
Beta-Glucan from S. cerevisiae Protected AOM-Induced Colon Cancer in cGAS-Deficient Mice Partly through Dectin-1-Manipulated Macrophage Cell Energy

 ,
, 
Abstract
:1. Introduction
2. Results
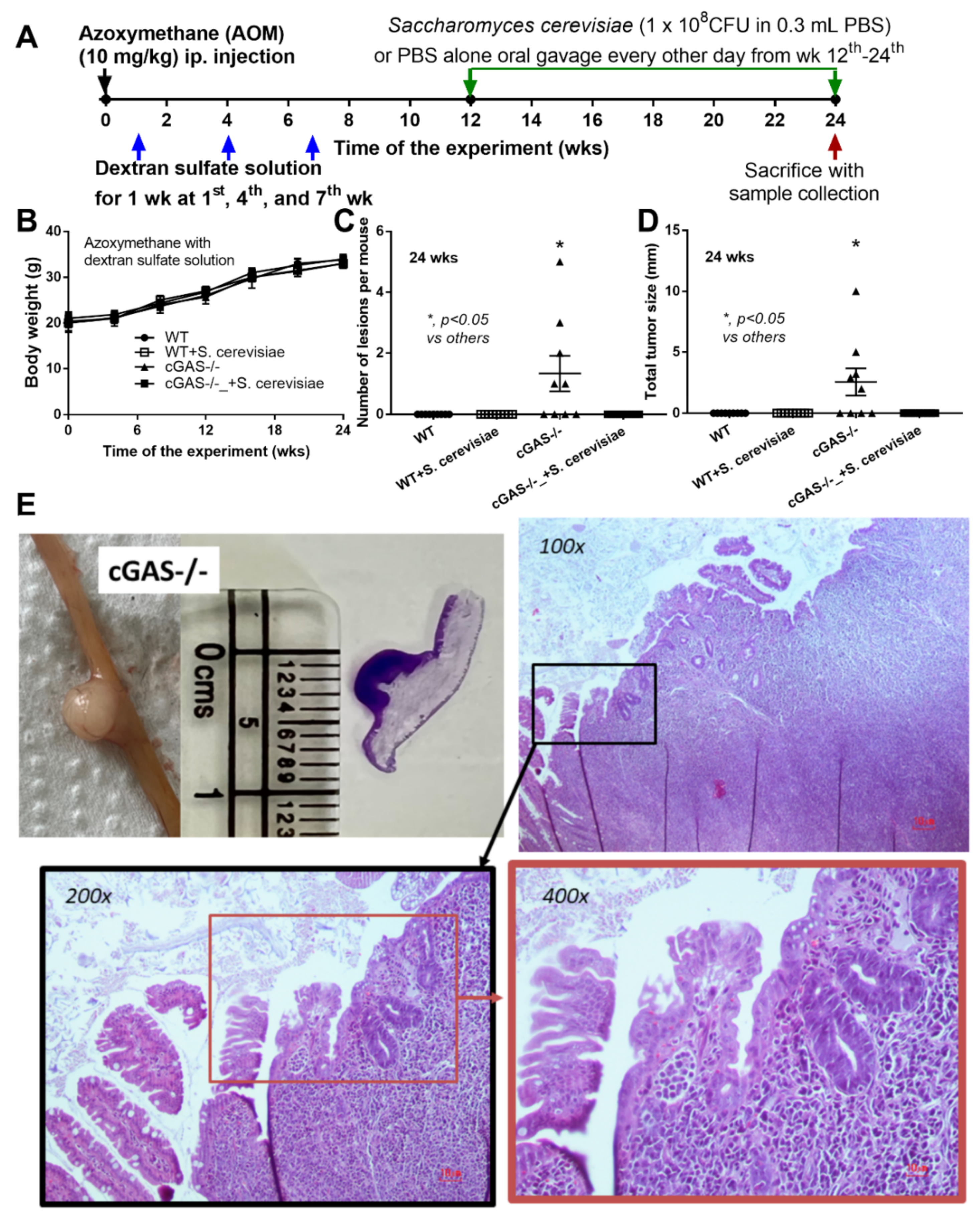
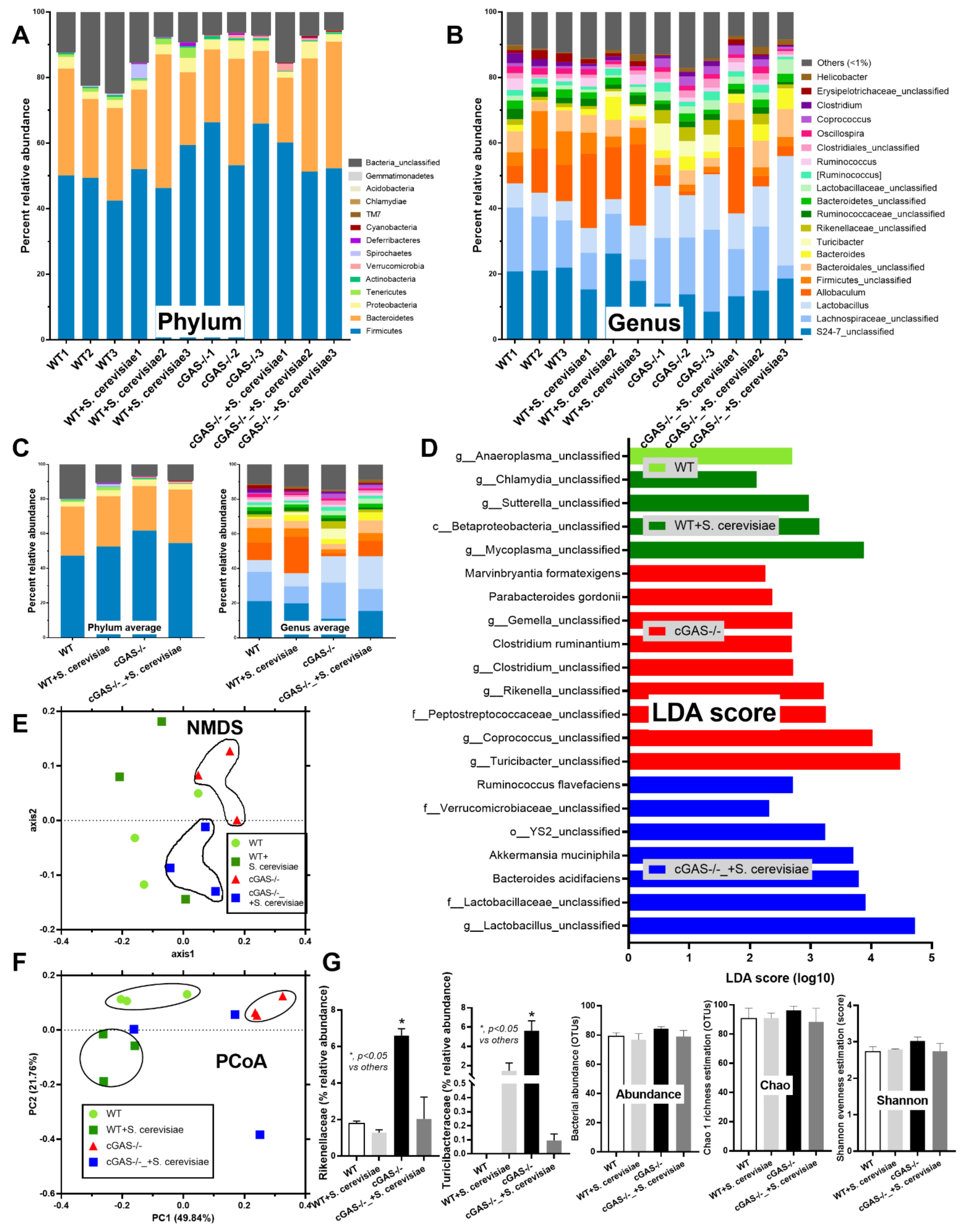
2.1. Azoxymethane Induced Colon Cancer Only in cGAS-Deficient (cGAS-/-) but Not in the Wild-Type nor S. cerevisiae-Administered cGAS-/- Mice Partly through Dysbiosis Attenuation
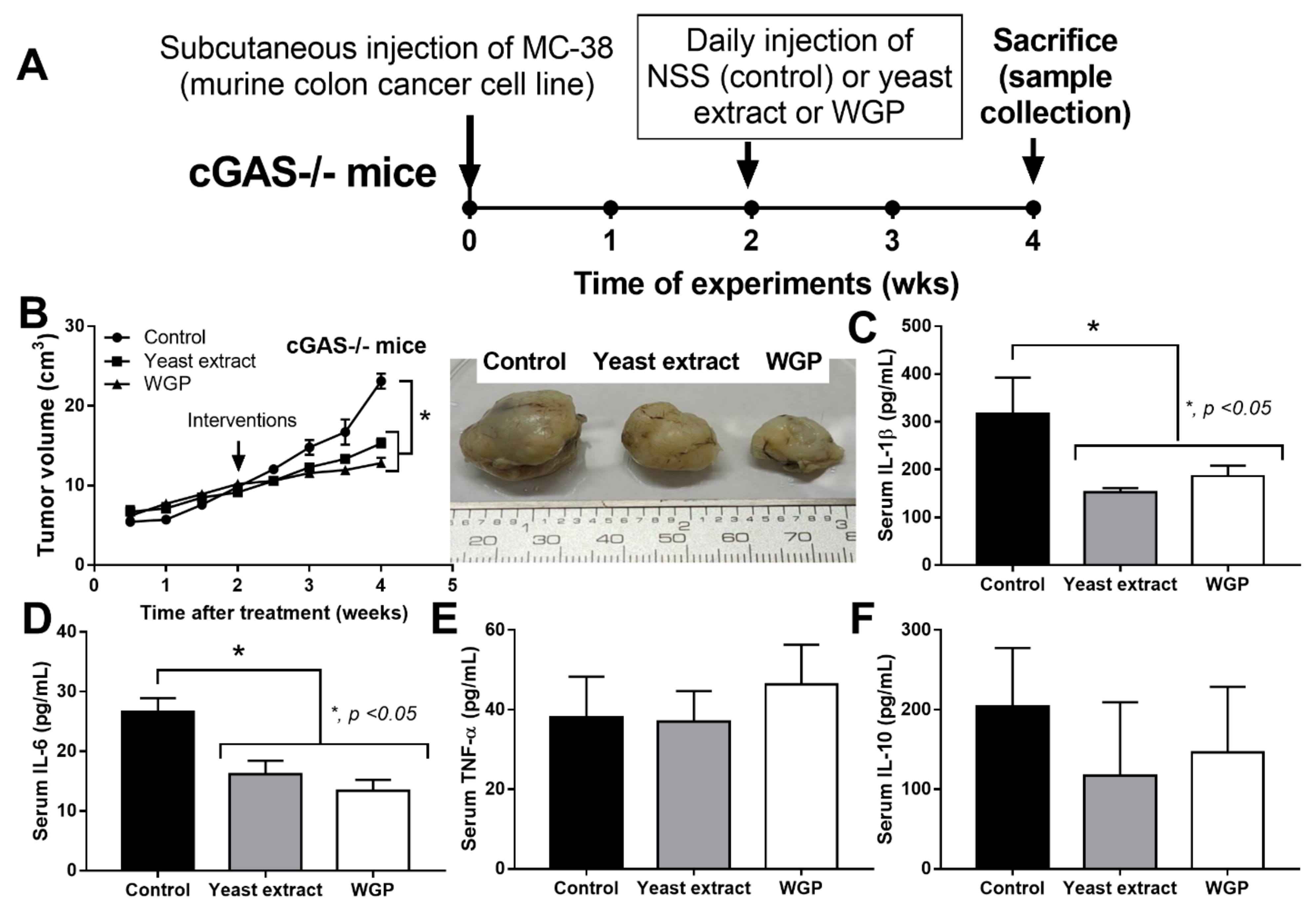
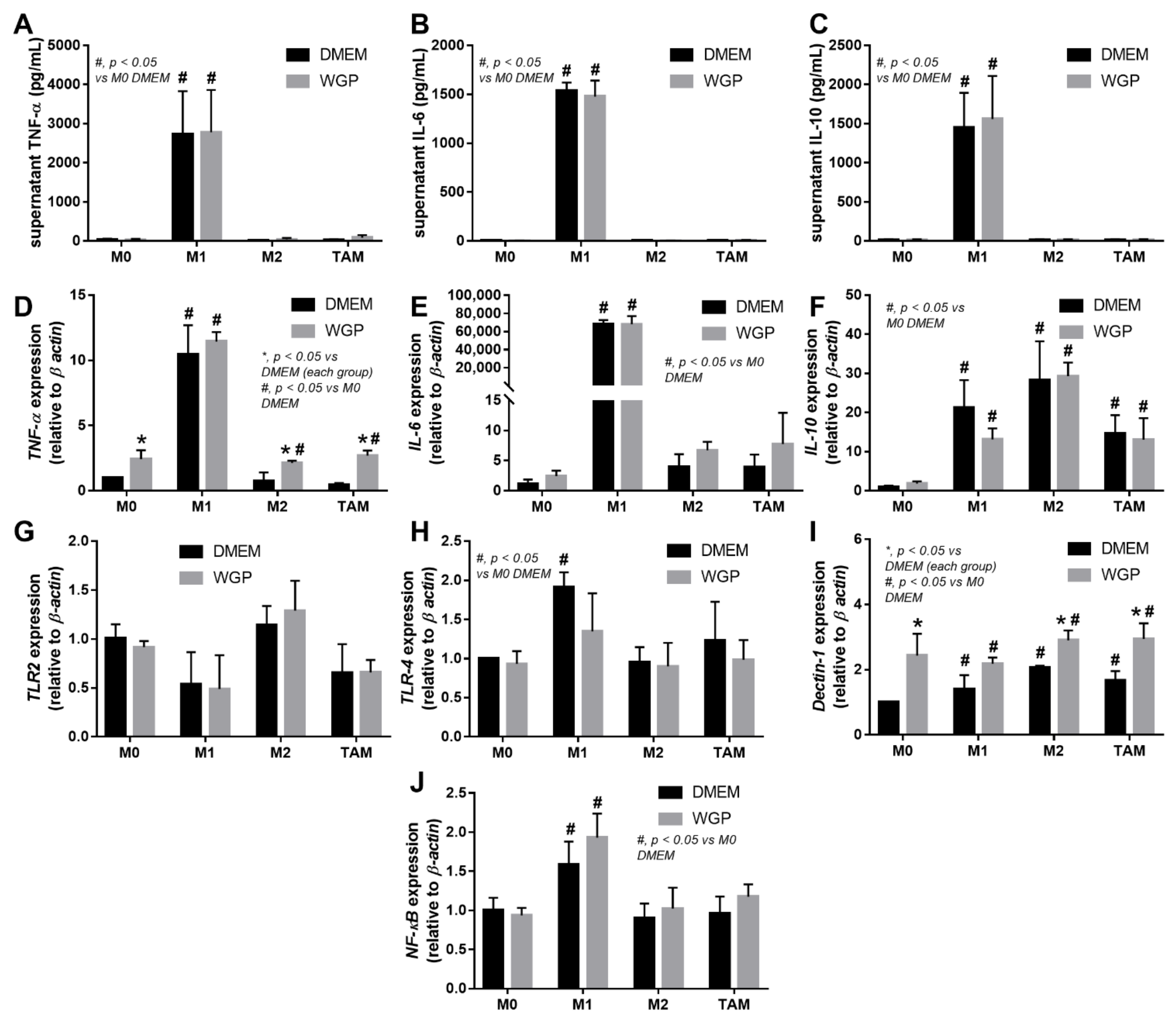
2.2. Intralesional Injection of the Extract from S. cerevisiae or Whole Glucan Particle (WGP) Attenuated Tumor Growth Partly through Macropahge Responses
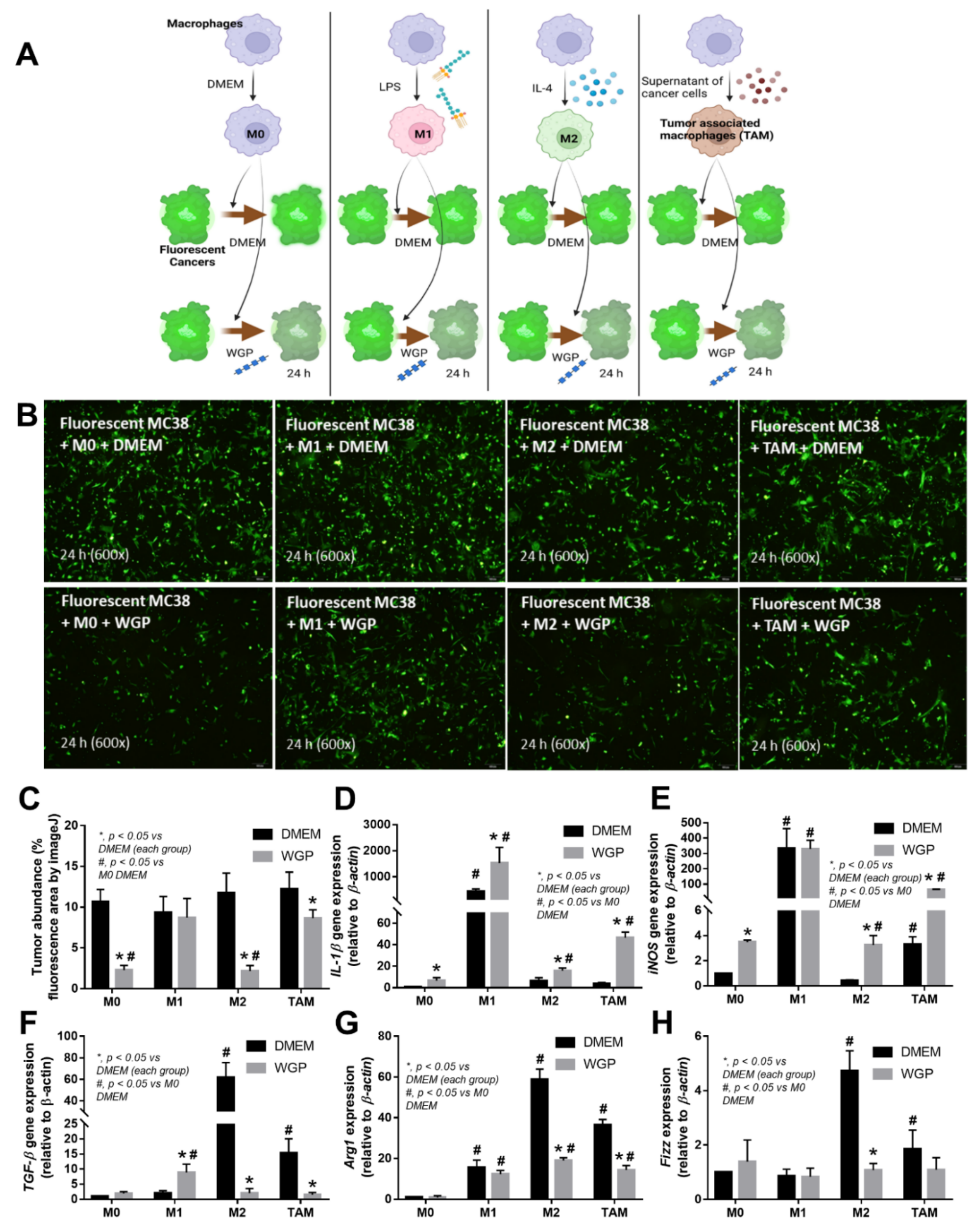
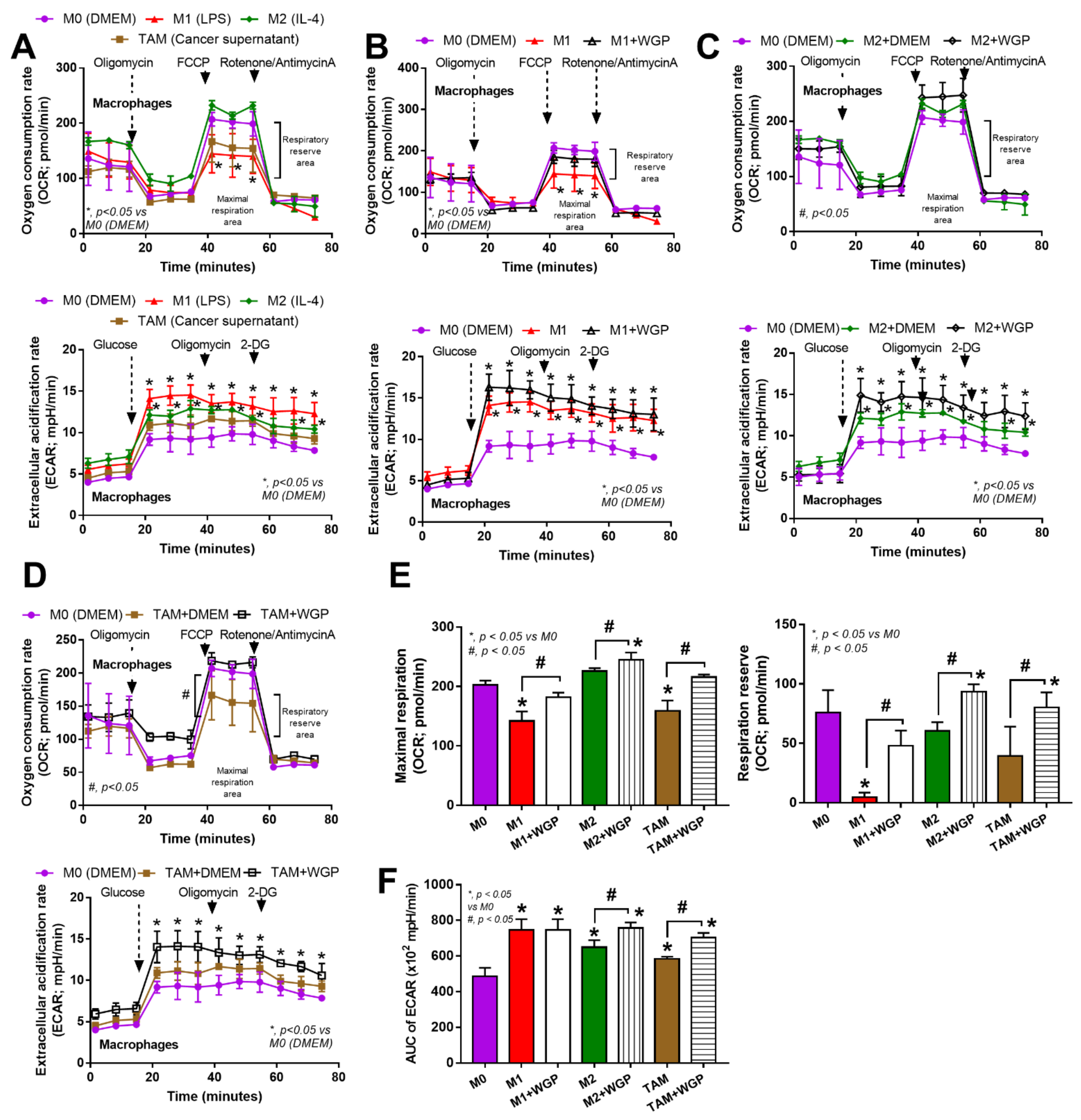
2.3. Cell Energy Status in Different Types of Macrophages and the Impact of WGP
3. Discussion
3.1. Impacts of Environemental Factors and Genetic Susceptibility in Spontaneous Colon Cancer in cGAS-/- Mice and Saccharomyces cerevisiae Administration
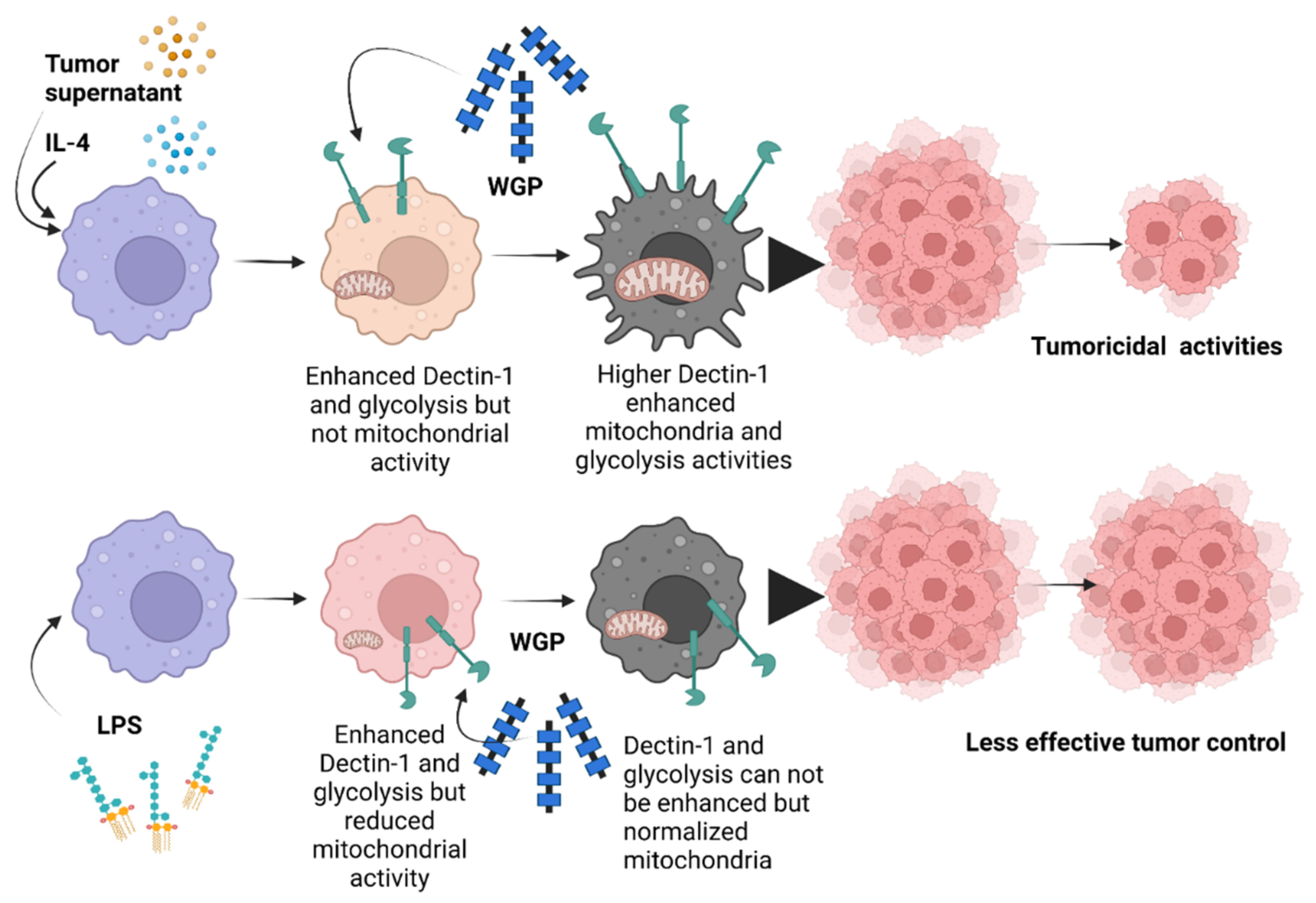
3.2. Beta-Glucan from the Cell Wall of Saccharomyces cerevisiae Attenuated Subcutaneous Tumor Growth Partly through Dectin-1-Mediated Cell Energy Enhancement in Macrophages
4. Materials and Methods
4.1. Animal and Animal Models
4.2. Fecal Microbiome Analysis
4.3. Macrophage Experiments and Fluorescent Labelling Cancer Cells
4.4. Gene Expression, Supernatant Cytokines, and Extracellular Flux Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sambrani, R.; Abdolalizadeh, J.; Kohan, L.; Jafari, B. Saccharomyces Cerevisiae Inhibits Growth and Metastasis and Stimulates Apoptosis In HT-29 Colorectal Cancer Cell Line. Comp. Clin. Pathol. 2018, 28, 985–995. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef]
- Powell, J.B.; Ghotbaddini, M. Cancer-Promoting and Inhibiting Effects of Dietary Compounds: Role of the Aryl Hydrocarbon Receptor (AhR). Biochem. Pharmacol. Open Access 2014, 3, 1000131. [Google Scholar] [CrossRef]
- Górska, A.; Przystupski, D.; Niemczura, M.J.; Kulbacka, J. Probiotic Bacteria: A Promising Tool in Cancer Prevention and Therapy. Curr. Microbiol. 2019, 76, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.J.; Aldhous, M.C.; Drummond, H.E.; Smith, B.R.K.; Nimmo, E.R.; Arnott, I.D.R.; Satsangi, J. Anti-Saccharomyces Cerevisiae Antibodies (ASCA) in Crohn’s Disease are Associated with Disease Severity but Not NOD2/CARD15 Mutations. Clin. Exp. Immunol. 2004, 135, 490–496. [Google Scholar] [CrossRef]
- Li, J.Q.; Xie, Y.H.; Wang, Y.; Shen, X.N.; Qian, Y.; Han, J.X.; Chen, Y.X.; Fang, J. Saccharomyces cerevisiae May Serve as a Probiotic in Colorectal Cancer by Promoting Cancer Cell Apoptosis. J. Dig. Dis. 2020, 21, 571–582. [Google Scholar] [CrossRef]
- Chou, W.K.; Park, J.; Carey, J.B.; McIntyre, D.R.; Berghman, L.R. Immunomodulatory Effects of Saccharomyces cerevisiae Fermentation Product Supplementation on Immune Gene Expression and Lymphocyte Distribution in Immune Organs in Broilers. Front. Veter Sci. 2017, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Meriggi, N.; Di Paola, M.; Vitali, F.; Rivero, D.; Cappa, F.; Turillazzi, F.; Gori, A.; Dapporto, L.; Beani, L.; Turillazzi, S.; et al. Saccharomyces cerevisiae Induces Immune Enhancing and Shapes Gut Microbiota in Social Wasps. Front. Microbiol. 2019, 10, 2320. [Google Scholar] [CrossRef]
- Seif, M.; Hoppstädter, J.; Breinig, F.; Kiemer, A.K. Yeast-Mediated Mrna Delivery Polarizes Immuno-Suppressive Macrophages Towards an Immuno-Stimulatory Phenotype. Eur. J. Pharm. Biopharm. 2017, 117, 1–13. [Google Scholar] [CrossRef]
- Issara-Amphorn, J.; Dang, C.; Saisorn, W.; Limbutara, K.; Leelahavanichkul, A. Candida Administration in Bilateral Nephrectomy Mice Elevates Serum (1→3)-β-D-glucan That Enhances Systemic Inflammation Through Energy Augmentation in Macrophages. Int. J. Mol. Sci. 2021, 22, 5031. [Google Scholar] [CrossRef]
- Li, B.; Cai, Y.; Qi, C.; Hansen, R.; Ding, C.; Mitchell, T.C.; Yan, J. Orally Administered Particulate β-Glucan Modulates Tumor-Capturing Dendritic Cells and Improves Antitumor T-Cell Responses in CancerParticulate β-Glucan Elicits Antitumor T-Cell Responses. Clin. Cancer Res. 2010, 16, 5153–5164. [Google Scholar] [CrossRef]
- Mata-Martínez, P.; Bergón-Gutiérrez, M.; Del Fresno, C. Dectin-1 Signaling Update: New Perspectives for Trained Immunity. Front. Immunol. 2022, 365, 812148. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.; Vinayagam, R.; Anand, M.A.V.; Isa, N.M.; Ponnaiyan, R. Biochemical and Molecular Aspects of 1,2-Dimethylhydrazine (DMH)-Induced Colon Carcinogenesis: A Review. Toxicol. Res. 2020, 9, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Arnesen, H.; Müller, M.H.B.; Aleksandersen, M.; Østby, G.C.; Carlsen, H.; Paulsen, J.E.; Boysen, P. Induction of Colorectal Carcinogenesis in the C57BL/6J And A/J Mouse Strains with a Reduced DSS Dose in the AOM/DSS Model. Lab. Anim. Res. 2021, 37, 1–10. [Google Scholar] [CrossRef]
- Abedi, J.; Saatloo, M.V.; Nejati, V.; Hobbenaghi, R.; Tukmechi, A.; Nami, Y.; Khosroushahi, A.Y. Selenium-Enriched Saccharomyces cerevisiae Reduces the Progression of Colorectal Cancer. Biol. Trace Element Res. 2018, 185, 424–432. [Google Scholar] [CrossRef]
- Garland, K.M.; Rosch, J.C.; Carson, C.S.; Wang-Bishop, L.; Hanna, A.; Sevimli, S.; Van Kaer, C.; Balko, J.M.; Ascano, M.; Wilson, J.T. Pharmacological Activation of cGAS for Cancer Immunotherapy. Front. Immunol. 2021, 12, 753472. [Google Scholar] [CrossRef] [PubMed]
- Wang-Bishop, L.; Wehbe, M.; Shae, D.; James, J.; Hacker, B.C.; Garland, K.; Chistov, P.P.; Rafat, M.; Balko, J.M.; Wilson, J.T. Potent STING Activation Stimulates Immunogenic Cell Death to Enhance Antitumor Immunity in Neuroblastoma. J. Immunother. Cancer 2019, 8, e000282. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Allendorf, D.J.; Brandley, B. Yeast Whole Glucan Particle (WGP) Β-Glucan in Conjunction with Antitumour Monoclonal Antibodies to Treat Cancer. Expert Opin. Biol. Ther. 2005, 5, 691–702. [Google Scholar] [CrossRef]
- Poh, A.R.; Ernst, M. Targeting Macrophages in Cancer: From Bench to Bedside. Front. Oncol. 2018, 8, 49. [Google Scholar] [CrossRef]
- Hu, S.; Fang, Y.; Chen, X.; Cheng, T.; Zhao, M.; Du, M.; Li, T.; Li, M.; Zeng, Z.; Wei, Y.; et al. cGAS Restricts Colon Cancer Development by Protecting Intestinal Barrier Integrity. Proc. Natl. Acad. Sci. USA 2021, 118, e2105747118. [Google Scholar] [CrossRef]
- Ducray, H.A.G.; Globa, L.; Pustovyy, O.; Morrison, E.; Vodyanoy, V.; Sorokulova, I. Yeast Fermentate Prebiotic Improves Intestinal Barrier Integrity During Heat Stress by Modulation of the Gut Microbiota in Rats. J. Appl. Microbiol. 2019, 127, 1192–1206. [Google Scholar] [CrossRef] [PubMed]
- Sivinski, S.E.; Mamedova, L.K.; Rusk, R.A.; Elrod, C.C.; Swartz, T.H.; McGill, J.M.; Bradford, B.J. Development of an In Vitro Macrophage Screening System on the Immunomodulating Effects of Feed Components. J. Anim. Sci. Biotechnol. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Liu, M.; Luo, F.; Ding, C.; Albeituni, S.; Hu, X.; Ma, Y.; Cai, Y.; McNally, L.R.; Sanders, M.A.; Jain, D.; et al. Dectin-1 Activation by a Natural Product β-Glucan Converts Immunosuppressive Macrophages into an M1-like Phenotype. J. Immunol. 2015, 195, 5055–5065. [Google Scholar] [CrossRef]
- Suzuki, R.; Kohno, H.; Sugie, S.; Nakagama, H.; Tanaka, T. Strain Differences in the Susceptibility to Azoxymethane and Dextran Sodium Sulfate-Induced Colon Carcinogenesis in Mice. Carcinogenesis 2005, 27, 162–169. [Google Scholar] [CrossRef]
- Neufert, C.; Becker, C.; Neurath, M.F. An Inducible Mouse Model of Colon Carcinogenesis for the Analysis of Sporadic and Inflammation-Driven Tumor Progression. Nat. Protoc. 2007, 2, 1998–2004. [Google Scholar] [CrossRef]
- De Robertis, M.; Massi, E.; Poeta, M.L.; Carotti, S.; Morini, S.; Cecchetelli, L.; Signori, E.; Fazio, V.M. The AOM/DSS Murine Model for the Study of Colon Carcinogenesis: From Pathways to Diagnosis and Therapy Studies. J. Carcinog. 2011, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Metzger, R.; Maruskova, M.; Krebs, S.; Janssen, K.P.; Krug, A.B. Increased Incidence of Colon Tumors in AOM-Treated Apc 1638N/+ Mice Reveals Higher Frequency of Tumor Associated Neutrophils in Colon Than Small Intestine. Front. Oncol. 2019, 9, 1001. [Google Scholar] [CrossRef]
- Du, J.-M.; Qian, M.-J.; Yuan, T.; Chen, R.-H.; He, Q.-J.; Yang, B.; Ling, Q.; Zhu, H. cGAS and Cancer Therapy: A Double-Edged Sword. Acta Pharmacol. Sin. 2022, 43, 1–10. [Google Scholar] [CrossRef]
- Boonhai, S.; Bootdee, K.; Saisorn, W.; Takkavatakarn, K.; Sitticharoenchai, P.; Tungsanga, S.; Tiranathanagul, K.; Leelahavanichkul, A. TMAO Reductase, A Biomarker for Gut Permeability Defect Induced Inflammation, In Mouse Model of Chronic Kidney Disease and Dextran Sulfate Solution-Induced Mucositis. Asian Pac. J. Allergy Immunol. 2021. [Google Scholar] [CrossRef]
- Amornphimoltham, P.; Yuen, P.S.T.; Star, R.A.; Leelahavanichkul, A. Gut Leakage of Fungal-Derived Inflammatory Mediators: Part of a Gut-Liver-Kidney Axis in Bacterial Sepsis. Am. J. Dig. Dis. 2019, 64, 2416–2428. [Google Scholar] [CrossRef] [PubMed]
- Ondee, T.; Pongpirul, K.; Janchot, K.; Kanacharoen, S.; Lertmongkolaksorn, T.; Wongsaroj, L.; Somboonna, N.; Ngamwongsatit, N.; Leelahavanichkul, A. Lactiplantibacillus Plantarum Dfa1 Outperforms Enterococcus Faecium Dfa1 on Anti-Obesity in High Fat-Induced Obesity Mice Possibly Through the Differences in Gut Dysbiosis Attenuation, Despite the Similar Anti-Inflammatory Properties. Nutrients 2021, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Ondee, T.; Pongpirul, K.; Visitchanakun, P.; Saisorn, W.; Kanacharoen, S.; Wongsaroj, L.; Kullapanich, C.; Ngamwongsatit, N.; Settachaimongkon, S.; Somboonna, N.; et al. Lactobacillus Acidophilus LA5 Improves Saturated Fat-Induced Obesity Mouse Model Through the Enhanced Intestinal Akkermansia Muciniphila. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Panpetch, W.; Sawaswong, V.; Chanchaem, P.; Ondee, T.; Dang, C.P.; Payungporn, S.; Tumwasorn, S.; Leelahavanichkul, A. Candida Administration Worsens Cecal Ligation and Puncture-Induced Sepsis in Obese Mice Through Gut Dysbiosis Enhanced Systemic Inflammation, Impact of Pathogen-Associated Molecules from Gut Translocation and Saturated Fatty Acid. Front. Immunol. 2020, 11, 2278. [Google Scholar] [CrossRef]
- Tungsanga, S.; Panpetch, W.; Bhunyakarnjanarat, T.; Udompornpitak, K.; Katavetin, P.; Chancharoenthana, W.; Chattanathon, P.; Somboona, N.; Tungsaga, K.; Tumwasorn, S.; et al. Uremia-Induced Gut Barrier Defect In 5/6 Nephrectomized Mice Is Worsened by Candida Administration Through a Synergy of Uremic Toxin, Lipopolysaccharide, and (1➔3)-Β-D-Glucan but Is Attenuated by Lacticaseibacillus Rhamnosus L34. Int. J. Mol. Sci. 2022, 23, 2511. [Google Scholar] [CrossRef]
- Panpetch, W.; Kullapanich, C.; Dang, C.P.; Visitchanakun, P.; Saisorn, W.; Wongphoom, J.; Wannigama, D.L.; Thim-uam, A.; Patarakul, K.; Somboonna, N.; et al. Candida Administration Worsens Uremia-Induced Gut Leakage in Bilateral Nephrectomy Mice, An Impact of Gut Fungi and Organismal Molecules in Uremia. Msystems 2021, 6, e01187-20. [Google Scholar] [CrossRef]
- Visitchanakun, P.; Panpetch, W.; Saisorn, W.; Chatthanathon, P.; Wannigama, D.L.; Thim-Uam, A.; Svasti, S.; Fucharoen, S.; Somboonna, N.; Leelahavanichkul, A. Increased Susceptibility to Dextran Sulfate-Induced Mucositis of Iron-Overload Β-Thalassemia Mice, Another Endogenous Cause of Septicemia in Thalassemia. Clin. Sci. 2021, 135, 1467–1486. [Google Scholar] [CrossRef]
- Charoensappakit, A.; Sae-Khow, K.; Leelahavanichkul, A. Gut Barrier Damage and Gut Translocation of Pathogen Molecules in Lupus, an Impact of Innate Immunity (Macrophages and Neutrophils) in Autoimmune Disease. Int. J. Mol. Sci. 2022, 23, 8223. [Google Scholar] [CrossRef]
- Issara-Amphorn, J.; Somboonna, N.; Pisitkun, P.; Hirankarn, N.; Leelahavanichkul, A. Syk inhibitor Attenuates Inflammation in Lupus Mice from Fcgriib Deficiency but Not in Pristane Induction: The Influence of Lupus Pathogenesis on the Therapeutic Effect. Lupus 2020, 29, 1248–1262. [Google Scholar] [CrossRef]
- Issara-Amphorn, J.; Surawut, S.; Worasilchai, N.; Thim-Uam, A.; Finkelman, M.; Chindamporn, A.; Palaga, T.; Hirankarn, N.; Pisitkun, P.; Leelahavanichkul, A. The Synergy of Endotoxin and (1→3)-β-D-Glucan, from Gut Translocation, Worsens Sepsis Severity in a Lupus Model of Fc Gamma Receptor IIb-Deficient Mice. J. Innate Immun. 2018, 10, 189–201. [Google Scholar] [CrossRef]
- Bhunyakarnjanarat, T.; Udompornpitak, K.; Saisorn, W.; Chantraprapawat, B.; Visitchanakun, P.; Dang, C.; Issara-Amphorn, J.; Leelahavanichkul, A. Prominent Indomethacin-Induced Enteropathy in Fcgriib Defi-cient lupus Mice: An Impact of Macrophage Responses and Immune Deposition in Gut. Int. J. Mol. Sci. 2021, 22, 1377. [Google Scholar] [CrossRef] [PubMed]
- Thim-Uam, A.; Makjaroen, J.; Issara-Amphorn, J.; Saisorn, W.; Wannigama, D.L.; Chancharoenthana, W.; Leelahavanichkul, A. Enhanced Bacteremia in Dextran Sulfate-Induced Colitis in Splenectomy Mice Correlates with Gut Dysbiosis and LPS Tolerance. Int. J. Mol. Sci. 2022, 23, 1676. [Google Scholar] [CrossRef] [PubMed]
- McClure, E.E.; Chávez, A.S.O.; Shaw, D.K.; Carlyon, J.A.; Ganta, R.R.; Noh, S.M.; Wood, D.O.; Bavoil, P.M.; Brayton, K.A.; Martinez, J.J.; et al. Engineering of Obligate Intracellular Bacteria: Progress, Challenges and Paradigms. Nat. Rev. Genet. 2017, 15, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, 1356540. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, R.; Tschismarov, R.; Meissner, F.; Prabakaran, T.; Krissanaprasit, A.; Farahani, E.; Zhang, B.-C.; Assil, S.; Martin, A.; Bertrams, W.; et al. Intracellular Bacteria Engage A STING–TBK1–MVB12b Pathway to Enable Paracrine Cgas–STING Signalling. Nat. Microbiol. 2019, 4, 701–713. [Google Scholar] [CrossRef]
- Graf, J. The Family Rikenellaceae. In The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 857–859. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, J.; Ou, X.; Han, Y. Anti-cancer Substances and Safety of Lactic Acid Bacteria in Clinical Treatment. Front. Microbiol. 2021, 12, 722052. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, P.; Du, H.; Chu, W.; Sun, R.; Qin, S.; Tian, Y.; Zhang, Z.; Xu, F. Akkermansia Muciniphila Potentiates the Antitumor Efficacy of FOLFOX in Colon Cancer. Front. Pharmacol. 2021, 12, 725583. [Google Scholar] [CrossRef]
- Surcel, M.; Constantin, C.; Caruntu, C.; Zurac, S.; Neagu, M. Inflammatory Cytokine Pattern Is Sex-Dependent in Mouse Cutaneous Melanoma Experimental Model. J. Immunol. Res. 2017, 2017, 9212134. [Google Scholar] [CrossRef]
- Mazzone, M.; Menga, A.; Castegna, A. Metabolism and TAM functions-it takes two to tango. FEBS J. 2017, 285, 700–716. [Google Scholar] [CrossRef] [Green Version]
- Lubitz, G.S.; Brody, J.D. Not just neighbours: Positive feedback between tumour-associated macrophages and exhausted T cells. Nat. Rev. Immunol. 2021, 22, 3. [Google Scholar] [CrossRef]
- Shalapour, S.; Karin, M. Pas de Deux: Control of Anti-tumor Immunity by Cancer-Associated Inflammation. Immunity 2019, 51, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.T.H.; Tummamunkong, P.; Padungros, P.; Ponpakdee, P.; Boonprakong, L.; Saisorn, W.; Leelahavanichkul, A.; Kueanjinda, P.; Ritprajak, P. Interaction Between Dendritic Cells and Candida krusei β-Glucan Partially Depends on Dectin-1 and It Promotes High IL-10 Production by T Cells. Front. Cell. Infect. Microbiol. 2021, 10, 566661. [Google Scholar] [CrossRef] [PubMed]
- Hiengrach, P.; Visitchanakun, P.; Finkelman, M.A.; Chancharoenthana, W.; Leelahavanichkul, A. More Prominent Inflammatory Response to Pachyman than to Whole-Glucan Particle and Oat-β-Glucans in Dextran Sulfate-Induced Mucositis Mice and Mouse Injection through Proinflammatory Macrophages. Int. J. Mol. Sci. 2022, 23, 4026. [Google Scholar] [CrossRef] [PubMed]
- Rogers, H.; Williams, D.W.; Feng, G.-J.; Lewis, M.A.O.; Wei, X.-Q. Role of Bacterial Lipopolysaccharide in Enhancing Host Immune Response to Candida albicans. Clin. Dev. Immunol. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Walachowski, S.; Tabouret, G.; Foucras, G. Triggering Dectin-1-Pathway Alone Is Not Sufficient to Induce Cytokine Production by Murine Macrophages. PLoS ONE 2016, 11, e0148464. [Google Scholar] [CrossRef]
- Willment, J.A.; Lin, H.H.; Reid, D.M.; Taylor, P.R.; Williams, D.L.; Wong, S.Y.C.; Gordon, S.; Brown, G.D. Dectin-1 Expression and Function Are Enhanced on Alternatively Activated And GM-CSF-Treated Macrophages and Are Negatively Regulated by IL-10, Dexamethasone, and Lipopolysaccharide. J. Immunol. 2003, 171, 4569–4573. [Google Scholar] [CrossRef]
- Shah, V.B.; Williams, D.L.; Keshvara, L. β-Glucan Attenuates TLR2-And TLR4-Mediated Cytokine Production by Microglia. Neurosci. Lett. 2009, 458, 111–115. [Google Scholar] [CrossRef]
- Daley, D.; Mani, V.R.; Mohan, N.; Akkad, N.; Ochi, A.; Heindel, D.W.; Lee, K.B.; Zambirinis, C.P.; Pandian, G.S.B.; Savadkar, S.; et al. Dectin 1 Activation on Macrophages by Galectin 9 Promotes Pancreatic Carcinoma and Peritumoral Immune Tolerance. Nat. Med. 2017, 23, 556–567. [Google Scholar] [CrossRef]
- Chiba, S.; Ikushima, H.; Ueki, H.; Yanai, H.; Kimura, Y.; Hangai, S.; Nishio, J.; Negishi, H.; Tamura, T.; Saijo, S.; et al. Recognition of Tumor Cells by Dectin-1 Orchestrates Innate Immune Cells for Anti-Tumor Responses. Elife 2014, 3, e04177. [Google Scholar] [CrossRef]
- Chen, F.; Chen, J.; Yang, L.; Liu, J.; Zhang, X.; Zhang, Y.; Tu, Q.; Yin, D.; Lin, D.; Wong, P.P.; et al. Extracellular vesicle-packaged HIF-1α-stabilizing lncRNA from tumour-associated macrophages regulates aerobic glycolysis of breast cancer cells. Nat. Cell Biol. 2019, 21, 498–510. [Google Scholar] [CrossRef]
- Mojsilovic, S.S.; Mojsilovic, S.; Villar, V.H.; Santibanez, J.F. The Metabolic Features of Tumor-Associated Macrophages: Opportunities for Immunotherapy? Anal. Cell. Pathol. 2021, 2021, 5523055. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Jeong, H.; Bae, Y.; Shin, K.; Kang, S.; Kim, H.; Oh, J.; Bae, H. Targeting of M2-Like Tumor-Associated Macrophages with a Melittin-Based Pro-Apoptotic Peptide. J. Immunother. Cancer 2019, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Guo, Y.; Li, B.; Li, X.; Wang, Y.; Han, S.; Cheng, D.; Shuai, X. M2-Like Tumor-Associated Macrophage-Targeted Codelivery of STAT6 Inhibitor and IKKβ siRNA Induces M2-to-M1 Repolarization for Cancer Immunotherapy with Low Immune Side Effects. ACS Central Sci. 2020, 6, 1208–1222. [Google Scholar] [CrossRef]
- Hansen, M.E.; Simmons, K.J.; Tippetts, T.S.; Thatcher, M.O.; Saito, R.R.; Hubbard, S.T.; Trumbull, A.M.; Parker, B.A.; Taylor, O.J.; Bikman, B.T. Lipopolysaccharide Disrupts Mitochondrial Physiology in Skeletal Muscle via Disparate Effects on Sphingolipid Metabolism. Shock 2015, 44, 585–592. [Google Scholar] [CrossRef]
- Kuwabara, T.; Imajoh-Ohmi, S. LPS-Induced Apoptosis Is Dependent Upon Mitochondrial Dysfunction. Apoptosis 2004, 9, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.-S.; Kroemer, G.; Galluzzi, L. Mitochondrial Metabolism and Cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef]
- Klein, K.; He, K.; Younes, A.I.; Barsoumian, H.B.; Chen, D.; Ozgen, T.; Mosaffa, S.; Patel, R.R.; Gu, M.; Novaes, J.; et al. Role of Mitochondria in Cancer Immune Evasion and Potential Therapeutic Approaches. Front. Immunol. 2020, 11, 573326. [Google Scholar] [CrossRef]
- Bhat, T.A.; Kumar, S.; Chaudhary, A.K.; Yadav, N.; Chandra, D. Restoration of Mitochondria Function as a Target for Cancer Therapy. Drug Discov. Today 2015, 20, 635–643. [Google Scholar] [CrossRef]
- Visitchanakun, P.; Kaewduangduen, W.; Chareonsappakit, A.; Susantitaphong, P.; Pisitkun, P.; Ritprajak, P.; Townamchai, N.; Leelahavanichkul, A. Interference on Cytosolic DNA Activation Attenuates Sepsis Severity: Experiments on Cyclic GMP–AMP Synthase (cGAS) Deficient Mice. Int. J. Mol. Sci. 2021, 22, 11450. [Google Scholar] [CrossRef]
- Leelahavanichkul, K.; Amornphimoltham, P.; Molinolo, A.A.; Basile, J.R.; Koontongkaew, S.; Gutkind, J.S. A role for p38 MAPK in head and neck cancer cell growth and tumor-induced angiogenesis and lymphangiogenesis. Mol. Oncol. 2013, 8, 105–118. [Google Scholar] [CrossRef]
- Pengkumsri, N.; Sivamaruthi, B.S.; Sirilun, S.; Peerajan, S.; Kesika, P.; Chaiyasut, K.; Chaiyasut, C. Extraction of β-glucan from Saccharomyces cerevisiae: Comparison of different extraction methods and in vivo assessment of immunomodulatory effect in mice. Food Sci. Technol. 2016, 37, 124–130. [Google Scholar] [CrossRef]
- Chancharoenthana, W.; Kamolratanakul, S.; Ariyanon, W.; Thanachartwet, V.; Phumratanaprapin, W.; Wilairatana, P.; Leelahavanichkul, A. Abnormal Blood Bacteriome, Gut Dysbiosis, and Progression to Severe Dengue Disease. Front. Cell. Infect. Microbiol. 2022, 12, 890817. [Google Scholar] [CrossRef] [PubMed]
- Panpetch, W.; Visitchanakun, P.; Saisorn, W.; Sawatpanich, A.; Chatthanathon, P.; Somboonna, N.; Tumwasorn, S.; Leelahavanichkul, A. Lactobacillus rhamnosus attenuates Thai chili extracts induced gut inflammation and dysbiosis despite capsaicin bactericidal effect against the probiotics, a possible toxicity of high dose capsaicin. PLoS ONE 2021, 16, e0261189. [Google Scholar] [CrossRef]
- Panpetch, W.; Phuengmaung, P.; Hiengrach, P.; Issara-Amphorn, J.; Cheibchalard, T.; Somboonna, N.; Tumwasorn, S.; Leelahavanichkul, A. Candida Worsens Klebsiella pneumoniae Induced-Sepsis in a Mouse Model with Low Dose Dextran Sulfate Solution through Gut Dysbiosis and Enhanced Inflammation. Int. J. Mol. Sci. 2022, 23, 7050. [Google Scholar] [CrossRef]
- Hiengrach, P.; Panpetch, W.; Chindamporn, A.; Leelahavanichkul, A. Macrophage depletion alters bacterial gut microbiota partly through fungal overgrowth in feces that worsens cecal ligation and puncture sepsis mice. Sci. Rep. 2022, 12, 1–17. [Google Scholar] [CrossRef]
- Tungsanga, S.; Katavetin, P.; Panpetch, W.; Udompornpitak, K.; Saisorn, W.; Praditpornsilpa, K.; Eiam-Ong, S.; Tungsanga, K.; Tumwasorn, S.; Leelahavanichkul, A. Lactobacillus Rhamnosus L34 Attenuates Chronic Kidney Disease Progression In 5/6 Nephrectomy Mouse Model Through the Excretion of Anti-Inflammatory Molecules. Nephrol. Dial. Transplant. 2022, 37, 1429–1442. [Google Scholar] [CrossRef] [PubMed]
- Kaewduangduen, W.; Visitchanakun, P.; Saisorn, W.; Phawadee, A.; Manonitnantawat, C.; Chutimaskul, C.; Susantitaphong, P.; Ritprajak, P.; Somboonna, N.; Cheibchalard, T.; et al. Blood Bacteria-Free DNA in Septic Mice Enhances LPS-Induced Inflammation in Mice through Macrophage Response. Int. J. Mol. Sci. 2022, 23, 1907. [Google Scholar] [CrossRef]
- Dang, C.P.; Leelahavanichkul, A. Over-expression of miR-223 induces M2 macrophage through glycolysis alteration and attenuates LPS-induced sepsis mouse model, the cell-based therapy in sepsis. PLoS ONE 2020, 15, e0236038. [Google Scholar] [CrossRef]
- Benner, B.; Scarberry, L.; Suarez-Kelly, L.P.; Duggan, M.C.; Campbell, A.R.; Smith, E.; Lapurga, G.; Jiang, K.; Butchar, J.P.; Tridandapani, S.; et al. Generation of monocyte-derived tumor-associated macrophages using tumor-conditioned media provides a novel method to study tumor-associated macrophages in vitro. J. Immunother. Cancer 2019, 7, 140. [Google Scholar] [CrossRef] [Green Version]
- Makjaroen, J.; Thim-Uam, A.; Dang, C.P.; Pisitkun, T.; Somparn, P.; Leelahavanichkul, A. A Comparison Between 1 Day versus 7 Days of Sepsis in Mice with the Experiments on LPS-Activated Macrophages Support the Use of Intravenous Immunoglobulin for Sepsis Attenuation. J. Inflamm. Res. 2021, ume 14, 7243–7263. [Google Scholar] [CrossRef]
- Dang, C.P.; Issara-Amphorn, J.; Charoensappakit, A.; Udompornpitak, K.; Bhunyakarnjanarat, T.; Saisorn, W.; Sae-Khow, K.; Leelahavanichkul, A. BAM15, a Mitochondrial Uncoupling Agent, Attenuates Inflammation in the LPS Injection Mouse Model: An Adjunctive Anti-Inflammation on Macrophages and Hepatocytes. J. Innate Immun. 2021, 13, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Hiengrach, P.; Visitchanakun, P.; Tongchairawewat, P.; Tangsirisatian, P.; Jungteerapanich, T.; Ritprajak, P.; Wannigama, D.L.; Tangtanatakul, P.; Leelahavanichkul, A. Sepsis Encephalopathy Is Partly Mediated by miR370-3p-Induced Mitochondrial Injury but Attenuated by BAM15 in Cecal Ligation and Puncture Sepsis Male Mice. Int. J. Mol. Sci. 2022, 23, 5445. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward | Reverse |
|---|---|---|
| Tumor necrosis factor-alpha (TNF-α) | 5′ -CCTCACACTCAGATCATCTTCTC- 3′ | 5′ -AGATCCATGCCGTTGGCCAG- 3′ |
| Interleukin-6 (IL-6) | 5′ -TACCACTTCACAAGTCGGAGGc- 3′ | 5′ -CTGCAAGTGCATCATCGTTGTTC- 3′ |
| Interleukin-10 (IL-10) | 5′ -GCTCTTACTGACTGGCATGAG- 3′ | 5′ -CGCAGCTCTAGGAGCATGTG- 3′ |
| Inducible nitric oxide synthase (iNOS) | 5′ -ACCCACATCTGGCAGAATGAG- 3′ | 5′ -AGCCATGACCTTTCGCATTAG- 3′ |
| Interleukin-1ß (IL-1ß) | 5′ -GAAATGCCACCTTTTGACAGTG- 3′ | 5′ -TGGATGCTCTCATCAGGACAG- 3′ |
| Arginase-1 (Arg-1) | 5′ -CTTGGCTTGCTTCGGAACTC- 3′ | 5′ -GGAGAAGGCGTTTGCTTAGTTC- 3′ |
| Transforming Growth Factor-β (TGF-β) | 5′ -CAGAGCTGCGCTTGCAGAG- 3′ | 5′ -GTCAGCAGCCGGTTACCAAG- 3′ |
| Resistin-like molecule-α (FIZZ-1) | 5′ -GCCAGGTCCTGGAACCTTTC- 3′ | 5′ -GGAGCAGGGAGATGCAGATGA- 3′ |
| Nuclear factor-κB (NF-κB) | 5′ -CTTCCTCAGCCATGGTACCTCT- 3′ | 5′ -CAAGTCTTCATCAGCATCAAACTG- 3′ |
| Toll like receptor-2 (TLR-2) | 5′ -ACAGCAAGGTCTTCCTGGTTCC- 3′ | 5′ -GCTCCCTTACAGGCTGAGTTCT- 3′ |
| Toll like receptor-4 (TLR-4) | 5′ -GGCAGCAGGTGGAATTGTAT- 3′ | 5′ -AGGCCCCAGAGTTTTGTTCT- 3′ |
| Dectin-1 | 5′ -TCCCGCAATCAGAGTGAAG- 3′ | 5′ -GTGCAGTAAGCTTTCCTGGG- 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binmama, S.; Dang, C.P.; Visitchanakun, P.; Hiengrach, P.; Somboonna, N.; Cheibchalard, T.; Pisitkun, P.; Chindamporn, A.; Leelahavanichkul, A. Beta-Glucan from S. cerevisiae Protected AOM-Induced Colon Cancer in cGAS-Deficient Mice Partly through Dectin-1-Manipulated Macrophage Cell Energy. Int. J. Mol. Sci. 2022, 23, 10951. https://doi.org/10.3390/ijms231810951
Binmama S, Dang CP, Visitchanakun P, Hiengrach P, Somboonna N, Cheibchalard T, Pisitkun P, Chindamporn A, Leelahavanichkul A. Beta-Glucan from S. cerevisiae Protected AOM-Induced Colon Cancer in cGAS-Deficient Mice Partly through Dectin-1-Manipulated Macrophage Cell Energy. International Journal of Molecular Sciences. 2022; 23(18):10951. https://doi.org/10.3390/ijms231810951
Chicago/Turabian StyleBinmama, Sulaiman, Cong Phi Dang, Peerapat Visitchanakun, Pratsanee Hiengrach, Naraporn Somboonna, Thanya Cheibchalard, Prapaporn Pisitkun, Ariya Chindamporn, and Asada Leelahavanichkul. 2022. "Beta-Glucan from S. cerevisiae Protected AOM-Induced Colon Cancer in cGAS-Deficient Mice Partly through Dectin-1-Manipulated Macrophage Cell Energy" International Journal of Molecular Sciences 23, no. 18: 10951. https://doi.org/10.3390/ijms231810951
APA StyleBinmama, S., Dang, C. P., Visitchanakun, P., Hiengrach, P., Somboonna, N., Cheibchalard, T., Pisitkun, P., Chindamporn, A., & Leelahavanichkul, A. (2022). Beta-Glucan from S. cerevisiae Protected AOM-Induced Colon Cancer in cGAS-Deficient Mice Partly through Dectin-1-Manipulated Macrophage Cell Energy. International Journal of Molecular Sciences, 23(18), 10951. https://doi.org/10.3390/ijms231810951