
Hypochlorous Acid Chemistry in Mammalian Cells—Influence on Infection and Role in Various Pathologies
 ,
,  and
and
Abstract
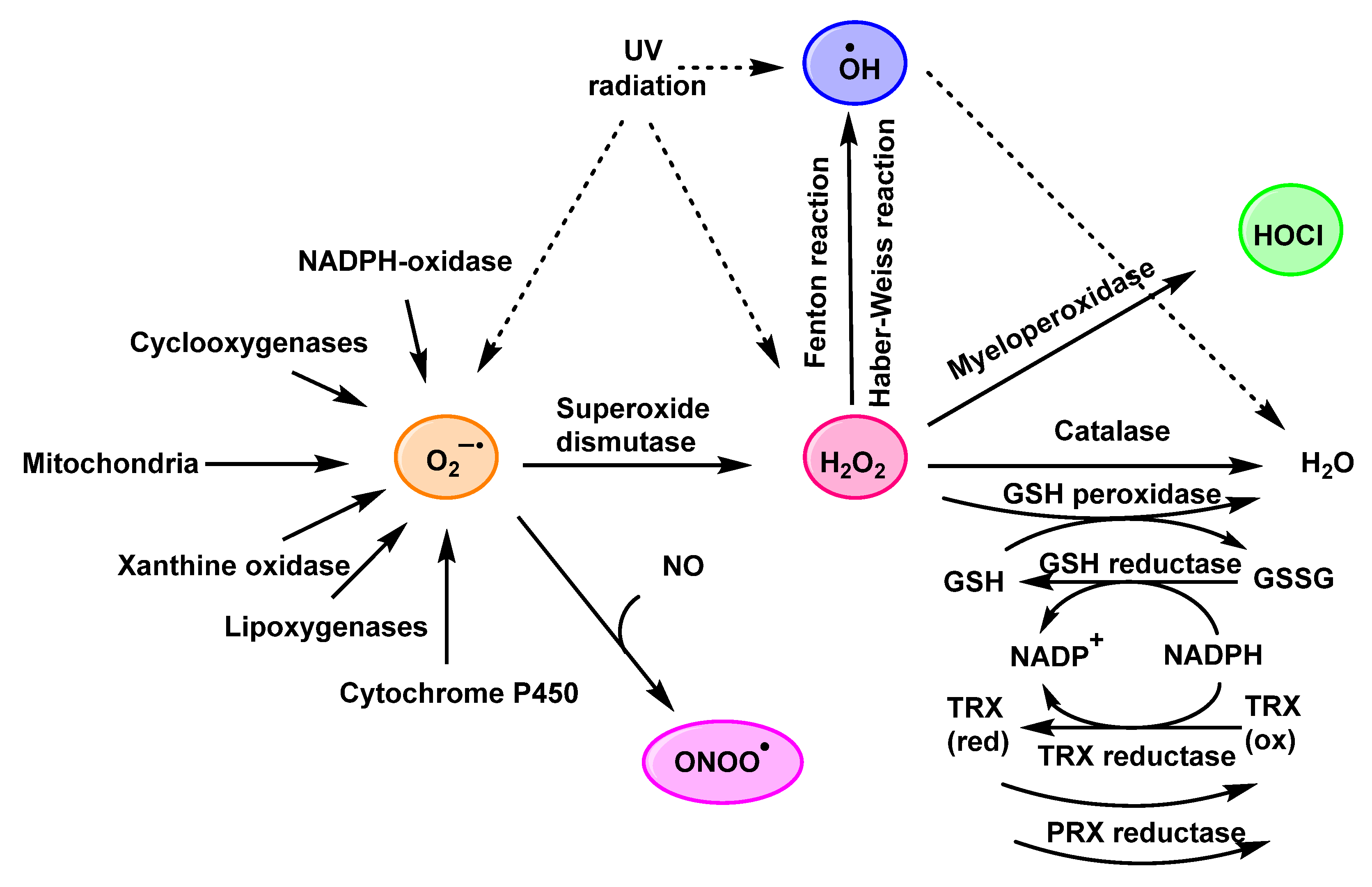
:1. Introduction
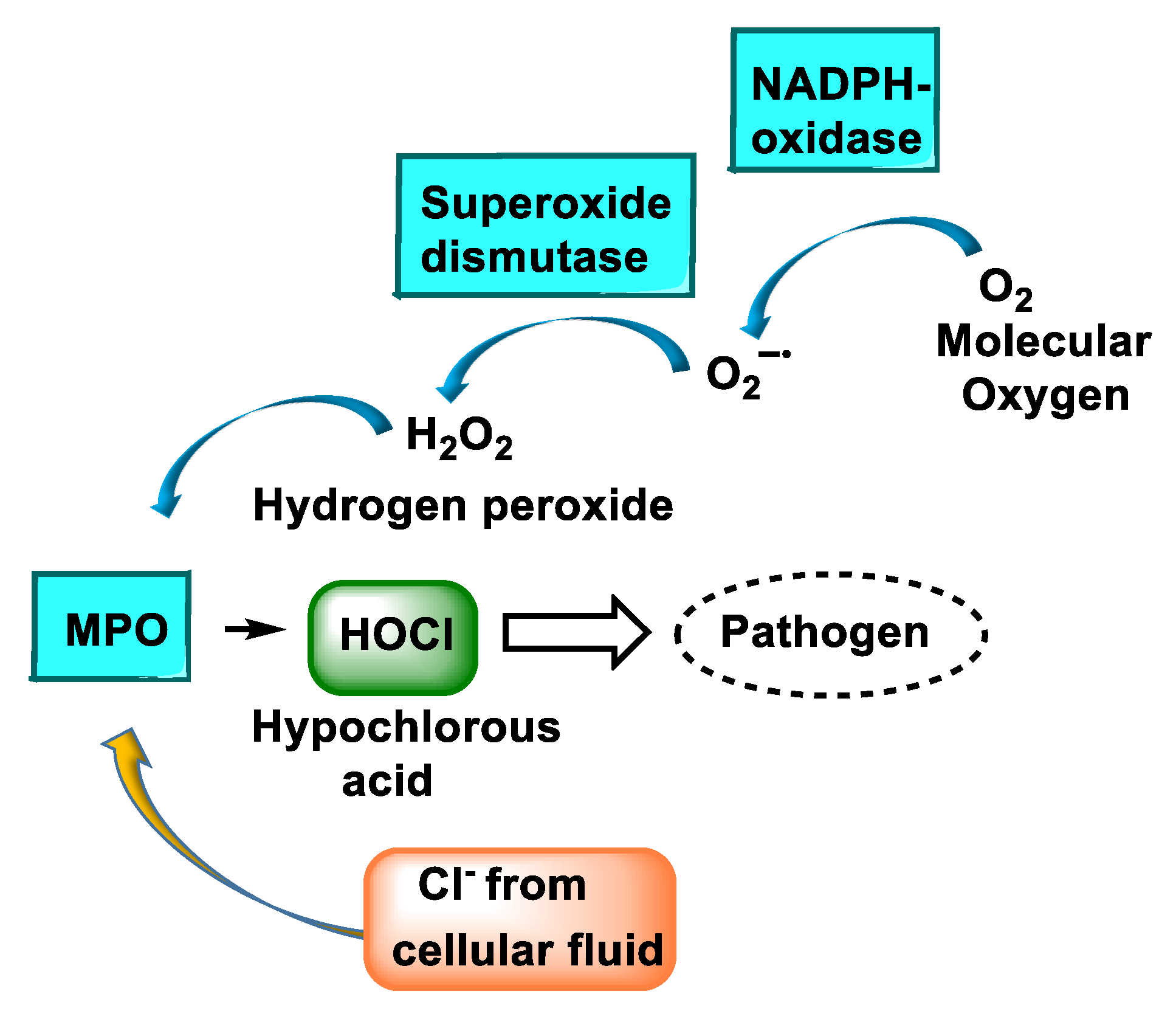
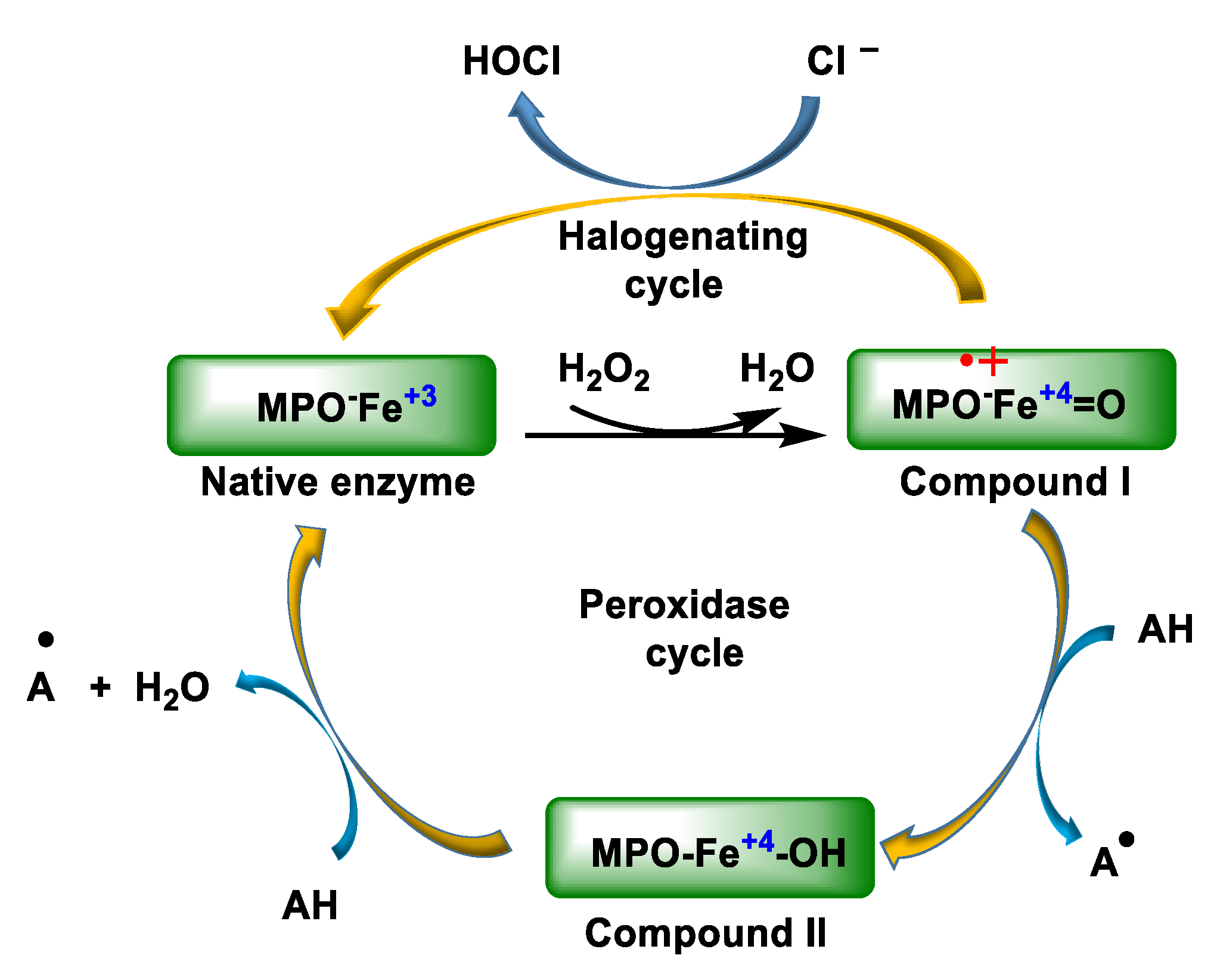
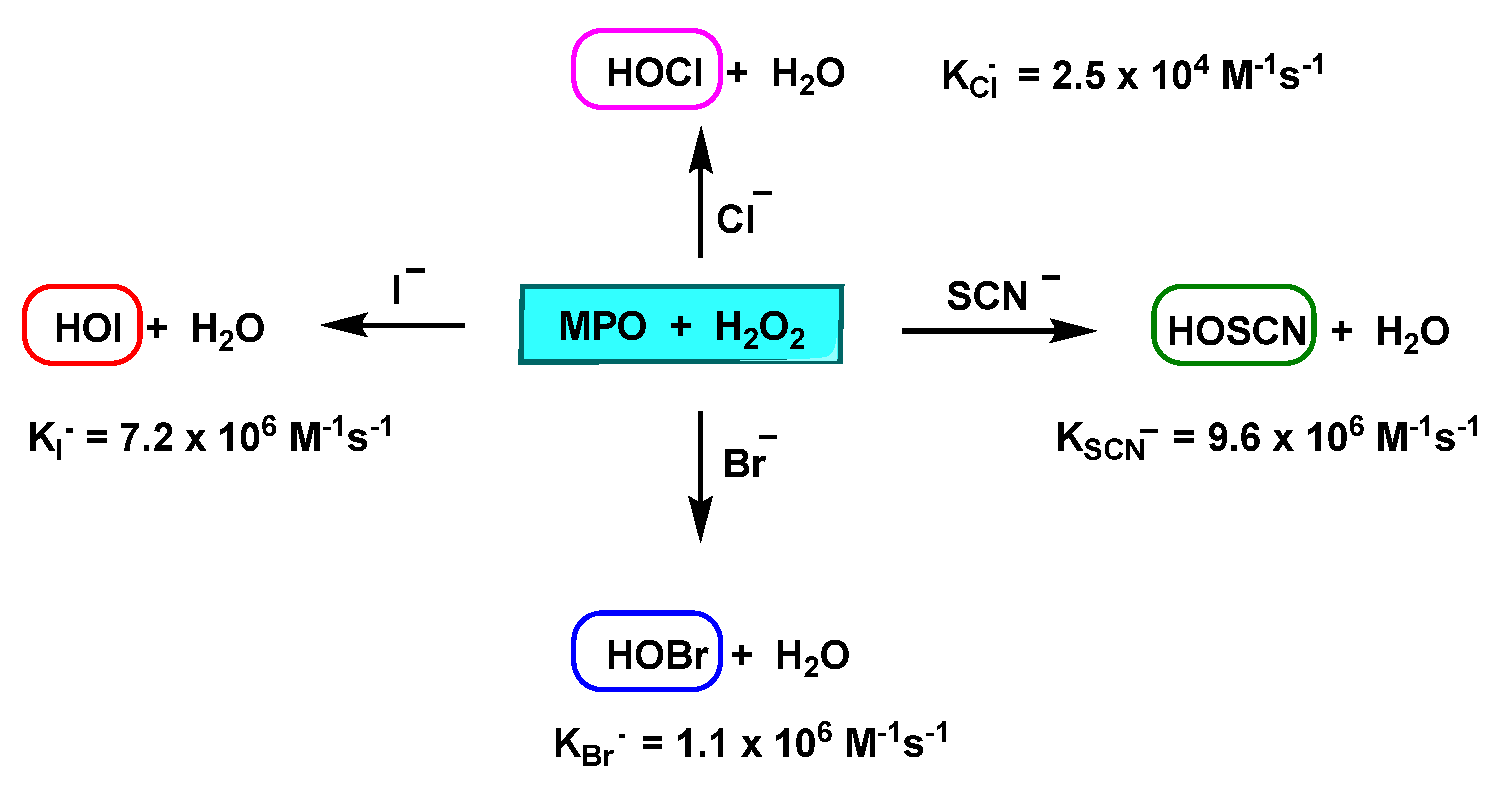
2. Enzyme Myeloperoxidase (MPO) and Its Catalytic Cycle
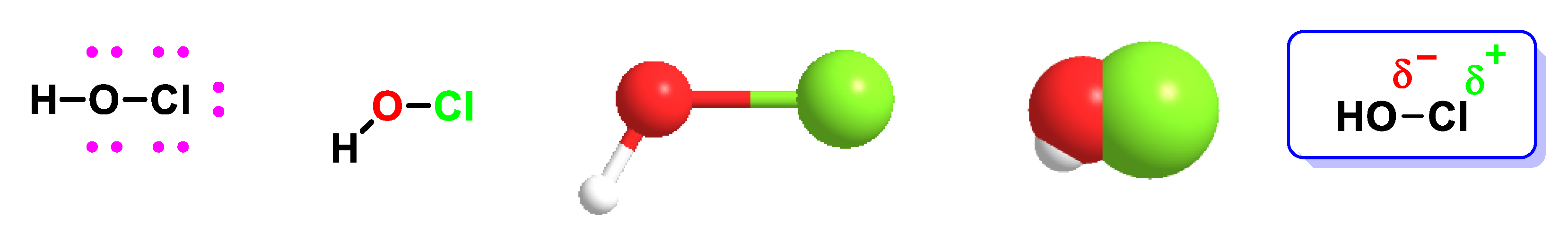
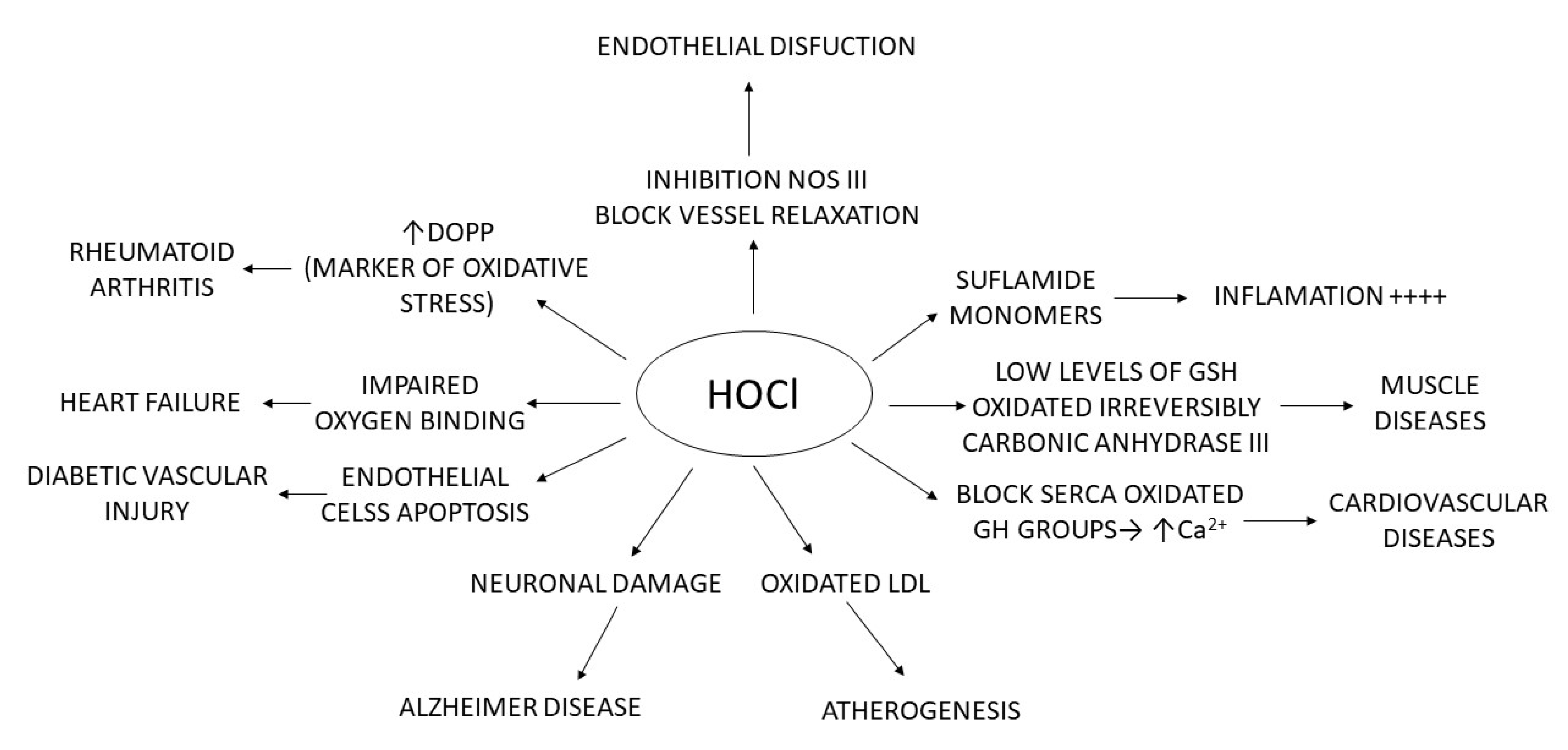
3. Formation and Features of Hypochlorous Acid HOCl
4. HOCl Reaction with Lipids
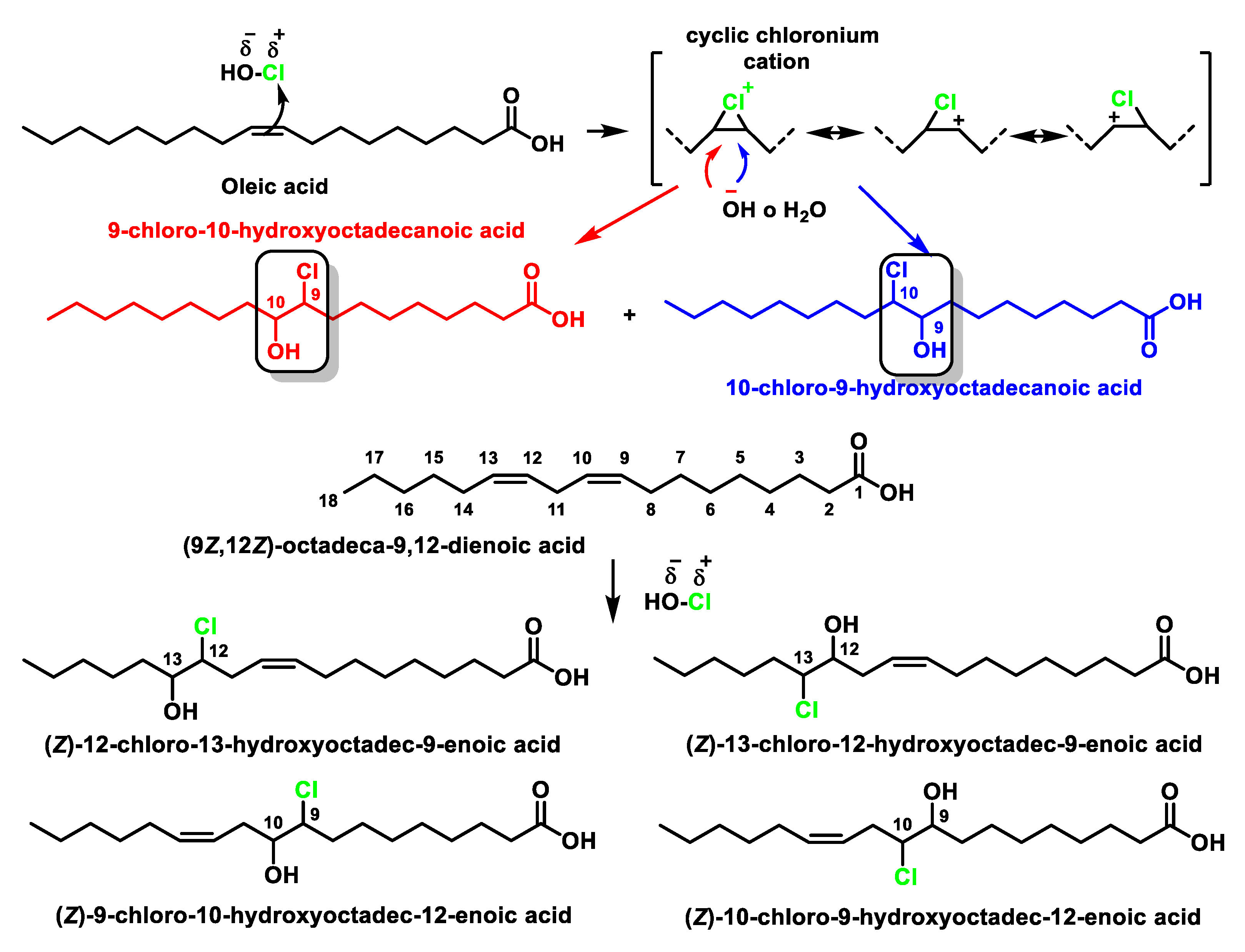
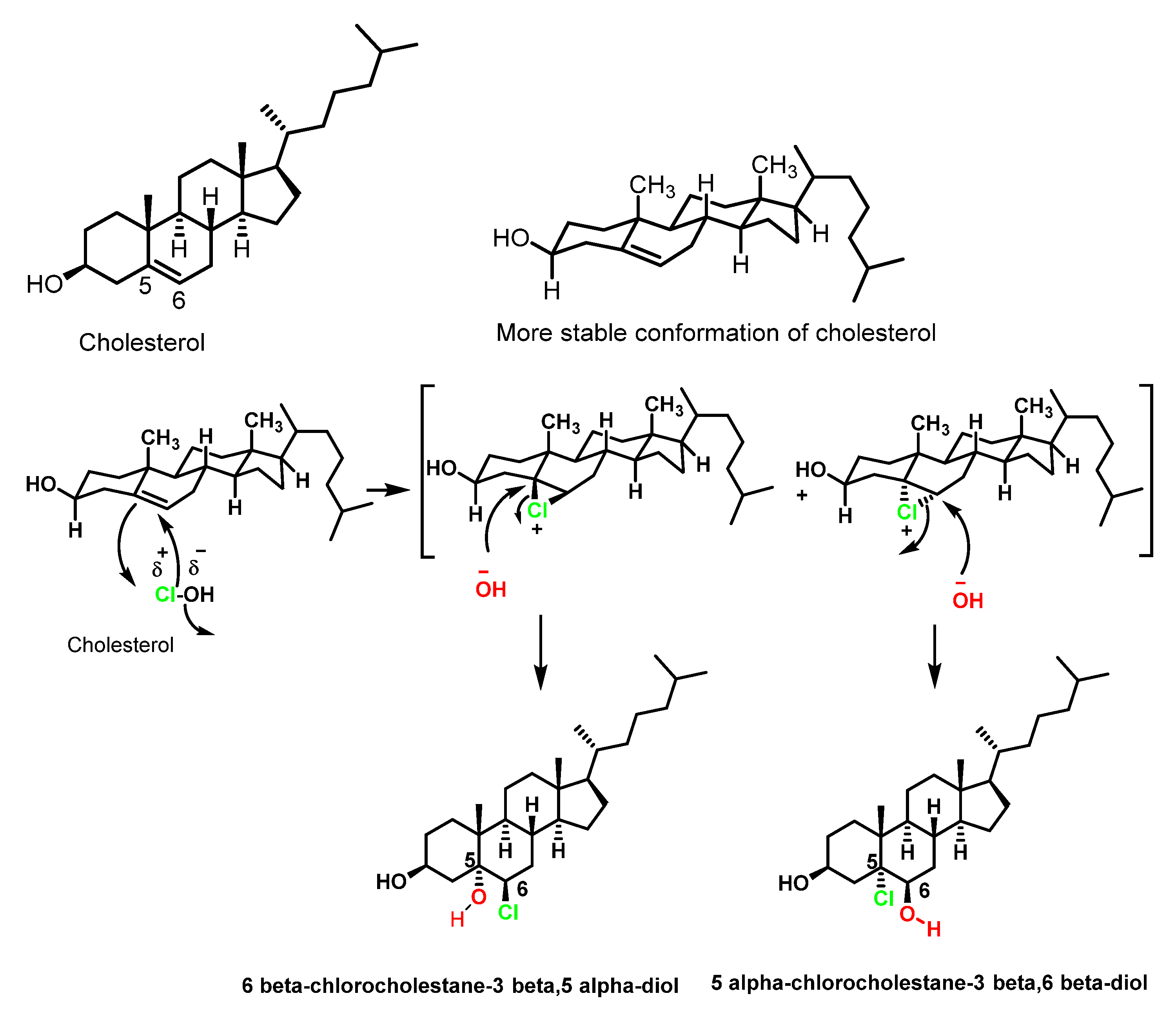
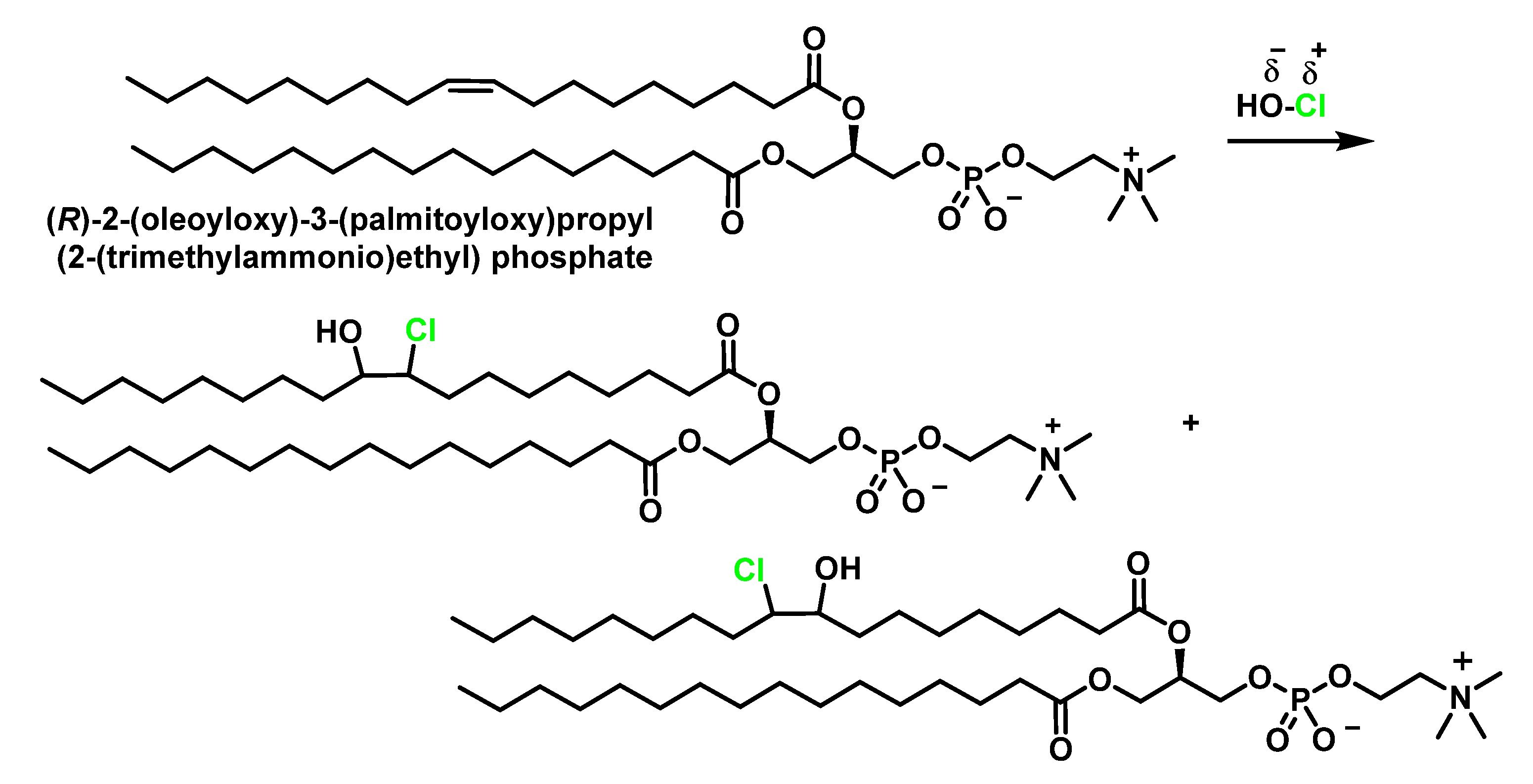
4.1. HOCl Reaction with Unsaturated Bonds
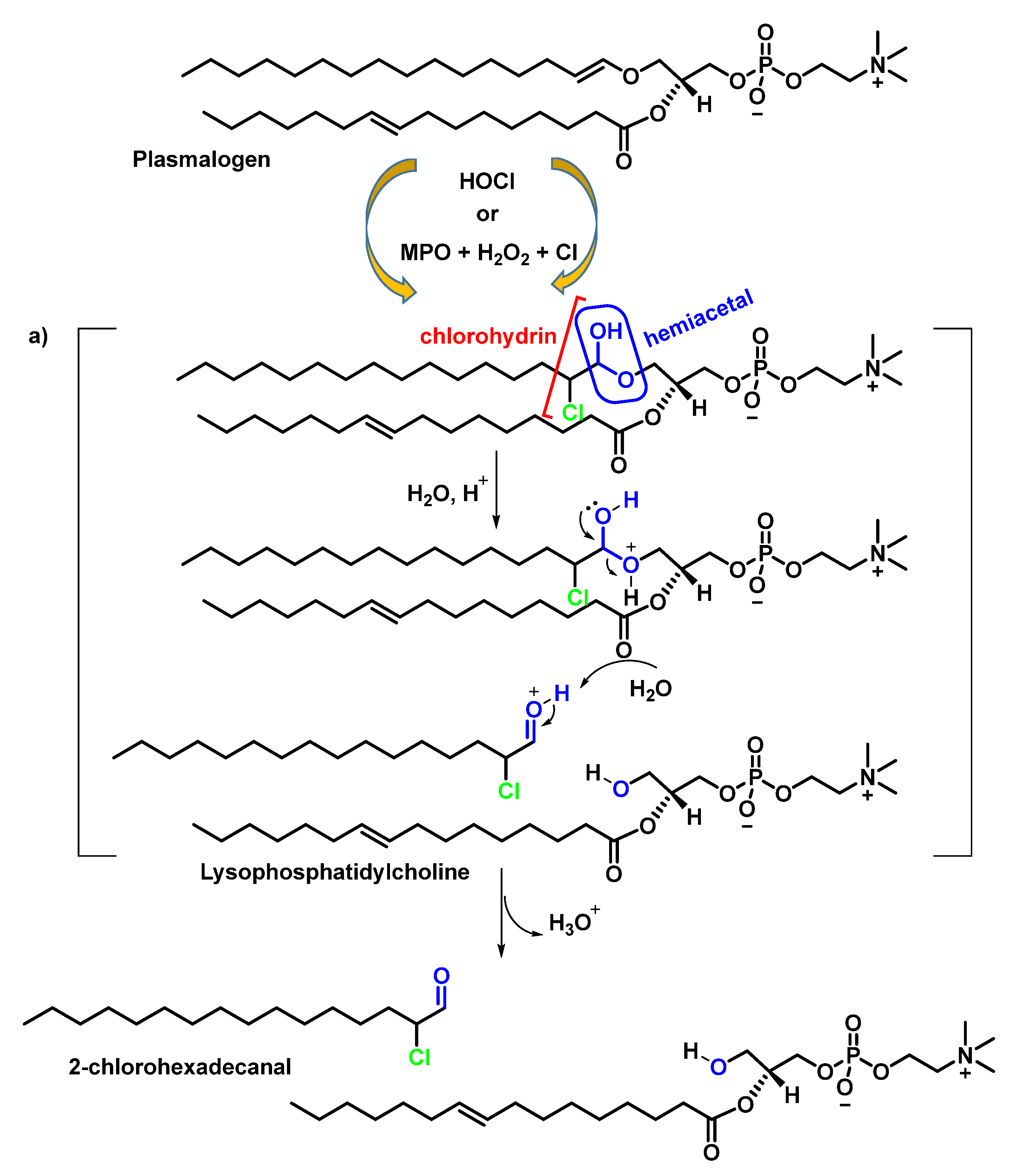
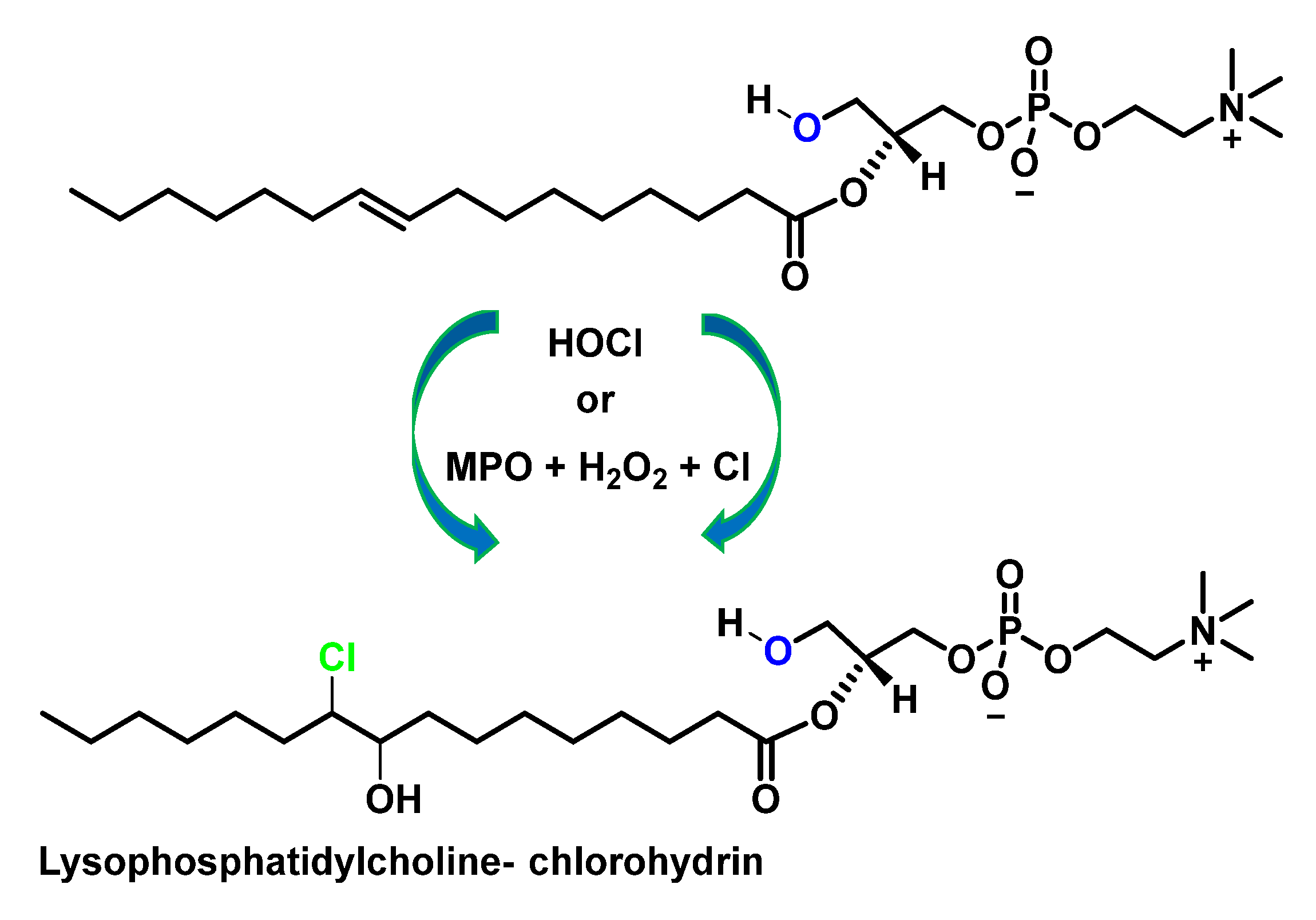
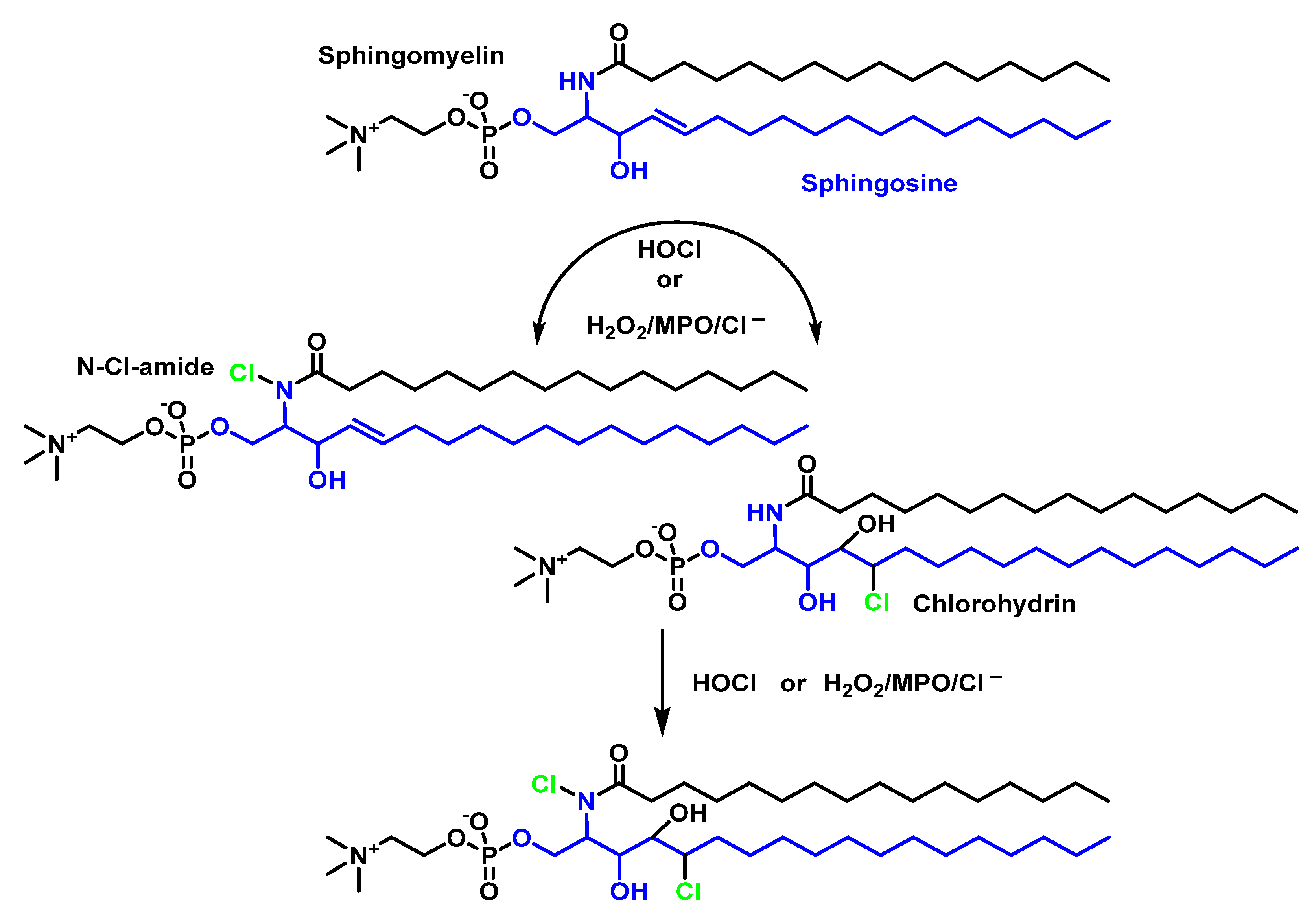
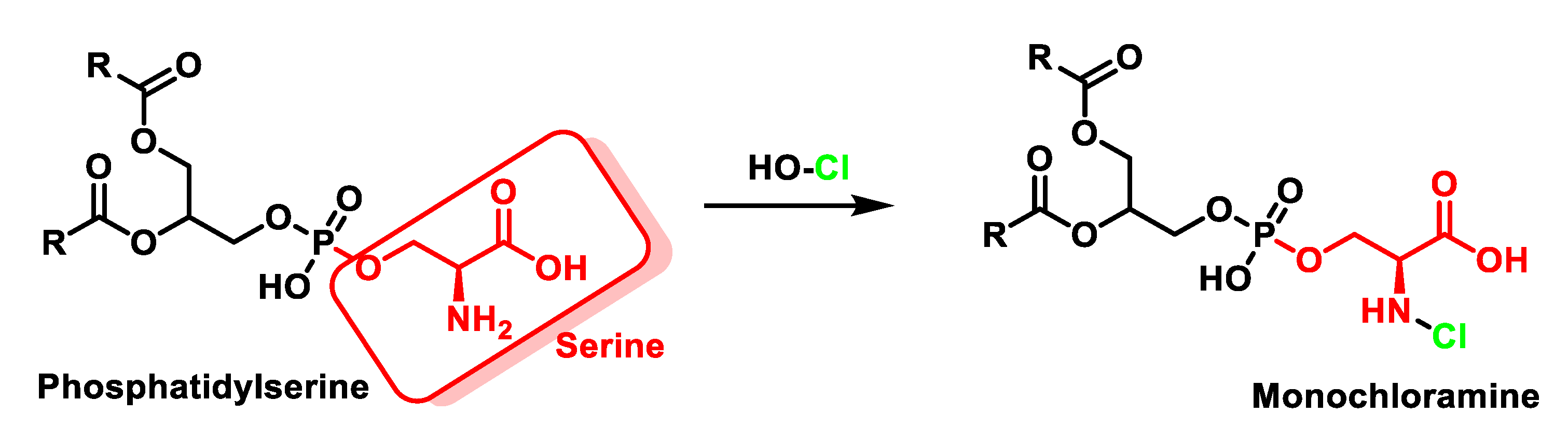
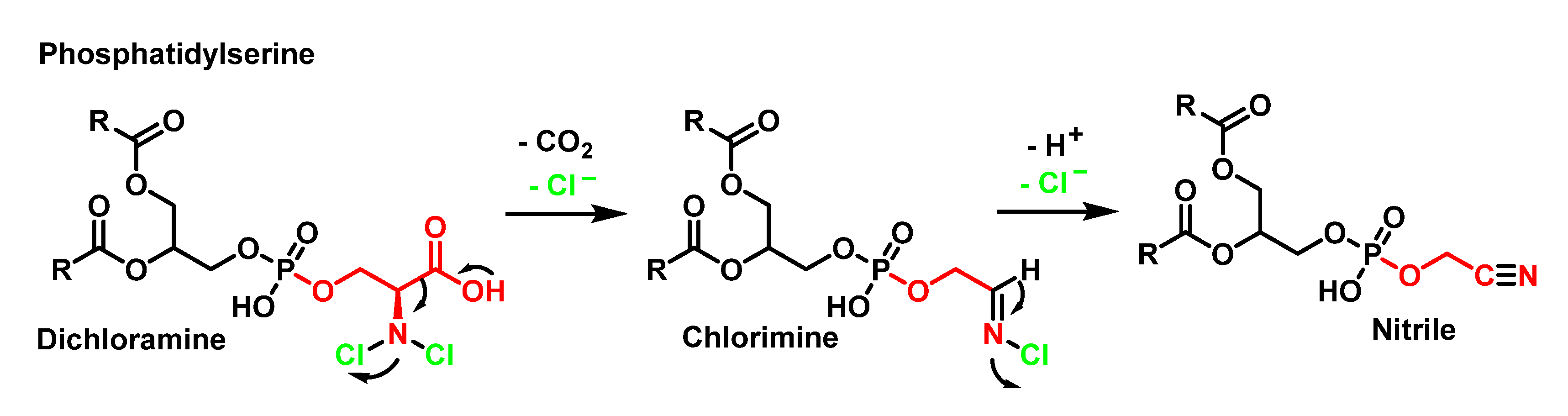
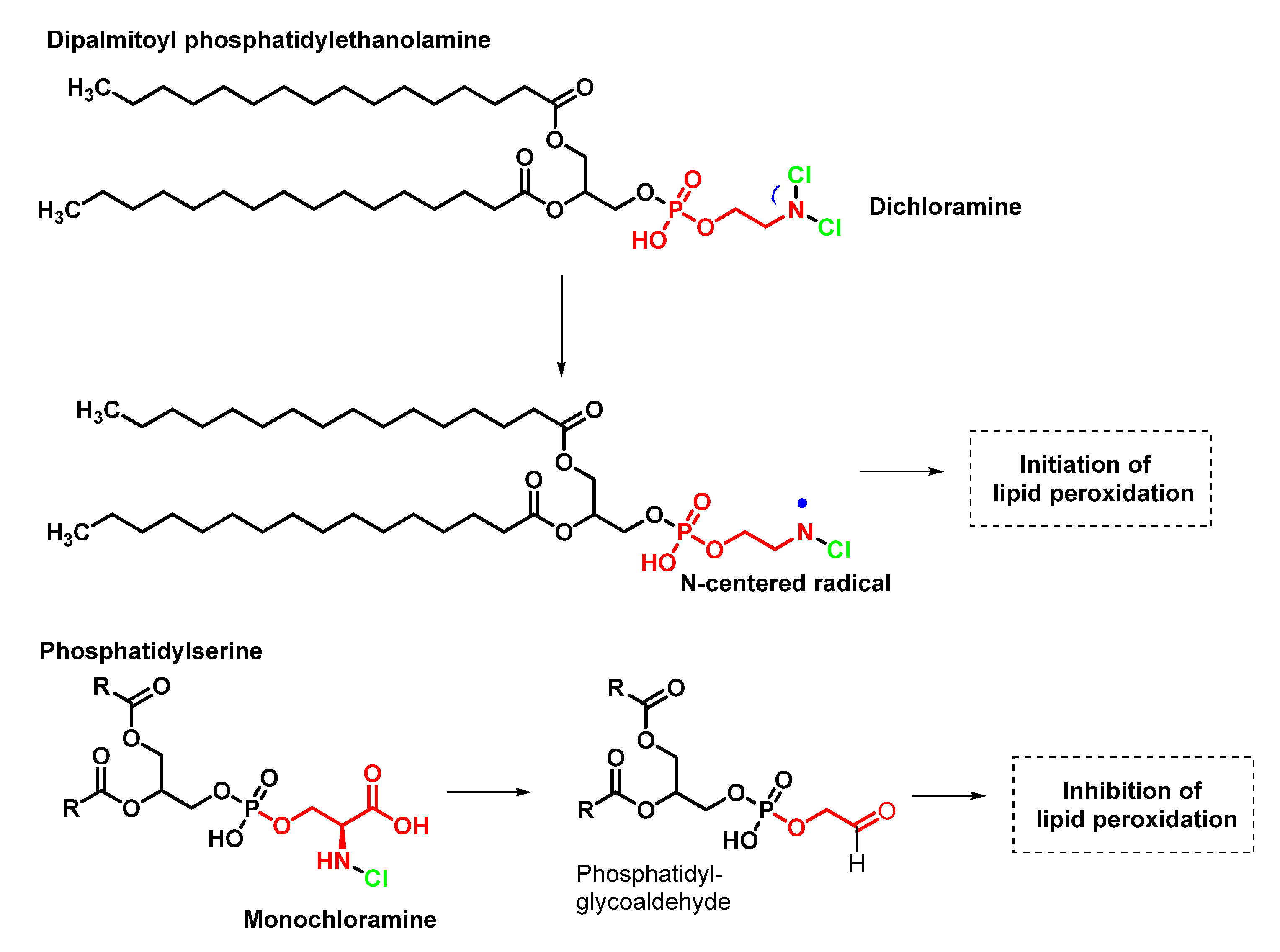
4.2. HOCl Reaction with Phospholipids
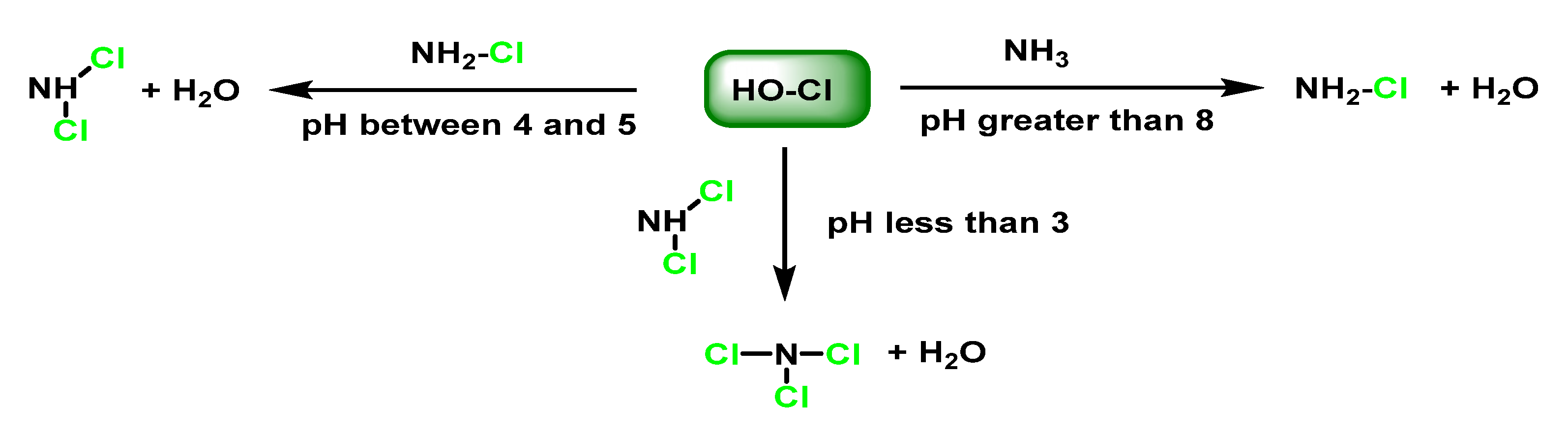
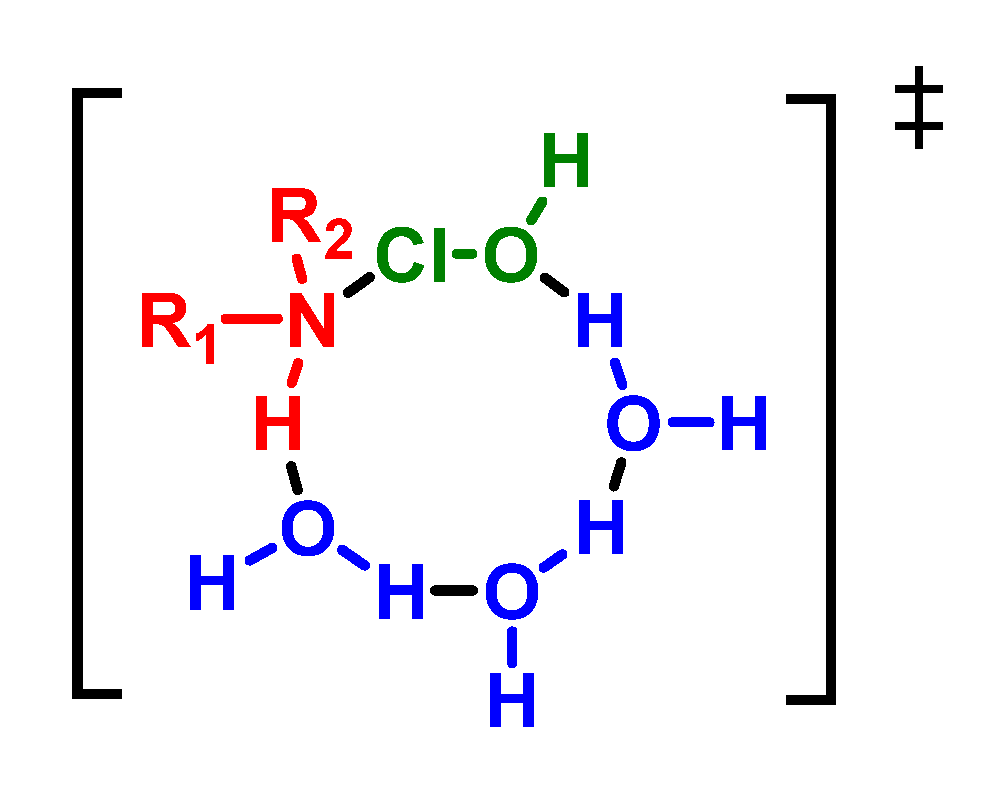
5. Reaction of HOCl with Ammonia and Aliphatic Amines
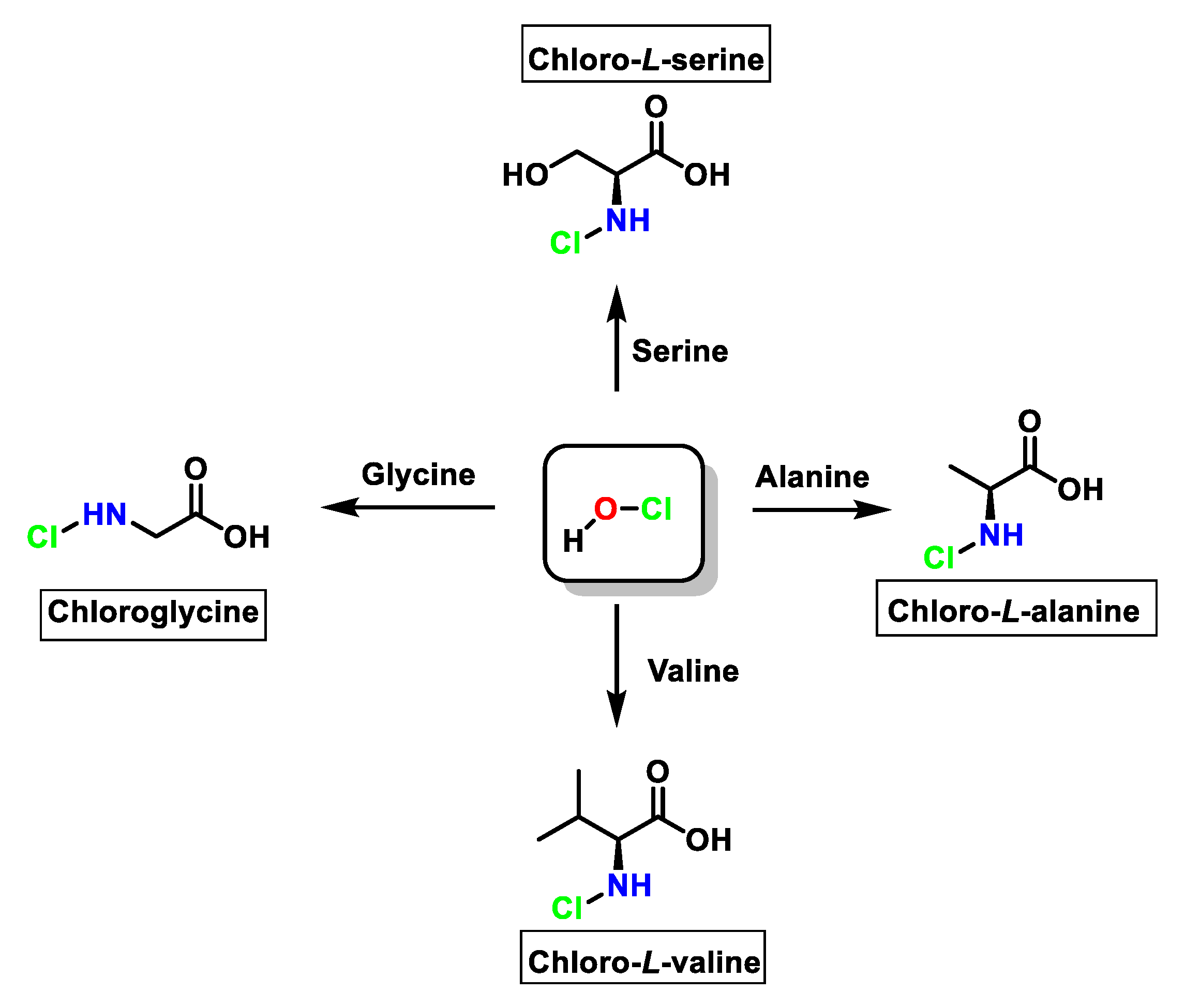
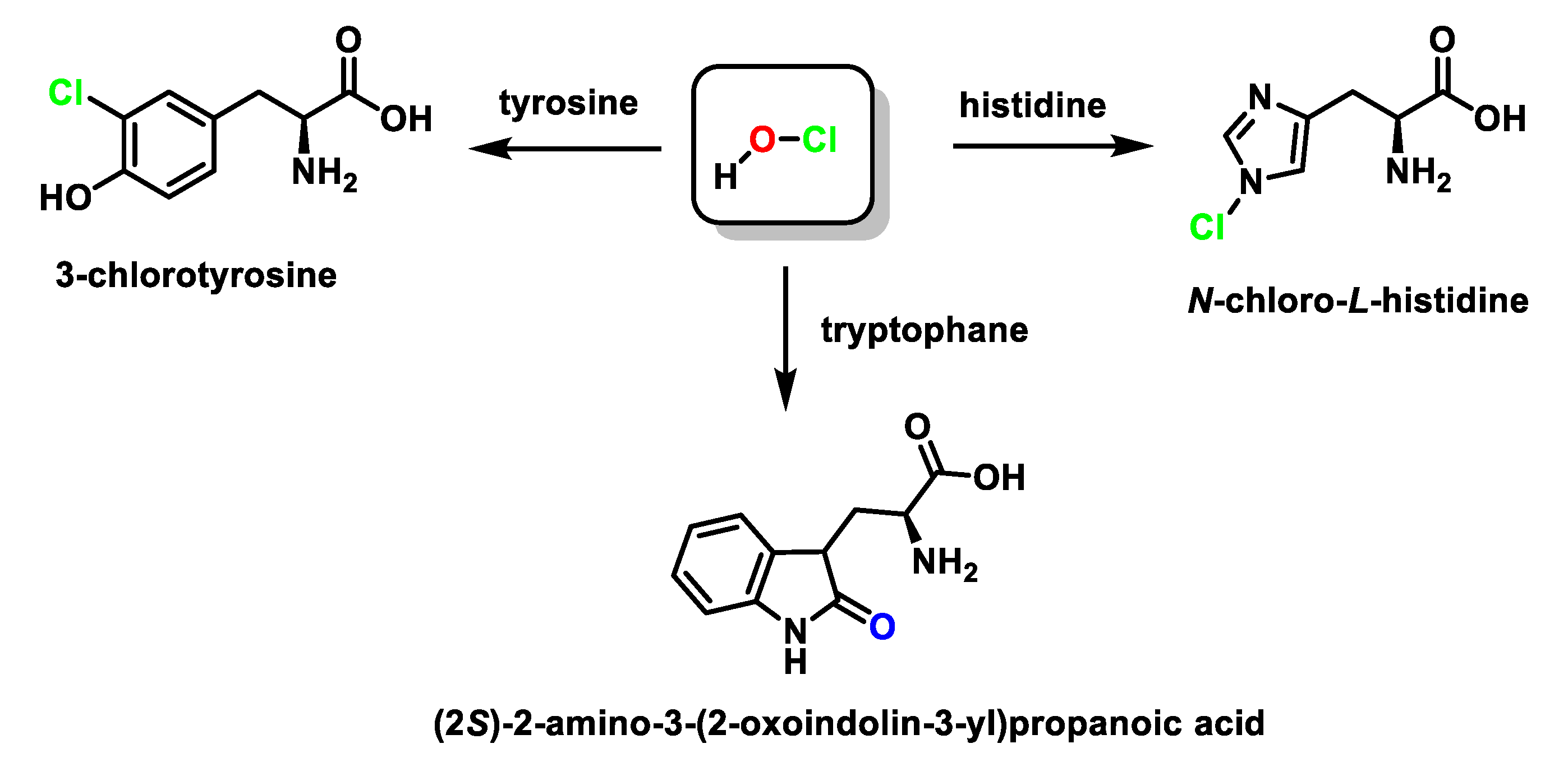
6. Reactions of HOCl with Amino Acids
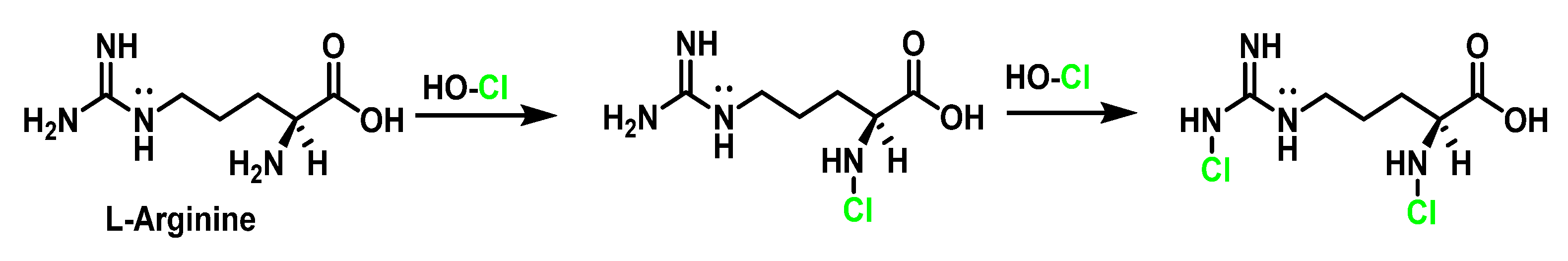
6.1. Reaction of HOCl with Nitrogen-Containing Compounds
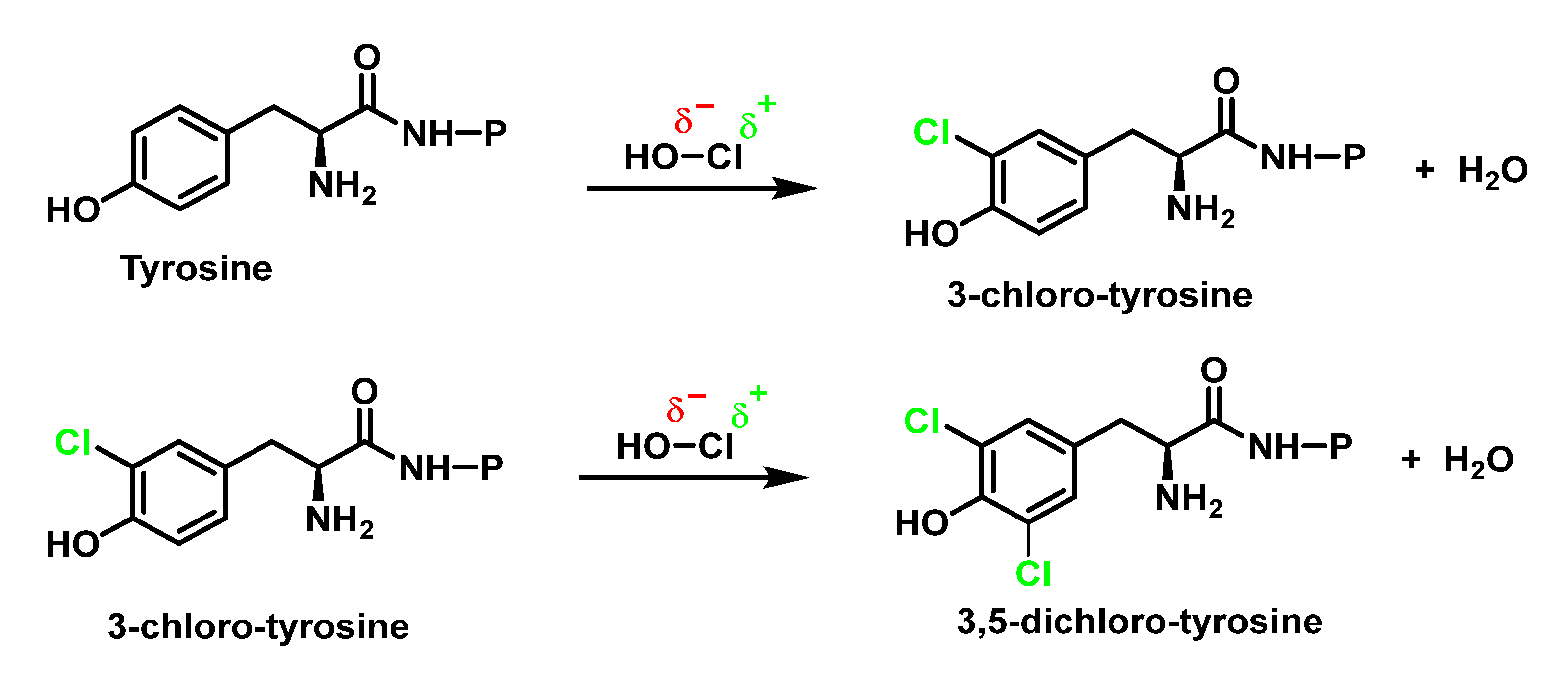
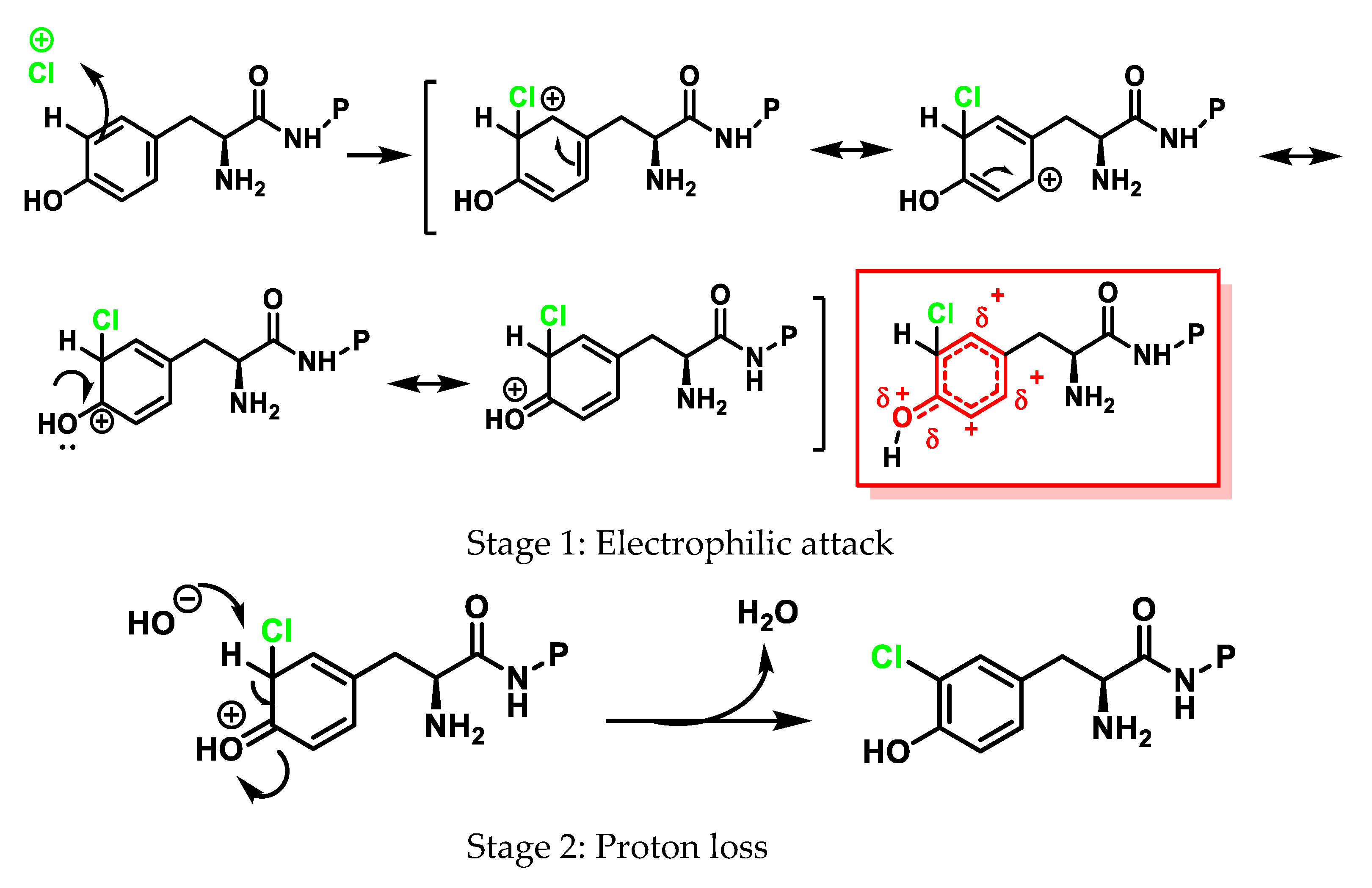
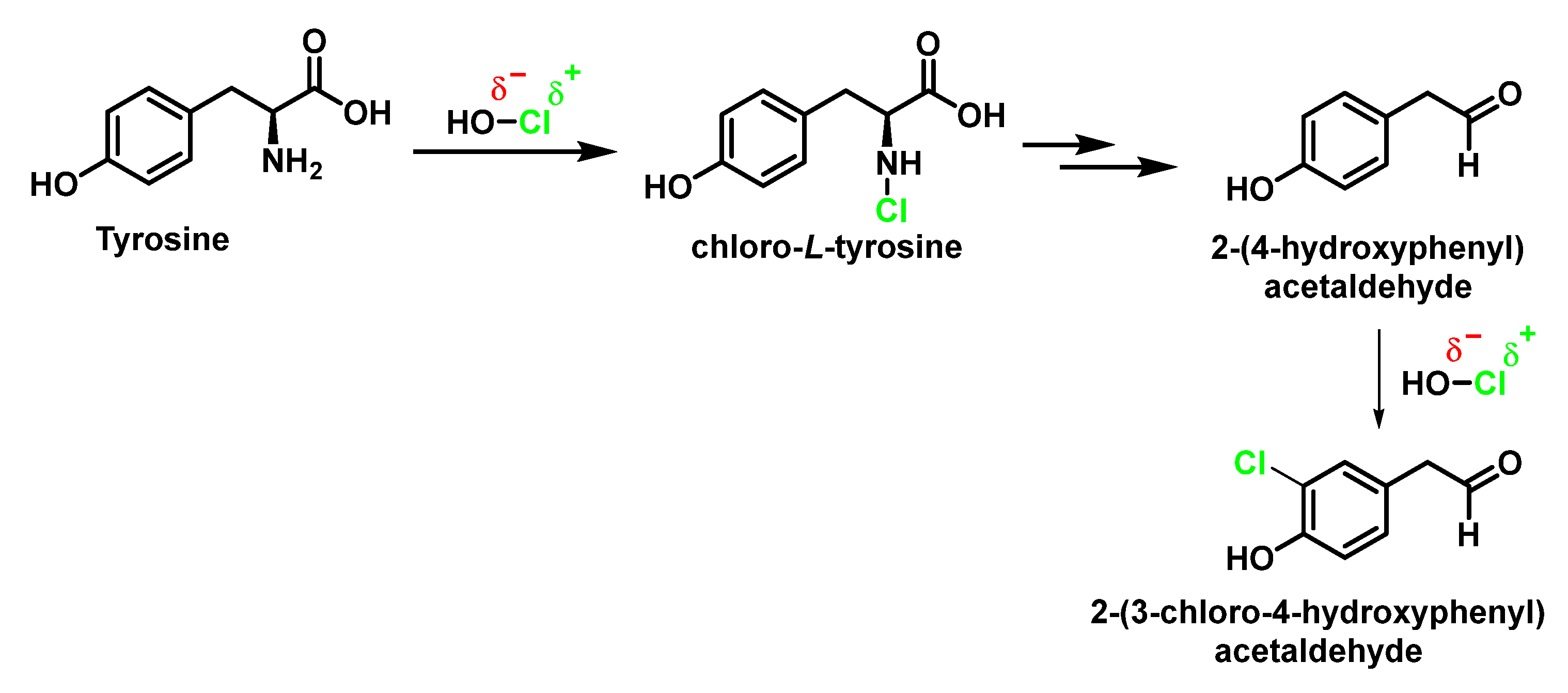
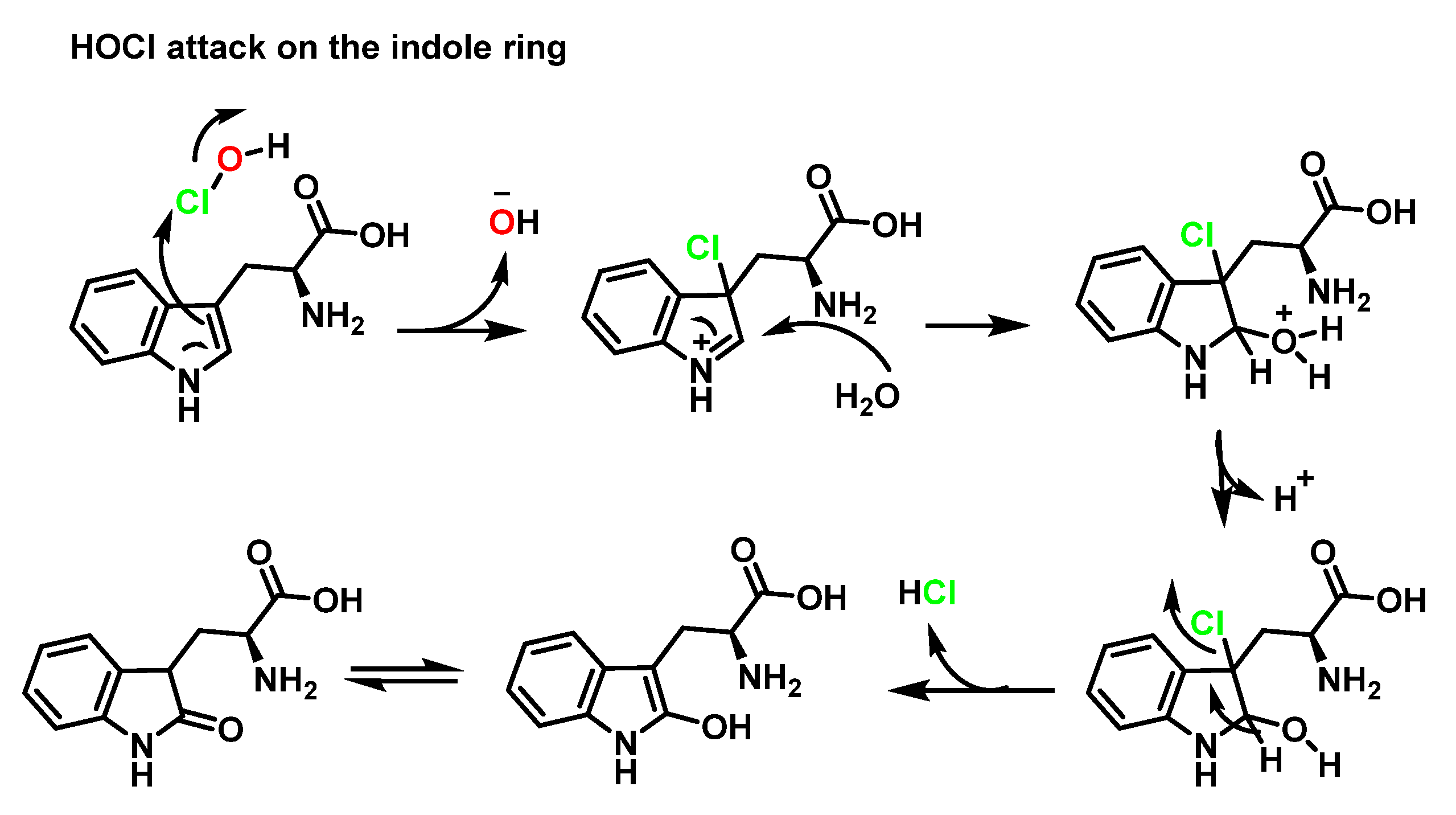
6.2. Reaction of HOCl with Aromatic Amino Acids
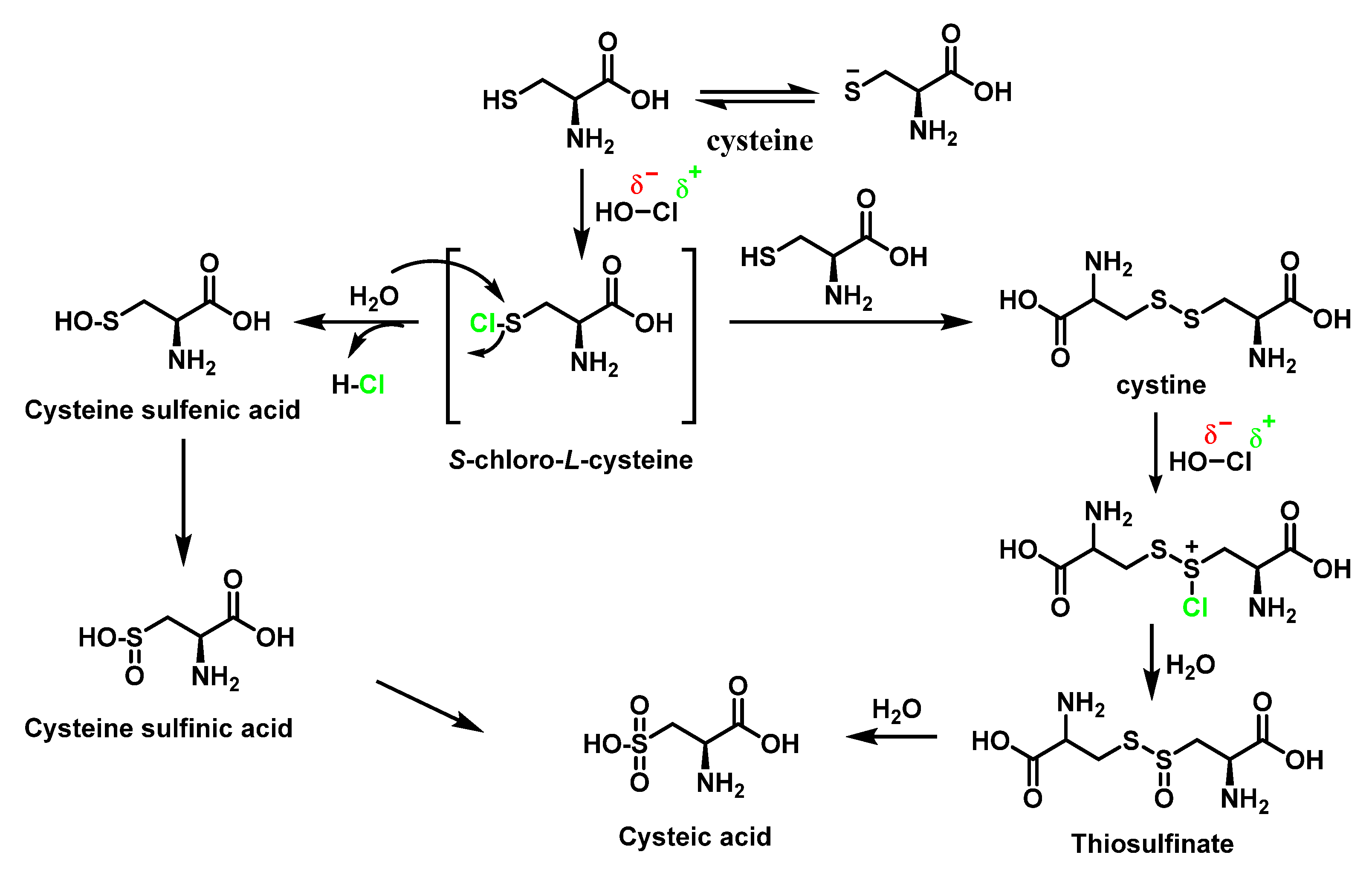
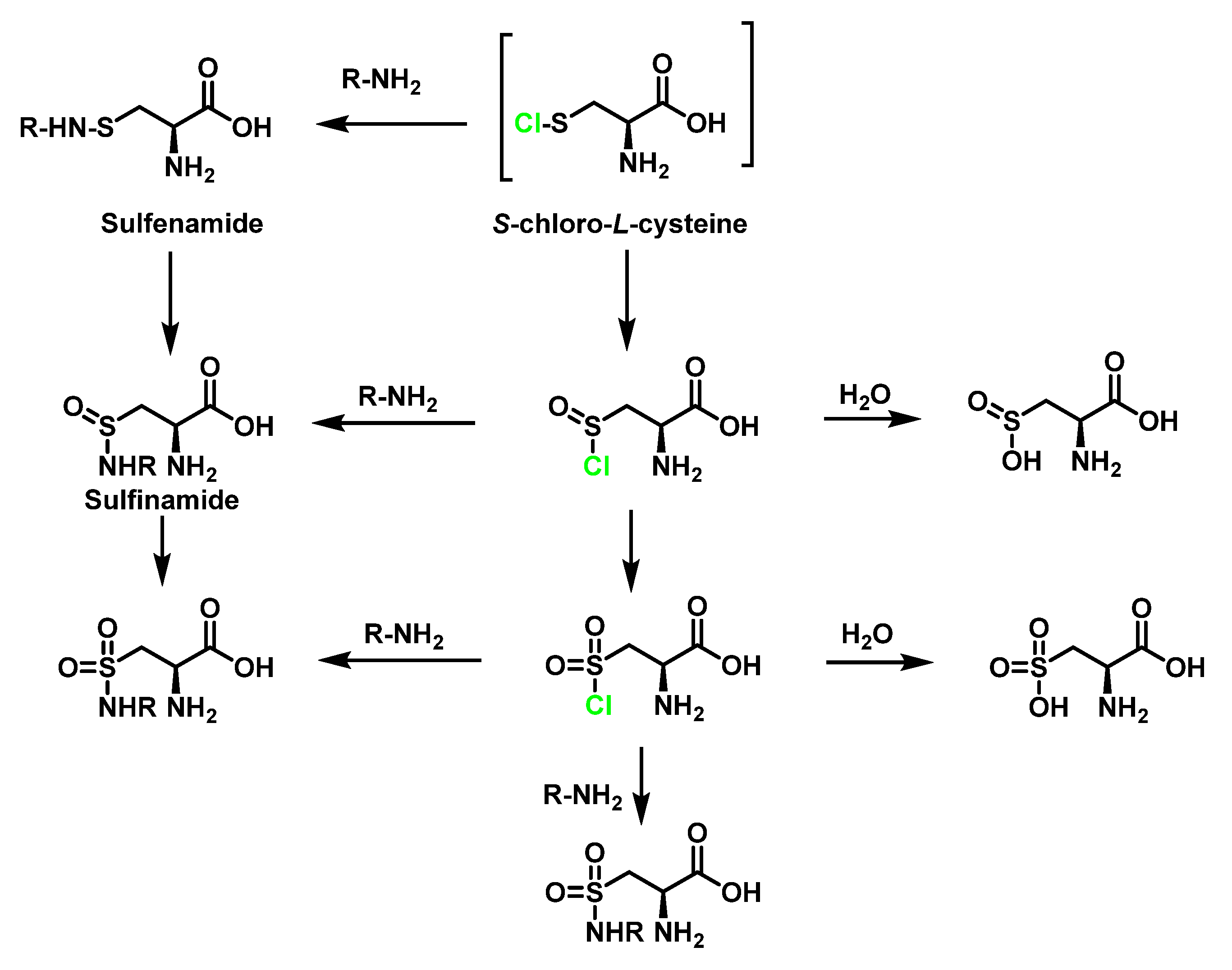
6.3. Reaction of HOCl with SULFUR-Containing Amino Acids
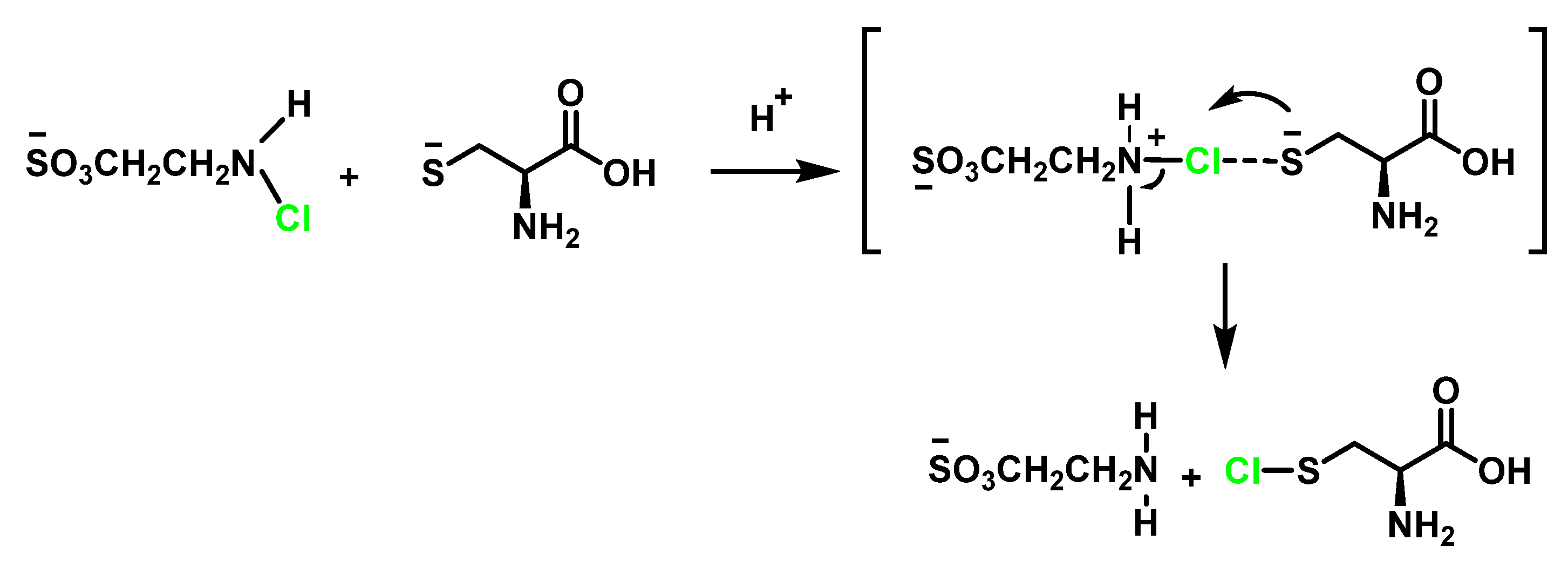
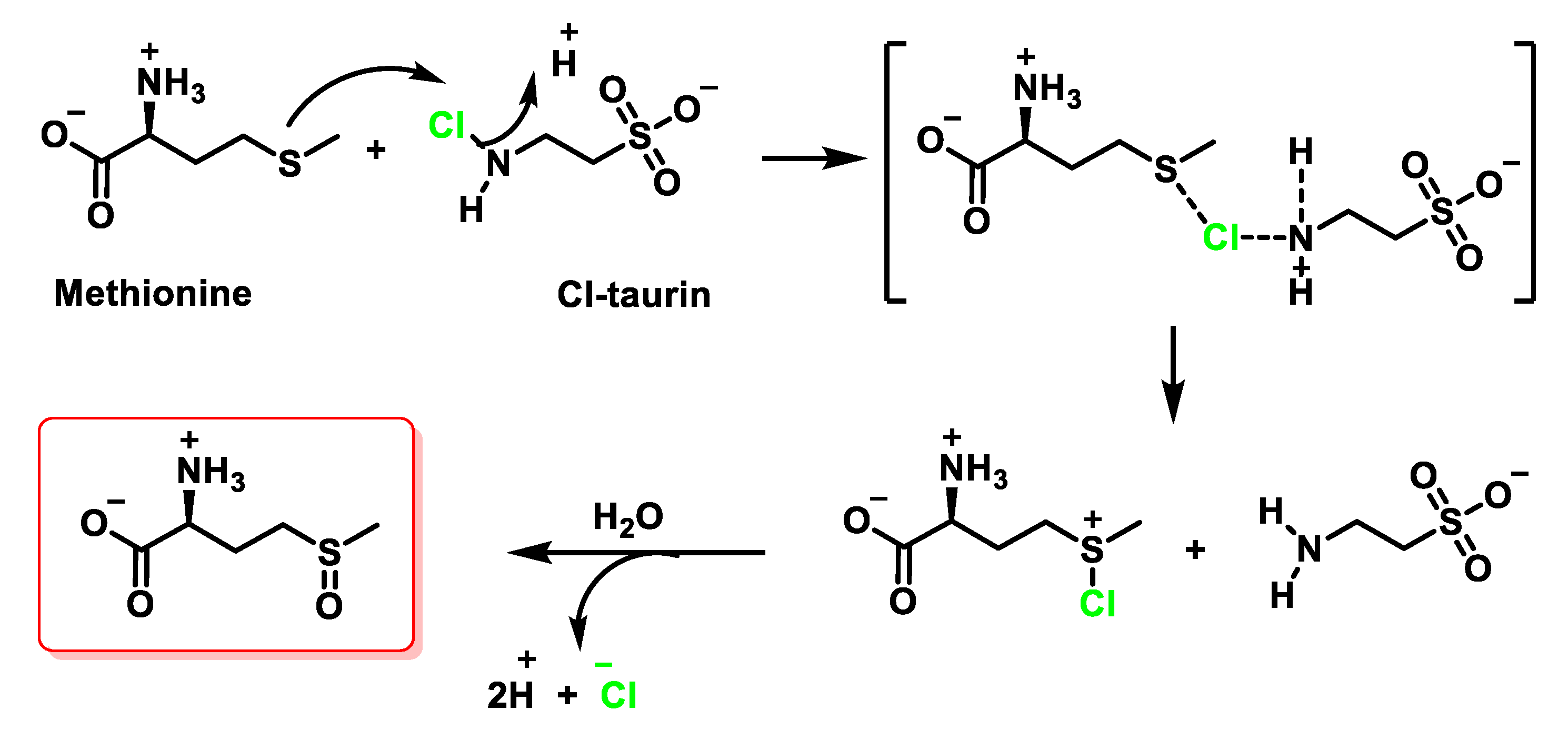
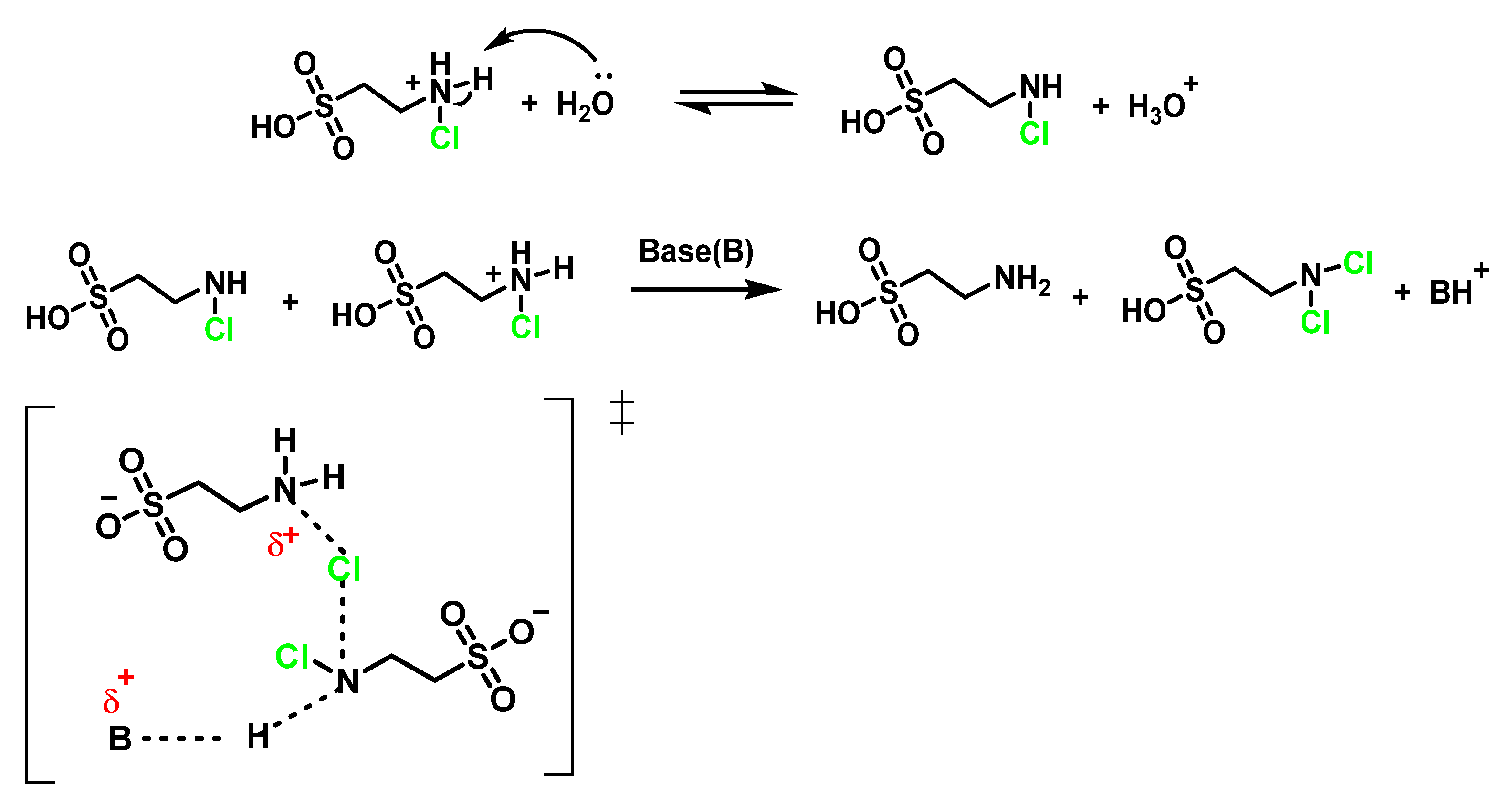
6.4. Reaction of HOCl with Taurine
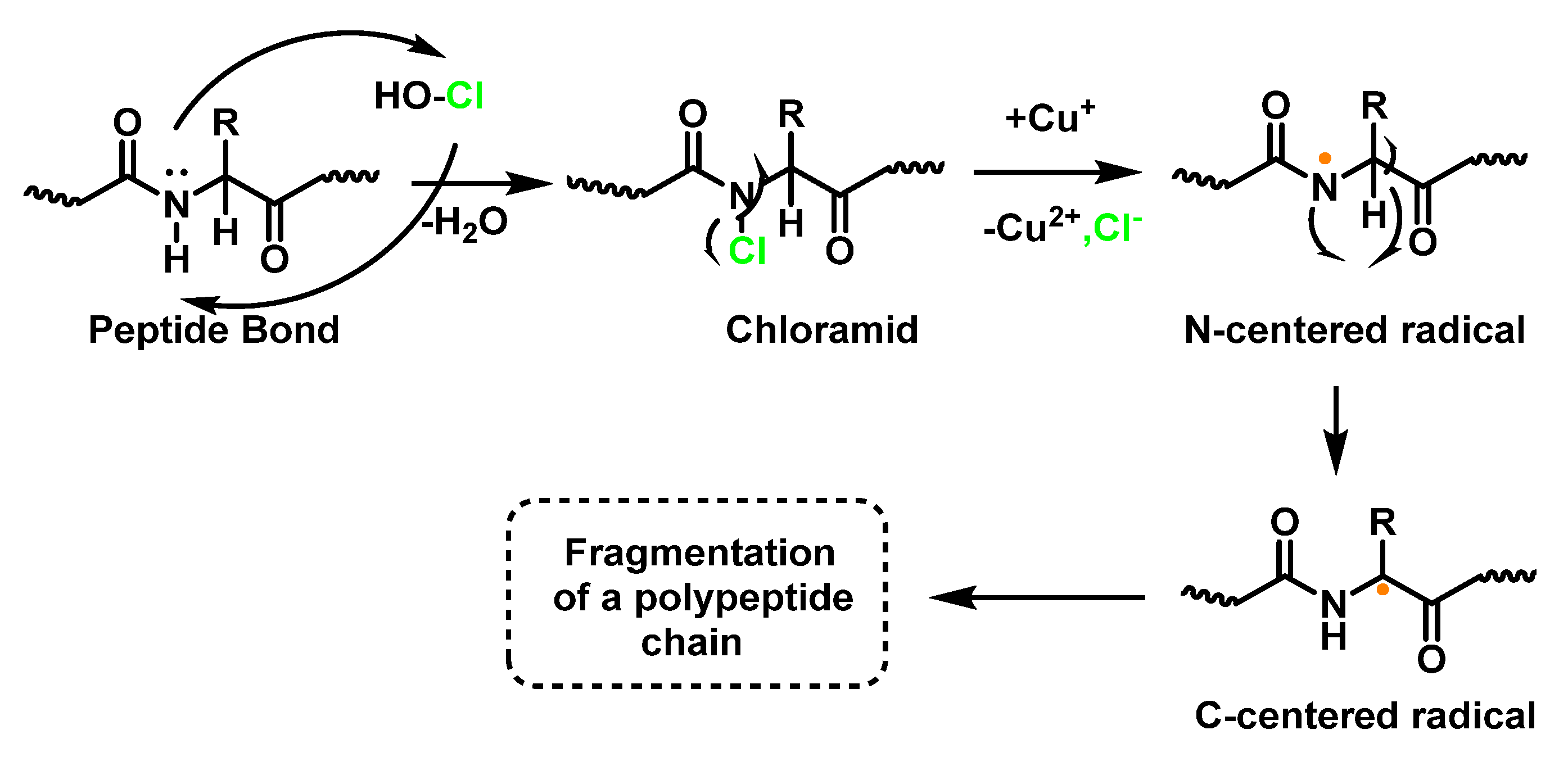
6.5. Reaction of HOCl with Peptides and Proteins
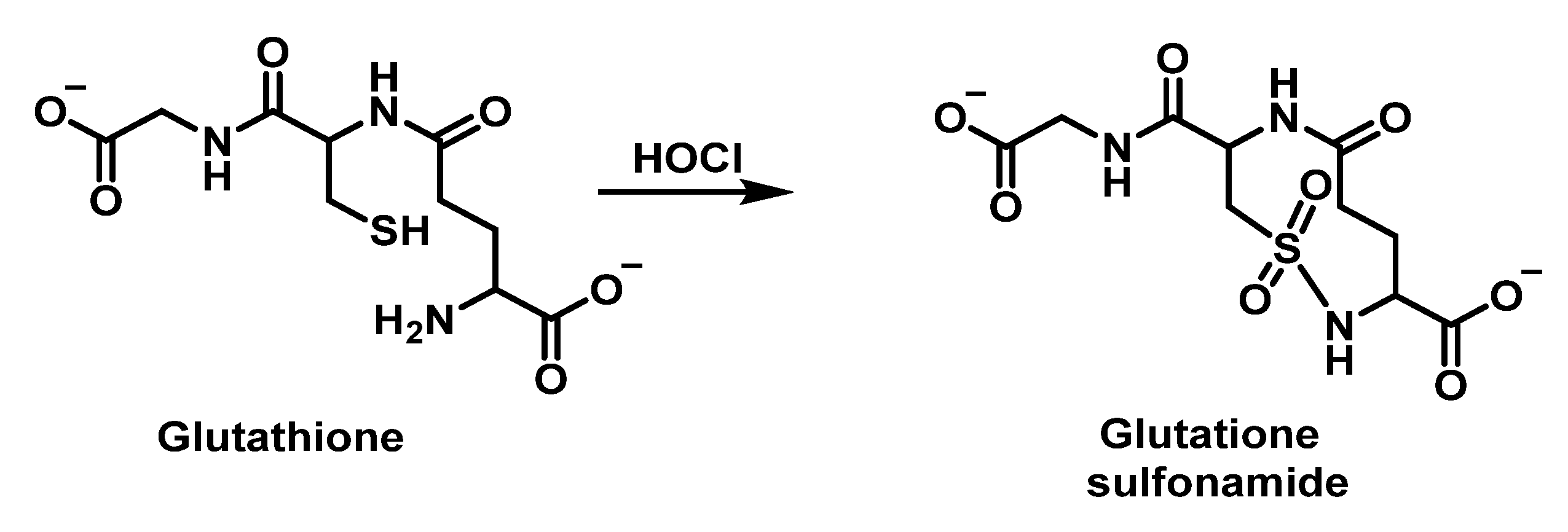
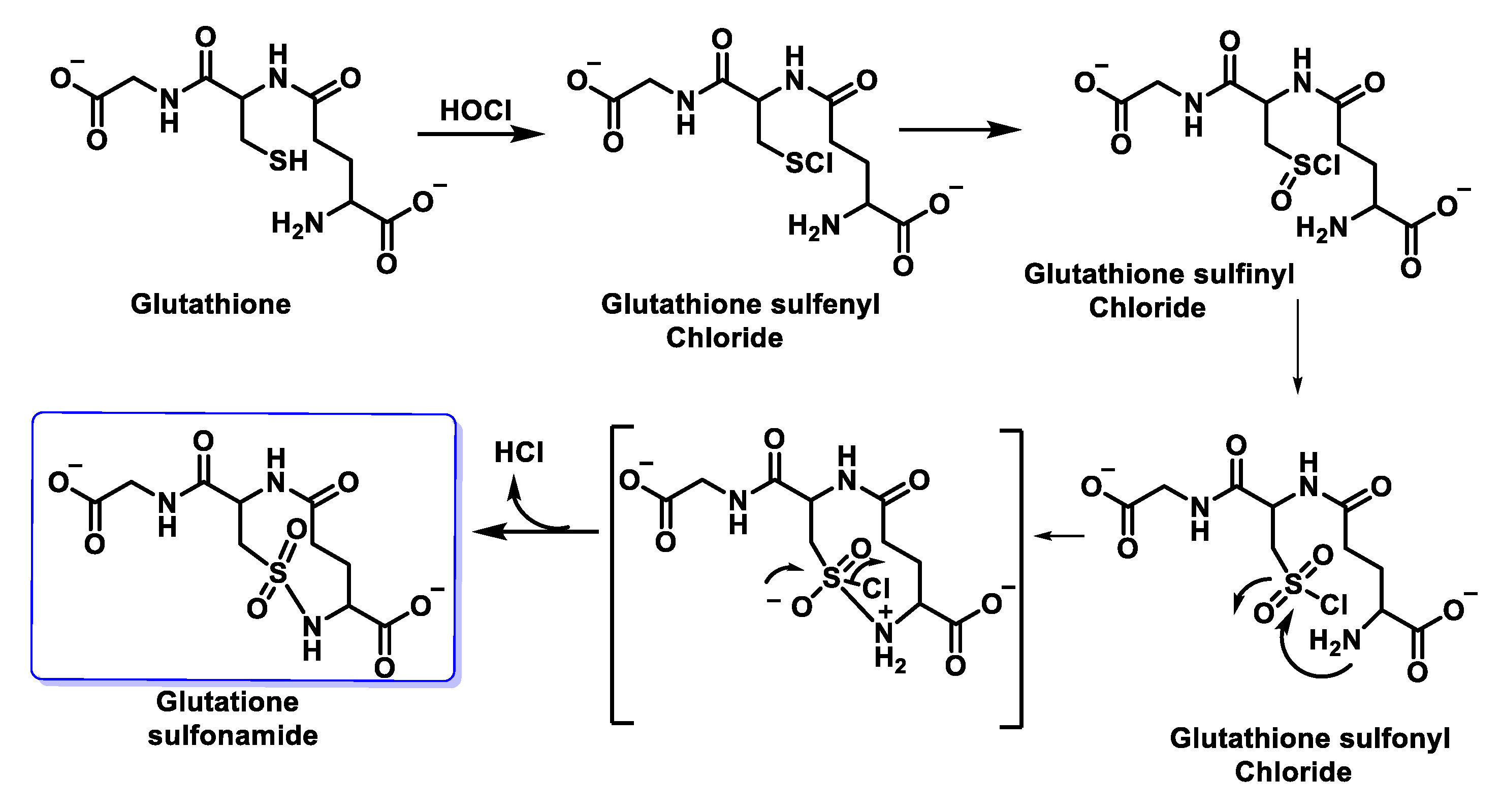
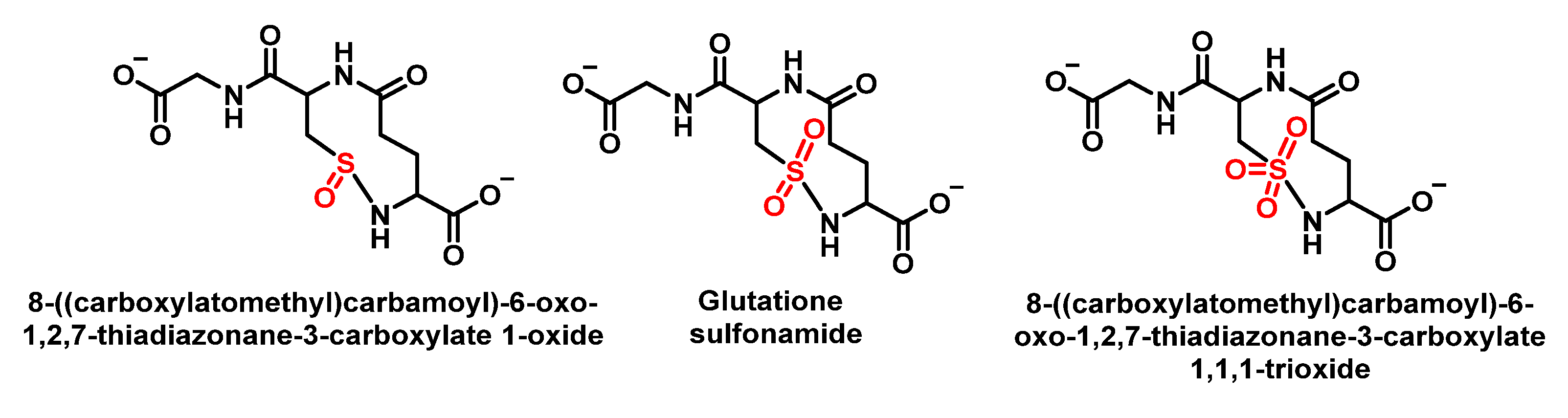
6.6. Reaction of HOCl with Glutathione
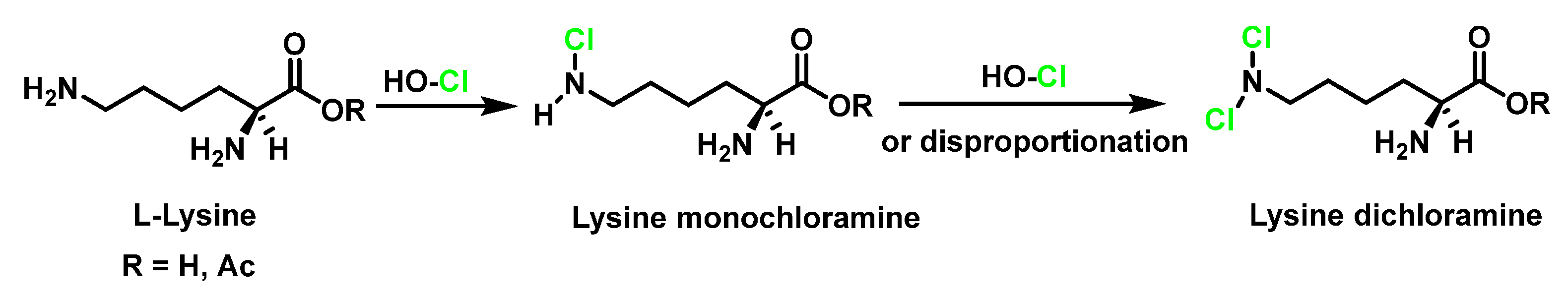
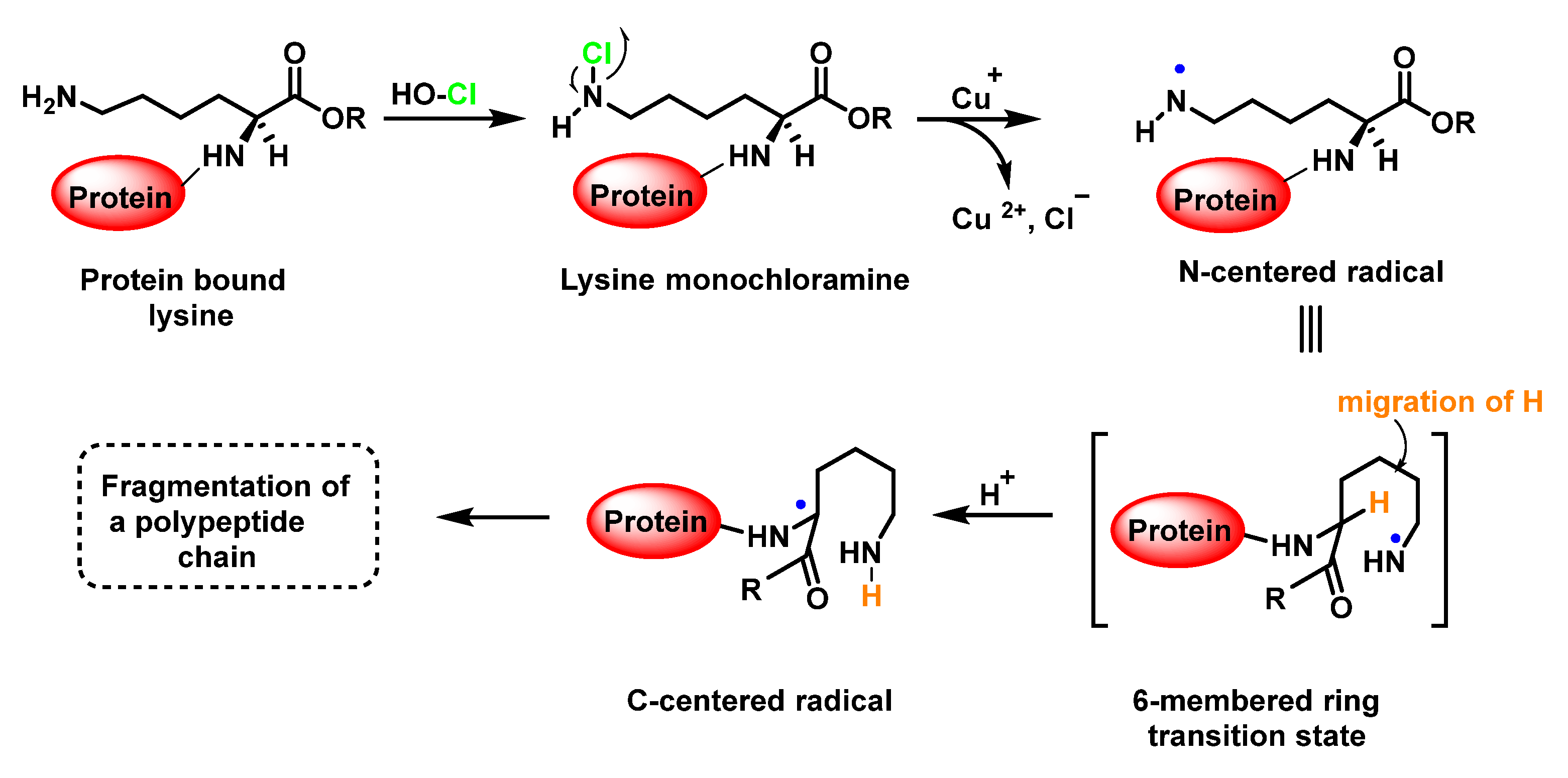
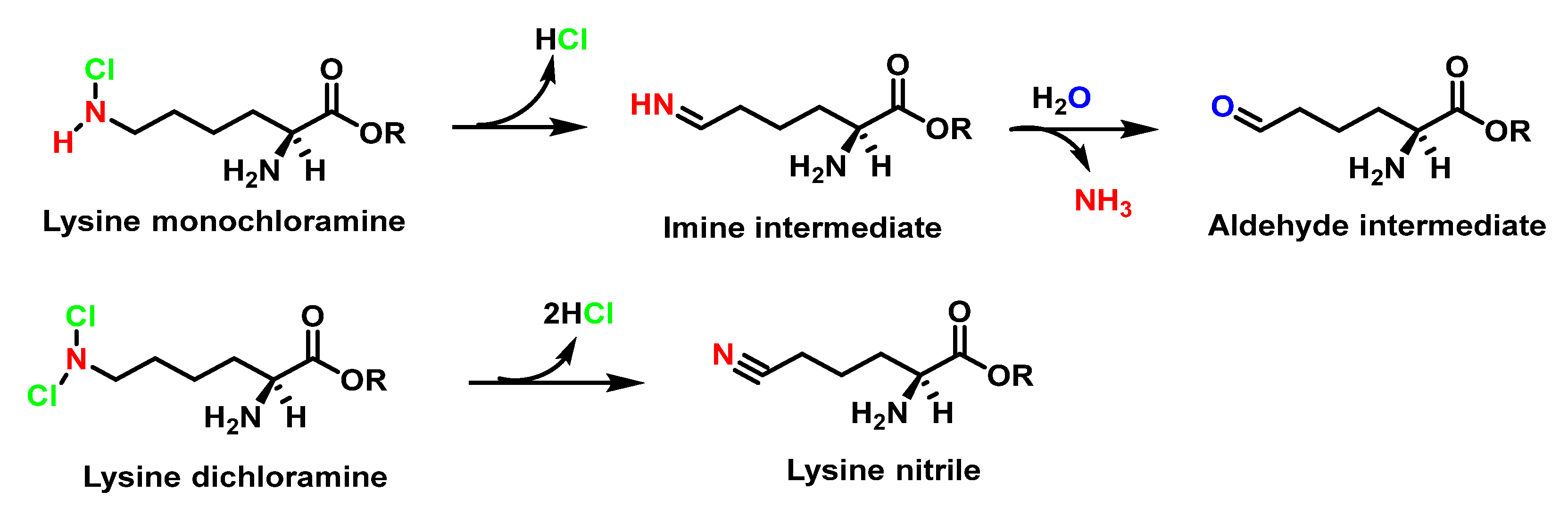
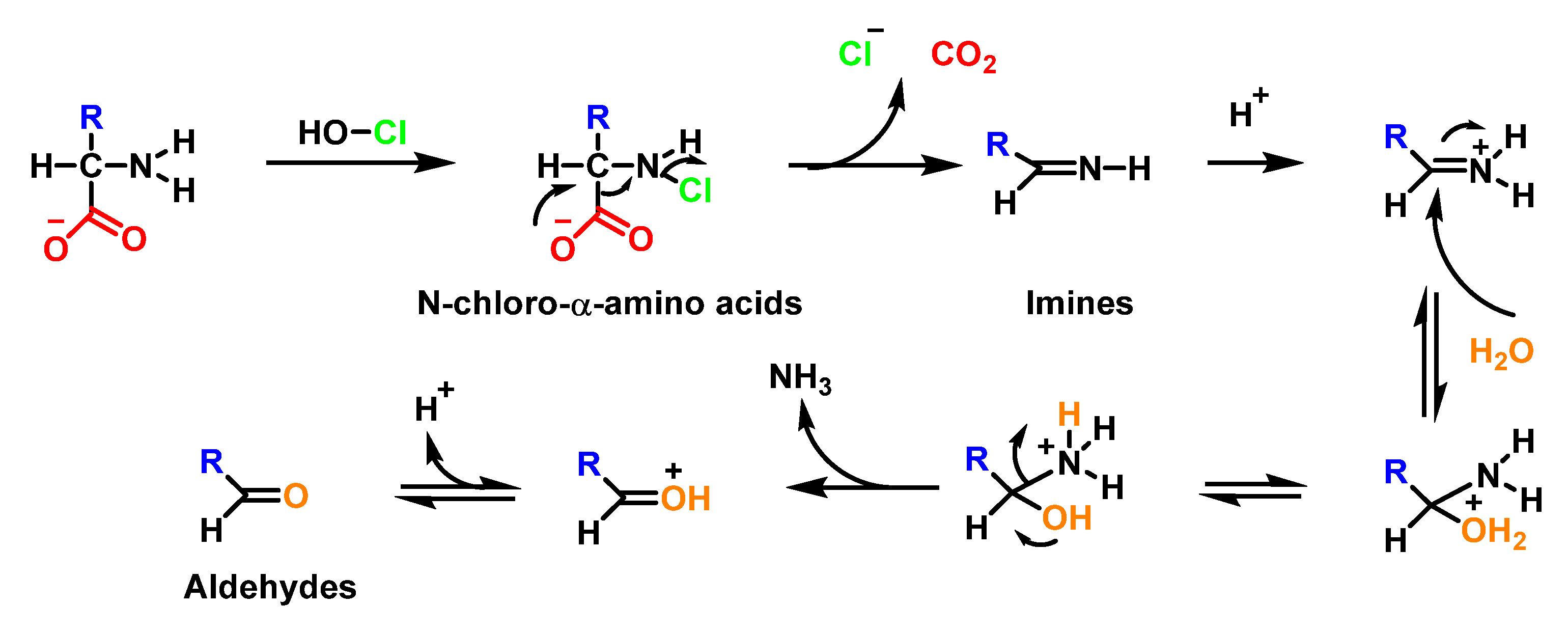
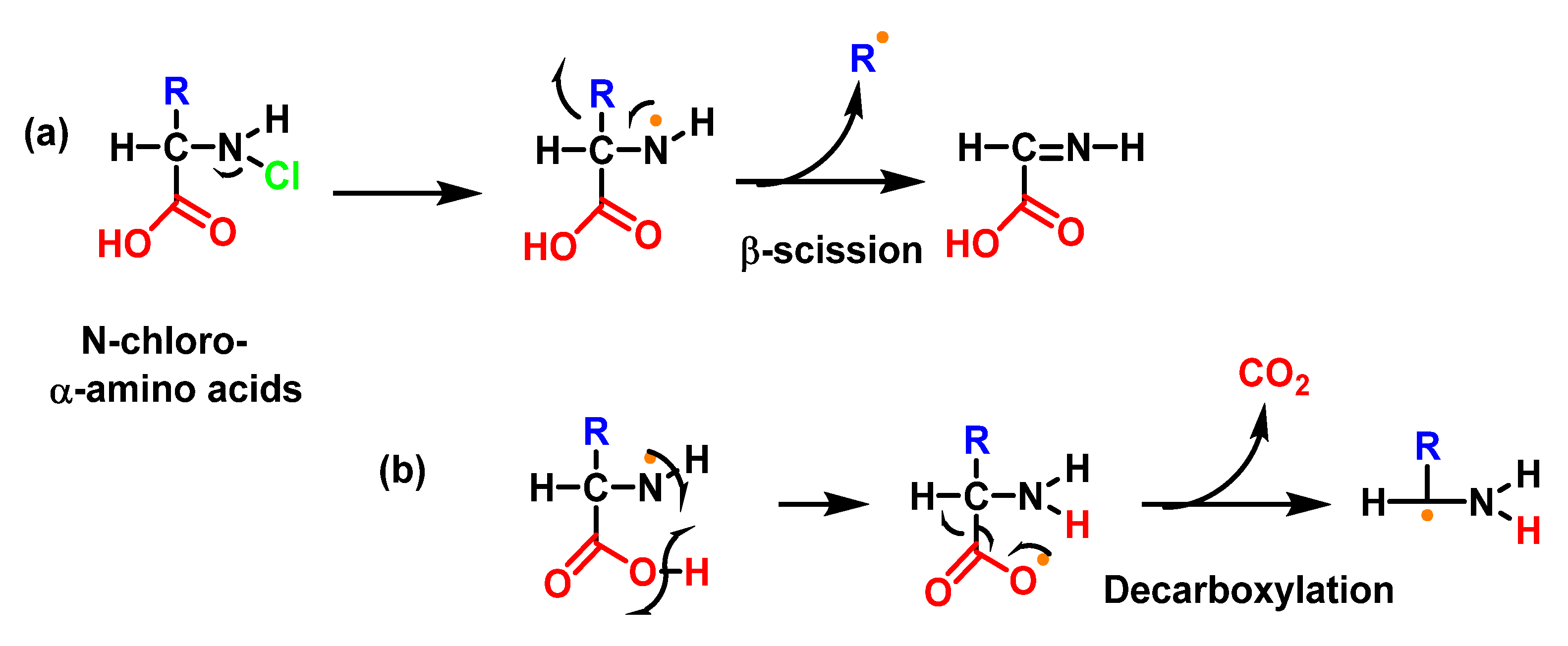
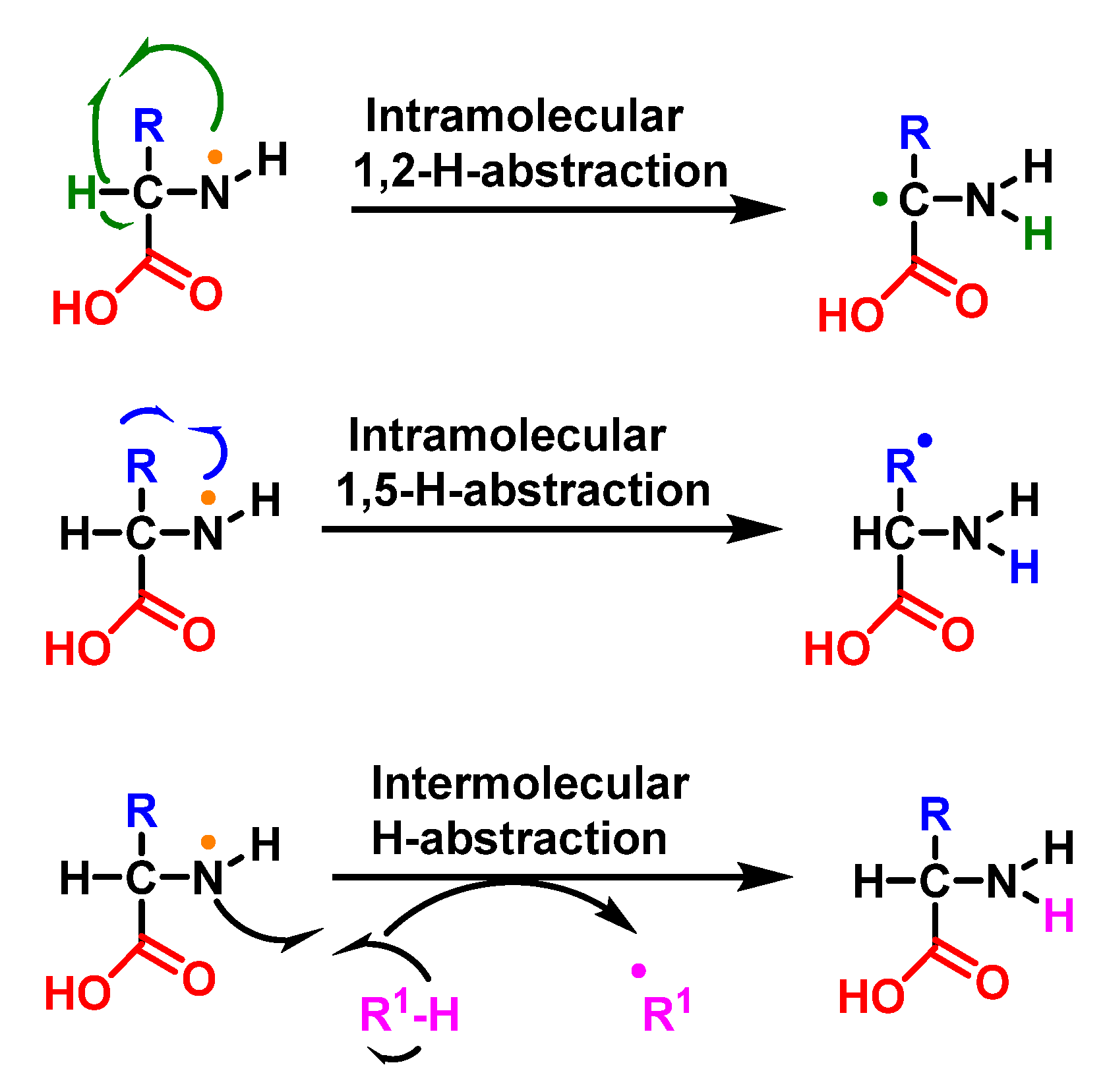
6.7. Degradation of Amines, N-Monochloro-α-Amino Acids, and Lys
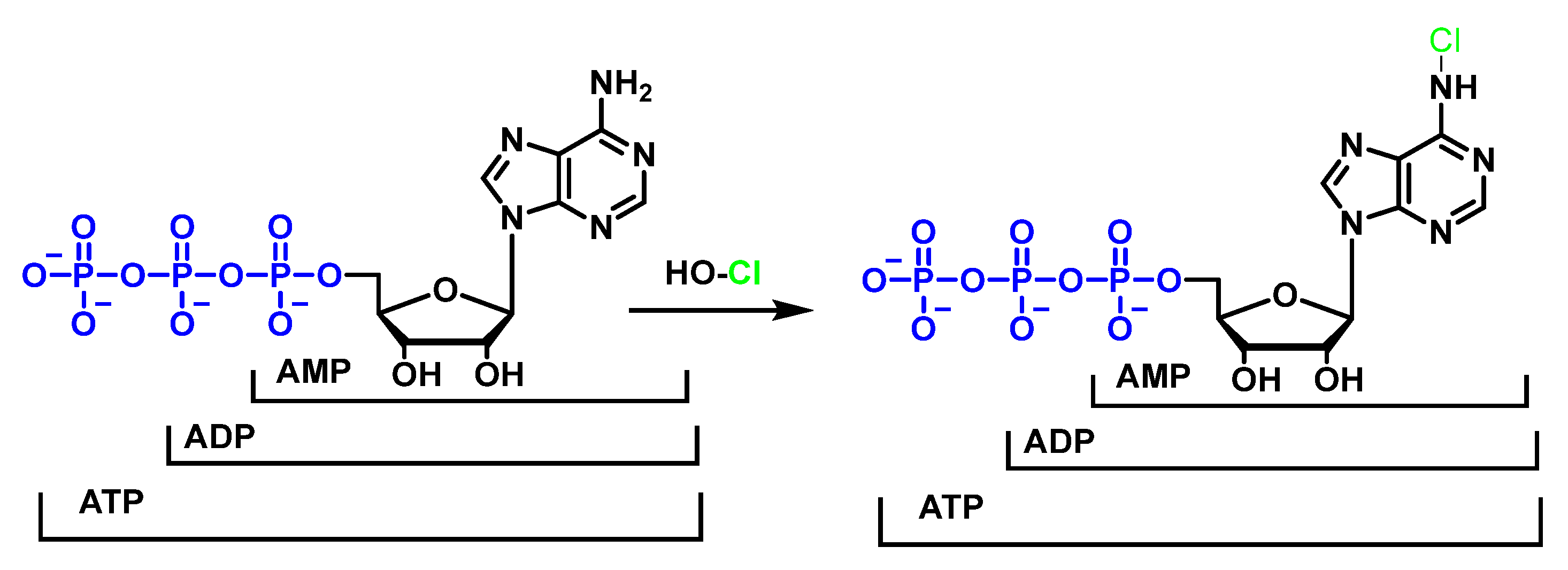
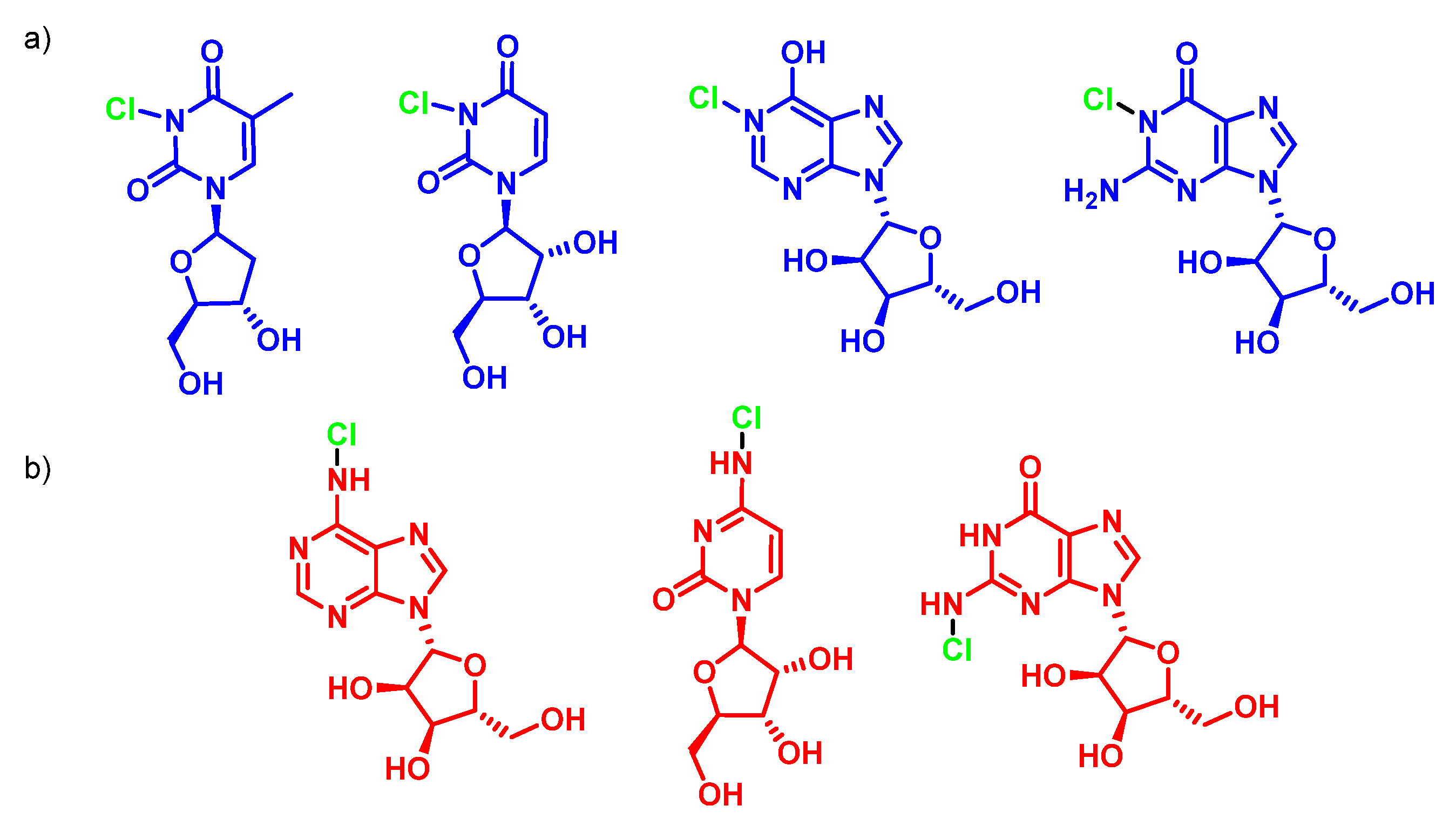
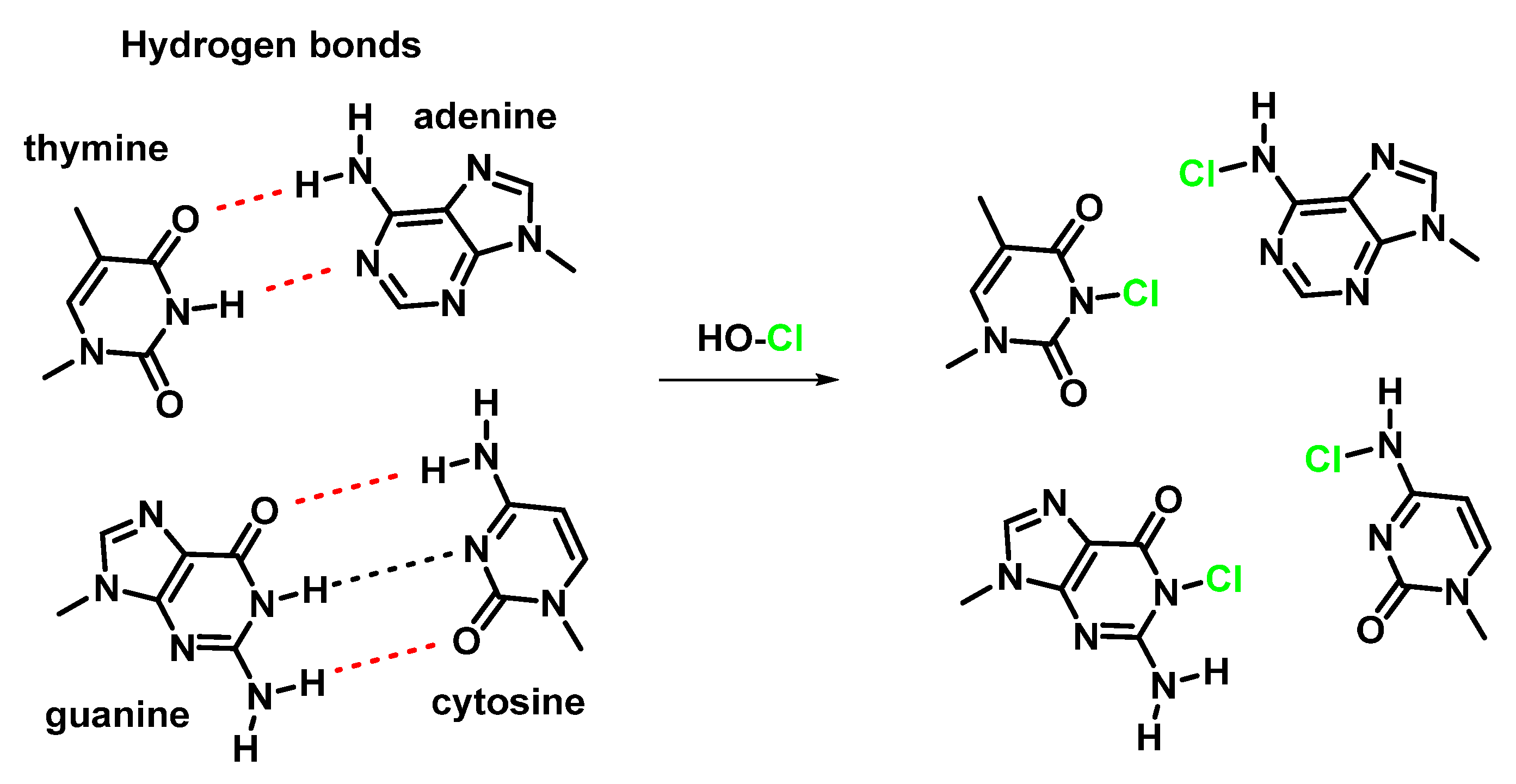
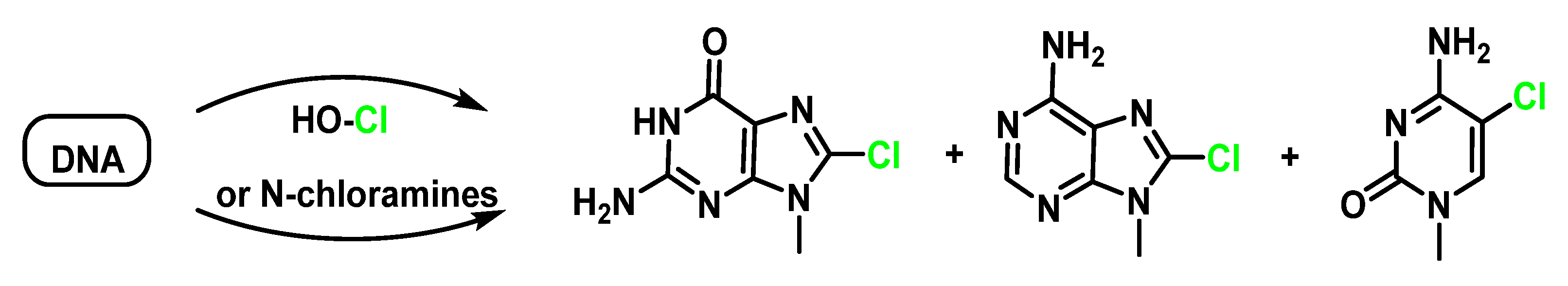
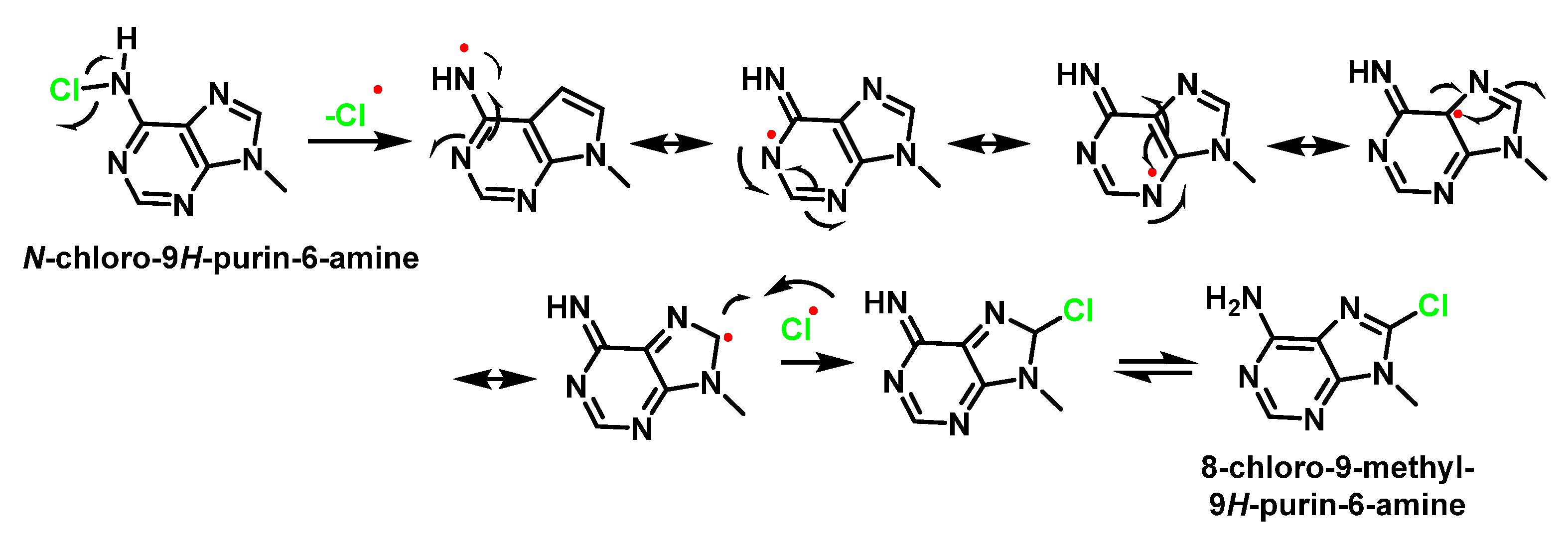
7. Reactions of HOCl with Nucleotides and Nucleic Acids
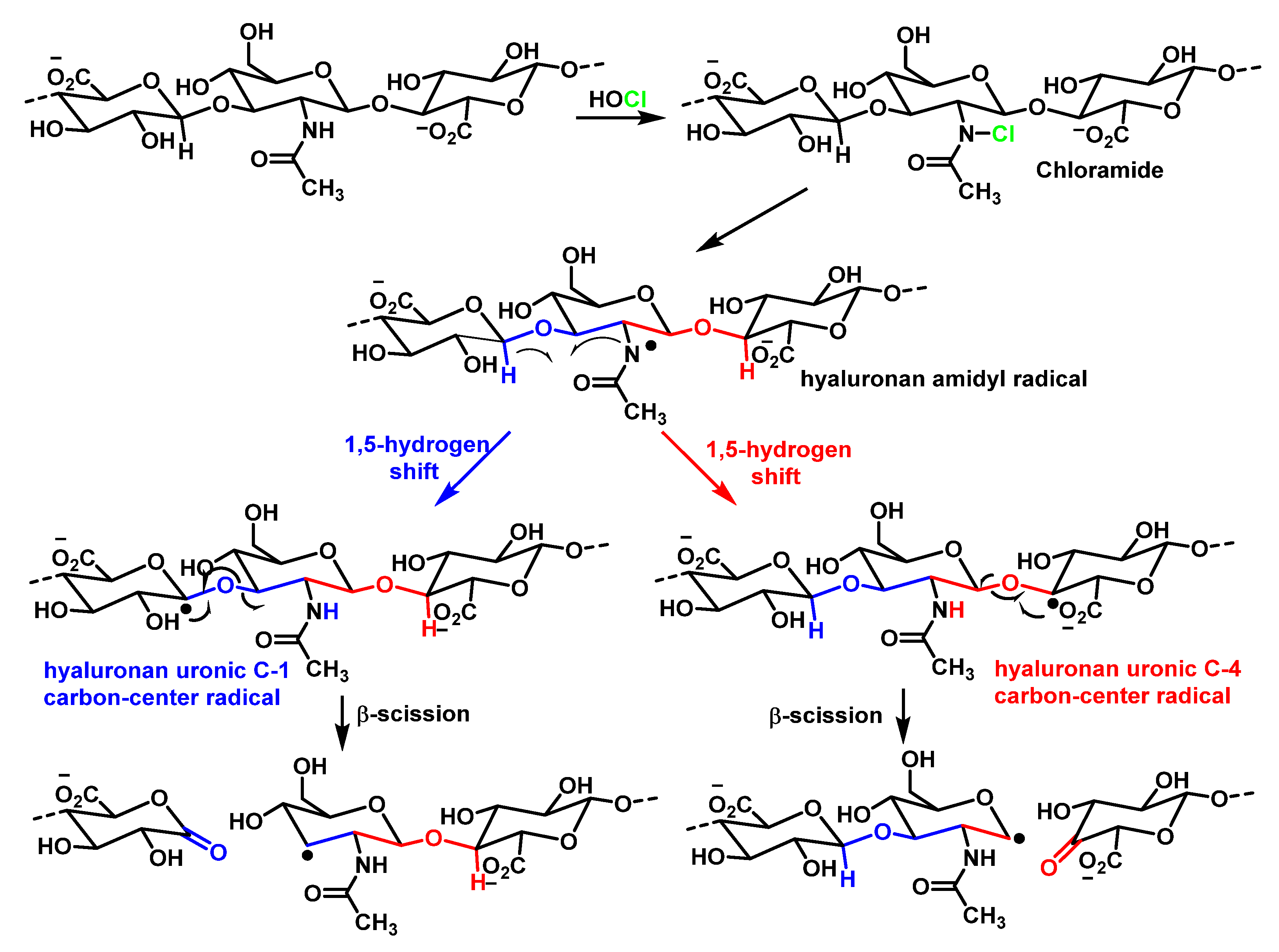
8. Reaction of HOCl with Carbohydrates
9. Role of MPO in Human Diseases and Inflammation
9.1. MPO in Human Diseases
9.2. MPO and Organ Inflammation
9.3. Summary Table of the Relationship between MPO and Different Diseases
9.4. Myeloperoxidase as a Disease Biomarker
9.5. Measurement of MPO Activity
9.6. Inhibitors of MPO
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- da Cruz Nizer, W.S.; Inkovskiy, V.; Overhage, J. Surviving reactive chlorine stress: Responses of gram-negative bacteria to hypochlorous acid. Microorganisms 2020, 8, 1220. [Google Scholar] [CrossRef] [PubMed]
- Kettle, A.; Winterbourn, C. Myeloperoxidase: A key regulator of neutrophil oxidant production. Redox Rep. 1997, 3, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Concept and some practical aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef]
- Malle, E.; Furtmüller, P.; Sattler, W.; Obinger, C. Myeloperoxidase: A target for new drug development? Br. J. Pharmacol. 2007, 152, 838–854. [Google Scholar] [CrossRef]
- Khan, A.A.; Alsahli, M.A.; Rahmani, A.H. Myeloperoxidase as an active disease biomarker: Recent biochemical and pathological perspectives. Med. Sci. 2018, 6, 33. [Google Scholar] [CrossRef]
- Hurst, J.K. What really happens in the neutrophil phagosome? Free Radic. Biol. Med. 2012, 53, 508–520. [Google Scholar] [CrossRef]
- Davies, M.J. Myeloperoxidase-derived oxidation: Mechanisms of biological damage and its prevention. J. Clin. Biochem. Nutr. 2010, 48, 8–19. [Google Scholar] [CrossRef]
- Shankar, S.; Mahadevan, A.; Satishchandra, P.; Uday Kumar, R.; Yasha, T.; Santosh, V.; Chandramuki, A.; Ravi, V.; Nath, A. Neuropathology of HIV/AIDS with an overview of the Indian scene. Indian J. Med. Res. 2005, 121, 468–488. [Google Scholar]
- Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Inside the neutrophil phagosome: Oxidants, myeloperoxidase, and bacterial killing. Blood J. Am. Soc. Hematol. 1998, 92, 3007–3017. [Google Scholar]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of protein modification by oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef]
- Lane, A.E.; Tan, J.T.; Hawkins, C.L.; Heather, A.K.; Davies, M.J. The myeloperoxidase-derived oxidant HOSCN inhibits protein tyrosine phosphatases and modulates cell signalling via the mitogen-activated protein kinase (MAPK) pathway in macrophages. Biochem. J. 2010, 430, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Ramos, D.R.; García, M.V.; Canle, M.; Santaballa, J.A.; Furtmüller, P.G.; Obinger, C. Myeloperoxidase-catalyzed chlorination: The quest for the active species. J. Inorg. Biochem. 2008, 102, 1300–1311. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Hampton, M.B.; Livesey, J.H.; Kettle, A.J. Modeling the reactions of superoxide and myeloperoxidase in the neutrophil phagosome: Implications for microbial killing. J. Biol. Chem. 2006, 281, 39860–39869. [Google Scholar] [CrossRef]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef]
- Leung, A.M.; LaMar, A.; He, X.; Braverman, L.E.; Pearce, E.N. Iodine status and thyroid function of Boston-area vegetarians and vegans. J. Clin. Endocrinol. Metab. 2011, 96, E1303–E1307. [Google Scholar] [CrossRef]
- Senthilmohan, R.; Kettle, A.J. Bromination and chlorination reactions of myeloperoxidase at physiological concentrations of bromide and chloride. Arch. Biochem. Biophys. 2006, 445, 235–244. [Google Scholar] [CrossRef]
- van Dalen, C.J.; Whitehouse, M.W.; Winterbourn, C.C.; Kettle, A.J. Thiocyanate and chloride as competing substrates for myeloperoxidase. Biochem. J. 1997, 327, 487–492. [Google Scholar] [CrossRef]
- Zámocký, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent evolution of four heme peroxidase superfamilies. Arch Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef]
- Love, D.T.; Barrett, T.J.; White, M.Y.; Cordwell, S.J.; Davies, M.J.; Hawkins, C.L. Cellular targets of the myeloperoxidase-derived oxidant hypothiocyanous acid (HOSCN) and its role in the inhibition of glycolysis in macrophages. Free Radic. Biol. Med. 2016, 94, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Dunford, H.B.; Araiso, T.; Job, D.; Ricard, J.; Rutter, R.; Hager, L.P.; Wever, R.; Kast, W.M.; Boelens, R.; Ellfolk, N.; et al. Peroxidases; Springer: Dordrecht, The Netherlands, 1982; pp. 337–355. [Google Scholar]
- McKenna, S.M.; Davies, K.J. The inhibition of bacterial growth by hypochlorous acid. Possible role in the bactericidal activity of phagocytes. Biochem. J. 1988, 254, 685–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storkey, C.; Davies, M.J.; Pattison, D.I. Reevaluation of the rate constants for the reaction of hypochlorous acid (HOCl) with cysteine, methionine, and peptide derivatives using a new competition kinetic approach. Free Radic. Biol. Med. 2014, 73, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Schuller-Levis, G.B.; Park, E. Taurine and its chloramine: Modulators of immunity. Neurochem. Res. 2004, 29, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw. 2018, 18, e27. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, R.J.; Rawn, J.D. Organic Chemistry Study Guide: Key Concepts, Problems, and Solutions; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Winterbourn, C.C. Biological reactivity and biomarkers of the neutrophil oxidant, hypochlorous acid. Toxicology 2002, 181, 223–227. [Google Scholar] [CrossRef]
- Morris, J.C. The acid ionization constant of HOCl from 5 to 35. J. Phys. Chem. 1966, 70, 3798–3805. [Google Scholar] [CrossRef]
- Wang, L.; Bassiri, M.; Najafi, R.; Najafi, K.; Yang, J.; Khosrovi, B.; Hwong, W.; Barati, E.; Belisle, B.; Celeri, C. Hypochlorous acid as a potential wound care agent: Part I. Stabilized hypochlorous acid: A component of the inorganic armamentarium of innate immunity. J. Burn. Wounds 2007, 6, e5. [Google Scholar]
- Mütze, S.; Hebling, U.; Stremmel, W.; Wang, J.; Arnhold, J.r.; Pantopoulos, K.; Mueller, S. Myeloperoxidase-derived hypochlorous acid antagonizes the oxidative stress-mediated activation of iron regulatory protein 1. J. Biol. Chem. 2003, 278, 40542–40549. [Google Scholar]
- Weiss, S.J. Tissue destruction by neutrophils. N. Engl. J. Med. 1989, 320, 365–376. [Google Scholar]
- White, G.C. The Handbook of Chlorination and Alternative Disinfectant; Van Nostrand Reinhold International: London, UK, 1992. [Google Scholar]
- Ward, J.F. DNA damage produced by ionizing radiation in mammalian cells: Identities, mechanisms of formation, and reparability. Prog. Nucleic Acid Res. Mol. Biol. 1988, 35, 95–125. [Google Scholar]
- Stanley, N.R.; Pattison, D.I.; Hawkins, C.L. Ability of hypochlorous acid and N-chloramines to chlorinate DNA and its constituents. Chem. Res. Toxicol. 2010, 23, 1293–1302. [Google Scholar] [CrossRef]
- Del Rosso, J.Q.; Bhatia, N. Status report on topical hypochlorous acid: Clinical relevance of specific formulations, potential modes of action, and study outcomes. J. Clin. Aesthetic Dermatol. 2018, 11, 36. [Google Scholar]
- Xu, Q.; Lee, K.A.; Lee, S.; Lee, K.M.; Lee, W.J.; Yoon, J. A highly specific fluorescent probe for hypochlorous acid and its application in imaging microbe-induced HOCl production. J. Am. Chem. Soc. 2013, 135, 9944–9949. [Google Scholar] [CrossRef]
- Ford, D.A. Lipid oxidation by hypochlorous acid: Chlorinated lipids in atherosclerosis and myocardial ischemia. Clin. Lipidol. 2010, 5, 835–852. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J. Biomarkers of myeloperoxidase-derived hypochlorous acid. Free Radic. Biol. Med. 2000, 29, 403–409. [Google Scholar] [CrossRef]
- Schröter, J.; Schiller, J. Chlorinated phospholipids and fatty acids:(patho) physiological relevance, potential toxicity, and analysis of lipid chlorohydrins. Oxidative Med. Cell. Longev. 2016, 2016, 8386362. [Google Scholar] [CrossRef] [Green Version]
- Arnhold, J.; Osipov, A.N.; Spalteholz, H.; Panasenko, O.M.; Schiller, J. Formation of lysophospholipids from unsaturated phosphatidylcholines under the influence of hypochlorous acid. Biochim. Biophys. Acta BBA Gen. Subj. 2002, 1572, 91–100. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; van den Berg, J.J.; Roitman, E.; Kuypers, F.A. Chlorohydrin formation from unsaturated fatty acids reacted with hypochlorous acid. Arch. Biochem. Biophys. 1992, 296, 547–555. [Google Scholar] [CrossRef]
- Heinecke, J.W.; Li, W.; Mueller, D.M.; Bohrer, A.; Turk, J. Cholesterol chlorohydrin synthesis by the myeloperoxidase-hydrogen peroxide-chloride system: Potential markers for lipoproteins oxidatively damaged by phagocytes. Biochemistry 1994, 33, 10127–10136. [Google Scholar] [CrossRef]
- Carr, A.C.; van den Berg, J.J.; Winterbourn, C.C. Chlorination of cholesterol in cell membranes by hypochlorous acid. Arch. Biochem. Biophys. 1996, 332, 63–69. [Google Scholar] [CrossRef]
- Hazen, S.L.; Hsu, F.F.; Duffin, K.; Heinecke, J.W. Molecular chlorine generated by the myeloperoxidase-hydrogen peroxide-chloride system of phagocytes converts low density lipoprotein cholesterol into a family of chlorinated sterols. J. Biol. Chem. 1996, 271, 23080–23088. [Google Scholar] [CrossRef]
- Shao, B.; Oda, M.N.; Oram, J.F.; Heinecke, J.W. Myeloperoxidase: An oxidative pathway for generating dysfunctional high-density lipoprotein. Chem. Res. Toxicol. 2010, 23, 447–454. [Google Scholar] [CrossRef]
- Panasenko, O.; Torkhovskaya, T.; Gorudko, I.; Sokolov, A. The role of halogenative stress in atherogenic modification of low-density lipoproteins. Biochemistry 2020, 85, 34–55. [Google Scholar] [CrossRef]
- Spickett, C.M.; Jerlich, A.; Panasenko, O.M.; Arnhold, J.; Pitt, A.R.; Stelmaszyńska, T.; Schaur, R.J. The reactions of hypochlorous acid, the reactive oxygen species produced by myeloperoxidase, with lipids. Acta Biochim. Pol. 2000, 47, 889–899. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Subbaiah, P.V. Regulation of hepatic lipase activity by sphingomyelin in plasma lipoproteins. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2015, 1851, 1327–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusshold, C.; Kollroser, M.; Köfeler, H.; Rechberger, G.; Reicher, H.; Üllen, A.; Bernhart, E.; Waltl, S.; Kratzer, I.; Hermetter, A. Hypochlorite modification of sphingomyelin generates chlorinated lipid species that induce apoptosis and proteome alterations in dopaminergic PC12 neurons in vitro. Free Radic. Biol. Med. 2010, 48, 1588–1600. [Google Scholar] [CrossRef] [PubMed]
- Spalteholz, H.; Wenske, K.; Arnhold, J. Interaction of hypohalous acids and heme peroxidases with unsaturated phosphatidylcholines. Biofactors 2005, 24, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Greig, F.H.; Kennedy, S.; Spickett, C.M. Physiological effects of oxidized phospholipids and their cellular signaling mechanisms in inflammation. Free Radic. Biol. Med. 2012, 52, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Kanno, K.; Wu, M.K.; Scapa, E.F.; Roderick, S.L.; Cohen, D.E. Structure and function of phosphatidylcholine transfer protein (PC-TP)/StarD2. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2007, 1771, 654–662. [Google Scholar] [CrossRef]
- Kawai, Y.; Kiyokawa, H.; Kimura, Y.; Kato, Y.; Tsuchiya, K.; Terao, J. Hypochlorous acid-derived modification of phospholipids: Characterization of aminophospholipids as regulatory molecules for lipid peroxidation. Biochemistry 2006, 45, 14201–14211. [Google Scholar] [CrossRef]
- Villamena, F.A. Reactive Species Detection in Biology; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Nauseef, W.M. Myeloperoxidase in human neutrophil host defence. Cell. Microbiol. 2014, 16, 1146–1155. [Google Scholar] [CrossRef]
- Thomas, E.L. Myeloperoxidase, hydrogen peroxide, chloride antimicrobial system: Nitrogen-chlorine derivatives of bacterial components in bactericidal action against Escherichia coli. Infect. Immun. 1979, 23, 522–531. [Google Scholar] [CrossRef]
- Corbett, R.; Metcalf, W.; Soper, F. 395. Studies of N-halogeno-compounds. Part IV. The reaction between ammonia and chlorine in aqueous solution, and the hydrolysis constants of chloroamines. J. Chem. Soc. Resumed 1953, 1927–1929. [Google Scholar] [CrossRef]
- Chapin, R.M. Dichloro-amine. J. Am. Chem. Soc. 1929, 51, 2112–2117. [Google Scholar] [CrossRef]
- Abia, L.; Armesto, X.; Canle, M.; Garcia, M.; Santaballa, J. Oxidation of aliphatic amines by aqueous chlorine. Tetrahedron 1998, 54, 521–530. [Google Scholar] [CrossRef]
- Armesto, X.; García, M.; Santaballa, J. Aqueous chemistry of N-halo-compounds. Chem. Soc. Rev. 1998, 27, 453–460. [Google Scholar] [CrossRef]
- Fernández-Pérez, M.-I. Mecanismos de Oxidación y Fotooxidación de Compuestos de Relevancia Bioquímica y Medioambiental: 1. Halogenación de Aminoácidos Azufrados. 2. Fotodegradación de Biocidas Tipo S. Triazina. Ph.D. Thesis, University of Coruña, A Coruña, Spain, 2002. Available online: https://ruc.udc.es/dspace/handle/2183/11706 (accessed on 4 August 2022).
- Bochkov, V.N.; Oskolkova, O.V.; Birukov, K.G.; Levonen, A.-L.; Binder, C.J.; Stöckl, J. Generation and biological activities of oxidized phospholipids. Antioxid. Redox Signal. 2010, 12, 1009–1059. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L.; Pattison, D.I.; Rees, M.D. Mammalian heme peroxidases: From molecular mechanisms to health implications. Antioxid. Redox Signal. 2008, 10, 1199–1234. [Google Scholar] [CrossRef]
- Lloyd, M.M.; van Reyk, D.M.; Davies, M.J.; Hawkins, C.L. Hypothiocyanous acid is a more potent inducer of apoptosis and protein thiol depletion in murine macrophage cells than hypochlorous acid or hypobromous acid. Biochem. J. 2008, 414, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Pattison, D.; Davies, M. Reactions of myeloperoxidase-derived oxidants with biological substrates: Gaining chemical insight into human inflammatory diseases. Curr. Med. Chem. 2006, 13, 3271–3290. [Google Scholar] [CrossRef] [PubMed]
- Ulfig, A.; Leichert, L.I. The effects of neutrophil-generated hypochlorous acid and other hypohalous acids on host and pathogens. Cell. Mol. Life Sci. 2021, 78, 385–414. [Google Scholar] [CrossRef] [PubMed]
- Kettle, A.J. Detection of 3-chlorotyrosine in proteins exposed to neutrophil oxidants. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 300, pp. 111–120. [Google Scholar]
- Olszowski, S.; Olszowska, E.; Stelmaszyńska, T.; Krawczyk, A.; Marcinkiewicz, J.; Baczek, N. Oxidative modification of ovalbumin. Acta Biochim. Pol. 1996, 43, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Domigan, N.M.; Charlton, T.S.; Duncan, M.W.; Winterbourn, C.C.; Kettle, A.J. Chlorination of tyrosyl residues in peptides by myeloperoxidase and human neutrophils. J. Biol. Chem. 1995, 270, 16542–16548. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.L.; Hsu, F.F.; Mueller, D.M.; Crowley, J.R.; Heinecke, J.W. Human neutrophils employ chlorine gas as an oxidant during phagocytosis. J. Clin. Investig. 1996, 98, 1283–1289. [Google Scholar] [CrossRef]
- Davies, M.; Fu, S.; Wang, H.; Dean, R. Reactions of hypochlorous acid with tyrosine and peptidyl-tyrosyl residues give dichlorinated and aldehydic products in addition to 3-chlorotyrosine. J. Biol. Chem. 2000, 275, 10851–10858. [Google Scholar]
- Kang, J.; Joseph, I.; Neidigh, J.W. Hypochlorous acid damages histone proteins forming 3-chlorotyrosine and 3, 5-dichlorotyrosine. Chem. Res. Toxicol. 2008, 21, 1028–1038. [Google Scholar] [CrossRef]
- Hawkins, C.; Pattison, D.; Davies, M. Hypochlorite-induced oxidation of amino acids, peptides and proteins. Amino Acids 2003, 25, 259–274. [Google Scholar] [CrossRef]
- Kettle, A.J. Neutrophils convert tyrosyl residues in albumin to chlorotyrosine. FEBS Lett. 1996, 379, 103–106. [Google Scholar] [CrossRef]
- Hazen, S.L.; Hsu, F.F.; Heinecke, J.W. p-Hydroxyphenylacetaldehyde is the major product of L-tyrosine oxidation by activated human phagocytes: A chloride-dependent mechanism for the conversion of free amino acids into reactive aldehydes by myeloperoxidase. J. Biol. Chem. 1996, 271, 1861–1867. [Google Scholar] [CrossRef]
- Pereira, W.; Hoyano, Y.; Summons, R.; Bacon, V.; Duffield, A. Chlorination studies II. The reaction of aqueous hypochlorous acid with α-amino acids and dipeptides. Biochim. Biophys. Acta BBA Gen. Subj. 1973, 313, 170–180. [Google Scholar] [CrossRef]
- Zhang, C.; Reiter, C.; Eiserich, J.P.; Boersma, B.; Parks, D.A.; Beckman, J.S.; Barnes, S.; Kirk, M.; Baldus, S.; Darley-Usmar, V.M. L-arginine chlorination products inhibit endothelial nitric oxide production. J. Biol. Chem. 2001, 276, 27159–27165. [Google Scholar] [CrossRef]
- Pattison, D.I.; Davies, M.J. Absolute rate constants for the reaction of hypochlorous acid with protein side chains and peptide bonds. Chem. Res. Toxicol. 2001, 14, 1453–1464. [Google Scholar] [CrossRef]
- DrozdŹ, R.; Naskalski, J.W.; Sznajd, J. Oxidation of amino acids and peptides in reaction with myeloperoxidase, chloride and hydrogen peroxide. Biochim. Biophys. Acta BBA Protein Struct. Mol. Enzymol. 1988, 957, 47–52. [Google Scholar] [CrossRef]
- Gupta, V.; Carroll, K.S. Sulfenic acid chemistry, detection and cellular lifetime. Biochim. Biophys. Acta BBA Gen. Subj. 2014, 1840, 847–875. [Google Scholar] [CrossRef]
- Fu, X.; Mueller, D.M.; Heinecke, J.W. Generation of intramolecular and intermolecular sulfenamides, sulfinamides, and sulfonamides by hypochlorous acid: A potential pathway for oxidative cross-linking of low-density lipoprotein by myeloperoxidase. Biochemistry 2002, 41, 1293–1301. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L. Hypochlorite-induced oxidation of thiols: Formation of thiyl radicals and the role of sulfenyl chlorides as intermediates. Free Radic. Res. 2000, 33, 719–729. [Google Scholar] [CrossRef]
- Naskalski, J. Oxidative modification of protein structures under the action of myeloperoxidase and the hydrogen peroxide and chloride system. Ann. Biol. Clin. 1994, 52, 451–456. [Google Scholar]
- Hawkins, C.L. Hypochlorous acid-mediated modification of proteins and its consequences. Essays Biochem. 2020, 64, 75–86. [Google Scholar] [CrossRef]
- Tiedemann, F.; Gmelin, L. Einige neue Bestandtheile der Galle des Ochsen. Ann. Phys. 1827, 85, 326–337. [Google Scholar] [CrossRef]
- Merckx, C.; De Paepe, B. The Role of Taurine in Skeletal Muscle Functioning and Its Potential as a Supportive Treatment for Duchenne Muscular Dystrophy. Metabolites 2022, 12, 193. [Google Scholar] [CrossRef]
- Marcinkiewicz, J. Taurine bromamine (TauBr)-its role in immunity and new perspectives for clinical use. J. Biomed. Sci. 2010, 17, S3. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Walczewska, M. Neutrophils as sentinel cells of the immune system: A role of the MPO-halide-system in innate and adaptive immunity. Curr. Med. Chem. 2020, 27, 2840–2851. [Google Scholar] [CrossRef]
- Antelo, J.M.; Arce, F.; Calvo, P.; Crugeiras, J.; Ríos, A. General acid–base catalysis in the reversible disproportionation reaction of N-chlorotaurine. J. Chem. Soc. Perkin Trans. 2 2000, 2109–2114. [Google Scholar] [CrossRef]
- Gottardi, W.; Nagl, M. N-chlorotaurine, a natural antiseptic with outstanding tolerability. J. Antimicrob. Chemother. 2010, 65, 399–409. [Google Scholar] [CrossRef]
- Gottardi, W.; Hagleitner, M.; Nagl, M. N, N-Dichlorotaurine: Chemical and Bactericidal Properties. Arch. Pharm. Int. J. Pharm. Med. Chem. 2005, 338, 473–483. [Google Scholar] [CrossRef]
- Seol, S.-I.; Kim, H.J.; Choi, E.B.; Kang, I.S.; Lee, H.-K.; Lee, J.-K.; Kim, C. Taurine protects against postischemic brain injury via the antioxidant activity of taurine chloramine. Antioxidants 2021, 10, 372. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. The role of reactive N-bromo species and radical intermediates in hypobromous acid-induced protein oxidation. Free Radic. Biol. Med. 2005, 39, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Peskin, A.V.; Winterbourn, C.C. Kinetics of the reactions of hypochlorous acid and amino acid chloramines with thiols, methionine, and ascorbate. Free Radic. Biol. Med. 2001, 30, 572–579. [Google Scholar] [CrossRef]
- Harlan, J.; Levine, J.; Callahan, K.; Schwartz, B.; Harker, L. Glutathione redox cycle protects cultured endothelial cells against lysis by extracellularly generated hydrogen peroxide. J. Clin. Investig. 1984, 73, 706–713. [Google Scholar] [CrossRef]
- Vissers, M.C.; Winterbourn, C.C. Oxidation of intracellular glutathione after exposure of human red blood cells to hypochlorous acid. Biochem. J. 1995, 307, 57–62. [Google Scholar] [CrossRef]
- Folkes, L.K.; Candeias, L.P.; Wardman, P. Kinetics and mechanisms of hypochlorous acid reactions. Arch. Biochem. Biophys. 1995, 323, 120–126. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Brennan, S.O. Characterization of the oxidation products of the reaction between reduced glutathione and hypochlorous acid. Biochem. J. 1997, 326, 87–92. [Google Scholar] [CrossRef]
- Haenen, G.R.; Bast, A. Glutathione revisited: A better scavenger than previously thought. Front. Pharmacol. 2014, 5, 260. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-induced damage to proteins: Formation of nitrogen-centred radicals from lysine residues and their role in protein fragmentation. Biochem. J. 1998, 332, 617–625. [Google Scholar] [CrossRef]
- Thomas, E.L.; Grisham, M.B.; Jefferson, M.M. Preparation and characterization of chloramines. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1986; Volume 132, pp. 569–585. [Google Scholar]
- Zhao, H.; Zhou, Y.; Han, C.; Liu, Y.D.; Zhong, R. Degradation mechanisms and substituent effects of N-chloro-α-amino acids: A computational study. Environ. Sci. Technol. 2020, 54, 2635–2645. [Google Scholar] [CrossRef]
- Prütz, W.A. Hypochlorous acid interactions with thiols, nucleotides, DNA, and other biological substrates. Arch. Biochem. Biophys. 1996, 332, 110–120. [Google Scholar] [CrossRef]
- Prütz, W. Interactions of hypochlorous acid with pyrimidine nucleotides, and secondary reactions of chlorinated pyrimidines with GSH, NADH, and other substrates. Arch. Biochem. Biophys. 1998, 349, 183–191. [Google Scholar] [CrossRef]
- Albrich, J.M.; McCarthy, C.A.; Hurst, J.K. Biological reactivity of hypochlorous acid: Implications for microbicidal mechanisms of leukocyte myeloperoxidase. Proc. Natl. Acad. Sci. USA 1981, 78, 210–214. [Google Scholar] [CrossRef]
- Bernofsky, C. Nucleotide chloramines and neutrophil-mediated cytotoxicity. FASEB J. 1991, 5, 295–300. [Google Scholar] [CrossRef]
- Prütz, W. Consecutive halogen transfer between various functional groups induced by reaction of hypohalous acids: NADH oxidation by halogenated amide groups. Arch. Biochem. Biophys. 1999, 371, 107–114. [Google Scholar] [CrossRef]
- Prütz, W.A.; Kissner, R.; Koppenol, W.H.; Rüegger, H. On the irreversible destruction of reduced nicotinamide nucleotides by hypohalous acids. Arch. Biochem. Biophys. 2000, 380, 181–191. [Google Scholar] [CrossRef]
- Hayatsu, H.; Pan, S.; Ukita, T. Reaction of sodium hypochlorite with nucleic acids and their constituents. Chem. Pharm. Bull. 1971, 19, 2189–2192. [Google Scholar] [CrossRef]
- Patton, W.; Bacon, V.; Duffield, A.; Halpern, B.; Hoyano, Y.; Pereira, W.; Lederberg, J. Chlorination studies. I. The reaction of aqueous hypochlorous acid with cytosine. Biochem. Biophys. Res. Commun. 1972, 48, 880–884. [Google Scholar] [CrossRef]
- Gould, J.; Richards, J.; Miles, M. The formation of stable organic chloramines during the aqueous chlorination of cytosine and 5-methylcytosine. Water Res. 1984, 18, 991–999. [Google Scholar] [CrossRef]
- Gould, J.; Richards, J.; Miles, M. The kinetics and primary products of uracil chlorination. Water Res. 1984, 18, 205–212. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-induced damage to nucleosides: Formation of chloramines and nitrogen-centered radicals. Chem. Res. Toxicol. 2001, 14, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.E.; Thompson, R.J. Extracellular matrix proteoglycan degradation by human alveolar macrophages and neutrophils. J. Appl. Physiol. 1989, 66, 400–409. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.E. Mechanisms of extracellular matrix proteoglycan degradation by human neutrophils. Am. J. Respir. Cell Mol. Biol. 1990, 2, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Panasenko, O.; Gorudko, I.; Sokolov, A. Hypochlorous acid as a precursor of free radicals in living systems. Biochemistry 2013, 78, 1466–1489. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Degradation of hyaluronic acid, poly-and mono-saccharides, and model compounds by hypochlorite: Evidence for radical intermediates and fragmentation. Free Radic. Biol. Med. 1998, 24, 1396–1410. [Google Scholar] [CrossRef]
- Rees, M.D.; Hawkins, C.L.; Davies, M.J. Hypochlorite-mediated fragmentation of hyaluronan, chondroitin sulfates, and related N-acetyl glycosamines: Evidence for chloramide intermediates, free radical transfer reactions, and site-specific fragmentation. J. Am. Chem. Soc. 2003, 125, 13719–13733. [Google Scholar] [CrossRef]
- Woods, A.A.; Davies, M.J. Fragmentation of extracellular matrix by hypochlorous acid. Biochem. J. 2003, 376, 219–227. [Google Scholar] [CrossRef]
- Heinecke, J.W. Mechanisms of oxidative damage by myeloperoxidase in atherosclerosis and other inflammatory disorders. J. Lab. Clin. Med. 1999, 133, 321–325. [Google Scholar] [CrossRef]
- Baskol, G.; Demir, H.; Baskol, M.; Kilic, E.; Ates, F.; Karakukcu, C.; Ustdal, M. Investigation of protein oxidation and lipid peroxidation in patients with rheumatoid arthritis. Cell Biochem. Funct. Cell. Biochem. Modul. Act. Agents Dis. 2006, 24, 307–311. [Google Scholar] [CrossRef]
- Andreadis, A.A.; Hazen, S.L.; Comhair, S.A.; Erzurum, S.C. Oxidative and nitrosative events in asthma. Free Radic. Biol. Med. 2003, 35, 213–225. [Google Scholar] [CrossRef]
- Cantin, A.; Woods, D.E. Protection by antibiotics against myeloperoxidase-dependent cytotoxicity to lung epithelial cells in vitro. J. Clin. Investig. 1993, 91, 38–45. [Google Scholar] [CrossRef]
- Ndrepepa, G. Myeloperoxidase–A bridge linking inflammation and oxidative stress with cardiovascular disease. Clin. Chim. Acta 2019, 493, 36–51. [Google Scholar] [CrossRef]
- Kuzuya, T.; Fuji, H.; Hoshida, S.; Nishida, M.; Goshima, K.; Hori, M.; Kamada, T.; Tada, M. Polymorphonuclear leukocytes-induced injury in hypoxic cardiac myocytes. Free Radic. Biol. Med. 1994, 17, 501–510. [Google Scholar] [CrossRef]
- Leone, A.; Palmer, R.; Knowles, R.; Francis, P.; Ashton, D.; Moncada, S. Constitutive and inducible nitric oxide synthases incorporate molecular oxygen into both nitric oxide and citrulline. J. Biol. Chem. 1991, 266, 23790–23795. [Google Scholar] [CrossRef]
- Cooper, C.E.; Odell, E. Interaction of human myeloperoxidase with nitrite. FEBS Lett. 1992, 314, 58–60. [Google Scholar] [CrossRef]
- Klebanoff, S.J. Reactive nitrogen intermediates and antimicrobial activity: Role of nitrite. Free Radic. Biol. Med. 1993, 14, 351–360. [Google Scholar] [CrossRef]
- Van Der Vliet, A.; Eiserich, J.P.; Halliwell, B.; Cross, C.E. Formation of reactive nitrogen species during peroxidase-catalyzed oxidation of nitrite: A potential additional mechanism of nitric oxide-dependent toxicity. J. Biol. Chem. 1997, 272, 7617–7625. [Google Scholar] [CrossRef]
- O’Brien, P.J. Radical formation during the peroxidase catalyzed metabolism of carcinogens and xenobiotics: The reactivity of these radicals with GSH, DNA, and unsaturated lipid. Free Radic. Biol. Med. 1988, 4, 169–183. [Google Scholar] [CrossRef]
- Eastmond, D.; Smith, M.; Ruzo, L.; Ross, D. Metabolic activation of phenol by human myeloperoxidase and horseradish peroxidase. Mol. Pharmacol. 1986, 30, 674–679. [Google Scholar]
- Frimat, B.; Gressier, B.; Odou, P.; Brunet, C.; Dine, T.; Luycky, M.; Cazin, M.; Cazin, J. Metabolism of clozapine by human neutrophils: Evidence for a specific oxidation of clozapine by the myeloperoxidase system with inhibition of enzymatic chlorination cycle. Fundam. Clin. Pharmacol. 1997, 11, 267–274. [Google Scholar] [CrossRef]
- Gutierrez-Correa, J.; Stoppani, A. Inactivation of heart dihydrolipoamide dehydrogenase by copper Fenton systems. Effect of thiol compounds and metal chelators. Free Radic. Res. 1995, 22, 239–250. [Google Scholar] [CrossRef]
- Correa, J.G.; Stoppani, A. Catecholamines enhance dihydrolipoamide dehydrogenase inactivation by the copper Fenton system. Enzyme protection by copper chelators. Free Radic. Res. 1996, 24, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.G.; Stoppani, A. Inactivation of lipoamide dehydrogenase by cobalt (II) and iron (II) Fenton systems: Effect of metal chelators, thiol compounds and adenine nucleotides. Free Radic. Res. Commun. 1993, 19, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez Correa, J.; Biscardi, A.M.; Stoppani, A.O. Inactivación de la lipoamida deshidrogenasa de miocardio por catecolaminas: Protección por captopril y otros tioles. Med. B. Aires 1995, 55, 397–407. [Google Scholar]
- Anatoliotakis, N.; Deftereos, S.; Bouras, G.; Giannopoulos, G.; Tsounis, D.; Angelidis, C.; Kaoukis, A.; Stefanadis, C. Myeloperoxidase: Expressing inflammation and oxidative stress in cardiovascular disease. Curr. Top. Med. Chem. 2013, 13, 115–138. [Google Scholar] [CrossRef] [PubMed]
- Eiserich, J.P.; Baldus, S.; Brennan, M.-L.; Ma, W.; Zhang, C.; Tousson, A.; Castro, L.; Lusis, A.J.; Nauseef, W.M.; White, C.R. Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science 2002, 296, 2391–2394. [Google Scholar] [CrossRef]
- Mollenhauer, M.; Friedrichs, K.; Lange, M.; Gesenberg, J.; Remane, L.; Kerkenpaß, C.; Krause, J.; Schneider, J.; Ravekes, T.; Maass, M. Myeloperoxidase mediates postischemic arrhythmogenic ventricular remodeling. Circ. Res. 2017, 121, 56–70. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef]
- Carr, A.C.; McCall, M.R.; Frei, B. Oxidation of LDL by myeloperoxidase and reactive nitrogen species: Reaction pathways and antioxidant protection. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1716–1723. [Google Scholar] [CrossRef]
- Roth Flach, R.J.; Su, C.; Bollinger, E.; Cortes, C.; Robertson, A.W.; Opsahl, A.C.; Coskran, T.M.; Maresca, K.P.; Keliher, E.J.; Yates, P.D. Myeloperoxidase inhibition in mice alters atherosclerotic lesion composition. PLoS ONE 2019, 14, e0214150. [Google Scholar] [CrossRef]
- Wang, Z.; Nicholls, S.J.; Rodriguez, E.R.; Kummu, O.; Hörkkö, S.; Barnard, J.; Reynolds, W.F.; Topol, E.J.; DiDonato, J.A.; Hazen, S.L. Protein carbamylation links inflammation, smoking, uremia and atherogenesis. Nat. Med. 2007, 13, 1176–1184. [Google Scholar] [CrossRef]
- Pham, P.; Pham, P.; Pham, P. Patients with diabetes mellitus type 2 and hypomagnesemia may have enhanced glomerular filtration via hypocalcemia. Clin. Nephrol. 2012, 78, 442–448. [Google Scholar] [CrossRef]
- Chami, B.; Martin, N.J.; Dennis, J.M.; Witting, P.K. Myeloperoxidase in the inflamed colon: A novel target for treating inflammatory bowel disease. Arch. Biochem. Biophys. 2018, 645, 61–71. [Google Scholar] [CrossRef]
- Garrity-Park, M.; Loftus, E.V., Jr.; Sandborn, W.J.; Smyrk, T.C. Myeloperoxidase immunohistochemistry as a measure of disease activity in ulcerative colitis: Association with ulcerative colitis-colorectal cancer, tumor necrosis factor polymorphism and RUNX3 methylation. Inflamm. Bowel Dis. 2012, 18, 275–283. [Google Scholar] [CrossRef]
- Chami, B.; Ahmad, G.; Schroder, A.; San Gabriel, P.; Witting, P. The role of myeloperoxidase and neutrophil extracellular traps in the pathogenesis of inflammatory bowel disease. Gastroenterology 2021, 160, S5–S6. [Google Scholar] [CrossRef]
- Wang, W.; Jian, Z.; Guo, J.; Ning, X. Increased levels of serum myeloperoxidase in patients with active rheumatoid arthritis. Life Sci. 2014, 117, 19–23. [Google Scholar] [CrossRef]
- Fernandes, R.M.S.N.; Silva, N.P.d.; Sato, E.I. Increased myeloperoxidase plasma levels in rheumatoid arthritis. Rheumatol. Int. 2012, 32, 1605–1609. [Google Scholar] [CrossRef]
- Telles, R.W.; Ferreira, G.A.; Silva, N.P.d.; Sato, E.I. Increased plasma myeloperoxidase levels in systemic lupus erythematosus. Rheumatol. Int. 2010, 30, 779–784. [Google Scholar] [CrossRef]
- Olson, S.; Lee, J.; Poirier, M.; Little, D.; Prince, L.; Baker, T.; Edison, J.; Abbott, K. Anti-myeloperoxidase antibodies associate with future proliferative lupus nephritis. Autoimmune Dis. 2017, 2017, 1872846. [Google Scholar] [CrossRef]
- Carrillo-Vázquez, D.A.; Jardón-Valadez, E.; Torres-Ruiz, J.; Juárez-Vega, G.; Maravillas-Montero, J.L.; Meza-Sánchez, D.E.; Domínguez-López, M.L.; Varela, J.C.A.; Gómez-Martín, D. Conformational changes in myeloperoxidase induced by ubiquitin and NETs containing free ISG15 from systemic lupus erythematosus patients promote a pro-inflammatory cytokine response in CD4+ T cells. J. Transl. Med. 2020, 18, 429. [Google Scholar] [CrossRef]
- Green, P.S.; Mendez, A.J.; Jacob, J.S.; Crowley, J.R.; Growdon, W.; Hyman, B.T.; Heinecke, J.W. Neuronal expression of myeloperoxidase is increased in Alzheimer’s disease. J. Neurochem. 2004, 90, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Tzikas, S.; Schlak, D.; Sopova, K.; Gatsiou, A.; Stakos, D.; Stamatelopoulos, K.; Stellos, K.; Laske, C. Increased myeloperoxidase plasma levels in patients with Alzheimer’s disease. J. Alzheimer’s Dis. 2014, 39, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Volkman, R.; Ben-Zur, T.; Kahana, A.; Garty, B.Z.; Offen, D. Myeloperoxidase deficiency inhibits cognitive decline in the 5XFAD mouse model of Alzheimer’s disease. Front. Neurosci. 2019, 13, 990. [Google Scholar] [CrossRef] [PubMed]
- Pulli, B.; Bure, L.; Wojtkiewicz, G.R.; Iwamoto, Y.; Ali, M.; Li, D.; Schob, S.; Hsieh, K.L.-C.; Jacobs, A.H.; Chen, J.W. Multiple sclerosis: Myeloperoxidase immunoradiology improves detection of acute and chronic disease in experimental model. Radiology 2015, 275, 480. [Google Scholar] [CrossRef]
- Gray, E.; Thomas, T.L.; Betmouni, S.; Scolding, N.; Love, S. Elevated myeloperoxidase activity in white matter in multiple sclerosis. Neurosci. Lett. 2008, 444, 195–198. [Google Scholar] [CrossRef]
- Nagra, R.M.; Becher, B.; Tourtellotte, W.W.; Antel, J.P.; Gold, D.; Paladino, T.; Smith, R.A.; Nelson, J.R.; Reynolds, W.F. Immunohistochemical and genetic evidence of myeloperoxidase involvement in multiple sclerosis. J. Neuroimmunol. 1997, 78, 97–107. [Google Scholar] [CrossRef]
- Pravalika, K.; Sarmah, D.; Kaur, H.; Wanve, M.; Saraf, J.; Kalia, K.; Borah, A.; Yavagal, D.R.; Dave, K.R.; Bhattacharya, P. Myeloperoxidase and neurological disorder: A crosstalk. ACS Chem. Neurosci. 2018, 9, 421–430. [Google Scholar] [CrossRef]
- Ray, R.; Katyal, A. Myeloperoxidase: Bridging the gap in neurodegeneration. Neurosci. Biobehav. Rev. 2016, 68, 611–620. [Google Scholar] [CrossRef]
- Gellhaar, S.; Sunnemark, D.; Eriksson, H.; Olson, L.; Galter, D. Myeloperoxidase-immunoreactive cells are significantly increased in brain areas affected by neurodegeneration in Parkinson’s and Alzheimer’s disease. Cell Tissue Res. 2017, 369, 445–454. [Google Scholar] [CrossRef]
- Choi, D.-K.; Pennathur, S.; Perier, C.; Tieu, K.; Teismann, P.; Wu, D.-C.; Jackson-Lewis, V.; Vila, M.; Vonsattel, J.-P.; Heinecke, J.W. Ablation of the inflammatory enzyme myeloperoxidase mitigates features of Parkinson’s disease in mice. J. Neurosci. 2005, 25, 6594–6600. [Google Scholar] [CrossRef]
- Jucaite, A.; Svenningsson, P.; Rinne, J.O.; Cselenyi, Z.; Varnäs, K.; Johnström, P.; Amini, N.; Kirjavainen, A.; Helin, S.; Minkwitz, M. Effect of the myeloperoxidase inhibitor AZD3241 on microglia: A PET study in Parkinson’s disease. Brain 2015, 138, 2687–2700. [Google Scholar] [CrossRef] [Green Version]
- Cojocaru, I.M.; Cojocaru, M.; Iliescu, I.; Botnaru, L.; Gurban, C.V.; Sfrijan, F.; Tanasescu, R. Plasma myeloperoxidase levels in patients with acute ischemic stroke. Rom. J. Intern. Med. 2010, 48, 101–104. [Google Scholar]
- Tay, A.; Tamam, Y.; Yokus, B.; Ustundag, M.; Orak, M. Serum myeloperoxidase levels in predicting the severity of stroke and mortality in acute ischemic stroke patients. Eur. Rev. Med. Pharm. Sci. 2015, 19, 1983–1988. [Google Scholar]
- Yu, G.; Liang, Y.; Huang, Z.; Jones, D.W.; Pritchard, K.A.; Zhang, H. Inhibition of myeloperoxidase oxidant production by N-acetyl lysyltyrosylcysteine amide reduces brain damage in a murine model of stroke. J. Neuroinflamm. 2016, 13, 119. [Google Scholar] [CrossRef]
- Li, S.-B.; Yang, F.; Jing, L.; Ma, J.; Jia, Y.-D.; Dong, S.-Y.; Zheng, W.-F.; Zhao, L.-S. Myeloperoxidase and risk of recurrence of atrial fibrillation after catheter ablation. J. Investig. Med. 2013, 61, 722–727. [Google Scholar] [CrossRef]
- Rudolph, V.; Andrié, R.P.; Rudolph, T.K.; Friedrichs, K.; Klinke, A.; Hirsch-Hoffmann, B.; Schwoerer, A.P.; Lau, D.; Fu, X.; Klingel, K. Myeloperoxidase acts as a profibrotic mediator of atrial fibrillation. Nat. Med. 2010, 16, 470–474. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Hazen, S.L. Myeloperoxidase, modified lipoproteins, and atherogenesis. J. Lipid Res. 2009, 50, S346–S351. [Google Scholar] [CrossRef]
- Tsimikas, S. Oxidative biomarkers in the diagnosis and prognosis of cardiovascular disease. Am. J. Cardiol. 2006, 98, S9–S17. [Google Scholar] [CrossRef]
- Zhang, R.; Brennan, M.-L.; Fu, X.; Aviles, R.J.; Pearce, G.L.; Penn, M.S.; Topol, E.J.; Sprecher, D.L.; Hazen, S.L. Association between myeloperoxidase levels and risk of coronary artery disease. JAMA 2001, 286, 2136–2142. [Google Scholar] [CrossRef]
- Klinke, A.; Berghausen, E.; Friedrichs, K.; Molz, S.; Lau, D.; Remane, L.; Berlin, M.; Kaltwasser, C.; Adam, M.; Mehrkens, D. Myeloperoxidase aggravates pulmonary arterial hypertension by activation of vascular Rho-kinase. JCI Insight 2018, 3, e97530. [Google Scholar] [CrossRef]
- Van der Zwan, L.P.; Scheffer, P.G.; Dekker, J.M.; Stehouwer, C.D.; Heine, R.J.; Teerlink, T. Hyperglycemia and oxidative stress strengthen the association between myeloperoxidase and blood pressure. Hypertension 2010, 55, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Penha, L.; Caldeira-Dias, M.; Tanus-Santos, J.E.; de Carvalho Cavalli, R.; Sandrim, V.C. Myeloperoxidase in hypertensive disorders of pregnancy and its relation with nitric oxide. Hypertension 2017, 69, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Mocatta, T.J.; Pilbrow, A.P.; Cameron, V.A.; Senthilmohan, R.; Frampton, C.M.; Richards, A.M.; Winterbourn, C.C. Plasma concentrations of myeloperoxidase predict mortality after myocardial infarction. J. Am. Coll. Cardiol. 2007, 49, 1993–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, M.D.; Pattison, D.I.; Davies, M.J. Oxidation of heparan sulphate by hypochlorite: Role of N-chloro derivatives and dichloramine-dependent fragmentation. Biochem. J. 2005, 391, 125–134. [Google Scholar] [CrossRef]
- Omran, M.M.; Zahran, F.M.; Kadry, M.; Belal, A.A.; Emran, T.M. Role of myeloperoxidase in early diagnosis of acute myocardial infarction in patients admitted with chest pain. J. Immunoass. Immunochem. 2018, 39, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Abdo, A.I.; Rayner, B.S.; van Reyk, D.M.; Hawkins, C.L. Low-density lipoprotein modified by myeloperoxidase oxidants induces endothelial dysfunction. Redox Biol. 2017, 13, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Manchanda, K.; Kolarova, H.; Kerkenpaß, C.; Mollenhauer, M.; Vitecek, J.; Rudolph, V.; Kubala, L.; Baldus, S.; Adam, M.; Klinke, A. MPO (myeloperoxidase) reduces endothelial glycocalyx thickness dependent on its cationic charge. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Vita, J.A.; Brennan, M.-L.; Gokce, N.; Mann, S.A.; Goormastic, M.; Shishehbor, M.H.; Penn, M.S.; Keaney, J.F., Jr.; Hazen, S.L. Serum myeloperoxidase levels independently predict endothelial dysfunction in humans. Circulation 2004, 110, 1134–1139. [Google Scholar] [CrossRef]
- Caramori, G.; Papi, A. Oxidants and asthma. Thorax 2004, 59, 170–173. [Google Scholar] [CrossRef]
- Ekmekci, O.; Donma, O.; Sardoğan, E.; Yildirim, N.; Uysal, O.; Demirel, H.; Demir, T. Iron, nitric oxide, and myeloperoxidase in asthmatic patients. Biochemistry 2004, 69, 462–467. [Google Scholar] [CrossRef]
- Jatakanon, A.; Uasuf, C.; Maziak, W.; Lim, S.; Chung, K.F.; Barnes, P.J. Neutrophilic inflammation in severe persistent asthma. Am. J. Respir. Crit. Care Med. 1999, 160, 1532–1539. [Google Scholar] [CrossRef]
- O’Donnell, C.; Newbold, P.; White, P.; Thong, B.; Stone, H.; Stockley, R.A. 3-Chlorotyrosine in sputum of COPD patients: Relationship with airway inflammation. COPD J. Chronic Obstr. Pulm. Dis. 2010, 7, 411–417. [Google Scholar] [CrossRef]
- Zhu, A.; Ge, D.; Zhang, J.; Teng, Y.; Yuan, C.; Huang, M.; Adcock, I.M.; Barnes, P.J.; Yao, X. Sputum myeloperoxidase in chronic obstructive pulmonary disease. Eur. J. Med. Res. 2014, 19, 12. [Google Scholar] [CrossRef] [Green Version]
- Andelid, K.; Glader, P.; Jirholt, P.; Gjertsson, I.; Jansson, A.E.; Lindén, A. Systemic myeloperoxidase in COPD. Respir. Med. 2013, 107, S4. [Google Scholar] [CrossRef]
- Kettle, A.J.; Chan, T.; Osberg, I.; Senthilmohan, R.; Chapman, A.L.; Mocatta, T.J.; Wagener, J.S. Myeloperoxidase and protein oxidation in the airways of young children with cystic fibrosis. Am. J. Respir. Crit. Care Med. 2004, 170, 1317–1323. [Google Scholar] [CrossRef]
- Van der Vliet, A.; Nguyen, M.N.; Shigenaga, M.K.; Eiserich, J.P.; Marelich, G.P.; Cross, C.E. Myeloperoxidase and protein oxidation in cystic fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L537–L546. [Google Scholar] [CrossRef]
- Vasu, V.T.; De Cruz, S.J.; Houghton, J.S.; Hayakawa, K.A.; Morrissey, B.M.; Cross, C.E.; Eiserich, J.P. Evaluation of thiol-based antioxidant therapeutics in cystic fibrosis sputum: Focus on myeloperoxidase. Free Radic. Res. 2011, 45, 165–176. [Google Scholar] [CrossRef]
- Casciaro, M.; Di Salvo, E.; Pace, E.; Ventura-Spagnolo, E.; Navarra, M.; Gangemi, S. Chlorinative stress in age-related diseases: A literature review. Immun. Ageing 2017, 14, 21. [Google Scholar] [CrossRef]
- Alikhan, M.A.; Jaw, J.; Shochet, L.R.; Robson, K.J.; Ooi, J.D.; Brouwer, E.; Heeringa, P.; Holdsworth, S.R.; Kitching, A.R. Ageing enhances cellular immunity to myeloperoxidase and experimental anti-myeloperoxidase glomerulonephritis. Rheumatology 2022, 61, 2132–2143. [Google Scholar] [CrossRef]
- Lu, H.; Xu, S.; Liang, X.; Dai, Y.; Huang, Z.; Ren, Y.; Lin, J.; Liu, X. Advanced glycated end products alter neutrophil effect on regulation of CD4+ T cell differentiation through induction of myeloperoxidase and neutrophil elastase activities. Inflammation 2019, 42, 559–571. [Google Scholar] [CrossRef]
- Fedeles, B.I.; Freudenthal, B.D.; Yau, E.; Singh, V.; Chang, S.-c.; Li, D.; Delaney, J.C.; Wilson, S.H.; Essigmann, J.M. Intrinsic mutagenic properties of 5-chlorocytosine: A mechanistic connection between chronic inflammation and cancer. Proc. Natl. Acad. Sci. USA 2015, 112, E4571–E4580. [Google Scholar] [CrossRef]
- Rymaszewski, A.L.; Tate, E.; Yimbesalu, J.P.; Gelman, A.E.; Jarzembowski, J.A.; Zhang, H.; Pritchard, K.A., Jr.; Vikis, H.G. The role of neutrophil myeloperoxidase in models of lung tumor development. Cancers 2014, 6, 1111–1127. [Google Scholar] [CrossRef]
- Weitzman, S.A.; Gordon, L.I. Inflammation and cancer: Role of phagocyte-generated oxidants in carcinogenesis. Blood 1990, 76, 655–663. [Google Scholar] [CrossRef]
- Kisic, B.; Miric, D.; Dragojevic, I.; Rasic, J.; Popovic, L. Role of myeloperoxidase in patients with chronic kidney disease. Oxidative Med. Cell. Longev. 2016, 2016, 1069743. [Google Scholar] [CrossRef]
- Lehners, A.; Lange, S.; Niemann, G.; Rosendahl, A.; Meyer-Schwesinger, C.; Oh, J.; Stahl, R.; Ehmke, H.; Benndorf, R.; Klinke, A. Myeloperoxidase deficiency ameliorates progression of chronic kidney disease in mice. Am. J. Physiol. Ren. Physiol. 2014, 307, F407–F417. [Google Scholar] [CrossRef]
- Madhusudhana Rao, A.; Anand, U.; Anand, C. Myeloperoxidase in chronic kidney disease. Indian J. Clin. Biochem. 2011, 26, 28–31. [Google Scholar] [CrossRef]
- Chen, S.; Chen, H.; Du, Q.; Shen, J. Targeting myeloperoxidase (MPO) mediated oxidative stress and inflammation for reducing brain ischemia injury: Potential application of natural compounds. Front. Physiol. 2020, 11, 433. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, Z.; Zhang, W.; Zhou, J.; Wu, Y.; Zhang, M.; Zhu, H.; Zou, M.-H. Myeloperoxidase deletion prevents high-fat diet–induced obesity and insulin resistance. Diabetes 2014, 63, 4172–4185. [Google Scholar] [CrossRef]
- Zaki, M.; Basha, W.; Reyad, H.; Mohamed, R.; Hassan, N.; Kholousi, S. Association between myeloperoxidase levels and risk of insulin resistance in Egyptian obese women. Open Access Maced. J. Med. Sci. 2018, 6, 629. [Google Scholar] [CrossRef]
- Qaddoumi, M.G.; Alanbaei, M.; Hammad, M.M.; Al Khairi, I.; Cherian, P.; Channanath, A.; Thanaraj, T.A.; Al-Mulla, F.; Abu-Farha, M.; Abubaker, J. Investigating the role of myeloperoxidase and angiopoietin-like protein 6 in obesity and diabetes. Sci. Rep. 2020, 10, 6170. [Google Scholar] [CrossRef]
- García, A.G.; Rodríguez, M.R.; Alonso, C.G.; Ochoa, D.Y.R.; Aguilar, C.A. Myeloperoxidase is associated with insulin resistance and inflammation in overweight subjects with first-degree relatives with type 2 diabetes mellitus. Diabetes Metab. J. 2015, 39, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Heinecke, J.W.; Goldberg, I.J. Myeloperoxidase: A therapeutic target for preventing insulin resistance and the metabolic sequelae of obesity? Diabetes 2014, 63, 4001–4003. [Google Scholar] [CrossRef] [PubMed]
- El Kebir, D.; József, L.; Pan, W.; Filep, J.n.G. Myeloperoxidase delays neutrophil apoptosis through CD11b/CD18 integrins and prolongs inflammation. Circ. Res. 2008, 103, 352–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorward, D.A.; Lucas, C.D.; Chapman, G.B.; Haslett, C.; Dhaliwal, K.; Rossi, A.G. The role of formylated peptides and formyl peptide receptor 1 in governing neutrophil function during acute inflammation. Am. J. Pathol. 2015, 185, 1172–1184. [Google Scholar] [CrossRef]
- Futosi, K.; Fodor, S.; Mócsai, A. Reprint of Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int. Immunopharmacol. 2013, 17, 1185–1197. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Pattison, D.I.; Stanley, N.R.; Davies, M.J. Tryptophan residues are targets in hypothiocyanous acid-mediated protein oxidation. Biochem. J. 2008, 416, 441–452. [Google Scholar] [CrossRef]
- Britigan, B.E.; Ratcliffe, H.R.; Buettner, G.R.; Rosen, G.M. Binding of myeloperoxidase to bacteria: Effect on hydroxyl radical formation and susceptibility to oxidant-mediated killing. Biochim. Biophys. Acta BBA Gen. Subj. 1996, 1290, 231–240. [Google Scholar] [CrossRef]
- Haegens, A.; Vernooy, J.H.; Heeringa, P.; Mossman, B.T.; Wouters, E.F. Myeloperoxidase modulates lung epithelial responses to pro-inflammatory agents. Eur. Respir. J. 2008, 31, 252–260. [Google Scholar] [CrossRef]
- Klinke, A.; Nussbaum, C.; Kubala, L.; Friedrichs, K.; Rudolph, T.K.; Rudolph, V.; Paust, H.-J.; Schröder, C.; Benten, D.; Lau, D. Myeloperoxidase attracts neutrophils by physical forces. Blood J. Am. Soc. Hematol. 2011, 117, 1350–1358. [Google Scholar] [CrossRef]
- Lazarevic-Pasti, T.; Leskovac, A.; Vasic, V. Myeloperoxidase inhibitors as potential drugs. Curr. Drug Metab. 2015, 16, 168–190. [Google Scholar] [CrossRef]
- Galijasevic, S. The development of myeloperoxidase inhibitors. Bioorg. Med. Chem. Lett. 2019, 29, 1–7. [Google Scholar] [CrossRef]
- Piek, A.; Koonen, D.P.; Schouten, E.-M.; Lindtstedt, E.L.; Michaëlsson, E.; de Boer, R.A.; Silljé, H.H. Pharmacological myeloperoxidase (MPO) inhibition in an obese/hypertensive mouse model attenuates obesity and liver damage, but not cardiac remodeling. Sci. Rep. 2019, 9, 18765. [Google Scholar] [CrossRef]
- Huang, J.; Smith, F.; Panizzi, J.R.; Goodwin, D.C.; Panizzi, P. Inactivation of myeloperoxidase by benzoic acid hydrazide. Arch. Biochem. Biophys. 2015, 570, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Furtmüller, P.G.; Arnhold, J.; Jantschko, W.; Pichler, H.; Obinger, C. Redox properties of the couples compound I/compound II and compound II/native enzyme of human myeloperoxidase. Biochem. Biophys. Res. Commun. 2003, 301, 551–557. [Google Scholar] [CrossRef]
- Bensalem, S.; Soubhye, J.; Aldib, I.; Bournine, L.; Nguyen, A.T.; Vanhaeverbeek, M.; Rousseau, A.; Boudjeltia, K.Z.; Sarakbi, A.; Kauffmann, J.M. Inhibition of myeloperoxidase activity by the alkaloids of Peganum harmala L.(Zygophyllaceae). J. Ethnopharmacol. 2014, 154, 361–369. [Google Scholar] [CrossRef]
- Segelmark, M.; Persson, B.; Hellmark, T.; Wieslander, J. Binding and inhibition of myeloperoxidase (MPO): A major function of ceruloplasmin? Clin. Exp. Immunol. 1997, 108, 167–174. [Google Scholar] [CrossRef]
- Kohnen, S.; Franck, T.; Van Antwerpen, P.; Zouaoui Boudjeltia, K.; Mouithys-Mickalad, A.; Deby, C.; Moguilevsky, N.; Deby-Dupont, G.; Lamy, M.; Serteyn, D. Resveratrol inhibits the activity of equine neutrophil myeloperoxidase by a direct interaction with the enzyme. J. Agric. Food Chem. 2007, 55, 8080–8087. [Google Scholar] [CrossRef]
- Zeraik, M.L.; Ximenes, V.F.; Regasini, L.O.; Dutra, L.; Silva, D.H.S.; Fonseca, L.; Coelho, D.; Machado, S.; Bolzani, V.d.S. 4′-Aminochalcones as novel inhibitors of the chlorinating activity of myeloperoxidase. Curr. Med. Chem. 2012, 19, 5405–5413. [Google Scholar] [CrossRef]
- Shiba, Y.; Kinoshita, T.; Chuman, H.; Taketani, Y.; Takeda, E.; Kato, Y.; Naito, M.; Kawabata, K.; Ishisaka, A.; Terao, J. Flavonoids as substrates and inhibitors of myeloperoxidase: Molecular actions of aglycone and metabolites. Chem. Res. Toxicol. 2008, 21, 1600–1609. [Google Scholar] [CrossRef]




















































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | K2 M−1 s−1 | Residue | K2 M−1 s−1 |
|---|---|---|---|
| Met | 3.8 × 107 | Lys | 5.0 × 103 |
| Cys | 3.0 × 107 | Tyr | 44 |
| Cistine | 1.6 × 105 | Arg | 26 |
| His | 1.0 × 105 | Backbone amides | <10 |
| α-amino | 1.0 × 105 | Asn | 0.03 |
| Trp | 1.1 × 104 | Gln | 0.03 |
| Disease Classification | Disease and References |
|---|---|
| Autoimmune Disease | Inflammatory bowel disease/colitis [150,151,152] Rheumatoid arthritis [153,154] Systemic lupus erythematosus [155,156,157] |
| Neuronal Pathology | Alzheimer’s disease [158,159,160] Multiple sclerosis [161,162,163] Neurodegenerative disease [164,165,166] Parkinson’s disease [167,168] Stroke [169,170,171] |
| Cardiovascular Pathology | Atrial fibrillation [172,173] Cardiovascular disease/atherosclerosis [174,175,176] Hypertension [177,178,179] Myocardial infarction [180,181,182] Vascular dysfunction [183,184,185] Asthma [186,187,188] |
| Pulmonary Pathology | Chronic obstructive pulmonary disease [189,190,191] Cystic fibrosis [192,193,194] |
| Miscellaneous | Ageing [195,196,197] Cancer [198,199,200] Chronic kidney disease [201,202,203] Inflammation [16,129,204] Lipoprotein modification [151,182,183] Metabolic syndrome/obesity [205,206,207] Type 2 diabetes [207,208,209] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Hypochlorous Acid Chemistry in Mammalian Cells—Influence on Infection and Role in Various Pathologies. Int. J. Mol. Sci. 2022, 23, 10735. https://doi.org/10.3390/ijms231810735
Andrés CMC, Pérez de la Lastra JM, Juan CA, Plou FJ, Pérez-Lebeña E. Hypochlorous Acid Chemistry in Mammalian Cells—Influence on Infection and Role in Various Pathologies. International Journal of Molecular Sciences. 2022; 23(18):10735. https://doi.org/10.3390/ijms231810735
Chicago/Turabian StyleAndrés, Celia María Curieses, José Manuel Pérez de la Lastra, Celia Andrés Juan, Francisco J. Plou, and Eduardo Pérez-Lebeña. 2022. "Hypochlorous Acid Chemistry in Mammalian Cells—Influence on Infection and Role in Various Pathologies" International Journal of Molecular Sciences 23, no. 18: 10735. https://doi.org/10.3390/ijms231810735
APA StyleAndrés, C. M. C., Pérez de la Lastra, J. M., Juan, C. A., Plou, F. J., & Pérez-Lebeña, E. (2022). Hypochlorous Acid Chemistry in Mammalian Cells—Influence on Infection and Role in Various Pathologies. International Journal of Molecular Sciences, 23(18), 10735. https://doi.org/10.3390/ijms231810735