
Circular RNAs Involved in the Regulation of the Age-Related Pathways

 , , and
, , and
Abstract
:1. Introduction
2. Biogenesis and Functional Mechanisms of circRNAs
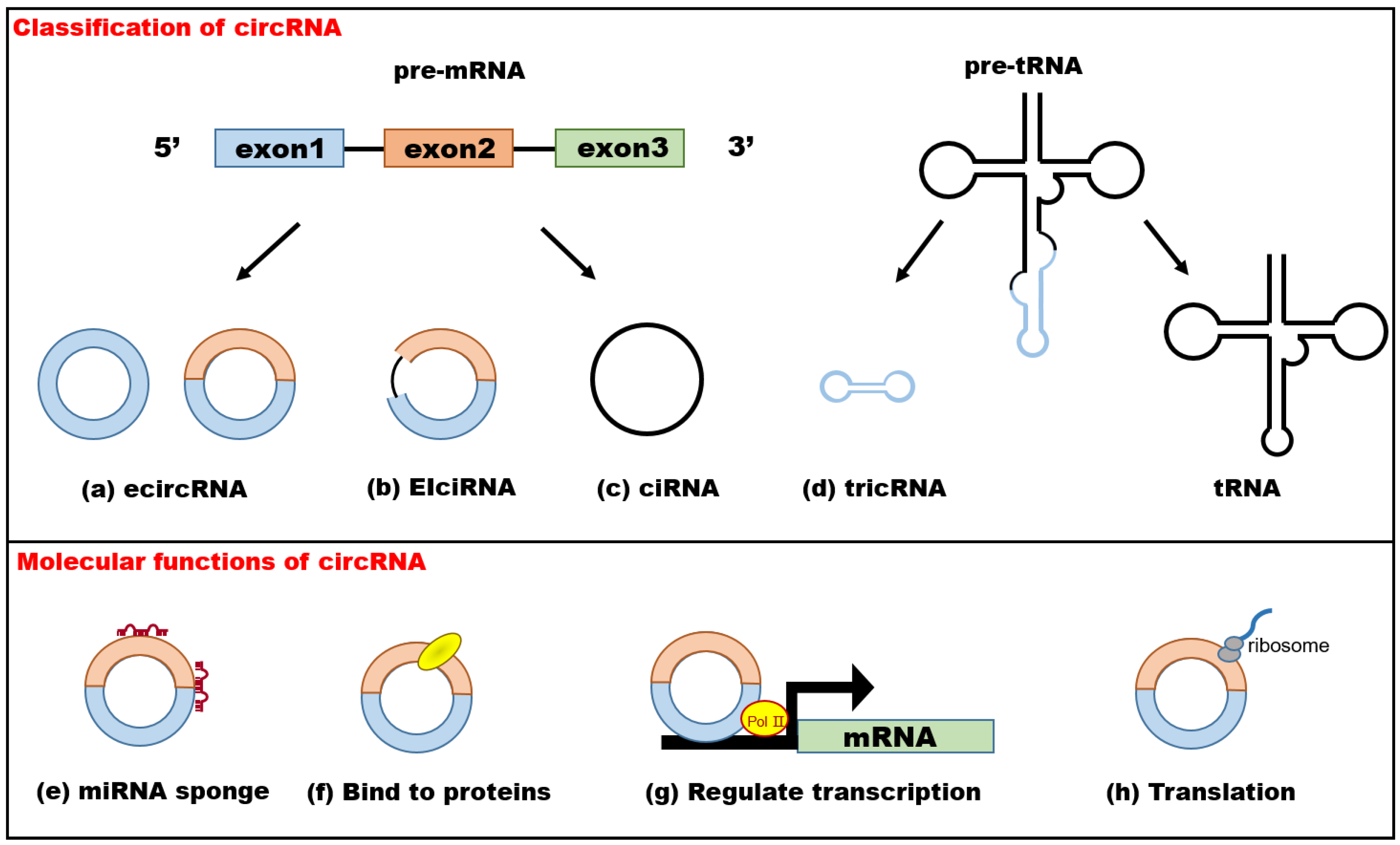
2.1. Classification of circRNAs
2.2. Functional Mechanisms of circRNAs
3. Age-Related Signaling Pathways
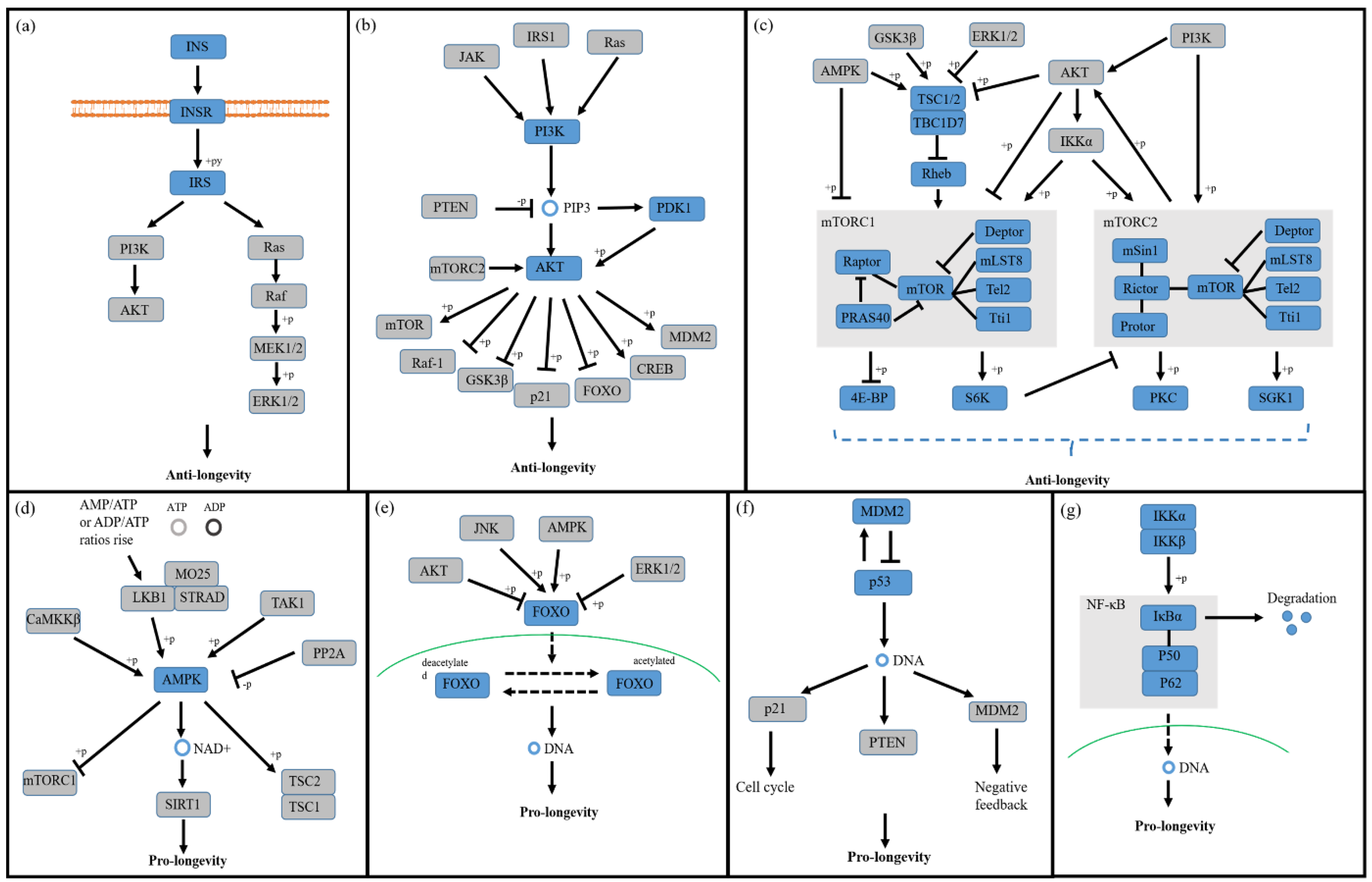
3.1. Insulin/Insulin-like Growth Factor-1 Signaling (IIS) Pathway
3.2. PI3K/Akt Signaling Pathway
3.3. mTOR Signaling Pathway
3.4. AMPK Signaling Pathway
3.5. FOXO Signaling Pathway
3.6. p53 Signaling Pathway
3.7. NF-κB Signaling Pathway
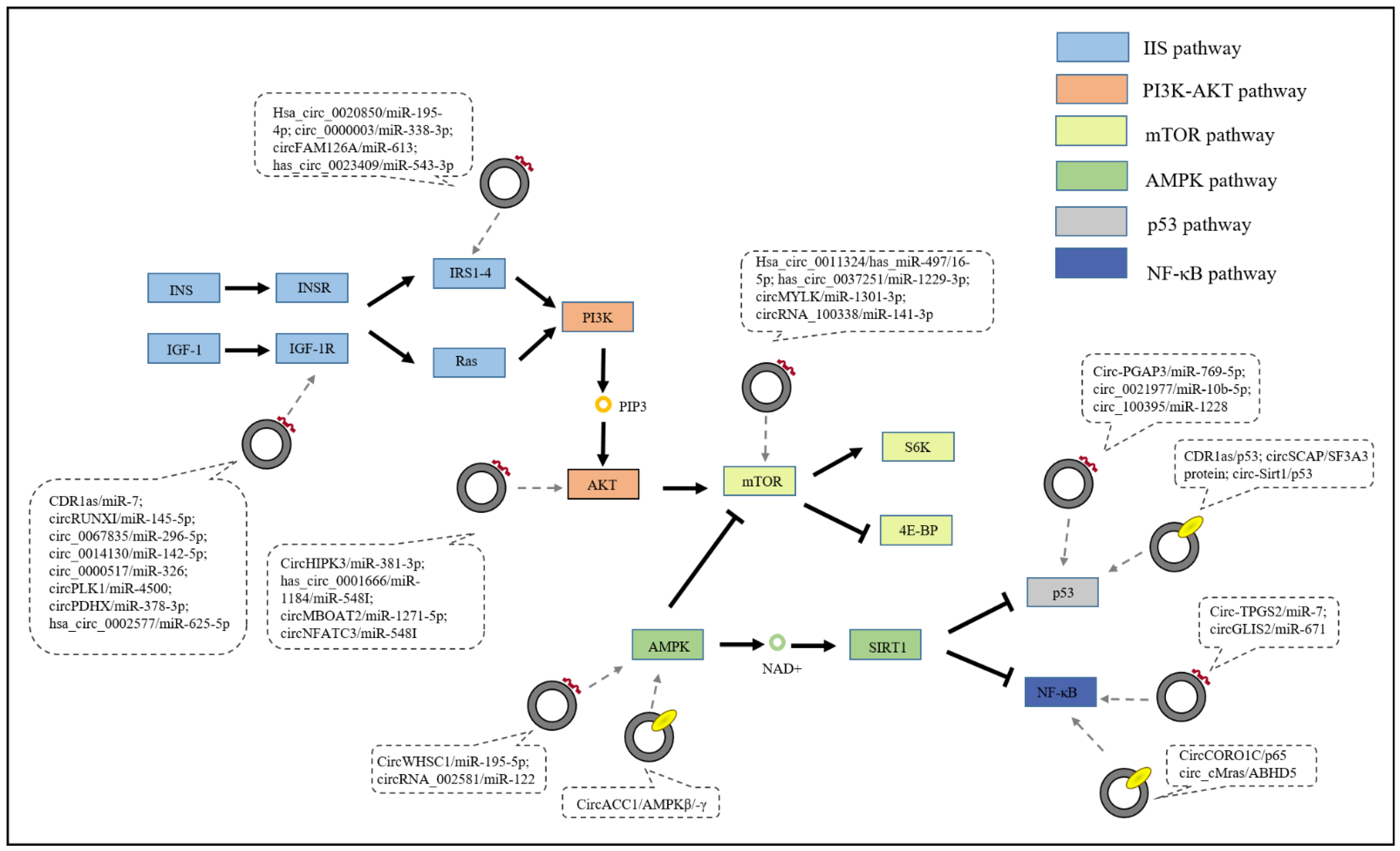
4. CircRNAs in Age-Related Signaling Pathways
4.1. IIS Pathway
4.1.1. CDR1as
4.1.2. Hsa_circ_0020850
4.1.3. Hsa_circ_0023409
4.2. PI3K/AKT Signaling Pathway
4.2.1. Pancreatic Cancer
4.2.2. Glioma
4.2.3. Gastric Cancer
4.2.4. Non-Small Cell Lung Cancer
4.2.5. Colorectal Cancer
4.2.6. Breast Cancer
4.2.7. Other Cancers
4.3. mTOR Signaling Pathway
4.3.1. AKT-mTOR Axis
4.3.2. AMPK-mTOR Axis
4.3.3. Targeting mTOR
4.4. AMPK Signaling Pathway
4.5. p53 Signaling Pathway
4.5.1. Circ-PGAP3
4.5.2. Circ-Sirt1
4.5.3. CircSCAP
4.5.4. Circ-MDM2
4.6. NF-κB Signaling Pathway
5. Summary and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Meyer, K.; Yankner, B.A. Slowing Down Aging. Cell Metab. 2017, 26, 592–593. [Google Scholar] [CrossRef]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef]
- Green, C.L.; Lamming, D.W.; Fontana, L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat. Rev. Mol. Cell Biol. 2022, 23, 56–73. [Google Scholar] [CrossRef]
- Suo, M.; Rommelfanger, M.K.; Chen, Y.; Amro, E.M.; Han, B.; Chen, Z.; Szafranski, K.; Chakkarappan, S.R.; Boehm, B.O.; MacLean, A.L.; et al. Age-dependent effects of Igf2bp2 on gene regulation, function, and aging of hematopoietic stem cells in mice. Blood 2022, 139, 2653–2665. [Google Scholar] [CrossRef]
- Dolati, S.; Shakouri, S.K.; Dolatkhah, N.; Yousefi, M.; Jadidi-Niaragh, F.; Sanaie, S. The role of exosomal non-coding RNAs in aging-related diseases. Biofactors 2021, 47, 292–310. [Google Scholar] [CrossRef]
- Cai, J.; Qi, H.; Yao, K.; Yao, Y.; Jing, D.; Liao, W.; Zhao, Z. Non-Coding RNAs Steering the Senescence-Related Progress, Properties, and Application of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 650431. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted LV, W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, present, and future of circRNAs. Embo J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zhang, Y.; Xiong, W.; Zhang, Z.; Wang, Z.; Lv, L.; Liu, C.; Hu, Z.; Zheng, Y.T.; Lu, L.; et al. CircGRIA1 shows an age-related increase in male macaque brain and regulates synaptic plasticity and synaptogenesis. Nat. Commun. 2020, 11, 3594. [Google Scholar] [CrossRef] [PubMed]
- Gruner, H.; Cortés-López, M.; Cooper, D.A.; Bauer, M.; Miura, P. CircRNA accumulation in the aging mouse brain. Sci. Rep. 2016, 6, 38907. [Google Scholar] [CrossRef] [PubMed]
- Cortés-López, M.; Gruner, M.R.; Cooper, D.A.; Gruner, H.N.; Voda, A.I.; van der Linden, A.M.; Miura, P. Global accumulation of circRNAs during aging in Caenorhabditis elegans. BMC Genom. 2018, 19, 8. [Google Scholar] [CrossRef]
- Westholm, J.O.; Miura, P.; Olson, S.; Shenker, S.; Joseph, B.; Sanfilippo, P.; Celniker, S.E.; Graveley, B.R.; Lai, E.C. Genome-wide analysis of drosophila circular RNAs reveals their structural and sequence properties and age-dependent neural accumulation. Cell Rep. 2014, 9, 1966–1980. [Google Scholar] [CrossRef] [PubMed]
- Weigelt, C.M.; Sehgal, R.; Tain, L.S.; Cheng, J.; Eßer, J.; Pahl, A.; Dieterich, C.; Grönke, S.; Partridge, L. An Insulin-Sensitive Circular RNA that Regulates Lifespan in Drosophila. Mol. Cell 2020, 79, 268–279.e5. [Google Scholar] [CrossRef]
- Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. Int. J. Mol. Sci. 2019, 20, 3926. [Google Scholar] [CrossRef]
- Yuan, X.; Yuan, Y.; He, Z.; Li, D.; Zeng, B.; Ni, Q.; Yang, M.; Yang, D. The Regulatory Functions of Circular RNAs in Digestive System Cancers. Cancers 2020, 12, 770. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Schmidt, C.A.; Matera, A.G. tRNA introns: Presence, processing, and purpose. Wiley Interdiscip Rev. RNA 2020, 11, e1583. [Google Scholar] [CrossRef]
- Zhang, D.; Ni, N.; Wang, Y.; Tang, Z.; Gao, H.; Ju, Y.; Sun, N.; He, X.; Gu, P.; Fan, X. CircRNA-vgll3 promotes osteogenic differentiation of adipose-derived mesenchymal stem cells via modulating miRNA-dependent integrin α5 expression. Cell Death Differ. 2021, 28, 283–302. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Guo, S.; Li, W.; Yu, P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci. Rep. 2015, 5, 12453. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, H.; Zhao, Y.; Zhao, Y. CDR1as is overexpressed in laryngeal squamous cell carcinoma to promote the tumour’s progression via miR-7 signals. Cell Prolif. 2018, 51, e12521. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.; Meng, L.; Sang, Y.; Liu, S.; Ding, P.; Ju, Y.; Liu, F.; Gu, L.; Lian, Y.; Li, J.; et al. Circular RNA ciRS-7 accelerates ESCC progression through acting as a miR-876-5p sponge to enhance MAGE-A family expression. Cancer Lett. 2018, 426, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Wang, L.; Long, H.; Zheng, Q.; Bo, X.; Xiao, X.; Li, B. Circular RNA circRHOT1 promotes hepatocellular carcinoma progression by initiation of NR2F6 expression. Mol. Cancer 2019, 18, 119. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z. Efficient backsplicing produces translatable circular mRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, M.; Bonafè, M.; Franceschi, C.; Paolisso, G. Insulin/IGF-I-signaling pathway: An evolutionarily conserved mechanism of longevity from yeast to humans. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1064–E1071. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Ogg, S.; Paradis, S.; Gottlieb, S.; Patterson, G.I.; Lee, L.; Tissenbaum, H.A.; Ruvkun, G. The Fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 1997, 389, 994–999. [Google Scholar] [CrossRef]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef]
- Junnila, R.K.; List, E.O.; Berryman, D.E.; Murrey, J.W.; Kopchick, J.J. The GH/IGF-1 axis in ageing and longevity. Nat. Rev. Endocrinol. 2013, 9, 366–376. [Google Scholar] [CrossRef]
- Slack, C.; Tullet, J. Signal Transduction Pathways in Ageing. Subcell. Biochem. 2018, 90, 323–350. [Google Scholar]
- Selman, C.; Lingard, S.; Choudhury, A.I.; Batterham, R.L.; Claret, M.; Clements, M.; Ramadani, F.; Okkenhaug, K.; Schuster, E.; Blanc, E.; et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. Faseb. J. 2008, 22, 807–818. [Google Scholar] [CrossRef]
- Taguchi, A.; Wartschow, L.M.; White, M.F. Brain IRS2 signaling coordinates life span and nutrient homeostasis. Science 2007, 317, 369–372. [Google Scholar] [CrossRef]
- White, M.F. IRS proteins and the common path to diabetes. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E413–E422. [Google Scholar] [CrossRef]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Qian, Y.; Sun, Z.; Shen, X.; Cai, Y.; Li, L.; Wang, Z. Role of PI3K in the Progression and Regression of Atherosclerosis. Front. Pharmacol. 2021, 12, 632378. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Heo, S.; Brzostowski, J.; Kang, D. Endosomal mTORC2 Is Required for Phosphoinositide-Dependent AKT Activation in Platelet-Derived Growth Factor-Stimulated Glioma Cells. Cancers 2021, 13, 2405. [Google Scholar] [CrossRef]
- Dieterle, A.M.; Böhler, P.; Keppeler, H.; Alers, S.; Berleth, N.; Drießen, S.; Hieke, N.; Pietkiewicz, S.; Löffler, A.S.; Peter, C.; et al. PDK1 controls upstream PI3K expression and PIP3 generation. Oncogene 2014, 33, 3043–3053. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef]
- Wei, Y.; Zhou, J.; Yu, H.; Jin, X. AKT phosphorylation sites of Ser473 and Thr308 regulate AKT degradation. Biosci. Biotechnol. Biochem. 2019, 83, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Noorolyai, S.; Shajari, N.; Baghbani, E.; Sadreddini, S.; Baradaran, B. The relation between PI3K/AKT signalling pathway and cancer. Gene 2019, 698, 120–128. [Google Scholar] [CrossRef]
- Wang, X.; Hills, L.B.; Huang, Y.H. Lipid and Protein Co-Regulation of PI3K Effectors Akt and Itk in Lymphocytes. Front. Immunol. 2015, 6, 117. [Google Scholar] [CrossRef]
- Nazari, N.; Jafari, F.; Ghalamfarsa, G.; Hadinia, A.; Atapour, A.; Ahmadi, M.; Dolati, S.; Rostamzadeh, D. The emerging role of microRNA in regulating the mTOR signaling pathway in immune and inflammatory responses. Immunol. Cell Biol. 2021, 99, 814–832. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef]
- Kaeberlein, M.; Powers, R.W., 3rd; Steffen, K.K.; Westman, E.A.; Hu, D.; Dang, N.; Kerr, E.O.; Kirkland, K.T.; Fields, S.; Kennedy, B.K. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science 2005, 310, 1193–1196. [Google Scholar] [CrossRef] [Green Version]
- Kapahi, P.; Zid, B.M.; Harper, T.; Koslover, D.; Sapin, V.; Benzer, S. Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. Curr. Biol. 2004, 14, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Vellai, T.; Takacs-Vellai, K.; Zhang, Y.; Kovacs, A.L.; Orosz, L.; Müller, F. Genetics: Influence of TOR kinase on lifespan in C. elegans. Nature 2003, 426, 620. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.E.; Strong, R.; Sharp, Z.D.; Nelson, J.F.; Astle, C.M.; Flurkey, K.; Nadon, N.L.; Wilkinson, J.E.; Frenkel, K.; Carter, C.S.; et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 2009, 460, 392–395. [Google Scholar] [CrossRef]
- Burkewitz, K.; Zhang, Y.; Mair, W.B. AMPK at the nexus of energetics and aging. Cell Metab. 2014, 20, 10–25. [Google Scholar] [CrossRef]
- Apfeld, J.; O’Connor, G.; McDonagh, T.; DiStefano, P.S.; Curtis, R. The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans. Genes Dev. 2004, 18, 3004–3009. [Google Scholar] [CrossRef] [PubMed]
- Tabibzadeh, S. Signaling pathways and effectors of aging. Front. Biosci. 2021, 26, 50–96. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Age-related changes in AMPK activation: Role for AMPK phosphatases and inhibitory phosphorylation by upstream signaling pathways. Ageing Res. Rev. 2016, 28, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef]
- Mair, W.; Morantte, I.; Rodrigues, A.P.; Manning, G.; Montminy, M.; Shaw, R.J.; Dillin, A. Lifespan extension induced by AMPK and calcineurin is mediated by CRTC-1 and CREB. Nature 2011, 470, 404–408. [Google Scholar] [CrossRef]
- Martins, R.; Lithgow, G.J.; Link, W. Long live FOXO: Unraveling the role of FOXO proteins in aging and longevity. Aging Cell 2016, 15, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Burgering, B.M.; Kops, G.J. Cell cycle and death control: Long live Forkheads. Trends. Biochem. Sci. 2002, 27, 352–360. [Google Scholar] [CrossRef]
- van der Horst, A.; Burgering, B.M. Stressing the role of FoxO proteins in lifespan and disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Brunet, A. FOXO transcription factors. Curr. Biol. 2007, 17, R113–R114. [Google Scholar] [CrossRef]
- Webb, A.E.; Brunet, A. FOXO flips the longevity SWItch. Nat. Cell Biol. 2013, 15, 444–446. [Google Scholar] [CrossRef]
- Sharpless, N.E.; DePinho, R.A. p53, good cop/bad cop. Cell 2002, 110, 9–12. [Google Scholar] [CrossRef]
- Bartas, M.; Brázda, V.; Volná, A.; Červeň, J.; Pečinka, P.; Zawacka-Pankau, J.E. The Changes in the p53 Protein across the Animal Kingdom Point to Its Involvement in Longevity. Int. J. Mol. Sci. 2021, 22, 8512. [Google Scholar] [CrossRef]
- Tavernarakis, N.; Pasparaki, A.; Tasdemir, E.; Maiuri, M.C.; Kroemer, G. The effects of p53 on whole organism longevity are mediated by autophagy. Autophagy 2008, 4, 870–873. [Google Scholar] [CrossRef]
- Mayo, L.D.; Donner, D.B. The PTEN, Mdm2, p53 tumor suppressor-oncoprotein network. Trends. Biochem. Sci. 2002, 27, 462–467. [Google Scholar] [CrossRef]
- Chène, P. Inhibiting the p53-MDM2 interaction: An important target for cancer therapy. Nat. Rev. Cancer 2003, 3, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.; Garg, M.; Tergaonkar, V.; Tan, S.Y.; Sethi, G. Pharmacological significance of the non-canonical NF-κB pathway in tumorigenesis. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188449. [Google Scholar] [CrossRef]
- Hyder, U.; McCann, J.L.; Wang, J.; Fung, V.; Bayo, J.; D’Orso, I. The ARF tumor suppressor targets PPM1G/PP2Cγ to counteract NF-κB transcription tuning cell survival and the inflammatory response. Proc. Natl. Acad. Sci. USA 2020, 117, 32594–32605. [Google Scholar] [CrossRef] [PubMed]
- Smale, S.T. Hierarchies of NF-κB target-gene regulation. Nat. Immunol. 2011, 12, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Diamant, G.; Dikstein, R. Transcriptional control by NF-κB: Elongation in focus. Biochim. Biophys. Acta 2013, 1829, 937–945. [Google Scholar] [CrossRef]
- Willcox, B.J.; Donlon, T.A.; He, Q.; Chen, R.; Grove, J.S.; Yano, K.; Masaki, K.H.; Willcox, D.C.; Rodriguez, B.; Curb, J.D. FOXO3A genotype is strongly associated with human longevity. Proc. Natl. Acad. Sci. USA 2008, 105, 13987–13992. [Google Scholar] [CrossRef]
- Yeung, F.; Hoberg, J.E.; Ramsey, C.S.; Keller, M.D.; Jones, D.R.; Frye, R.A.; Mayo, M.W. Modulation of NF-kappaB-dependent transcription and cell survival by the SIRT1 deacetylase. Embo J. 2004, 23, 2369–2380. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Ji, M.; He, G.; Yang, L.; Niu, Z.; Jian, M.; Wei, Y.; Ren, L.; Xu, J. Silencing CDR1as inhibits colorectal cancer progression through regulating microRNA-7. Onco Targets Ther. 2017, 10, 2045–2056. [Google Scholar] [CrossRef]
- Chen, Z.L.; Li, X.N.; Ye, C.X.; Chen, H.Y.; Wang, Z.J. Elevated Levels of circRUNX1 in Colorectal Cancer Promote Cell Growth and Metastasis via miR-145-5p/IGF1 Signalling. Onco Targets Ther. 2020, 13, 4035–4048. [Google Scholar] [CrossRef]
- Wang, P.; Sun, Y.; Yang, Y.; Chen, Y.; Liu, H. Circ_0067835 Knockdown Enhances the Radiosensitivity of Colorectal Cancer by miR-296-5p/IGF1R Axis. Onco Targets Ther. 2021, 14, 491–502. [Google Scholar] [CrossRef]
- Wang, M.; Shi, J.; Jiang, H.; Xu, K.; Huang, Z. Circ_0014130 Participates in the Proliferation and Apoptosis of Nonsmall Cell Lung Cancer Cells via the miR-142-5p/IGF-1 Axis. Cancer Biother. Radiopharm. 2020, 35, 233–240. [Google Scholar] [CrossRef]
- He, S.; Yang, J.; Jiang, S.; Li, Y.; Han, X. Circular RNA circ_0000517 regulates hepatocellular carcinoma development via miR-326/IGF1R axis. Cancer Cell Int. 2020, 20, 404. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Wang, S.; Zhang, X.; Wang, D. Circular RNA circPLK1 promotes breast cancer cell proliferation, migration and invasion by regulating miR-4500/IGF1 axis. Cancer Cell Int. 2020, 20, 593. [Google Scholar] [CrossRef]
- Mao, Y.; Li, W.; Hua, B.; Gu, X.; Pan, W.; Chen, Q.; Xu, B.; Lu, C.; Wang, Z. Circular RNA_PDHX Promotes the Proliferation and Invasion of Prostate Cancer by Sponging MiR-378a-3p. Front. Cell Dev. Biol. 2020, 8, 602707. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, L.; Sun, X. CircRNA hsa_circ_0002577 accelerates endometrial cancer progression through activating IGF1R/PI3K/Akt pathway. J. Exp. Clin. Cancer Res. 2020, 39, 169. [Google Scholar] [CrossRef]
- Xin, T.; Li, S.; Zhang, Y.; Kamali, X.; Liu, H.; Jia, T. circRNA Hsa_circ_0020850 Silence Represses the Development of Lung Adenocarcinoma via Regulating miR-195-5p/IRS2 Axis. Cancer Manag. Res. 2020, 12, 10679–10692. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Y.; Xu, D.; Cao, L. hsa_circ_0023409 Accelerates Gastric Cancer Cell Growth and Metastasis Through Regulating the IRS4/PI3K/AKT Pathway. Cell Transpl. 2021, 30, 963689720975390. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Niu, X.; Li, H.; Liang, Y.; Sun, Z.; Yan, Y. Circ_0000003 promotes the proliferation and metastasis of non-small cell lung cancer cells via miR-338-3p/insulin receptor substrate 2. Cell Cycle 2019, 18, 3525–3539. [Google Scholar] [CrossRef]
- Wang, Y.; Ai, D.; Li, S. CirRNA circFAM126A Exerts Oncogenic Functions in NSCLC to Upregulate IRS2. Biochem. Genet. 2022. [Google Scholar] [CrossRef]
- Akhter, R. Circular RNA and Alzheimer’s Disease. Adv. Exp. Med. Biol. 2018, 1087, 239–243. [Google Scholar]
- Stoll, L.; Sobel, J.; Rodriguez-Trejo, A.; Guay, C.; Lee, K.; Venø, M.T.; Kjems, J.; Laybutt, D.R.; Regazzi, R. Circular RNAs as novel regulators of β-cell functions in normal and disease conditions. Mol. Metab. 2018, 9, 69–83. [Google Scholar] [CrossRef]
- Jiang, J.; Li, R.; Wang, J.; Hou, J.; Qian, H.; Xu, W. Circular RNA CDR1as Inhibits the Metastasis of Gastric Cancer through Targeting miR-876-5p/GNG7 Axis. Gastroenterol. Res. Pract. 2021, 2021, 5583029. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Gong, X.; Sun, L.; Zhou, Q.; Lu, B.; Zhu, L. The Circular RNA Cdr1as Act as an Oncogene in Hepatocellular Carcinoma through Targeting miR-7 Expression. PLoS ONE 2016, 11, e0158347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmenier, F.M.; Lasheen, D.S.; Abouzid KA, M. Phosphatidylinositol 3 kinase (PI3K) inhibitors as new weapon to combat cancer. Eur. J. Med. Chem. 2019, 183, 111718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, M.; Lu, H.; Peng, T. Up-Regulation of circEIF6 Contributes to Pancreatic Cancer Development Through Targeting miR-557/SLC7A11/PI3K/AKT Signaling. Cancer Manag. Res. 2021, 13, 247–258. [Google Scholar] [CrossRef]
- Kong, Y.; Li, Y.; Luo, Y.; Zhu, J.; Zheng, H.; Gao, B.; Guo, X.; Li, Z.; Chen, R.; Chen, C. circNFIB1 inhibits lymphangiogenesis and lymphatic metastasis via the miR-486-5p/PIK3R1/VEGF-C axis in pancreatic cancer. Mol. Cancer 2020, 19, 82. [Google Scholar] [CrossRef]
- Guo, X.; Zhou, Q.; Su, D.; Luo, Y.; Fu, Z.; Huang, L.; Li, Z.; Jiang, D.; Kong, Y.; Li, Z.; et al. Circular RNA circBFAR promotes the progression of pancreatic ductal adenocarcinoma via the miR-34b-5p/MET/Akt axis. Mol. Cancer 2020, 19, 83. [Google Scholar] [CrossRef]
- Zheng, K.; Xie, H.; Wu, W.; Wen, X.; Zeng, Z.; Shi, Y. CircRNA PIP5K1A promotes the progression of glioma through upregulation of the TCF12/PI3K/AKT pathway by sponging miR-515-5p. Cancer Cell Int. 2021, 21, 27. [Google Scholar] [CrossRef]
- Mutalifu, N.; Du, P.; Zhang, J.; Akbar, H.; Yan, B.; Alimu, S.; Tong, L.; Luan, X. Circ_0000215 Increases the Expression of CXCR2 and Promoted the Progression of Glioma Cells by Sponging miR-495-3p. Technol. Cancer Res. Treat. 2020, 19, 1533033820957026. [Google Scholar] [CrossRef]
- Qiao, J.; Liu, M.; Tian, Q.; Liu, X. Microarray analysis of circRNAs expression profile in gliomas reveals that circ_0037655 could promote glioma progression by regulating miR-214/PI3K signaling. Life Sci. 2020, 245, 117363. [Google Scholar] [CrossRef]
- Shi, F.; Shi, Z.; Zhao, Y.; Tian, J. CircRNA hsa-circ-0014359 promotes glioma progression by regulating miR-153/PI3K signaling. Biochem. Biophys. Res. Commun. 2019, 510, 614–620. [Google Scholar] [CrossRef]
- Xia, X.; Li, X.; Li, F.; Wu, X.; Zhang, M.; Zhou, H.; Huang, N.; Yang, X.; Xiao, F.; Liu, D.; et al. A novel tumor suppressor protein encoded by circular AKT3 RNA inhibits glioblastoma tumorigenicity by competing with active phosphoinositide-dependent Kinase-1. Mol. Cancer 2019, 18, 131. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Xu, Y.; Xu, T.; Fan, R.; Jiang, T.; Cao, M.; Shi, L.; Song, J. CircPIP5K1A activates KRT80 and PI3K/AKT pathway to promote gastric cancer development through sponging miR-671-5p. Biomed. Pharmacother. 2020, 126, 109941. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Sun, H.; Wang, Q.; Wang, X.; Quan, J.; Dong, D.; Lun, Y. Circular RNA circMAN2B2 promotes growth and migration of gastric cancer cells by down-regulation of miR-145. J. Clin. Lab. Anal. 2020, 34, e23215. [Google Scholar] [CrossRef]
- Liang, X.; Qin, C.; Yu, G.; Guo, X.; Cheng, A.; Zhang, H.; Wang, Z. Circular RNA circRAB31 acts as a miR-885-5psponge to suppress gastric cancer progressionvia the PTEN/PI3K/AKT pathway. Mol. Ther. Oncolytics 2021, 23, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, C. Upregulated hsa_circRNA_100269 inhibits the growth and metastasis of gastric cancer through inactivating PI3K/Akt axis. PLoS ONE 2021, 16, e0250603. [Google Scholar] [CrossRef]
- Chen, T.; Liu, Y.; Li, C.; Xu, C.; Ding, C.; Chen, J.; Zhao, J. Tumor-derived exosomal circFARSA mediates M2 macrophage polarization via the PTEN/PI3K/AKT pathway to promote non-small cell lung cancer metastasis. Cancer Treat. Res. Commun. 2021, 28, 100412. [Google Scholar] [CrossRef]
- Si, J.; Jin, J.; Sai, J.; Liu, X.; Luo, X.; Fu, Z.; Wang, J. Circular RNA circ-PLCD1 functions as a tumor suppressor in non-small cell lung cancer by inactivation of PI3K/AKT signaling pathway. Hum. Cell 2022, 35, 924–935. [Google Scholar] [CrossRef]
- Li, W.; Yang, P.; Zhong, C.; Shen, X.; Shi, X.; Li, X. The circ-PITX1 promotes non-small cell lung cancer development via the miR-30e-5p/ITGA6 axis. Cell Cycle 2022, 21, 304–321. [Google Scholar] [CrossRef]
- Li, C.; Li, X. circPTEN suppresses colorectal cancer progression through regulating PTEN/AKT pathway. Mol. Ther. Nucleic Acid. 2021, 26, 1418–1432. [Google Scholar] [CrossRef]
- Wang, J.; Luo, J.; Liu, G.; Li, X. Circular RNA hsa_circ_0008285 inhibits colorectal cancer cell proliferation and migration via the miR-382-5p/PTEN axis. Biochem. Biophys. Res. Commun. 2020, 527, 503–510. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Long, H.; Lin, W.; Wang, H.; Chen, Y.; Yuan, Q.; Li, X. circCDYL2, Overexpressed in Highly Migratory Colorectal Cancer Cells, Promotes Migration by Binding to Ezrin. Front. Oncol. 2021, 11, 716073. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.H.; Wang, Y.; Xu, D. Hsa_circ_001569 is an unfavorable prognostic factor and promotes cell proliferation and metastasis by modulating PI3K-AKT pathway in breast cancer. Cancer Biomark 2019, 25, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Rong, D.; Hui, B.; He, X.; Jiang, W.; Xu, Y.; Cao, H.; Xu, Z.; Tang, W. CircETFA upregulates CCL5 by sponging miR-612 and recruiting EIF4A3 to promote hepatocellular carcinoma. Cell Death Discov. 2021, 7, 321. [Google Scholar] [CrossRef]
- Fu, H.W.; Lin, X.; Zhu, Y.X.; Lan, X.; Kuang, Y.; Wang, Y.Z.; Ke, Z.G.; Yuan, T.; Chen, P. Circ-IGF1R has pro-proliferative and anti-apoptotic effects in HCC by activating the PI3K/AKT pathway. Gene 2019, 716, 144031. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Xu, X.; Zhang, X.; Zhou, Y. Circular RNA-9119 suppresses in ovarian cancer cell viability via targeting the microRNA-21-5p-PTEN-Akt pathway. Aging 2020, 12, 14314–14328. [Google Scholar] [CrossRef]
- Chen, W.; Cen, S.; Zhou, X.; Yang, T.; Wu, K.; Zou, L.; Luo, J.; Li, C.; Lv, D.; Mao, X. Circular RNA CircNOLC1, Upregulated by NF-KappaB, Promotes the Progression of Prostate Cancer via miR-647/PAQR4 Axis. Front. Cell Dev. Biol. 2020, 8, 624764. [Google Scholar] [CrossRef]
- Yao, J.; Qian, K.; Chen, C.; Liu, X.; Yu, D.; Yan, X.; Liu, T.; Li, S. ZNF139/circZNF139 promotes cell proliferation, migration and invasion via activation of PI3K/AKT pathway in bladder cancer. Aging 2020, 12, 9915–9934. [Google Scholar] [CrossRef]
- Hasan, S.; Jacob, R.; Manne, U.; Paluri, R. Advances in pancreatic cancer biomarkers. Oncol. Rev. 2019, 13, 410. [Google Scholar] [CrossRef]
- Mikheev, A.M.; Ramakrishna, R.; Stoll, E.A.; Mikheeva, S.A.; Beyer, R.P.; Plotnik, D.A.; Schwartz, J.L.; Rockhill, J.K.; Silber, J.R.; Born, D.E.; et al. Increased age of transformed mouse neural progenitor/stem cells recapitulates age-dependent clinical features of human glioma malignancy. Aging Cell 2012, 11, 1027–1035. [Google Scholar] [CrossRef]
- Stoll, E.A.; Horner, P.J.; Rostomily, R.C. The impact of age on oncogenic potential: Tumor-initiating cells and the brain microenvironment. Aging Cell 2013, 12, 733–741. [Google Scholar] [CrossRef]
- Jovčevska, I.; Kočevar, N.; Komel, R. Glioma and glioblastoma—How much do we (not) know? Mol. Clin. Oncol. 2013, 1, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Wu, Y.; Yang, J.; Yang, D.; Fang, X. Progress in the treatment of advanced gastric cancer. Tumour. Biol. 2017, 39, 1010428317714626. [Google Scholar] [CrossRef] [Green Version]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [PubMed]
- Xiao, B.; Zhang, X.; Li, X.; Zhao, Z. Circ_001569 regulates FLOT2 expression to promote the proliferation, migration, invasion and EMT of osteosarcoma cells through sponging miR-185-5p. Open Life Sci. 2020, 15, 476–487. [Google Scholar] [CrossRef]
- Liu, H.; Xue, L.; Song, C.; Liu, F.; Jiang, T.; Yang, X. Overexpression of circular RNA circ_001569 indicates poor prognosis in hepatocellular carcinoma and promotes cell growth and metastasis by sponging miR-411-5p and miR-432-5p. Biochem. Biophys. Res. Commun. 2018, 503, 2659–2665. [Google Scholar] [CrossRef]
- Xie, H.; Ren, X.; Xin, S.; Lan, X.; Lu, G.; Lin, Y.; Yang, S.; Zeng, Z.; Liao, W.; Ding, Y.Q.; et al. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget 2016, 7, 26680–26691. [Google Scholar] [CrossRef]
- Gu, F.; Zhang, J.; Yan, L.; Li, D. CircHIPK3/miR-381-3p axis modulates proliferation, migration, and glycolysis of lung cancer cells by regulating the AKT/mTOR signaling pathway. Open Life Sci. 2020, 15, 683–695. [Google Scholar] [CrossRef]
- Wang, X.; Li, R.; Feng, L.; Wang, J.; Qi, Q.; Wei, W.; Yu, Z. Hsa_circ_0001666 promotes non-small cell lung cancer migration and invasion through miR-1184/miR-548I/AGO1 axis. Mol. Ther. Oncolytics 2022, 24, 597–611. [Google Scholar] [CrossRef]
- Shi, J.; Liu, C.; Chen, C.; Guo, K.; Tang, Z.; Luo, Y.; Chen, L.; Su, Y.; Xu, K. Circular RNA circMBOAT2 promotes prostate cancer progression via a miR-1271-5p/mTOR axis. Aging 2020, 12, 13255–13280. [Google Scholar] [CrossRef]
- Jia, C.; Yao, Z.; Lin, Z.; Zhao, L.; Cai, X.; Chen, S.; Deng, M.; Zhang, Q. circNFATC3 sponges miR-548I acts as a ceRNA to protect NFATC3 itself and suppressed hepatocellular carcinoma progression. J. Cell Physiol. 2021, 236, 1252–1269. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, T.; Li, C.; Xu, C.; Ding, C.; Chen, J.; Ju, S.; Zhang, Z.; Liang, Z.; Cui, Z.; et al. A circular RNA hsa_circ_0079929 inhibits tumor growth in hepatocellular carcinoma. Cancer Manag. Res. 2019, 11, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Gao, J.; Zheng, R.; Yu, M.; Ren, Y.; Yan, T.; Huang, Y.; Li, Y. Antagonizing circRNA_002581-miR-122-CPEB1 axis alleviates NASH through restoring PTEN-AMPK-mTOR pathway regulated autophagy. Cell Death Dis. 2020, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, Z.; Ding, H.; Li, H.; Wang, W.; Pan, Z. CircWHSC1 Promotes Breast Cancer Progression by Regulating the FASN/AMPK/mTOR Axis Through Sponging miR-195-5p. Front. Oncol. 2021, 11, 649242. [Google Scholar] [CrossRef]
- Chen, R.; Mao, L.; Shi, R.; Wang, W.; Cheng, J. circRNA MYLK Accelerates Cervical Cancer via Up-Regulation of RHEB and Activation of mTOR Signaling. Cancer Manag. Res. 2020, 12, 3611–3621. [Google Scholar] [CrossRef]
- Huang, X.Y.; Huang, Z.L.; Zhang, P.B.; Huang, X.Y.; Huang, J.; Wang, H.C.; Xu, B.; Zhou, J.; Tang, Z.Y. CircRNA-100338 Is Associated With mTOR Signaling Pathway and Poor Prognosis in Hepatocellular Carcinoma. Front. Oncol. 2019, 9, 392. [Google Scholar] [CrossRef]
- Liu, D.; Bi, X.; Yang, Y. Circular RNA hsa_circ_0011324 is involved in endometrial cancer progression and the evolution of its mechanism. Bioengineered 2022, 13, 7485–7499. [Google Scholar] [CrossRef]
- Cao, Q.; Shi, Y.; Wang, X.; Yang, J.; Mi, Y.; Zhai, G.; Zhang, M. Circular METRN RNA hsa_circ_0037251 Promotes Glioma Progression by Sponging miR-1229-3p and Regulating mTOR Expression. Sci. Rep. 2019, 9, 19791. [Google Scholar] [CrossRef]
- Chen, X.; Liu, C.; Ji, L.; Wang, N.; Liu, Y.; Wang, M.; Ruan, L. The circACC1/miR-29c-3p/FOXP1 network plays a key role in gastric cancer by regulating cell proliferation. Biochem. Biophys. Res. Commun. 2021, 557, 221–227. [Google Scholar] [CrossRef]
- Zhu, B.; Ke, L.; Li, P.; Wang, X.; Yang, L.; Bai, M.; Chen, M. CircACC1 Promotes NSCLC Proliferation via miR-29c-3p/MCL-1 Signaling Pathway. Front. Genet. 2021, 12, 798587. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R.F.; Zhang, X.D.; Hu, W.; Wu, M. CircACC1 Regulates Assembly and Activation of AMPK Complex under Metabolic Stress. Cell Metab. 2019, 30, 157–173.e7. [Google Scholar] [CrossRef] [PubMed]
- Jun, T.; Chen, W.; Hailing, C.; Ning, W.; Qinxue, C. The novel circular RNA circ-PGAP3 retards cervical cancer growth by regulating the miR-769-5p/p53 axis. Hum. Cell 2021, 34, 878–888. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Jiang, W.; Hui, B.; Rong, D.; Fu, K.; Dong, C.; Tang, W.; Cao, H. The circ_0021977/miR-10b-5p/P21 and P53 regulatory axis suppresses proliferation, migration, and invasion in colorectal cancer. J. Cell Physiol. 2020, 235, 2273–2285. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lin, S.; Mo, Z.; Jiang, J.; Tang, H.; Wu, C.; Song, J. CircRNA_100395 inhibits cell proliferation and metastasis in ovarian cancer via regulating miR-1228/p53/epithelial-mesenchymal transition (EMT) axis. J. Cancer 2020, 11, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Hao, Y.; Lin, K.; Lyu, Y.; Chen, M.; Wang, H.; Zou, D.; Jiang, X.; Wang, R.; Jin, D.; et al. Circular RNA CDR1as disrupts the p53/MDM2 complex to inhibit Gliomagenesis. Mol. Cancer 2020, 19, 138. [Google Scholar] [CrossRef]
- Kong, P.; Li, C.L.; Dou, Y.Q.; Cao, L.; Zhang, X.Y.; Zhang, W.D.; Bi, Z.Q.; Peng, Z.Y.; Yan, A.Q.; Han, M. circ-Sirt1 Decelerates Senescence by Inhibiting p53 Activation in Vascular Smooth Muscle Cells, Ameliorating Neointima Formation. Front. Cardiovasc. Med. 2021, 8, 724592. [Google Scholar] [CrossRef]
- Chen, D.; Zhou, H.; Cai, Z.; Cai, K.; Liu, J.; Wang, W.; Miao, H.; Li, H.; Li, R.; Li, X.; et al. CircSCAP interacts with SF3A3 to inhibit the malignance of non-small cell lung cancer by activating p53 signaling. J. Exp. Clin. Cancer Res. 2022, 41, 120. [Google Scholar] [CrossRef]
- Chaudhary, R.; Muys, B.R.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Li, X.L.; Zhu, Y.; Daulatabad, S.V.; Tsitsipatis, D.; Meltzer, P.S.; et al. A Circular RNA from the MDM2 Locus Controls Cell Cycle Progression by Suppressing p53 Levels. Mol. Cell Biol. 2020, 40, e00473-19. [Google Scholar] [CrossRef]
- Li, Q.K.; Liu, Y.K.; Li, J.W.; Liu, Y.T.; Li, Y.F.; Li, B.H. Circ-sirt1 inhibits growth and invasion of gastric cancer by sponging miR-132-3p/miR-212-3p and upregulating sirt1 expression. Neoplasma 2021, 68, 780–787. [Google Scholar] [CrossRef]
- Huang, M.; Dong, Y.; Sun, G.; Yu, Y. Circ-Sirt1 inhibits vascular smooth muscle cells proliferation via the c-Myc/cyclin B1 axis. Cell Biol. Int. 2022, 46, 628–636. [Google Scholar] [CrossRef]
- Li, H.L.; Wei, Y.Y.; Li, X.H.; Zhang, S.S.; Zhang, R.T.; Li, J.H.; Ma, B.W.; Shao, S.B.; Lv, Z.W.; Ruan, H.; et al. Diosmetin has therapeutic efficacy in colitis regulating gut microbiota, inflammation, and oxidative stress via the circ-Sirt1/Sirt1 axis. Acta Pharmacol. Sin. 2022, 43, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Grootaert, M.O.; da Costa Martins, P.A.; Bitsch, N.; Pintelon, I.; De Meyer, G.R.; Martinet, W.; Schrijvers, D.M. Defective autophagy in vascular smooth muscle cells accelerates senescence and promotes neointima formation and atherogenesis. Autophagy 2015, 11, 2014–2032. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Feng, X.; Wang, Y.; Li, Q.; Li, X. Dysregulation of tumour microenvironment driven by circ-TPGS2/miR-7/TRAF6/NF-κB axis facilitates breast cancer cell motility. Autoimmunity 2021, 54, 284–293. [Google Scholar] [CrossRef]
- Chen, J.; Yang, X.; Liu, R.; Wen, C.; Wang, H.; Huang, L.; Li, W.; Zhu, Z.; Zhu, Y.; Liu, H. Circular RNA GLIS2 promotes colorectal cancer cell motility via activation of the NF-κB pathway. Cell Death Dis. 2020, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Sun, G.; Zheng, W.; Tang, W.; Cheng, Y.; Wu, L.; Li, X.; Tao, J.; Ma, S.; Cao, H. circCORO1C promotes the proliferation and metastasis of hepatocellular carcinoma by enhancing the expression of PD-L1 through NF-κB pathway. J. Clin. Lab. Anal. 2021, 35, e24003. [Google Scholar] [CrossRef]
- Zhou, Q.; Sun, Y. Circular RNA cMras Suppresses the Progression of Lung Adenocarcinoma Through ABHD5/ATGL Axis Using NF-κB Signaling Pathway. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Disease | CircRNA | Functional Mechanism | Targets | References |
|---|---|---|---|---|
| Pancreatic cancer | CircEIF6 | miRNA sponge | miR-557/SLC7A11 | [94] |
| PDAC | CircNFIB1 | miRNA sponge | miR-486-5p/PIK3R1 | [95] |
| PDAC | CircBFAR | miRNA sponge | miR-34b-5p/MET | [96] |
| Glioma | CircPIP5K1A | miRNA sponge | miR-515-5p/TCF12 | [97] |
| Glioma | Circ_0000215 | miRNA sponge | miR-495-3p/CXCR2 | [98] |
| Glioma | Circ_0037655 | miRNA sponge | miR-214 | [99] |
| Glioma | Hsa-circ-0014359 | miRNA sponge | miR-153 | [100] |
| GBM | Circ-AKT3 | translation to protein | PDK1 | [101] |
| GC | CircPIP5K1A | miRNA sponge | miR-671-5p | [102] |
| GC | CircMAN2B2 | miRNA sponge | miR-145 | [103] |
| GC | CircRAB31 | miRNA sponge | miR-885-5p | [104] |
| GC | Hsa_circRNA_100269 | unclear | [105] | |
| NSCLC | CircFARSA | interact with protein | PTEN | [106] |
| NSCLC | Circ-PLCD1 | miRNA sponge | miR-375/miR-1179/PTEN | [107] |
| NSCLC | Circ-PITX1 | miRNA sponge | miR-30E-5p/ITGA6 | [108] |
| CRC | CircPTEN | miRNA sponge | miR-4470/PTEN | [109] |
| CRC | Circ_0008285 | miRNA sponge | miR-382-5p/PTEN | [110] |
| CRC | CircCDYL2 | interact with protein | Ezrin | [111] |
| BC | Hsa_circ_001569 | unclear | [112] | |
| HCC | CircETFA | miRNA sponge | hsa-miR-612/CCL5 | [113] |
| HCC | CircIGF1R | unclear | [114] | |
| OC | CircRNA-9119 | miRNA sponge | miR-21-5p/PTEN | [115] |
| PCa | CircNOLC1 | miRNA sponge | miR-647/PAQR4 | [116] |
| Bladder cancer | CircZNF139 | unclear | [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Xiao, F.; Li, J.; Fan, X.; He, Z.; Yan, T.; Yang, M.; Yang, D. Circular RNAs Involved in the Regulation of the Age-Related Pathways. Int. J. Mol. Sci. 2022, 23, 10443. https://doi.org/10.3390/ijms231810443
Wang S, Xiao F, Li J, Fan X, He Z, Yan T, Yang M, Yang D. Circular RNAs Involved in the Regulation of the Age-Related Pathways. International Journal of Molecular Sciences. 2022; 23(18):10443. https://doi.org/10.3390/ijms231810443
Chicago/Turabian StyleWang, Siqi, Feng Xiao, Jiamei Li, Xiaolan Fan, Zhi He, Taiming Yan, Mingyao Yang, and Deying Yang. 2022. "Circular RNAs Involved in the Regulation of the Age-Related Pathways" International Journal of Molecular Sciences 23, no. 18: 10443. https://doi.org/10.3390/ijms231810443
APA StyleWang, S., Xiao, F., Li, J., Fan, X., He, Z., Yan, T., Yang, M., & Yang, D. (2022). Circular RNAs Involved in the Regulation of the Age-Related Pathways. International Journal of Molecular Sciences, 23(18), 10443. https://doi.org/10.3390/ijms231810443