
Serum microRNAs in Systemic Sclerosis, Associations with Digital Vasculopathy and Lung Involvement

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. SSc Patients Show a Defined Demographic and Clinical Profile
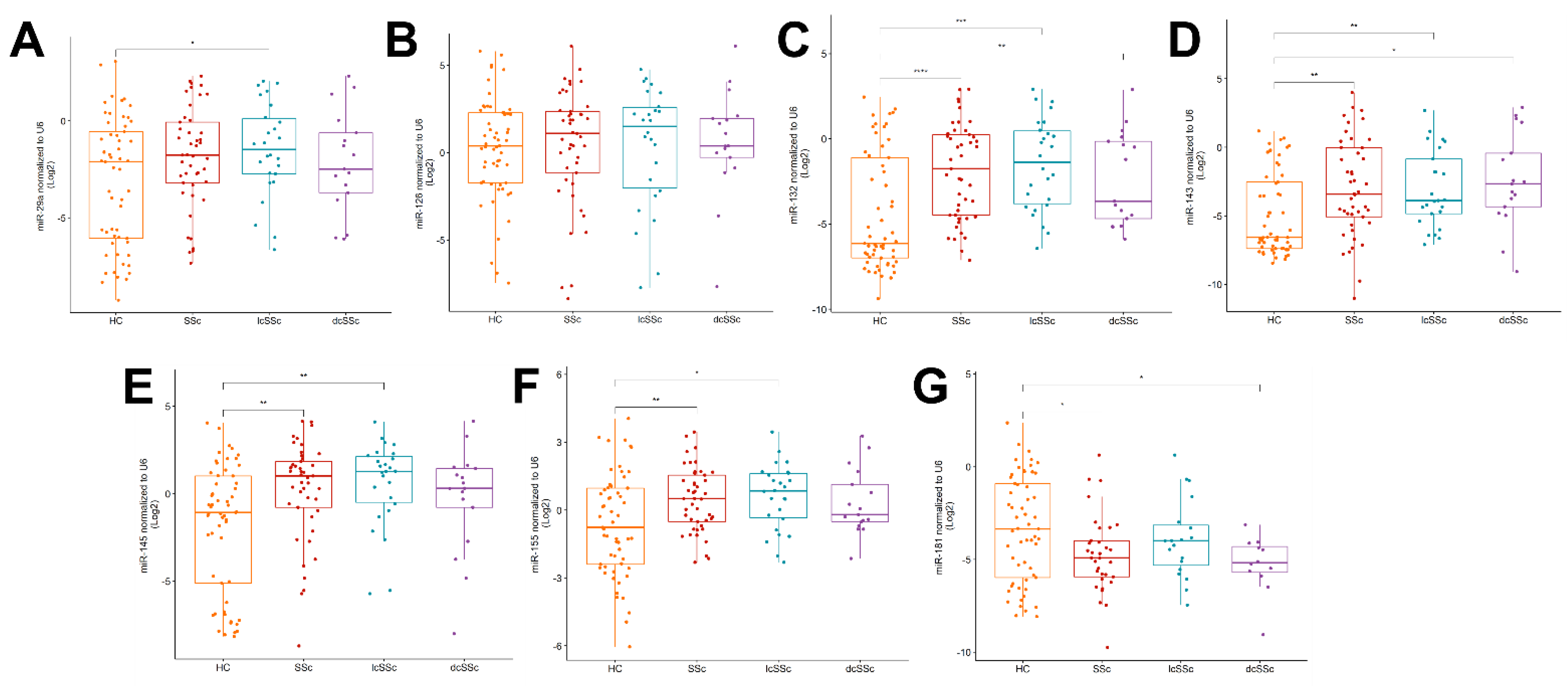
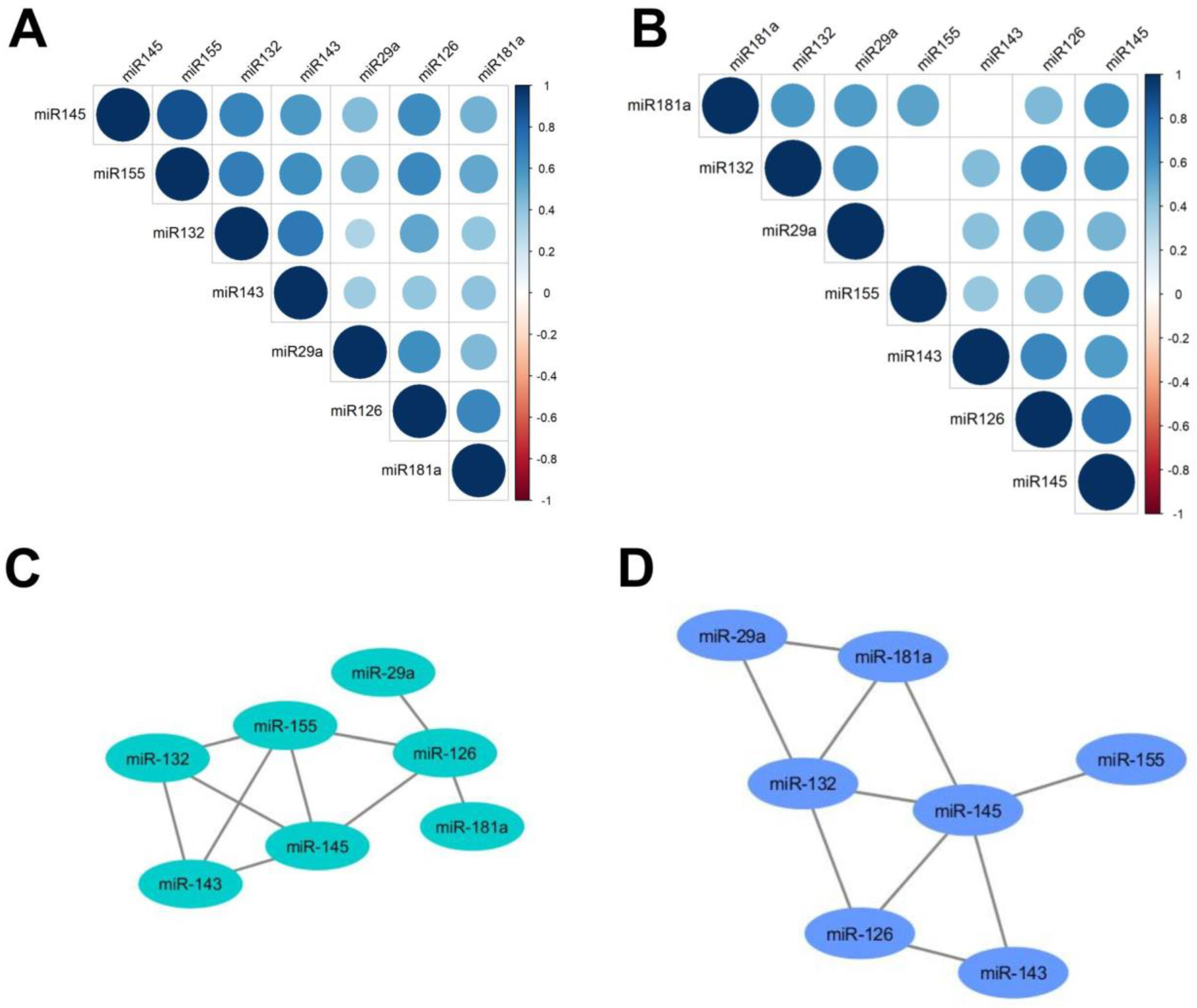
2.2. Upregulation of miR-132, -143, -145 and -155 and Downregulation of miR-181a in the Serum of SSc Patients
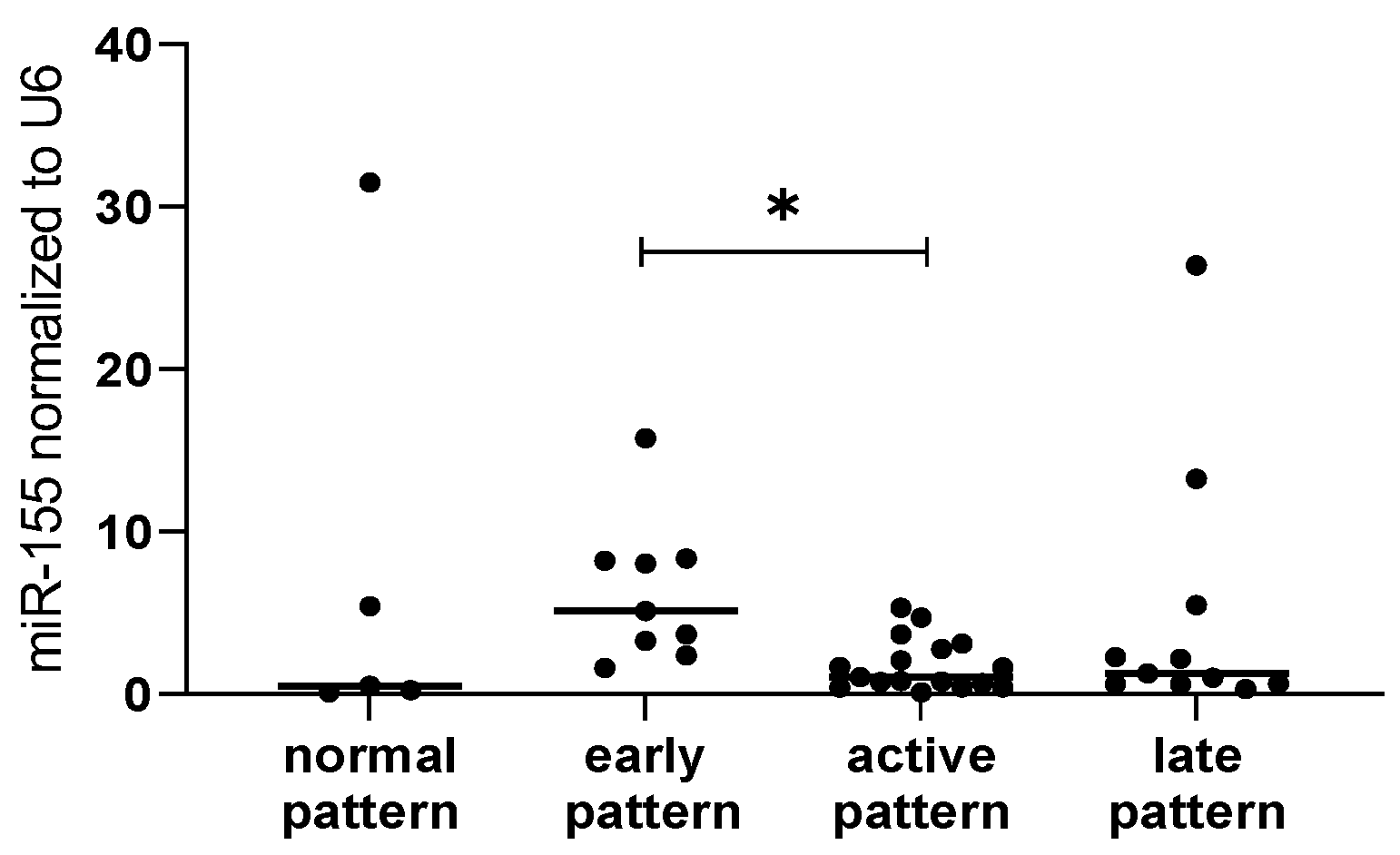
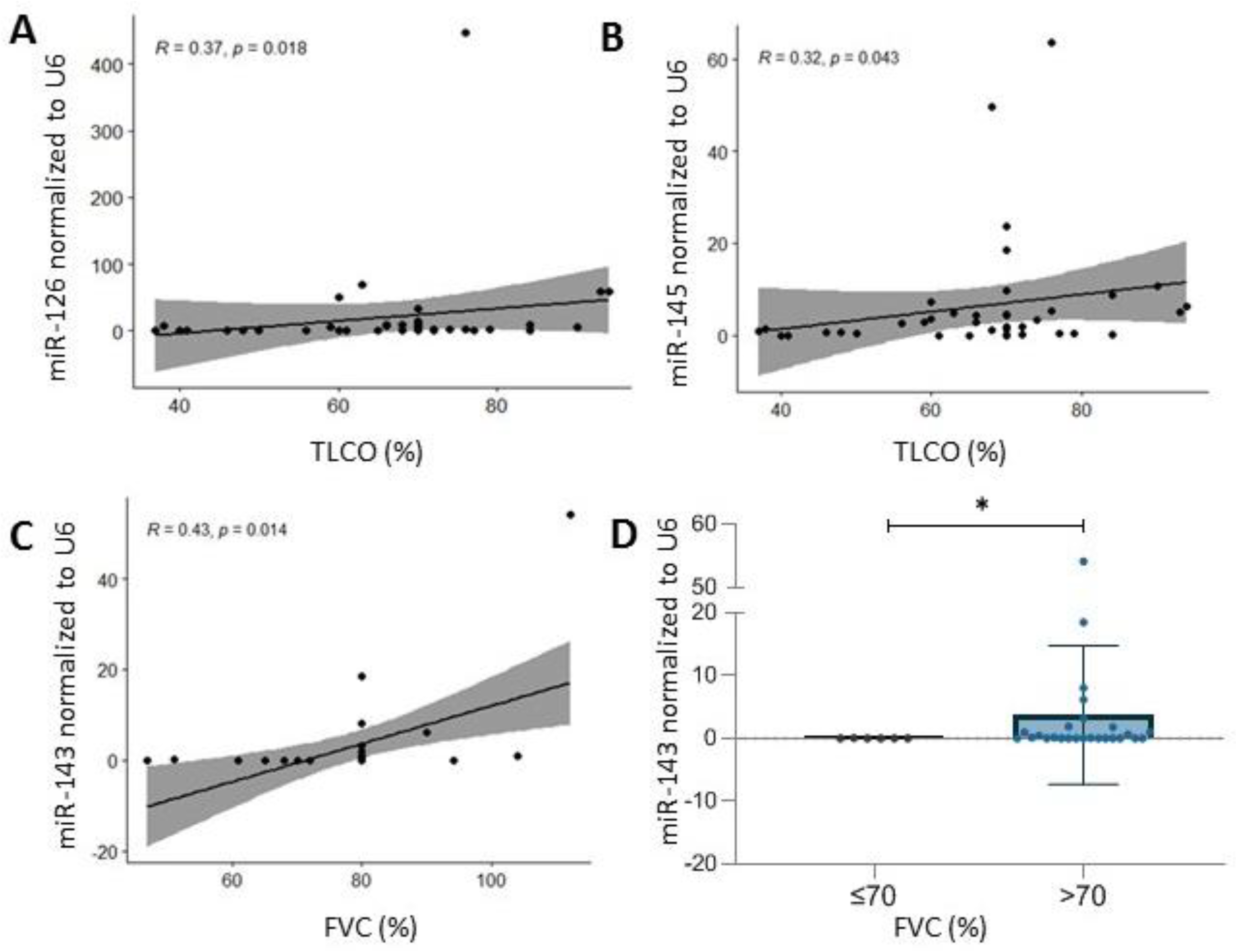
2.3. microRNA and Clinical Association-Specific microRNAs Correlate with Microvascular Damage, TLCO and FVC
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Analysis of miRNA Expression
4.3. Statystical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Denton, C.P.; Khanna, D. Systemic sclerosis. Lancet 2017, 390, 1685–1699. [Google Scholar] [CrossRef]
- Pattanaik, D.; Brown, M.; Postlethwaite, B.C.; Postlethwaite, A.E. Pathogenesis of systemic sclerosis. Front. Immunol. 2015, 6, 272. [Google Scholar] [CrossRef] [PubMed]
- Feghali-Bostwick, C.; Medsger, T.A.; Wright, T.M. Analysis of systemic sclerosis in twins reveals low concordance for disease and high concordance for the presence of antinuclear antibodies. Arthritis Rheum. 2003, 48, 1956–1963. [Google Scholar] [CrossRef]
- Juhl, P.; Bondesen, S.; Hawkins, C.L.; Karsdal, M.A.; Bay-Jensen, A.C.; Davies, M.J.; Siebuhr, A.S. Dermal fibroblasts have different extracellular matrix profiles induced by TGF-β, PDGF and IL-6 in a model for skin fibrosis. Sci. Rep. 2020, 10, 17300. [Google Scholar] [CrossRef] [PubMed]
- Wei, J. Regulation of Matrix Remodeling by Peroxisome Proliferator-Activated Receptor-γ: A Novel Link Between Metabolism and Fibrogenesis. Open Rheumatol. J. 2012, 6, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.N. Interactions between TGF-β1, canonical WNT/β-catenin pathway and PPAR γ in radiation-induced fibrosis. Oncotarget 2017, 8, 90579–90604. [Google Scholar] [CrossRef]
- Distler, O.; Del Rosso, A.; Giacomelli, R.; Cipriani, P.; Conforti, M.L.; Guiducci, S.; Gay, R.E.; Michel, B.A.; Brühlmann, P.; Müller-Ladner, U.; et al. Angiogenic and angiostatic factors in systemic sclerosis: Increased levels of vascular endothelial growth factor are a feature of the earliest disease stages and are associated with the absence of fingertip ulcers. Arthritis Res. 2002, 4, R11. [Google Scholar] [CrossRef]
- Truchetet, M.E.; Brembilla, N.C.; Chizzolini, C. Current Concepts on the Pathogenesis of Systemic Sclerosis. Clin. Rev. Allergy Immunol. 2021. [Google Scholar] [CrossRef]
- Kahaleh, B. Vascular Disease in Scleroderma: Mechanisms of Vascular Injury. Rheum. Dis. Clin. N. Am. 2008, 34, 57–71. [Google Scholar] [CrossRef]
- Moroncini, G.; Mori, S.; Tonnini, C.; Gabrielli, A. Role of viral infections in the etiopathogenesis of systemic sclerosis. Clin. Exp. Rheumatol. 2013, 2, 3–7. [Google Scholar]
- Farina, A.; Cirone, M.; York, M.; Lenna, S.; Padilla, C.; McLaughlin, S.; Faggioni, A.; Lafyatis, R.; Trojanowska, M.; Farina, G.A. Epstein-barr virus infection induces aberrant TLR activation pathway and fibroblast-myofibroblast conversion in Scleroderma. J. Investig. Dermatol. 2014, 134, 954–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, A.M.; Majidi, J.; Baradaran, B.; Yousefi, M. Toll-like receptors in the pathogenesis of autoimmune diseases. Adv. Pharm. Bull. 2015, 5, 605–614. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum microRNAs are promising novel biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef] [PubMed]
- Pascut, D.; Krmac, H.; Gilardi, F.; Patti, R.; Calligaris, R.; Crocè, L.S.; Tiribelli, C. A comparative characterization of the circulating miRNome in whole blood and serum of HCC patients. Sci. Rep. 2019, 9, 8265. [Google Scholar] [CrossRef] [PubMed]
- Stypinska, B.; Wajda, A.; Walczuk, E.; Olesinska, M.; Lewandowska, A.; Walczyk, M.; Paradowska-Gorycka, A. The Serum Cell-Free microRNA Expression Profile in MCTD, SLE, SSc, and RA Patients. J. Clin. Med. 2020, 9, 161. [Google Scholar] [CrossRef]
- Chouri, E.; Servaas, N.H.; Bekker, C.P.J.; Affandi, A.J.; Cossu, M.; Hillen, M.R.; Angiolilli, C.; Mertens, J.S.; van den Hoogen, L.L.; Silva-Cardoso, S.; et al. Serum microRNA screening and functional studies reveal miR-483-5p as a potential driver of fibrosis in systemic sclerosis. J. Autoimmun. 2018, 89, 162–170. [Google Scholar] [CrossRef]
- Steen, S.O.; Iversen, L.V.; Carlsen, A.L.; Burton, M.; Nielsen, C.T.; Jacobsen, S.; Heegaard, N.H.H. The circulating cell-free microRNA profile in systemic sclerosis is distinct from both healthy controls and systemic lupus erythematosus. J. Rheumatol. 2015, 42, 214–221. [Google Scholar] [CrossRef]
- Huang, Y.-X.; Li, F.; Liu, D.; Sun, Y.-Y.; Zhao, Q.-H.; Jiang, R.; Wang, L.; Yuan, P.; Liu, J.-M.; Wu, Y.; et al. The expression profiling of microRNA in systemic sclerosis-associated pulmonary arterial hypertension. Ann. Transl. Med. 2021, 9, 1458. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, H.; Zhao, M.; Lu, Q. Meta-analysis of differentially expressed microRNAs in systemic sclerosis. Int. J. Rheum. Dis. 2020, 23, 1297–1304. [Google Scholar] [CrossRef]
- Maurer, B.; Stanczyk, J.; Jüngel, A.; Akhmetshina, A.; Trenkmann, M.; Brock, M.; Kowal-Bielecka, O.; Gay, R.E.; Michel, B.A.; Distler, J.H.W.; et al. MicroRNA-29, a key regulator of collagen expression in systemic sclerosis. Arthritis Rheum. 2010, 62, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Wuttge, D.M.; Carlsen, A.L.; Teku, G.; Steen, S.O.; Wildt, M.; Vihinen, M.; Hesselstrand, R.; Heegaard, N.H.H. Specific autoantibody profiles and disease subgroups correlate with circulating micro-RNA in systemic sclerosis. Rheumatology 2015, 54, 2100–2107. [Google Scholar] [CrossRef] [PubMed]
- Czömpöly, T.; Simon, D.; Czirják, L.; Németh, P. Anti-topoisomerase I autoantibodies in systemic sclerosis. Autoimmun. Rev. 2009, 8, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Shen, Y.; Hong, Y.; Zhang, Z.; Zhai, Q.; Fu, Z.; Li, H.; Lu, K.; Lin, J. miR-181a regulates the host immune response against Schistosoma japonicum infection through the TLR4 receptor pathway. Parasites Vectors 2021, 14, 548. [Google Scholar] [CrossRef]
- Xie, W.; Li, M.; Xu, N.; Lv, Q.; Huang, N.; He, J.; Zhang, Y. miR-181a Regulates Inflammation Responses in Monocytes and Macrophages. PLoS ONE 2013, 8, e58639. [Google Scholar] [CrossRef]
- Carlsen, A.L.; Schetter, A.J.; Nielsen, C.T.; Lood, C.; Knudsen, S.; Voss, A.; Harris, C.C.; Hellmark, T.; Jacobsen, S.; Bengtsson, A.A.; et al. Circulating microRNA expression profiles associated with systemic lupus erythematosus. Arthritis Rheum. 2013, 65, 1324–1334. [Google Scholar] [CrossRef]
- Li, H.S.; Ning, Y.; Li, S.B.; Shao, P.Y.; Chen, S.J.; Ye, Q.; Heng, X. Expression and clinical significance of miR-181a and miR-203 in systemic lupus erythematosus patients. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4790–4796. [Google Scholar]
- van den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A.J.; Carreira, P.E.; et al. 2013 classification criteria for systemic sclerosis: An American college of rheumatology/European league against rheumatism collaborative initiative. Ann. Rheum. Dis. 2013, 72, 1747–1755. [Google Scholar] [CrossRef]
- Cutolo, M.; Sulli, A.; Pizzorni, C.; Accardo, S. Nailfold videocapillaroscopy assessment of microvascular damage in systemic sclerosis. J. Rheumatol. 2000, 27, 155–160. [Google Scholar]
- Elton, T.S.; Selemon, H.; Elton, S.M.; Parinandi, N.L. Regulation of the MIR155 host gene in physiological and pathological processes. Gene 2013, 532, 1–12. [Google Scholar] [CrossRef]
- Eissa, M.G.; Artlett, C.M. The microRNA miR-155 is essential in fibrosis. Non-Coding RNA 2019, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Christmann, R.B.; Wooten, A.; Sampaio-Barros, P.; Borges, C.L.; Carvalho, C.R.R.; Kairalla, R.A.; Feghali-Bostwick, C.; Ziemek, J.; Mei, Y.; Goummih, S.; et al. miR-155 in the progression of lung fibrosis in systemic sclerosis. Arthritis Res. Ther. 2016, 18, 155. [Google Scholar] [CrossRef] [PubMed]
- Artlett, C.M.; Sassi-Gaha, S.; Hope, J.L.; Feghali-Bostwick, C.A.; Katsikis, P.D. Mir-155 is overexpressed in systemic sclerosis fibroblasts and is required for NLRP3 inflammasome-mediated collagen synthesis during fibrosis. Arthritis Res. Ther. 2017, 19, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankratz, F.; Bemtgen, X.; Zeiser, R.; Leonhardt, F.; Kreuzaler, S.; Hilgendorf, I.; Smolka, C.; Helbing, T.; Hoefer, I.; Esser, J.S.; et al. MicroRNA-155 Exerts Cell-Specific Antiangiogenic but Proarteriogenic Effects During Adaptive Neovascularization. Circulation 2015, 131, 1575–1589. [Google Scholar] [CrossRef] [PubMed]
- Vigorito, E.; Perks, K.L.; Abreu-Goodger, C.; Bunting, S.; Xiang, Z.; Kohlhaas, S.; Das, P.P.; Miska, E.A.; Rodriguez, A.; Bradley, A.; et al. microRNA-155 Regulates the Generation of Immunoglobulin Class-Switched Plasma Cells. Immunity 2007, 27, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Vigorito, E.; Clare, S.; Warren, M.V.; Couttet, P.; Soond, D.R.; Van Dongen, S.; Grocock, R.J.; Das, P.P.; Miska, E.A.; et al. Requirement of bic/microRNA-155 for normal immune function. Science 2007, 316, 608–611. [Google Scholar] [CrossRef]
- Tashkin, D.P.; Roth, M.D.; Clements, P.J.; Furst, D.E.; Khanna, D.; Kleerup, E.C.; Goldin, J.; Arriola, E.; Volkmann, E.R.; Kafaja, S.; et al. Mycophenolate mofetil versus oral cyclophosphamide in scleroderma-related interstitial lung disease (SLS II): A randomised controlled, double-blind, parallel group trial. Lancet Respir. Med. 2016, 4, 708–719. [Google Scholar] [CrossRef]
- Le Gouellec, N.; Duhamel, A.; Perez, T.; Hachulla, A.L.; Sobanski, V.; Faivre, J.B.; Morell-Dubois, S.; Lambert, M.; Hatron, P.Y.; Hachulla, E.; et al. Predictors of lung function test severity and outcome in systemic sclerosis-associated interstitial lung disease. PLoS ONE 2017, 12, e0181692. [Google Scholar] [CrossRef]
- Jordan, N.P.; Tingle, S.J.; Shuttleworth, V.G.; Cooke, K.; Redgrave, R.E.; Singh, E.; Glover, E.K.; Ahmad Tajuddin, H.B.; Kirby, J.A.; Arthur, H.M.; et al. Mir-126-3p is dynamically regulated in endothelial-to-mesenchymal transition during fibrosis. Int. J. Mol. Sci. 2021, 22, 8629. [Google Scholar] [CrossRef]
- Qu, M.-J.; Pan, J.-J.; Shi, X.-J.; Zhang, Z.-J.; Tang, Y.-H.; Yang, G.-Y. MicroRNA-126 is a prospective target for vascular disease. Neuroimmunol. Neuroinflammation 2018, 5, 10. [Google Scholar] [CrossRef]
- Liakouli, V.; Cipriani, P.; Di Benedetto, P.; Panzera, N.; Ruscitti, P.; Pantano, I.; Berardicurti, O.; Carubbi, F.; Esteves, F.; Mavria, G.; et al. Epidermal Growth Factor Like-domain 7 and miR-126 are abnormally expressed in diffuse Systemic Sclerosis fibroblasts. Sci. Rep. 2019, 9, 4589. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, Y.; Qu, S.; Luo, H.; Zhou, Y.; Wang, Y.; Zhao, H.; You, Y.; Xiao, X.; Zuo, X. MicroRNA expression abnormalities in limited cutaneous scleroderma and diffuse cutaneous scleroderma. J. Clin. Immunol. 2012, 32, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Ly, T.-D.; Riedel, L.; Fischer, B.; Schmidt, V.; Hendig, D.; Distler, J.; Kuhn, J.; Knabbe, C.; Faust, I. microRNA-145 mediates xylosyltransferase-I induction in myofibroblasts via suppression of transcription factor KLF4. Biochem. Biophys. Res. Commun. 2020, 523, 1001–1006. [Google Scholar] [CrossRef]
- Malaab, M.; Renaud, L.; Takamura, N.; Zimmerman, K.D.; da Silveira, W.A.; Ramos, P.S.; Haddad, S.; Peters-Golden, M.; Penke, L.R.; Wolf, B.; et al. Antifibrotic factor KLF4 is repressed by the miR-10/TFAP2A/TBX5 axis in dermal fibroblasts: Insights from twins discordant for systemic sclerosis. Ann. Rheum. Dis. 2022, 81, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Men, R.; Wen, M.; Zhao, M.; Dan, X.; Yang, Z.; Wu, W.; Wang, M.H.; Liu, X.; Yang, L. MircoRNA-145 promotes activation of hepatic stellate cells via targeting krüppel-like factor 4. Sci. Rep. 2017, 7, 40468. [Google Scholar] [CrossRef]
- Xiao, X.; Tang, W.; Yuan, Q.; Peng, L.; Yu, P. Epigenetic repression of Krüppel-like factor 4 through Dnmt1 contributes to EMT in renal fibrosis. Int. J. Mol. Med. 2015, 35, 1596–1602. [Google Scholar] [CrossRef]
- Lai, X.; Vera, J. MicroRNA Clusters. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.-H., Yokota, H., Eds.; Springer: New York, NY, USA, 2013; pp. 1310–1314. ISBN 978-1-4419-9863-7. [Google Scholar]
- Xin, M.; Small, E.M.; Sutherland, L.B.; Qi, X.; McAnally, J.; Plato, C.F.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. MicroRNAs miR-143 and miR-145 modulate cytoskeletal dynamics and responsiveness of smooth muscle cells to injury. Genes Dev. 2009, 23, 2166–2178. [Google Scholar] [CrossRef]
- Tu, H.; Chen, D.; Cai, C.; Du, Q.; Lin, H.; Pan, T.; Sheng, L.; Xu, Y.; Teng, T.; Tu, J.; et al. microRNA-143-3p attenuated development of hepatic fibrosis in autoimmune hepatitis through regulation of TAK1 phosphorylation. J. Cell. Mol. Med. 2020, 24, 1256–1267. [Google Scholar] [CrossRef]
- Diazzi, S.; Baeri, A.; Fassy, J.; Lecacheur, M.; Marin-Bejar, O.; Girard, C.A.; Lefevre, L.; Lacoux, C.; Irondelle, M.; Mounier, C.; et al. Blockade of the pro-fibrotic reaction mediated by the miR-143/-145 cluster enhances the responses to targeted therapy in melanoma. EMBO Mol. Med. 2022, 14, e15295. [Google Scholar] [CrossRef]
- Liu, X.; Gong, J.; Xu, B. miR-143 down-regulates TLR2 expression in hepatoma cells and inhibits hepatoma cell proliferation and invasion. Int. J. Clin. Exp. Pathol. 2015, 8, 12738–12747. [Google Scholar]
- Andonian, B.J.; Chou, C.-H.; Ilkayeva, O.R.; Koves, T.R.; Connelly, M.A.; Kraus, W.E.; Kraus, V.B.; Huffman, K.M. Plasma MicroRNAs in Established Rheumatoid Arthritis Relate to Adiposity and Altered Plasma and Skeletal Muscle Cytokine and Metabolic Profiles. Front. Immunol. 2019, 10, 1475. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.G.; Zhang, W.; Ma, H.J.; Guo, Z.Y.; Xu, Y. Silencing of LncRNA TCONS_00088786 reduces renal fibrosis through miR-132. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, R.; Ruan, Z.; Liu, L.; Li, Y.; Zhu, L. MicroRNA-132 attenuated cardiac fibrosis in myocardial infarction-induced heart failure rats. Biosci. Rep. 2020, 40, BSR20201696. [Google Scholar] [CrossRef] [PubMed]
- Täubel, J.; Hauke, W.; Rump, S.; Viereck, J.; Batkai, S.; Poetzsch, J.; Rode, L.; Weigt, H.; Genschel, C.; Lorch, U.; et al. Novel antisense therapy targeting microRNA-132 in patients with heart failure: Results of a first-in-human Phase 1b randomized, double-blind, placebo-controlled study. Eur. Heart J. 2020, 42, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Catalano, D.; Talis, A.; Szabo, G.; Bala, S. Protective effect of LNA-anti-miR-132 therapy on liver fibrosis in mice. Mol. Ther.-Nucleic Acids 2021, 25, 155–167. [Google Scholar] [CrossRef]
- Henderson, J.; Brown, M.; Horsburgh, S.; Duffy, L.; Wilkinson, S.; Worrell, J.; Stratton, R.; O’Reilly, S. Methyl cap binding protein 2: A key epigenetic protein in systemic sclerosis. Rheumatology 2019, 58, 527–535. [Google Scholar] [CrossRef]
- Fang, Q.; Tian, M.; Wang, F.; Zhang, Z.; Du, T.; Wang, W.; Yang, Y.; Li, X.; Chen, G.; Xiao, L.; et al. Amlodipine induces vasodilation via Akt2/Sp1-activated miR-21 in smooth muscle cells. Br. J. Pharmacol. 2019, 176, 2306–2320. [Google Scholar] [CrossRef]
- Yang, J.; Si, D.; Zhao, Y.; He, C.; Yang, P. S-amlodipine improves endothelial dysfunction via the RANK/RANKL/OPG system by regulating microRNA-155 in hypertension. Biomed. Pharmacother. 2019, 114, 108799. [Google Scholar] [CrossRef]
- Chiral Drugs–S-Amlodipine, Calcium Channel Blocker. Available online: http://www.chiralemcure.com/pop/S-amlodipine_globalpresence.html (accessed on 9 September 2022).
- Dalal, J.; Mohan, J.C.; Iyengar, S.S.; Hiremath, J.; Sathyamurthy, I.; Bansal, S.; Kahali, D.; Dasbiswas, A. S-Amlodipine: An Isomer with Difference-Time to Shift from Racemic Amlodipine. Int. J. Hypertens. 2018, 2018, 8681792. [Google Scholar] [CrossRef]
- Iwamoto, N.; Furukawa, K.; Endo, Y.; Shimizu, T.; Sumiyoshi, R.; Umeda, M.; Koga, T.; Kawashiri, S.Y.; Igawa, T.; Ichinose, K.; et al. Methotrexate alters the expression of microrna in fibroblast-like synovial cells in rheumatoid arthritis. Int. J. Mol. Sci. 2021, 22, 11561. [Google Scholar] [CrossRef]
- Yang, D.; Haemmig, S.; Zhou, H.; Pérez-Cremades, D.; Sun, X.; Chen, L.; Li, J.; Haneo-Mejia, J.; Yang, T.; Hollan, I.; et al. Methotrexate attenuates vascular inflammation through an adenosine-microRNA-dependent pathway. Elife 2021, 10, e58064. [Google Scholar] [CrossRef] [PubMed]
- McAlexander, M.A.; Phillips, M.J.; Witwer, K.W. Comparison of methods for miRNA extraction from plasma and quantitative recovery of RNA from cerebrospinal fluid. Front. Genet. 2013, 4, 83. [Google Scholar] [CrossRef] [PubMed]
- Dufourd, T.; Robil, N.; Mallet, D.; Carcenac, C.; Boulet, S.; Brishoual, S.; Rabois, E.; Houeto, J.L.; De La Grange, P.; Carnicella, S. Plasma or serum? A qualitative study on rodents and humans using high-throughput microRNA sequencing for circulating biomarkers. Biol. Methods Protoc. 2019, 4, bpz006. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.; Herrick, A.L.; Ingegnoli, F.; Damjanov, N.; De Angelis, R.; Denton, C.P.; Distler, O.; Espejo, K.; Foeldvari, I.; Frech, T.; et al. Standardisation of nailfold capillaroscopy for the assessment of patients with Raynaud’s phenomenon and systemic sclerosis. Autoimmun. Rev. 2020, 19, 102458. [Google Scholar] [CrossRef] [PubMed]
- Pasławski, M.; Kurys, E.; Złomaniec, J. Differentiation of linear and reticular opacities in high resolution computed tomography (HRCT) in interstitial lung diseases. Ann. Univ. Mariae Curie-Skłodowska. Sect. D Med. 2003, 58, 378–385. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | SSc | lcSSc | dcSSc |
|---|---|---|---|
| Sex, female: n (%) | 34 (75.56) | 22 (48.89) | 12 (26.67) |
| Sex, male: n (%) | 11 (24.44) | 5 (11.11) | 6 (13.33) |
| Age, years: mean ± SD | 51 ± 13 | 54 ± 10,4 | 47 ± 15 |
| BMI | 24.9 (17.1–40.9) | 26.7 ± 5.9 | 24.2 (17.1–40.9) |
| Disease duration, years: median (min–max) | 6 (0–38) | 11 (1–38) | 3 (0–20) |
| Immunosuppressive therapy: n (%) | 28 (56%) | 15 (51.85%) | 13 (58.82%) |
| mRSS median (min-max) | 7 (2–30) | 5 (2–19) | 11 (2–30) |
| Scleroderma pattern in NVC | 37 (88.1%) | 21 (84%) | 16 (94.12%) |
| Presence of RF, n (%) | 3 (9) | 3 (15) | 0 |
| Hepatitis type B | 0 | 0 | 0 |
| Hepatitis type C | 0 | 0 | 0 |
| Elevated ESR, n (%) | 24 (55) | 14 (53.85) | 10 (58.82) |
| ESR, median (min–max) | 14 (2–78) | 13 (2–78) | 14 (4–53) |
| Elevated CRP, n (%) | 6 (13) | 4 (14.81) | 2 (11.76) |
| CRP, median (min–max) | 3 (1–52) | 3 (1–52) | 5 (1–45) |
| Presence of ANA, n (%) | 43 (93.33) | 25 (92.59) | 18 (100) |
| ANA range (min–max) | 1:80–1:40960 | 1:80–1:40960 | 1:320–1:10280 |
| Anti-TOPO-I | 21 (54%) | 6 (33,3%) | 15 (82,35%) |
| CENP | 12 (31%) | 12 (33.3%) | 0 (0%) |
| Variable | lcSSc n (%) | dcSSc n (%) | p * | SSc n (%) |
|---|---|---|---|---|
| Raynaud’s phenomenon | 27 (100) | 18 (100) | >0.99 | 45 (100) |
| Digital ulcers | 11 (40.74) | 8 (44.44) | 0.76 | 19 (42.22) |
| Digestive system involvement | 12 (44.44) | 10 (55.56) | 0.77 | 22 (48.89) |
| Interstitial lung disease | 15 (55.56) | 15 (83.33) | 0.10 | 30 (66.67) |
| Pulmonary arterial hypertension | 2 (7.41) | 1 (5.56) | 0.36 | 3 (6.67) |
| Heart involvement | 9 (33.33) | 8 (44.44) | >0.99 | 17 (37.78) |
| Arthritis | 10 (37.04) | 6 (33.33) | 0.73 | 16 (35.56) |
| Myositis | 2 (7.41) | 1 (5.56) | 0.55 | 3 (6.67) |
| Renal involvement | 3 (11.11) | 1 (5.56) | >0.99 | 4 (8.89) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wajda, A.; Walczyk, M.; Dudek, E.; Stypińska, B.; Lewandowska, A.; Romanowska-Próchnicka, K.; Chojnowski, M.; Olesińska, M.; Paradowska-Gorycka, A. Serum microRNAs in Systemic Sclerosis, Associations with Digital Vasculopathy and Lung Involvement. Int. J. Mol. Sci. 2022, 23, 10731. https://doi.org/10.3390/ijms231810731
Wajda A, Walczyk M, Dudek E, Stypińska B, Lewandowska A, Romanowska-Próchnicka K, Chojnowski M, Olesińska M, Paradowska-Gorycka A. Serum microRNAs in Systemic Sclerosis, Associations with Digital Vasculopathy and Lung Involvement. International Journal of Molecular Sciences. 2022; 23(18):10731. https://doi.org/10.3390/ijms231810731
Chicago/Turabian StyleWajda, Anna, Marcela Walczyk, Ewa Dudek, Barbara Stypińska, Aleksandra Lewandowska, Katarzyna Romanowska-Próchnicka, Marek Chojnowski, Marzena Olesińska, and Agnieszka Paradowska-Gorycka. 2022. "Serum microRNAs in Systemic Sclerosis, Associations with Digital Vasculopathy and Lung Involvement" International Journal of Molecular Sciences 23, no. 18: 10731. https://doi.org/10.3390/ijms231810731
APA StyleWajda, A., Walczyk, M., Dudek, E., Stypińska, B., Lewandowska, A., Romanowska-Próchnicka, K., Chojnowski, M., Olesińska, M., & Paradowska-Gorycka, A. (2022). Serum microRNAs in Systemic Sclerosis, Associations with Digital Vasculopathy and Lung Involvement. International Journal of Molecular Sciences, 23(18), 10731. https://doi.org/10.3390/ijms231810731