

AQP4, Astrogenesis, and Hydrocephalus: A New Neurological Perspective

 , , , , and
, , , , and 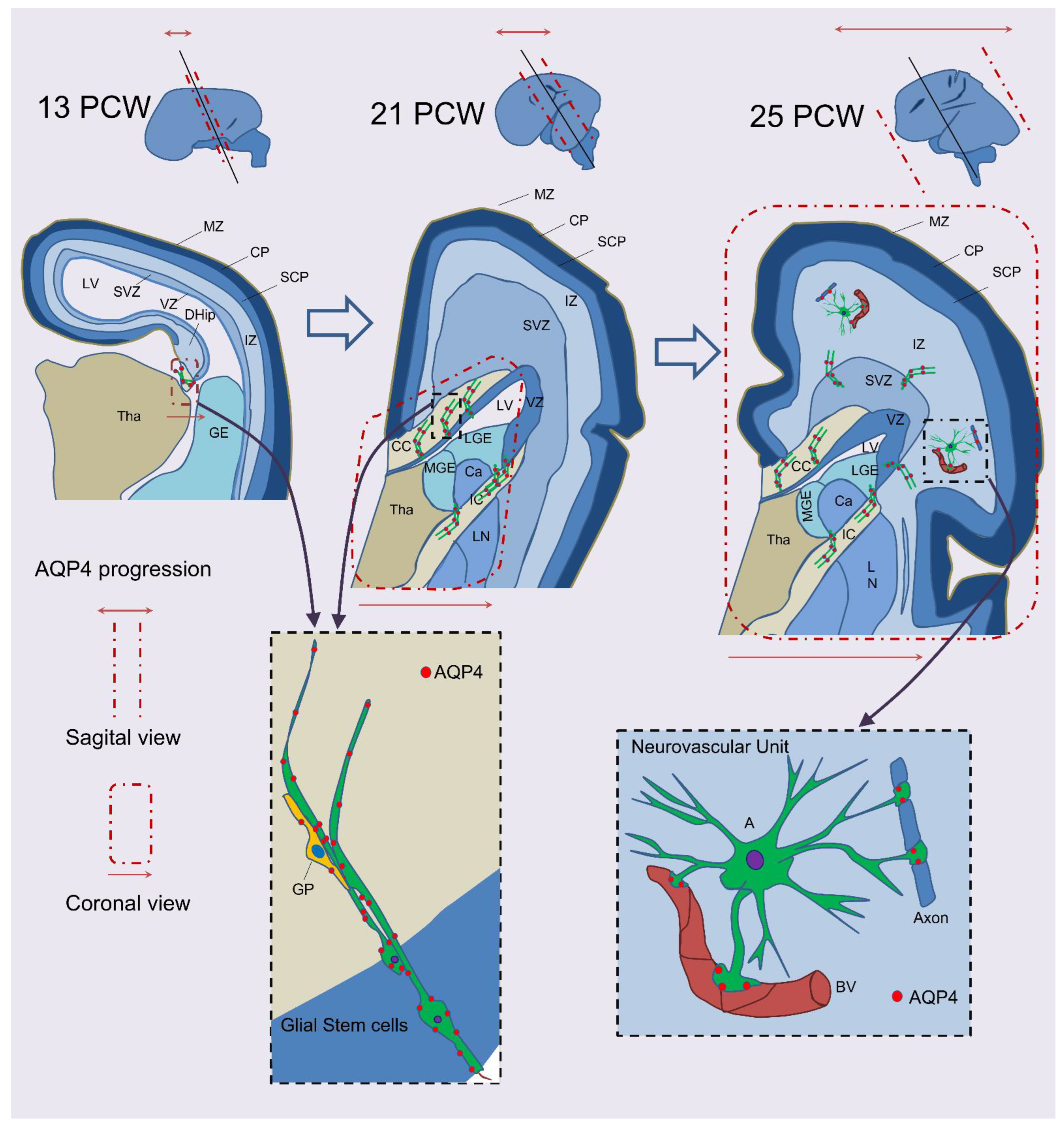{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. AQP4: A Possible Astrogenesis Marker
1.2. AQP4 in Pediatric Hydrocephalus: Neurodevelopmental Implications
2. Future Work
3. Search Strategy and Selection Criteria
4. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herculano-Houzel, S. The glia/neuron ratio: How it varies uniformly across brain structures and species and what that means for brain physiology and evolution. Glia 2014, 62, 1377–1391. [Google Scholar] [CrossRef] [PubMed]
- Morel, L.; Chiang, M.S.R.; Higashimori, H.; Shoneye, T.; Iyer, L.K.; Yelick, J.; Tai, A.; Yang, Y. Molecular and Functional Properties of Regional Astrocytes in the Adult Brain. J. Neurosci. 2017, 37, 8706–8717. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron 2017, 95, 531–549.e9. [Google Scholar] [CrossRef] [PubMed]
- John Lin, C.-C.; Yu, K.; Hatcher, A.; Huang, T.-W.; Lee, H.K.; Carlson, J.; Weston, M.C.; Chen, F.; Zhang, Y.; Zhu, W.; et al. Identification of diverse astrocyte populations and their malignant analogs. Nat. Neurosci. 2017, 20, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, E.S.; Huang, A.Y.-S.; Deneen, B. Astrocytogenesis: Where, when, and how. F1000Research 2020, 9, 233. [Google Scholar] [CrossRef]
- López-Muñoz, F.; Boya, J.; Alamo, C. Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramón y Cajal. Brain Res. Bull. 2006, 70, 391–405. [Google Scholar] [CrossRef]
- Zeisel, A.; Hochgerner, H.; Lönnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Häring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular Architecture of the Mouse Nervous System. Cell 2018, 174, 999–1014.e22. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.; Macosko, E.Z.; Wysoker, A.; Goldman, M.; Krienen, F.M.; de Rivera, H.; Bien, E.; Baum, M.; Bortolin, L.; Wang, S.; et al. Molecular Diversity and Specializations among the Cells of the Adult Mouse Brain. Cell 2018, 174, 1015–1030.e16. [Google Scholar] [CrossRef] [PubMed]
- Batiuk, M.Y.; Martirosyan, A.; Wahis, J.; de Vin, F.; Marneffe, C.; Kusserow, C.; Koeppen, J.; Viana, J.F.; Oliveira, J.F.; Voet, T.; et al. Identification of region-specific astrocyte subtypes at single cell resolution. Nat. Commun. 2020, 11, 1220. [Google Scholar] [CrossRef]
- Molofsky, A.V.; Krenick, R.; Ullian, E.; Tsai, H.-H.; Deneen, B.; Richardson, W.D.; Barres, B.A.; Rowitch, D.H. Astrocytes and disease: A neurodevelopmental perspective. Genes Dev. 2012, 26, 891–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarius, S.; Paul, F.; Weinshenker, B.G.; Levy, M.; Kim, H.J.; Wildemann, B. Neuromyelitis Optica. Nat. Rev. Dis. Primers 2020, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Gu, F.; Fu, L.; Ma, Y.-J. Aquaporin-4 in glioma invasion and an analysis of molecular mechanisms. J. Clin. Neurosci. 2010, 17, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.-L.; Wang, X.; Lou, J.-C.; Ma, X.-C.; Zhang, B. The potential roles of aquaporin 4 in malignant gliomas. Oncotarget 2017, 8, 32345–32355. [Google Scholar] [CrossRef]
- Molofsky, A.V.; Deneen, B. Astrocyte development: A Guide for the Perplexed. Glia 2015, 63, 1320–1329. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; King, L.S.; Christensen, B.M.; Agre, P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am. J. Physiol. 1997, 273, C1549–C1561. [Google Scholar] [CrossRef]
- Rash, J.E.; Yasumura, T.; Hudson, C.S.; Agre, P.; Nielsen, S. Direct immunogold labeling of aquaporin-4 in square arrays of astrocyte and ependymocyte plasma membranes in rat brain and spinal cord. Proc. Natl. Acad. Sci. USA 1998, 95, 11981–11986. [Google Scholar] [CrossRef]
- Cavazzin, C.; Ferrari, D.; Facchetti, F.; Russignan, A.; Vescovi, A.L.; La Porta, C.A.; Gritti, A. Unique expression and localization of aquaporin- 4 and aquaporin-9 in murine and human neural stem cells and in their glial progeny. Glia 2006, 53, 167–181. [Google Scholar] [CrossRef]
- Kong, H.; Fan, Y.; Xie, J.; Ding, J.; Sha, L.; Shi, X.; Sun, X.; Hu, G. AQP4 knockout impairs proliferation, migration and neuronal differentiation of adult neural stem cells. J. Cell Sci. 2008, 121, 4029–4036. [Google Scholar] [CrossRef]
- La Porta, C.A.; Gena, P.; Gritti, A.; Fascio, U.; Svelto, M.; Calamita, G. Adult murine CNS stem cells express aquaporin channels. Biol. Cell 2006, 98, 89–94. [Google Scholar] [CrossRef]
- Liang, R.; Yong, S.; Huang, X.; Kong, H.; Hu, G.; Fan, Y. Aquaporin-4 Mediates the Suppressive Effect of Lipopolysaccharide on Hippocampal Neurogenesis. Neuroimmunomodulation 2016, 23, 309–317. [Google Scholar] [CrossRef]
- Mack, A.F.; Künzle, H.; Lange, M.; Mages, B.; Reichenbach, A.; Härtig, W. Radial glial elements in the cerebral cortex of the lesser hedgehog tenrec. Brain Struct. Funct. 2018, 223, 3909–3917. [Google Scholar] [CrossRef] [PubMed]
- Gleiser, C.; Wagner, A.; Fallier-Becker, P.; Wolburg, H.; Hirt, B.; Mack, A.F. Aquaporin-4 in Astroglial Cells in the CNS and Supporting Cells of Sensory Organs—A Comparative Perspective. Int. J. Mol. Sci. 2016, 17, 1411. [Google Scholar] [CrossRef] [PubMed]
- Falcone, C.; Penna, E.; Hong, T.; Tarantal, A.F.; Hof, P.R.; Hopkins, W.D.; Sherwood, C.C.; Noctor, S.C.; Martínez-Cerdeño, V. Cortical Interlaminar Astrocytes Are Generated Prenatally, Mature Postnatally, and Express Unique Markers in Human and Nonhuman Primates. Cereb. Cortex 2021, 31, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Holst, C.B.; Brøchner, C.B.; Vitting-Seerup, K.; Møllgård, K. Astrogliogenesis in human fetal brain: Complex spatiotemporal immunoreactivity patterns of GFAP, S100, AQP 4 and YKL -40. J. Anat. 2019, 235, 590–615. [Google Scholar] [CrossRef]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Pardo, M.R.; Baz-Davila, R.; Lee, S.; Muhonen, M.; Borges, R.; Castañeyra-Perdomo, A. AQP4 labels a subpopulation of white matter-dependent glial radial cells affected by pediatric hydrocephalus, and its expression increased in glial microvesicles released to the cerebrospinal fluid in obstructive hydrocephalus. Acta Neuropathol. Commun. 2022, 10, 41. [Google Scholar] [CrossRef]
- Fallier-Becker, P.; Vollmer, J.P.; Bauer, H.; Noell, S.; Wolburg, H.; Mack, A.F. Onset of aquaporin-4 expression in the developing mouse brain. Int. J. Dev. Neurosci. 2014, 36, 81–89. [Google Scholar] [CrossRef]
- Mack, A.F.; Wolburg, H. A Novel Look at Astrocytes: Aquaporins, ionic homeostasis, and the role of the microenvironment for regeneration in the CNS. Neuroscientist 2013, 19, 195–207. [Google Scholar] [CrossRef]
- Gömöri, E.; Pál, J.; Ábrahám, H.; Vajda, Z.; Sulyok, E.; Seress, L.; Dóczi, T. Fetal development of membrane water channel proteins aquaporin-1 and aquaporin-4 in the human brain. Int. J. Dev. Neurosci. 2006, 24, 295–305. [Google Scholar] [CrossRef]
- Deoni, S.C.L.; Mercure, E.; Blasi, A.; Gasston, D.; Thomson, A.; Johnson, M.; Williams, S.; Murphy, D. Mapping Infant Brain Myelination with Magnetic Resonance Imaging. J. Neurosci. 2011, 31, 784–791. [Google Scholar] [CrossRef]
- Kier, E.L.; Truwit, C.L. The normal and abnormal genu of the corpus callosum: An evolutionary, embryologic, anatomic, and MR analysis. AJNR Am. J. Neuroradiol. 1996, 17, 1631–1641. [Google Scholar]
- Huang, H.; Xue, R.; Zhang, J.; Ren, T.; Richards, L.J.; Yarowsky, P.; Miller, M.I.; Mori, S. Anatomical Characterization of Human Fetal Brain Development with Diffusion Tensor Magnetic Resonance Imaging. J. Neurosci. 2009, 29, 4263–4273. [Google Scholar] [CrossRef] [PubMed]
- Barkovich, A.J.; Lyon, G.; Evrard, P. Formation, maturation, and disorders of white matter. AJNR Am. J. Neuroradiol. 1992, 13, 447–461. [Google Scholar] [PubMed]
- Paul, L.K. Developmental malformation of the corpus callosum: A review of typical callosal development and examples of developmental disorders with callosal involvement. J. Neurodev. Disord. 2011, 3, 3–27. [Google Scholar] [CrossRef]
- Rakic, P.; Yakovlev, P.I. Development of the corpus callosum and cavum septi in man. J. Comp. Neurol. 1968, 132, 45–72. [Google Scholar] [CrossRef] [PubMed]
- Luders, E.; Thompson, P.; Toga, A.W. The Development of the Corpus Callosum in the Healthy Human Brain. J. Neurosci. 2010, 30, 10985–10990. [Google Scholar] [CrossRef]
- Anthony, T.E.; Klein, C.; Fishell, G.; Heintz, N. Radial Glia Serve as Neuronal Progenitors in All Regions of the Central Nervous System. Neuron 2004, 41, 881–890. [Google Scholar] [CrossRef]
- Owada, Y.; Yoshimoto, T.; Kondo, H. Spatio-temporally differential expression of genes for three members of fatty acid binding proteins in developing and mature rat brains. J. Chem. Neuroanat. 1996, 12, 113–122. [Google Scholar] [CrossRef]
- Pringle, N.P.; Yu, W.-P.; Howell, M.; Colvin, J.S.; Ornitz, D.M.; Richardson, W.D. Fgfr3 expression by astrocytes and their precursors: Evidence that astrocytes and oligodendrocytes originate in distinct neuroepithelial domains. Development 2003, 130, 93–102. [Google Scholar] [CrossRef]
- Shibata, T.; Yamada, K.; Watanabe, M.; Ikenaka, K.; Wada, K.; Tanaka, K.; Inoue, Y. Glutamate Transporter GLAST Is Expressed in the Radial Glia–Astrocyte Lineage of Developing Mouse Spinal Cord. J. Neurosci. 1997, 17, 9212–9219. [Google Scholar] [CrossRef]
- Rekate, H.L. A Contemporary Definition and Classification of Hydrocephalus. Semin. Pediatr. Neurol. 2009, 16, 9–15. [Google Scholar] [CrossRef]
- Kahle, K.T.; Kulkarni, A.V.; Limbrick, D.D., Jr.; Warf, B.C. Hydrocephalus in children. Lancet 2016, 387, 788–799. [Google Scholar] [CrossRef]
- Karimy, J.K.; Zhang, J.; Kurland, D.B.; Theriault, B.C.; Duran, D.; Stokum, J.A.; Furey, C.G.; Zhou, X.; Mansuri, M.S.; Montejo, J.; et al. Inflammation-dependent cerebrospinal fluid hypersecretion by the choroid plexus epithelium in posthemorrhagic hydrocephalus. Nat. Med. 2017, 23, 997–1003. [Google Scholar] [CrossRef]
- Karimy, J.K.; Reeves, B.C.; Damisah, E.; Duy, P.Q.; Antwi, P.; David, W.; Wang, K.; Schiff, S.J.; Limbrick, D.D., Jr.; Alper, S.L.; et al. Inflammation in acquired hydrocephalus: Pathogenic mechanisms and therapeutic targets. Nat. Rev. Neurol. 2020, 16, 285–296. [Google Scholar] [CrossRef]
- Harris, C.A.; Morales, D.M.; Arshad, R.; McAllister, J.P., II; Limbrick, D.D., Jr. Cerebrospinal fluid biomarkers of neuroinflammation in children with hydrocephalus and shunt malfunction. Fluids Barriers CNS 2021, 18, 4. [Google Scholar] [CrossRef]
- Garcia-Bonilla, M.; Castaneyra-Ruiz, L.; Zwick, S.; Talcott, M.; Otun, A.; Isaacs, A.M.; Morales, D.M.; Limbrick, D.D.; McAllister, J.P. Acquired hydrocephalus is associated with neuroinflammation, progenitor loss, and cellular changes in the subventricular zone and periventricular white matter. Fluids Barriers CNS 2022, 19, 17. [Google Scholar] [CrossRef]
- Isaacs, A.M.; Neil, J.J.; McAllister, J.P.; Dahiya, S.; Castaneyra-Ruiz, L.; Merisaari, H.; Botteron, H.E.; Alexopoulous, D.; George, A.; Sun, P.; et al. Microstructural Periventricular White Matter Injury in Post-hemorrhagic Ventricular Dilatation. Neurology 2021, 98, e364–e375. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, T.; Wu, G.; McBride, D.W.; Xu, N.; Klebe, D.W.; Zhang, Y.; Li, Q.; Tang, J.; Zhang, J.H. Astrogliosis inhibition attenuates hydrocephalus by increasing cerebrospinal fluid reabsorption through the glymphatic system after germinal matrix hemorrhage. Exp. Neurol. 2019, 320, 113003. [Google Scholar] [CrossRef]
- Eide, P.K.; Hansson, H.-A. Astrogliosis and impaired aquaporin-4 and dystrophin systems in idiopathic normal pressure hydrocephalus. Neuropathol. Appl. Neurobiol. 2018, 44, 474–490. [Google Scholar] [CrossRef]
- Garcia, C.A.B.; Catalão, C.H.R.; Machado, H.; Júnior, I.M.; Romeiro, T.H.; Peixoto-Santos, J.E.; Santos, M.V.; Lopes, L.D.S. Edaravone reduces astrogliosis and apoptosis in young rats with kaolin-induced hydrocephalus. Child’s Nerv. Syst. 2016, 33, 419–428. [Google Scholar] [CrossRef]
- Miller, J.M.; McAllister, J.P., II. Reduction of astrogliosis and microgliosis by cerebrospinal fluid shunting in experimental hydrocephalus. Cereb. Fluid Res. 2007, 4, 10–186. [Google Scholar] [CrossRef]
- Suryaningtyas, W.; Parenrengi, M.A.; Bajamal, A.H.; Rantam, F.A. Lipid Peroxidation Induces Reactive Astrogliosis by Activating WNT/β-Catenin Pathway in Hydrocephalus. Malays. J. Med. Sci. 2020, 27, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Tida, J.A.; Catalão, C.H.R.; Garcia, C.A.B.; dos Santos, A.C.; Salmon, C.E.G.; Lopes, L.D.S. Acupuncture at ST36 Exerts Neuroprotective Effects via Inhibition of Reactive Astrogliosis in Infantile Rats with Hydrocephalus. Acupunct. Med. 2018, 36, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Deren, K.E.; Packer, M.; Forsyth, J.; Milash, B.; Abdullah, O.M.; Hsu, E.W.; McAllister, J.P. Reactive astrocytosis, microgliosis and inflammation in rats with neonatal hydrocephalus. Exp. Neurol. 2010, 226, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Roales-Buján, R.; Páez, P.; Guerra, M.; Rodríguez, S.; Vío, K.; Ho-Plagaro, A.; García-Bonilla, M.; Rodríguez-Pérez, L.-M.; Domínguez-Pinos, M.-D.; Rodríguez, E.-M.; et al. Astrocytes acquire morphological and functional characteristics of ependymal cells following disruption of ependyma in hydrocephalus. Acta Neuropathol. 2012, 124, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Del Bigio, M.R. Neuropathology and structural changes in hydrocephalus. Dev. Disabil. Res. Rev. 2010, 16, 16–22. [Google Scholar] [CrossRef]
- Jin, S.C.; Dong, W.; Kundishora, A.J.; Panchagnula, S.; Moreno-De-Luca, A.; Furey, C.G.; Allocco, A.A.; Walker, R.L.; Nelson-Williams, C.; Smith, H.; et al. Exome sequencing implicates genetic disruption of prenatal neuro-gliogenesis in sporadic congenital hydrocephalus. Nat. Med. 2020, 26, 1754–1765. [Google Scholar] [CrossRef]
- Guerra, M.M.; Henzi, R.; Ortloff, A.; Lichtin, N.; Vío, K.; Jiménez, A.J.; Dominguez-Pinos, M.D.; González, C.; Jara, M.C.; Hinostroza, F.; et al. Cell Junction Pathology of Neural Stem Cells Is Associated With Ventricular Zone Disruption, Hydrocephalus, and Abnormal Neurogenesis. J. Neuropathol. Exp. Neurol. 2015, 74, 653–671. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Guerra, M.M.; Vío, K.; González, C.; Ortloff, A.; Bátiz, L.F.; Rodríguez, S.; Jara, M.C.; Muñoz, R.I.; Ortega, E.; et al. A cell junction pathology of neural stem cells leads to abnormal neurogenesis and hydrocephalus. Biol. Res. 2012, 45, 231–241. [Google Scholar] [CrossRef]
- Domínguez-Pinos, M.D.; Páez, P.; Jiménez, A.-J.; Weil, B.; Arráez, M.-A.; Pérez-Fígares, J.-M.; Rodríguez, E.-M. Ependymal Denudation and Alterations of the Subventricular Zone Occur in Human Fetuses With a Moderate Communicating Hydrocephalus. J. Neuropathol. Exp. Neurol. 2005, 64, 595–604. [Google Scholar] [CrossRef]
- McAllister, J.P.; Guerra, M.M.; Ruiz, L.C.; Jimenez, A.J.; Dominguez-Pinos, D.; Sival, D.; Dunnen, W.D.; Morales, D.; Schmidt, R.E.; Rodríguez, E.; et al. Ventricular Zone Disruption in Human Neonates with Intraventricular Hemorrhage. J. Neuropathol. Exp. Neurol. 2017, 76, 358–375. [Google Scholar] [CrossRef]
- Fernández-Muñoz, B.; Rosell-Valle, C.; Ferrari, D.; Alba-Amador, J.; Montiel, M.; Campos-Cuerva, R.; Lopez-Navas, L.; Muñoz-Escalona, M.; Martín-López, M.; Profico, D.C.; et al. Retrieval of germinal zone neural stem cells from the cerebrospinal fluid of premature infants with intraventricular hemorrhage. Stem Cells Transl. Med. 2020, 9, 1085–1101. [Google Scholar] [CrossRef] [PubMed]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Meyer, G.; Castañeyra-Perdomo, A. A Distal to Proximal Gradient of Human Choroid Plexus Development, with Antagonistic Expression of Glut1 and AQP1 in Mature Cells vs. Calbindin and PCNA in Proliferative Cells. Front. Neuroanat. 2016, 10, 87. [Google Scholar] [CrossRef] [PubMed]
- Castañeyra-Ruiz, L.; González-Marrero, I.; González-Toledo, J.M.; Castañeyra-Ruiz, A.; de Paz-Carmona, H.; Castañeyra-Perdomo, A.; Carmona-Calero, E.M. Aquaporin-4 expression in the cerebrospinal fluid in congenital human hydrocephalus. Fluids Barriers CNS 2013, 10, 18. [Google Scholar] [CrossRef]
- Limbrick, D.D., Jr.; Castaneyra-Ruiz, L.; Han, R.H.; Berger, D.; McAllister, J.P.; Morales, D.M. Cerebrospinal Fluid Biomarkers of Pediatric Hydrocephalus. Pediatr. Neurosurg. 2017, 52, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Carmona-Calero, E.M.; Abreu-Gonzalez, P.; Lecuona, M.; Brage, L.; Rodríguez, E.M.; Castañeyra-Perdomo, A. Cerebrospinal fluid levels of tumor necrosis factor alpha and aquaporin 1 in patients with mild cognitive impairment and idiopathic normal pressure hydrocephalus. Clin. Neurol. Neurosurg. 2016, 146, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Castañeyra-Ruiz, L.; Hernández-Abad, L.G.; Carmona-Calero, E.M.; Castañeyra-Perdomo, A.; González-Marrero, I. AQP1 Overexpression in the CSF of Obstructive Hydrocephalus and Inversion of Its Polarity in the Choroid Plexus of a Chiari Malformation Type II Case. J. Neuropathol. Exp. Neurol. 2019, 78, 641–647. [Google Scholar] [CrossRef]
- Jeon, T.; Park, K.-S.; Park, S.-H.; Hwang, J.-H.; Hwang, S.K. Expression of Aquaporin 1 and 4 in the Choroid Plexus and Brain Parenchyma of Kaolin-Induced Hydrocephalic Rats. Korean J. Neurotrauma 2017, 13, 68–75. [Google Scholar] [CrossRef]
- Kim, J.; Jung, Y. Increased aquaporin-1 and Na+-K+-2Cl- cotransporter 1 expression in choroid plexus leads to blood-cerebrospinal fluid barrier disruption and necrosis of hippocampal CA1 cells in acute rat models of hyponatremia. J. Neurosci. Res. 2012, 90, 1437–1444. [Google Scholar] [CrossRef]
- Paul, L.; Madan, M.; Rammling, M.; Chigurupati, S.; Chan, S.L.; Pattisapu, J.V. Expression of Aquaporin 1 and 4 in a Congenital Hydrocephalus Rat Model. Neurosurgery 2011, 68, 462–473. [Google Scholar] [CrossRef]
- Carmona-Calero, E.M.; Gonzalez-Toledo, J.M.; Castañeyra-Ruiz, L.; González-Marrero, I.; Castañeyra-Ruiz, M.; Rancel-Torres, N.; Castañeyra-Ruiz, A.; Castañeyra-Perdomo, A. Expression of aquaporin-1 in the choroid plexus in communicating and non-communicating hydrocephalic rats. Eur. J. Anat. 2015, 19, 125–129. [Google Scholar]
- Castañeyra-Ruiz, L.; González-Marrero, I.; Castañeyra-Ruiz, M.; González-Toledo, J.M.; Carmona-Calero, E.M. Los canales de agua. Acuaporinas 1 y 4 en el sistema nervioso central y su relación con la hidrocefalia. Majorensis 2014, 10, 1–6. [Google Scholar]
- Küppers, E.; Gleiser, C.; Brito, V.; Wachter, B.; Pauly, T.; Hirt, B.; Grissmer, S. AQP4 expression in striatal primary cultures is regulated by dopamine—Implications for proliferation of astrocytes. Eur. J. Neurosci. 2008, 28, 2173–2182. [Google Scholar] [CrossRef] [PubMed]
- Oria, M.; Pathak, B.; Li, Z.; Bakri, K.; Gouwens, K.; Varela, M.F.; Lampe, K.; Murphy, K.P.; Lin, C.-Y.; Peiro, J.L. Premature Neural Progenitor Cell Differentiation Into Astrocytes in Retinoic Acid-Induced Spina Bifida Rat Model. Front. Mol. Neurosci. 2022, 15, 888351. [Google Scholar] [CrossRef] [PubMed]
- Tully, H.M.; Dobyns, W.B. Infantile hydrocephalus: A review of epidemiology, classification and causes. Eur. J. Med. Genet. 2014, 57, 359–368. [Google Scholar] [CrossRef]
- Demyanenko, G.P.; Tsai, A.Y.; Maness, P.F. Abnormalities in Neuronal Process Extension, Hippocampal Development, and the Ventricular System of L1 Knockout Mice. J. Neurosci. 1999, 19, 4907–4920. [Google Scholar] [CrossRef]
- Hannay, H.J. Functioning of the corpus callosum in children with early hydrocephalus. J. Int. Neuropsychol. Soc. 2000, 6, 351–361. [Google Scholar] [CrossRef]
- Castaneyra-Ruiz, L.; Morales, M.D.M.; McAllister, J.P.; Brody, S.L.; Isaacs, A.M.; Strahle, J.M.; Dahiya, S.M.; Limbrick, D.D. Blood Exposure Causes Ventricular Zone Disruption and Glial Activation In Vitro. J. Neuropathol. Exp. Neurol. 2018, 77, 803–813. [Google Scholar] [CrossRef]
- Castaneyra-Ruiz, L.; McAllister, J.P., II; Morales, D.M.; Brody, S.L.; Isaacs, A.M.; Limbrick, D.D. Preterm intraventricular hemorrhage in vitro: Modeling the cytopathology of the ventricular zone. Fluids Barriers CNS 2020, 17, 46. [Google Scholar] [CrossRef]
- Dong, Y.; Zhong, J.; Dong, L. The Role of Decorin in Autoimmune and Inflammatory Diseases. J. Immunol. Res. 2022, 2022, 1283383. [Google Scholar] [CrossRef]
- Botfield, H.; Gonzalez, A.M.; Abdullah, O.; Skjolding, A.D.; Berry, M.; McAllister, J.P.; Logan, A. Decorin prevents the development of juvenile communicating hydrocephalus. Brain 2013, 136, 2842–2858. [Google Scholar] [CrossRef]
- Aojula, A.; Botfield, H.; McAllister, J.P., II; Gonzalez, A.M.; Abdullah, O.; Logan, A.; Sinclair, A. Diffusion tensor imaging with direct cytopathological validation: Characterisation of decorin treatment in experimental juvenile communicating hydrocephalus. Fluids Barriers CNS 2016, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- McAllister, J.P.; Aojula, A.; Botfield, H.; Abdullah, O.; Gonzalez, A.M.; Ragan, D.; Logan, A.; Sinclair, A. Decorin reduces white matter pathology in experimental hydrocephalus: A diffusion tensor imaging and immunohistochemical study. Fluids Barriers CNS 2015, 12, O59. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 74, 790–799. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castañeyra-Ruiz, L.; González-Marrero, I.; Hernández-Abad, L.G.; Lee, S.; Castañeyra-Perdomo, A.; Muhonen, M. AQP4, Astrogenesis, and Hydrocephalus: A New Neurological Perspective. Int. J. Mol. Sci. 2022, 23, 10438. https://doi.org/10.3390/ijms231810438
Castañeyra-Ruiz L, González-Marrero I, Hernández-Abad LG, Lee S, Castañeyra-Perdomo A, Muhonen M. AQP4, Astrogenesis, and Hydrocephalus: A New Neurological Perspective. International Journal of Molecular Sciences. 2022; 23(18):10438. https://doi.org/10.3390/ijms231810438
Chicago/Turabian StyleCastañeyra-Ruiz, Leandro, Ibrahim González-Marrero, Luis G. Hernández-Abad, Seunghyun Lee, Agustín Castañeyra-Perdomo, and Michael Muhonen. 2022. "AQP4, Astrogenesis, and Hydrocephalus: A New Neurological Perspective" International Journal of Molecular Sciences 23, no. 18: 10438. https://doi.org/10.3390/ijms231810438
APA StyleCastañeyra-Ruiz, L., González-Marrero, I., Hernández-Abad, L. G., Lee, S., Castañeyra-Perdomo, A., & Muhonen, M. (2022). AQP4, Astrogenesis, and Hydrocephalus: A New Neurological Perspective. International Journal of Molecular Sciences, 23(18), 10438. https://doi.org/10.3390/ijms231810438