrossRef]
- Micera, A.; Puxeddu, I.; Lambiase, A.; Antonelli, A.; Bonini, S.; Aloe, L.; Pe’Er, J.; Levi-Schaffer, F. The pro-fibrogenic effect of nerve growth factor on conjunctival fibroblasts is mediated by transforming growth factor-beta. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2005, 35, 650–656. [Google Scholar] [CrossRef]
- Micera, A.; Lambiase, A.; Stampachiacchiere, B.; Bonini, S.; Levi-Schaffer, F. Nerve growth factor and tissue repair remodeling: trkANGFR and p75NTR, two receptors one fate. Cytokine Growth Factor Rev. 2007, 18, 245–256. [Google Scholar] [CrossRef]
- Di Girolamo, N.; Kumar, R.K.; Coroneo, M.T.; Wakefield, D. UVB-mediated induction of interleukin-6 and -8 in pterygia and cultured human pterygium epithelial cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3430–3437. [Google Scholar]
- Di Girolamo, N.; Wakefield, D.; Coroneo, M.T. UVB-Mediated Induction of Cytokines and Growth Factors in Pterygium Epithelial Cells Involves Cell Surface Receptors and Intracellular Signaling. Investig. Opthalmology Vis. Sci. 2006, 47, 2430–2437. [Google Scholar] [CrossRef]
- Kondo, S.; Kono, T.; Sauder, D.N.; McKenzie, R.C. IL-8 Gene Expression and Production in Human Keratinocytes and Their Modulation by UVB. J. Investig. Dermatol. 1993, 101, 690–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delic, N.C.; Lyons, G.; Di Girolamo, N.; Halliday, G.M. Damaging Effects of Ultraviolet Radiation on the Cornea. Photochem. Photobiol. 2017, 93, 920–929. [Google Scholar] [CrossRef] [Green Version]
- Qazi, Y.; Wong, G.; Monson, B.; Stringham, J.; Ambati, B.K. Corneal transparency: Genesis, maintenance and dysfunction. Brain Res. Bull. 2010, 81, 198–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doutch, J.J.; Quantock, A.J.; Joyce, N.C.; Meek, K.M. Ultraviolet Light Transmission through the Human Corneal Stroma Is Reduced in the Periphery. Biophys. J. 2012, 102, 1258–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, D.A.; Rocha, E.M.; Aragona, P.; Clayton, J.A.; Ding, J.; Golebiowski, B.; Hampel, U.; McDermott, A.M.; Schaumberg, D.A.; Srinivasan, S.; et al. TFOS DEWS II Sex, gender, and hormones report. Ocul. Surf. 2017, 15, 284–333. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Q.; Liu, F. DNA Methyltransferase Inhibitors and their Therapeutic Potential. Curr. Top. Med. Chem. 2019, 18, 2448–2457. [Google Scholar] [CrossRef]
- Vaissière, T.; Sawan, C.; Herceg, Z. Epigenetic interplay between histone modifications and DNA methylation in gene silencing. Mutat. Res. Mutat. Res. 2008, 659, 40–48. [Google Scholar] [CrossRef]
- Brunmeir, R.; Lagger, S.; Seiser, C. Histone deacetylase HDAC1/HDAC2-controlled embryonic development and cell differentiation. Int. J. Dev. Biol. 2009, 53, 275–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thambyrajah, R.; Fadlullah, M.Z.; Proffitt, M.; Patel, R.; Cowley, S.; Kouskoff, V.; Lacaud, G. HDAC1 and HDAC2 Modulate TGF-β Signaling during Endothelial-to-Hematopoietic Transition. Stem Cell Rep. 2018, 10, 1369–1383, PMCID:PMC5998800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xu, Y.; Huang, X.; Ye, D.; Han, M.; Wang, H.-L. Regulatory Roles of Histone Deacetylases 1 and 2 in Pb-induced Neurotoxicity. Toxicol. Sci. 2018, 162, 688–701. [Google Scholar] [CrossRef] [Green Version]
- Mimura, J.; Kosaka, K.; Maruyama, A.; Satoh, T.; Harada, N.; Yoshida, H.; Satoh, K.; Yamamoto, M.; Itoh, K. Nrf2 regulates NGF mRNA induction by carnosic acid in T98G glioblastoma cells and normal human astrocytes. J. Biochem. 2011, 150, 209–217. [Google Scholar] [CrossRef]
- Bergmanson, J.P.; Sheldon, T.M. Ultraviolet radiation revisited. CLAO J. 1997, 23, 196–204. [Google Scholar]
- Palazzo, M.; Vizzarri, F.; Ondruška, L.; Rinaldi, M.; Pacente, L.; Guerra, G.; Merolla, F.; Caruso, C.; Costagliola, C. Corneal UV Protective Effects of a Topical Antioxidant Formulation: A Pilot Study on In Vivo Rabbits. Int. J. Mol. Sci. 2020, 21, 5426. [Google Scholar] [CrossRef] [PubMed]
- Naik, E.M.; Michalak, E.; Villunger, A.; Adams, J.M.; Strasser, A. Ultraviolet radiation triggers apoptosis of fibroblasts and skin keratinocytes mainly via the BH3-only protein Noxa. J. Cell Biol. 2007, 176, 415–424, PMCID:PMC2063977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, M.F.; Echenique, J.; López, J.M.; Medina, E.; Irós, M.; Serra, H.M.; Fini, M.E. Transcriptome Analysis of Pterygium and Pinguecula Reveals Evidence of Genomic Instability Associated with Chronic Inflammation. Int. J. Mol. Sci. 2021, 22, 12090, PMCID:PMC8584501. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R. Corneal Changes Associated with Chronic UV Irradiation. Arch. Ophthalmol. 1989, 107, 1481–1484. [Google Scholar] [CrossRef]
- Roberts, J.E. Ultraviolet Radiation as a Risk Factor for Cataract and Macular Degeneration. Eye Contact Lens 2011, 37, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Volatier, T.; Schumacher, B.; Cursiefen, C.; Notara, M. UV Protection in the Cornea: Failure and Rescue. Biology 2022, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Behboudifard, S.; Kluth, M.A.; Maßlo, C.; Ganss, C.; Frank, M.H.; Schumacher, B.; Cursiefen, C. UV light-blocking contact lenses protect against short-term UVB-induced limbal stem cell niche damage and inflammation. Sci. Rep. 2018, 8, 12564, PMCID:PMC6105637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonini, M.; Gaudenzi, D.; Spelta, S.; Sborgia, G.; Poddi, M.; Micera, A.; Sgrulletta, R.; Coassin, M.; Di Zazzo, A. Ocular Surface Failure in Urban Syndrome. J. Clin. Med. 2021, 10, 3048. [Google Scholar] [CrossRef]
- R Studio Team. R Studio: Integrated Development for R; R Studio, Inc.: Boston, MA, USA, 2016; Available online: http://www.rstudio.com (accessed on 1 February 2021).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Rieger, G. Anti-oxidative capacity of various artificial tear preparations. Graefes Arch. Clin. Exp. Ophthalmol. 2001, 239, 222–226. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Accession Number | Sequence (For/Rev; 5′-3′) | AT/Amplicon |
|---|---|---|
| IL8 BC013615 | 5′- TCT CTT GGC AGC CTT CCT G -3′ 5′- TGG GGT GGA AAG GTT TGG -3′ | 59 °C/116 bps |
| VEGF AF022375 | 5′- CTC CGT AGT AGC CGT GGT CT -3′ 5′- CCC CTC TCC TCT TCC TTC TC -3′ | 61 °C/131 bps |
| IL33 AY905581 | 5′- TGA GTC TCA ACA CCC CTC AA-3′ 5′- AAG ACA AAG AAG GCC TGG T-3′ | 59 °C/136 bps |
| OPN J04765 | 5′- GAA ACC CAC AGC CAC AAG C -3′ 5′- CTG TGG AAT TCA CGG CTG AC -3′ | 60 °C/139 bps |
| CYR61 BC009199 | 5′- CAC CCT TCT CAC TTG ACC-3′ 5′- CGT TTT GCT GCA GTC CTC-3′ | 59 °C/106 bps |
| DNMT3a C032392 | 5′-GCA CTC AAG GGC AGC AGA TA-3′ 5′-TTC CAG GCT TCC AGG GTT AG-3′ | 59 °C/129 bps |
| HDAC1 U50079 | 5′- GGG ATC GGT TAG GTT GCT TC-3′ 5′- AGG GCC ACA GCT GTC CTC ATA-3′ | 59 °C/100 bps |
| NRF2 BC011558 | 5′- ACA CGG TCC ACA GCT CAT C-3′ 5′- TGC CTC CAA AGT ATG TCA ATC A-3′ | 58 °C/107 bps |
| KEAP1 BC015945 | 5′- GGG TCC CCT ACA GCC AAG-3′ 5′- TGG GGT TCC AGA AGA TAA GC-3′ | 58 °C/106 bps |
| NGF V01511 | 5′- CTG GCC ACA CTG AGG TCG AT-3′ 5′- TCC TGC AGG GAC ATT GCT CTC-3′ | 53 °C/120 bps |
| GAPDH BC013310 | 5′-GAA GGG GTC ATT GAT GGC AAC-3′ 5′-GGG AAG GTG AAG GTC GAG AGT C-3′ | 60 °C/111 bps |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, G.; Balzamino, B.O.; Rocco, M.L.; Aloe, L.; Micera, A. Nerve Growth Factor (NGF) as Partaker in the Modulation of UV-Response in Cultured Human Conjunctival Fibroblasts. Int. J. Mol. Sci. 2022, 23, 6337. https://doi.org/10.3390/ijms23116337
Esposito G, Balzamino BO, Rocco ML, Aloe L, Micera A. Nerve Growth Factor (NGF) as Partaker in the Modulation of UV-Response in Cultured Human Conjunctival Fibroblasts. International Journal of Molecular Sciences. 2022; 23(11):6337. https://doi.org/10.3390/ijms23116337
Chicago/Turabian StyleEsposito, Graziana, Bijorn Omar Balzamino, Maria Luisa Rocco, Luigi Aloe, and Alessandra Micera. 2022. "Nerve Growth Factor (NGF) as Partaker in the Modulation of UV-Response in Cultured Human Conjunctival Fibroblasts" International Journal of Molecular Sciences 23, no. 11: 6337. https://doi.org/10.3390/ijms23116337
APA StyleEsposito, G., Balzamino, B. O., Rocco, M. L., Aloe, L., & Micera, A. (2022). Nerve Growth Factor (NGF) as Partaker in the Modulation of UV-Response in Cultured Human Conjunctival Fibroblasts. International Journal of Molecular Sciences, 23(11), 6337. https://doi.org/10.3390/ijms23116337