
Drimane Sesquiterpene Aldehydes Control Candida Yeast Isolated from Candidemia in Chilean Patients
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Identification of Yeast Samples by MALDI-TOF-MS
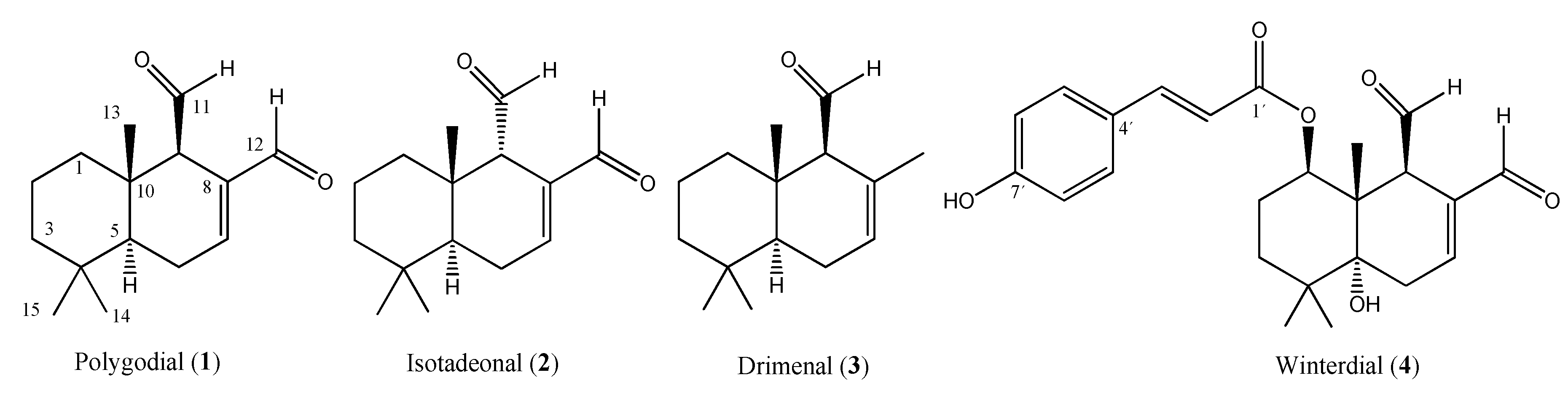
2.2. Secondary Metabolites Isolated from Drimys winteri
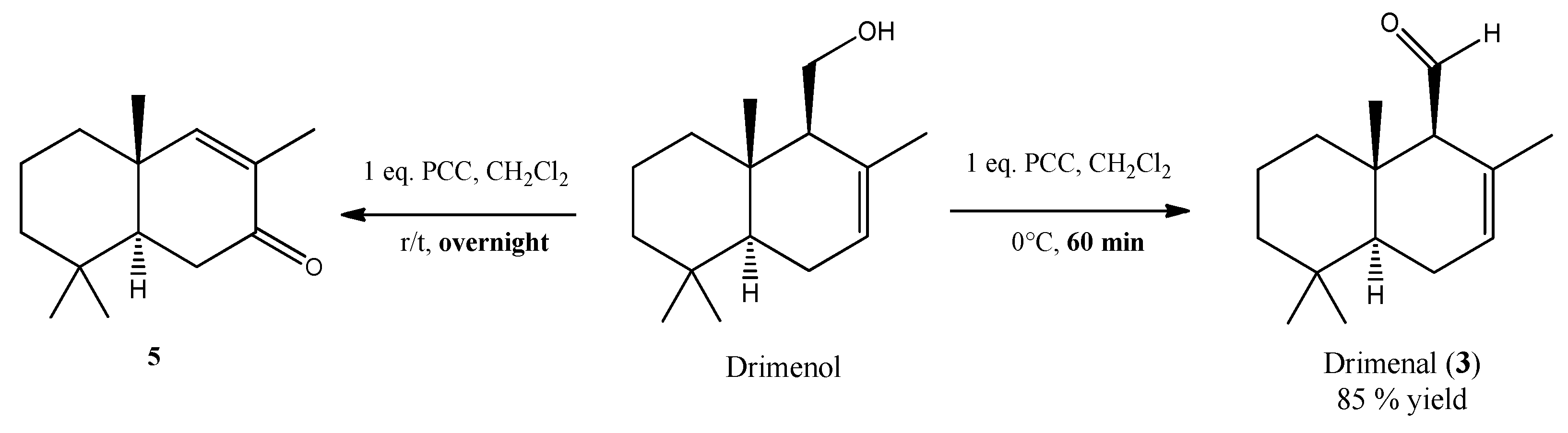
2.3. Oxidation of Drimenol with Pyridinium Chlorochromate
2.4. Anti-Candida Activity
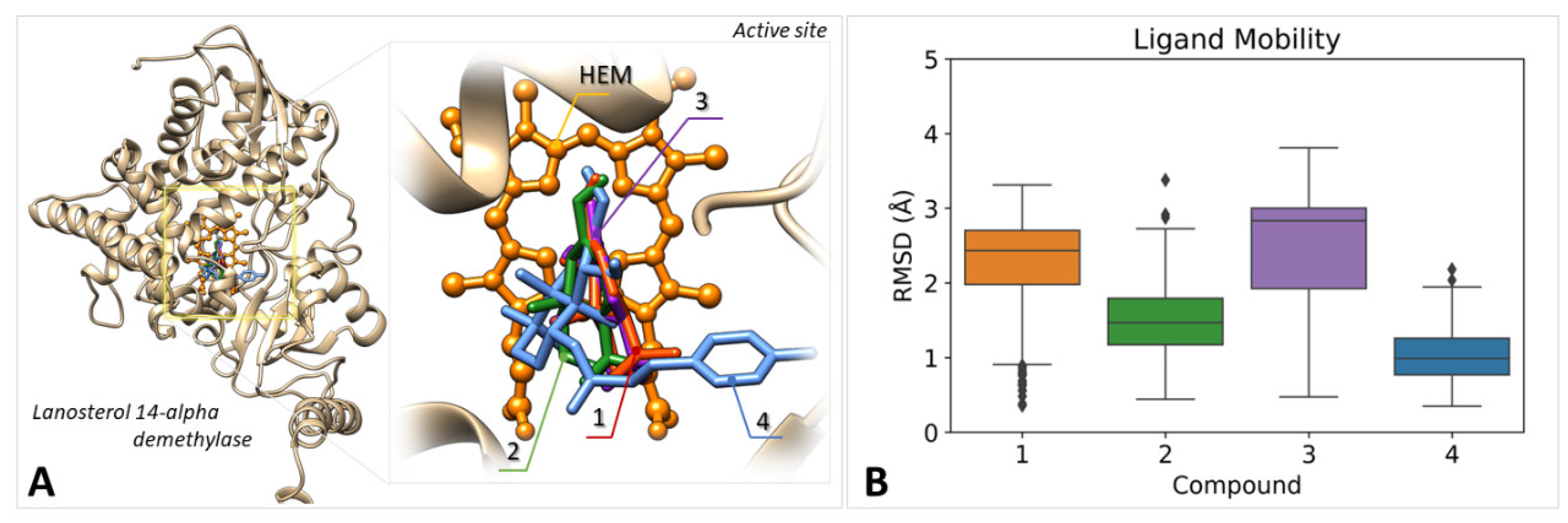
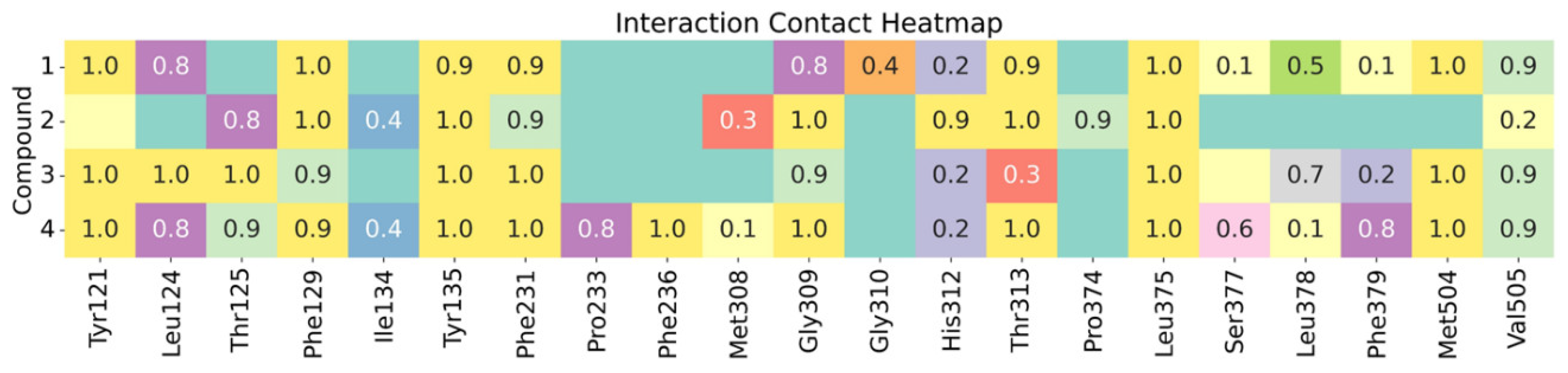
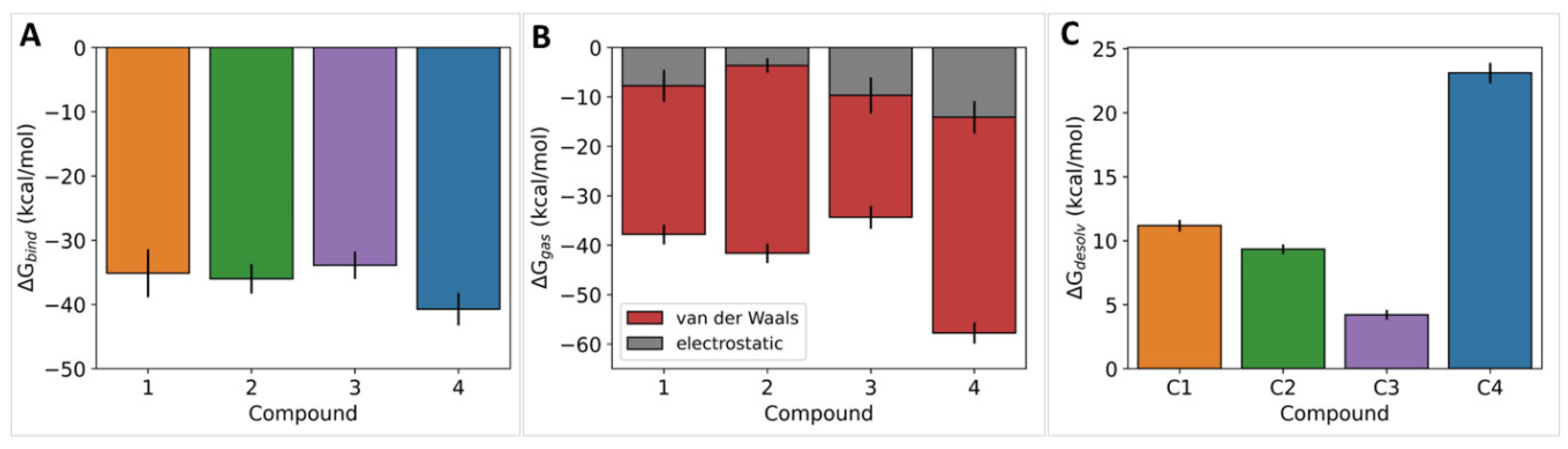
2.5. In Silico Studies
3. Discussion
4. Materials and Methods
4.1. Pathogens
4.2. Proteome Fingerprinting of Candida Yeast
4.3. Vegetal Material
4.4. General Information
4.5. Purification of Drimane Sesquiterpenoids
4.6. Oxidation of Drimenol with Pyridinium Chlorochromate
4.7. NMR Analysis of Drimane Sesquiterpenoids
4.8. Antifungal Assay against Candida Species
4.9. Molecular Simulation
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- McCarty, T.P.; White, C.M.; Pappas, P.G. Candidemia and Invasive Candidiasis. Infect. Dis. Clin. N. Am. 2021, 35, 389–413. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Hermosilla, G.; Abarca, C. Nosocomial candiduria in women undergoing urinary catheterization. Clonal relationship betweenstrains isolated from vaginal tract and urine. Med. Mycol. 2007, 45, 645–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucci, M.; Queiroz-Telles, F.; Alvarado-Matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J.; et al. Epidemiology of Candidemia in Latin America: A Laboratory-Based Survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alburquenque, O.C.; Silva, A.V.; Fuentes, G.M.; Tapia, C.C.; Silva, V.V. Susceptibilidad in vitro a anidulafungina en 100 cepas de especies de Candida aisladas previo a la introducción de esta equinocandina en Chile. J. Rev. Chil. Infectol. 2011, 28, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Krcmery, V.; Kalavsky, E. Antifungal drug discovery, six new molecules patented after 10 years of feast: Why do we need new patented drugs apart from new strategies? Recent Pat. Anti-Infect. Drug Discov 2007, 2, 182–187. [Google Scholar] [CrossRef]
- Krishnasamy, L.; Krishnakumar, S.; Kumaramanickavel, G.; Saikumar, C. Molecular Mechanisms of Antifungal Drug Resistance in Candida Species. J. Clin. Diagn. Res. 2018, 12, 1–6. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Muraszko, J.; Derkacz, D.; Łukaszewicz, M.; Bernat, P.; Krasowska, A. The Role of Ergosterol and Sphingolipids in the Localization and Activity of Candida albicans’ Multidrug Transporter Cdr1p and Plasma Membrane ATPase Pma1p. Int. J. Mol. Sci. 2022, 23, 9975. [Google Scholar] [CrossRef]
- Al-Baqsami, Z.F.; Ahmad, S.; Khan, Z. Antifungal drug susceptibility, molecular basis of resistance to echinocandins and molecular epidemiology of fluconazole resistance among clinical Candida glabrata isolates in Kuwait. Sci. Rep. 2020, 10, 6238. [Google Scholar] [CrossRef] [Green Version]
- Bombaça, A.C.S.; Von Dossow, D.; Barbosa, J.M.C.; Paz, C.; Burgos, V.; Menna-Barreto, R.F.S. Trypanocidal Activity of Natural Sesquiterpenoids Involves Mitochondrial Dysfunction, ROS Production and Autophagic Phenotype in Trypanosomacruzi. Molecules 2018, 23, 2800. [Google Scholar] [CrossRef] [Green Version]
- Paz, C.; Burgos, V.; Iturra, A.; Rebolledo, R.; Ortiz, L.; Baggio, R.; Becerra, J.; Cespedes-Acuña, C.L. Assessment of insecticidal responses of extracts and compounds of Drimys winteri, Lobelia tupa, Viola portalesia and Vestia foetida against the granary weevil Sitophilus granarius. Ind. Crop. Prod. 2018, 122, 232–238. [Google Scholar] [CrossRef]
- Cárcamo, G.; Silva, M.; Becerra, J.; Urrutia, H.; Sossa, K.; Paz, C. Inhibition of quorum sensing by drimane lactones from Chilean flora. J. Chil. Chem. Soc. 2014, 59, 2622–2624. [Google Scholar] [CrossRef] [Green Version]
- Paz, C.; Cárcamo, G.; Silva, M.; Becerra, J.; Urrutia, H.; Sossa, K. Drimendiol, A Drimane Sesquiterpene with Quorum Sensing Inhibition Activity. Nat. Prod. Commun. 2013, 8, 1934578X1300800201. [Google Scholar] [CrossRef] [Green Version]
- Arias, H.R.; Feuerbach, D.; Schmidt, B.; Heydenreich, M.; Paz, C.; Ortells, M.O. Drimane Sesquiterpenoids Noncompetitively Inhibit Human α4β2 Nicotinic Acetylcholine Receptors with Higher Potency Compared to Human α3β4 and α7 Subtypes. J. Nat. Prod. 2018, 81, 811–817. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Tanabe, M.; Kayama, Y.; Abe, M.; Kashio, M.; Koizumi, K.; Okumura, Y.; Morimitsu, Y.; Tominaga, M.; Ozawa, Y.; et al. Miogadial and miogatrial with α,β-unsaturated 1,4-dialdehyde moieties—Novel and potent TRPA1 agonists. Life Sci. 2009, 85, 60–69. [Google Scholar] [CrossRef]
- Beltran, L.R.; Dawid, C.; Beltrán, M.; Gisselmann, G.; Degenhardt, K.; Mathie, K.; Hofmann, T.; Hatt, H. The pungent substances piperine, capsaicin, 6-gingerol and polygodial inhibit the human two-pore domain potassium channels TASK-1, TASK-3 and TRESK. Front. Pharmacol. 2013, 4, 141. [Google Scholar] [CrossRef] [Green Version]
- Paz, C.; Ortiz, L.; Deuis, J.R.; Vetter, I. Polygodial, a drimane sesquiterpenoid dialdehyde purified from Drimys winteri, inhibits voltage-gated sodium channels. Nat. Prod. Res. 2022, 1–6. [Google Scholar] [CrossRef]
- Robles-Kelly, C.; Rubio, J.; Thomas, M.; Sedán, C.; Martinez, R.; Olea, A.F.; Carrasco, H.; Taborga, L.; Silva-Moreno, E. Effect of drimenol and synthetic derivatives on growth and germination of Botrytis cinerea: Evaluation of possible mechanism of action. Pestic. Biochem. Physiol. 2017, 141, 50–56. [Google Scholar] [CrossRef]
- Paz, C.; Viscardi, S.; Iturra, A.; Marin, V.; Miranda, F.; Barra, P.J.; Mendez, I.; Duran, P. Antifungal Effects of Drimane Sesquiterpenoids Isolated from Drimys winteri against Gaeumannomyces graminis var. tritici. Appl. Environ. Microbiol. 2020, 86, e01834-20. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.R.; Lunde, C.S.; Kubo, I. In Vitro Antifungal Susceptibilities of Candida albicans and Other Fungal Pathogens to Polygodial, a Sesquiterpene Dialdehyde. Planta Med. 1999, 65, 204–208. [Google Scholar] [CrossRef]
- Fujita, K.-I.; Kubo, I. Multifunctional action of antifungal polygodial against Saccharomyces cerevisiae: Involvement of pyrrole formation on cell surface in antifungal action. Bioorg. Med. Chem. 2005, 13, 6742–6747. [Google Scholar] [CrossRef]
- Kubo, I.; Lee, S.H.; Shimizu, K. Combination Effect of Miconazole with Polygodial against Candida albicans. Open J. Med. Microbiol. 2011, 1, 7–11. [Google Scholar] [CrossRef]
- Kubo, I.; Fujita, K.; Lee, S.H. Antifungal Mechanism of Polygodial. J. Agric. Food Chem. 2001, 49, 1607–1611. [Google Scholar] [CrossRef]
- Kubo, I.; Himejima, M. Anethole, a synergist of polygodial against filamentous microorganisms. J. Agric. Food Chem. 1991, 39, 2290–2292. [Google Scholar] [CrossRef]
- Marin, V.; Iturra, A.; Opazo, A.; Schmidt, B.; Heydenreich, M.; Ortiz, L.; Jiménez, V.A.; Paz, C. Oxidation of Isodrimeninol with PCC Yields Drimane Derivatives with Activity against Candida Yeast by Inhibition of Lanosterol 14-Alpha Demethylase. Biomolecules 2020, 10, 1101. [Google Scholar] [CrossRef]
- Opiyo, S.A. A review of 13C NMR spectra of drimane sesquiterpenes. Trends Phytochem. Res. 2019, 3, 147–180. [Google Scholar]
- Zárraga, M.; Zárraga, A.M.; Rodríguez, B.; Pérez, C.; Paz, C.; Paz, P.; Sanhueza, C.A. Synthesis of a new nitrogenated drimane derivative with antifungal activity. Tetrahedron Lett. 2008, 49, 4775–4776. [Google Scholar] [CrossRef]
- Cuellar, M.A.; Salas, C.O.; Cortés, M.J.; Morello, A.; Maya, J.D.; Preite, M.D. Synthesis and in vitro trypanocide activity of several polycyclic drimane-quinone derivatives. Bioorg. Med. Chem. 2003, 11, 2489–2497. [Google Scholar] [CrossRef]
- Mathie, K.; Lainer, J.; Spreng, S.; Dawid, C.; Andersson, D.A.; Bevan, S.; Hofmann, T. Structure–Pungency Relationships and TRP Channel Activation of Drimane Sesquiterpenes in Tasmanian Pepper (Tasmannia lanceolata). J. Agric. Food Chem. 2017, 65, 5700–5712. [Google Scholar] [CrossRef] [Green Version]
- Bahia, P.K.; Parks, T.A.; Stanford, K.R.; Mitchell, D.A.; Varma, S.; Stevens, S.M., Jr.; Taylor-Clark, T.E. The exceptionally high reactivity of Cys 621 is critical for electrophilic activation of the sensory nerve ion channel TRPA1. J. Gen. Physiol. 2016, 147, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Hinman, A.; Chuang, H.-H.; Bautista, D.M.; Julius, D. TRP channel activation by reversible covalent modification. Proc. Natl. Acad. Sci. USA 2006, 103, 19564–19568. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Sun, L.; Meng, L.; Wang, M.; Xu, J.; Bartlam, M.; Guo, Y. Sesquiterpenes from Carpesium macrocephalum inhibit Candida albicans biofilm formation and dimorphism. Bioorg. Med. Chem. Lett. 2015, 25, 5409–5411. [Google Scholar] [CrossRef] [PubMed]
- Santos, C. The Chilean Network of Microbial Culture Collections: Establishment and Operation. Boletín Micológico 2016, 31, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Gobom, J.; Schuerenberg, M.; Mueller, M.; Theiss, D.; Lehrach, H.; Nordhoff, E. Alpha-cyano-4-hydroxycinnamic acid affinity sample preparation. A protocol for MALDI-MS peptide analysis in proteomics. Anal. Chem. 2001, 73, 434–438. [Google Scholar] [CrossRef]
- Monk, B.C.; Tomasiak, T.M.; Keniya, M.V.; Huschmann, F.U.; Tyndall, J.D.A.; O’Connell, J.D., III; Cannon, R.D.; McDonald, J.G.; Rodriguez, A.; Finer-Moore, J.S.; et al. Architecture of a single membrane spanning cytochrome P450 suggests constraints that orient the catalytic domain relative to a bilayer. Proc. Natl. Acad. Sci. USA 2014, 111, 3865–3870. [Google Scholar] [CrossRef] [Green Version]
- Anandakrishnan, R.; Aguilar, B.; Onufriev, A.V. H++ 3.0: Automating pK prediction and the preparation of biomolecular structures for atomistic molecular modeling and simulations. Nucleic Acids Res. 2012, 40, W537–W541. [Google Scholar] [CrossRef] [Green Version]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.-X.; Cao, Y. CB-Dock2: Improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef]
- Yang, X.; Liu, Y.; Gan, J.; Xiao, Z.-X.; Cao, Y. FitDock: Protein–ligand docking by template fitting. Brief. Bioinform. 2022, 23, bbac087. [Google Scholar] [CrossRef]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.; Brozell, S.R.; Cerutti, D.S.; Cheatham, I.I.I.T.E.; Cruzeiro, V.W.; Darden, T.A.; Duke, R.E.; et al. AMBER 2021; University of California: San Francisco, CA, USA.
- Miller, B.R.; McGee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA.py: An Efficient Program for End-State Free Energy Calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Identification | Log (Score) * |
|---|---|---|
| 8 | Candida albicans | 2.180 |
| 6 | Candida krusei | 2.610 |
| 11 | Candida glabrata | 2.130 |
| 2 | Candida tropicalis | 2.260 |
| 3 | Candida parapsilopsis | 2.200 |
| 7 | Candida lusitaneae | 2.250 |
| Position | Compound | ||||
|---|---|---|---|---|---|
| Drimenol | 1 | 2 | 3 | 4 | |
| 1 | 1.06, td, J = 13.1, 3.7 | 1.39, td, J = 13.2, 2.5 | 1.77, m | 1.28, td, J = 13.3, 4.3 | |
| 1.95, ov | 1.84, dq, J = 13.2, 2.5 | 1.47, ov | 1.67, m | - | |
| 2 | 1.45, ov | 1.52, 2H, m | 1.62, dt, J = 16.0, 2.8 | 1.55, m | 1.74, m |
| 1.56, qt, J = 13.6, 3.1 | 1.47, ov | 1.43, ov | 1.83, m | ||
| 3 | 1.42, ov | 1.49, m | 1.17, m | 1.43, ov | 2.52, m |
| 1.18, ov | 1.24, m | 1.47, ov | 1.22, dd, J = 13.5, 3.5 | 2.93, dt, J = 20.4, 2.9 | |
| 4 | - | - | - | - | - |
| 5 | 1.17, ov | 1.26, m | 1.55, dd, J = 6.8, 4.9 | 1.16, dd, J = 12.1, 4.7 | - |
| 6 | 1.98, ov | 2.51, ddddd, J = 20.3, 5.5, 4.5, 2.1, 0.8 | 2.55, dt, J = 20.6, 4.6 | 2.07, d, J = 17.7 | 1.27, m |
| 1.86, ov | 2.32, dddd, J = 20.4, 12.1, 3.9, 2.4 | 2.20, ddt, J = 20.6, 11.8, 2.2 | 1.96, m | 2.02, ov | |
| 7 | 5.54, m | 7.14, dt, J = 5.0, 2.2 | 7.08, dd, J= 2.6, 2.3 | 5.68, m | 7.01, dd, J = 5.4, 2.7 |
| 8 | - | - | - | - | - |
| 9 | 1.84, ov | 2.83, ddd, J = 6.1, 4.1, 2.1 | 3.25, t, J = 1.91 | 2.59, s | 3.74, s |
| 10 | - | - | - | - | - |
| 11 | 3.85, dd, J = 11.3, 3.4 | 9.53, d, J = 4.4 | |||
| 3.73, dd, J = 11.3, 5.0 | 9.84, d, J = 2.6 | 9.68, d, J = 5.6 | 9.87, d, J = 3.3 | ||
| 12 | 1.78, s | 9.46, s | 9.40, s | 1.63, s | 9.36, s |
| 13 | 0.86, s | 0.92, s | 0.95, s | 1.06, s | 1.17, s |
| 14 | 0.88, s | 0.96, s | 0.92, s | 0.93, s | 1.06, s |
| 15 | 0.85, s | 0.95, s | 0.90, s | 0.89, s | 1.23, s |
| 2’ | 6.29, d, J = 16.2 | ||||
| 3’ | 7.55, ov | ||||
| 5’ | 7.55, ov | ||||
| 6’ | 6.89, d, J = 8.7 | ||||
| Carbon Number | Compound | ||||
|---|---|---|---|---|---|
| Drimenol | 1 | 2 | 3 | 4 | |
| 1 | 40.0 | 39.5 | 37.2 | 40.3 | 77.6 |
| 2 | 18.7 | 18.0 | 18.5 | 18.4 | 25.1 |
| 3 | 42.2 | 41.7 | 42.2 | 41.9 | 32.7 |
| 4 | 33.1 | 33.1 | 33.0 | 33.2 | 38.9 |
| 5 | 50.0 | 48.9 | 44.4 | 49.2 | 78.2 |
| 6 | 23.7 | 25.2 | 25.7 | 23.8 | 35.2 |
| 7 | 124.3 | 154.3 | 153.6 | 125.6 | 150.6 |
| 8 | 133.0 | 138.2 | 137.5 | 127.9 | 141.5 |
| 9 | 57.4 | 60.3 | 58.6 | 67.7 | 56.4 |
| 10 | 36.1 | 36.8 | 37.8 | 37.1 | 47.5 |
| 11 | 61.1 | 202.0 | 202.3 | 206.8 | 201.2 |
| 12 | 22.1 | 193.2 | 192.9 | 21.6 | 193.7 |
| 13 | 33.5 | 33.1 | 21.6 | 15.8 | 13.4 |
| 14 | 22.2 | 22.9 | 22.0 | 22.1 | 27.8 |
| 15 | 15.1 | 15.2 | 32.8 | 33.3 | 25.2 |
| 1’ | 166.6 | ||||
| 2’ | 116.0 | ||||
| 3’ | 131.0 | ||||
| 4’ | 127.3 | ||||
| 5’ | 131.0 | ||||
| 6’ | 116.7 | ||||
| 7’ | 160.7 | ||||
| Compound | MIC/MFC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| Candida Yeast Type | ||||||
| C. albicans | C. krusei | C. glabrata | C. tropicalis | C. parapsilopsis | C. lusitaneae | |
| 1 | 3.75/- | 7.5/- | 15.0/- | 15.0/- | 7.5/- | 7.5/- |
| 2 | 15.0/60 | 3.75/50 | 3.75/60 | 7.5/50 | 15.0/60 | 1.9/50 |
| 3 | 50.0/50 | 25.0/25.0 | 25.0/25 | 50.0/60 | 50.0/60 | 6.25/6.25 |
| 4 | -/- | -/- | -/- | -/- | 50.0/- | -/- |
| Control | 0.8 | 12.5 | 3.13 | 0.8 | 0.8 | 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marín, V.; Bart, B.; Cortez, N.; Jiménez, V.A.; Silva, V.; Leyton, O.; Cabrera-Pardo, J.R.; Schmidt, B.; Heydenreich, M.; Burgos, V.; et al. Drimane Sesquiterpene Aldehydes Control Candida Yeast Isolated from Candidemia in Chilean Patients. Int. J. Mol. Sci. 2022, 23, 11753. https://doi.org/10.3390/ijms231911753
Marín V, Bart B, Cortez N, Jiménez VA, Silva V, Leyton O, Cabrera-Pardo JR, Schmidt B, Heydenreich M, Burgos V, et al. Drimane Sesquiterpene Aldehydes Control Candida Yeast Isolated from Candidemia in Chilean Patients. International Journal of Molecular Sciences. 2022; 23(19):11753. https://doi.org/10.3390/ijms231911753
Chicago/Turabian StyleMarín, Víctor, Bryan Bart, Nicole Cortez, Verónica A. Jiménez, Víctor Silva, Oscar Leyton, Jaime R. Cabrera-Pardo, Bernd Schmidt, Matthias Heydenreich, Viviana Burgos, and et al. 2022. "Drimane Sesquiterpene Aldehydes Control Candida Yeast Isolated from Candidemia in Chilean Patients" International Journal of Molecular Sciences 23, no. 19: 11753. https://doi.org/10.3390/ijms231911753
APA StyleMarín, V., Bart, B., Cortez, N., Jiménez, V. A., Silva, V., Leyton, O., Cabrera-Pardo, J. R., Schmidt, B., Heydenreich, M., Burgos, V., & Paz, C. (2022). Drimane Sesquiterpene Aldehydes Control Candida Yeast Isolated from Candidemia in Chilean Patients. International Journal of Molecular Sciences, 23(19), 11753. https://doi.org/10.3390/ijms231911753