
The TZM-bl Reporter Cell Line Expresses Kynureninase That Can Neutralize 2F5-like Antibodies in the HIV-1 Neutralization Assay


Abstract
:
1. Introduction
2. Results
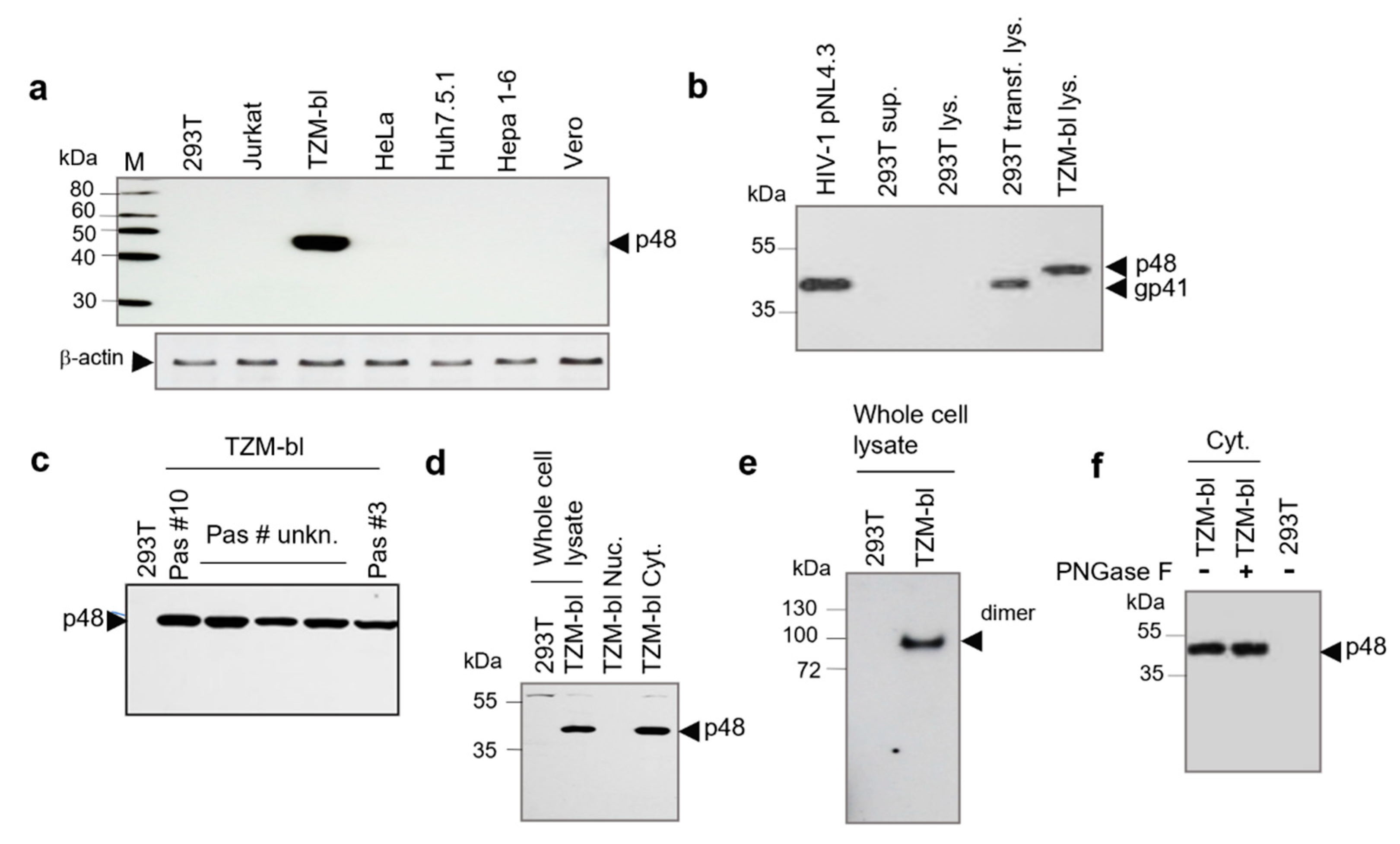
2.1. Detection and Characterization of p48 from TZM-bl Cells
2.2. Analysis of the p48 in Cells by Confocal Microscopy and Isolation of p48 from Cell-Free Supernatant
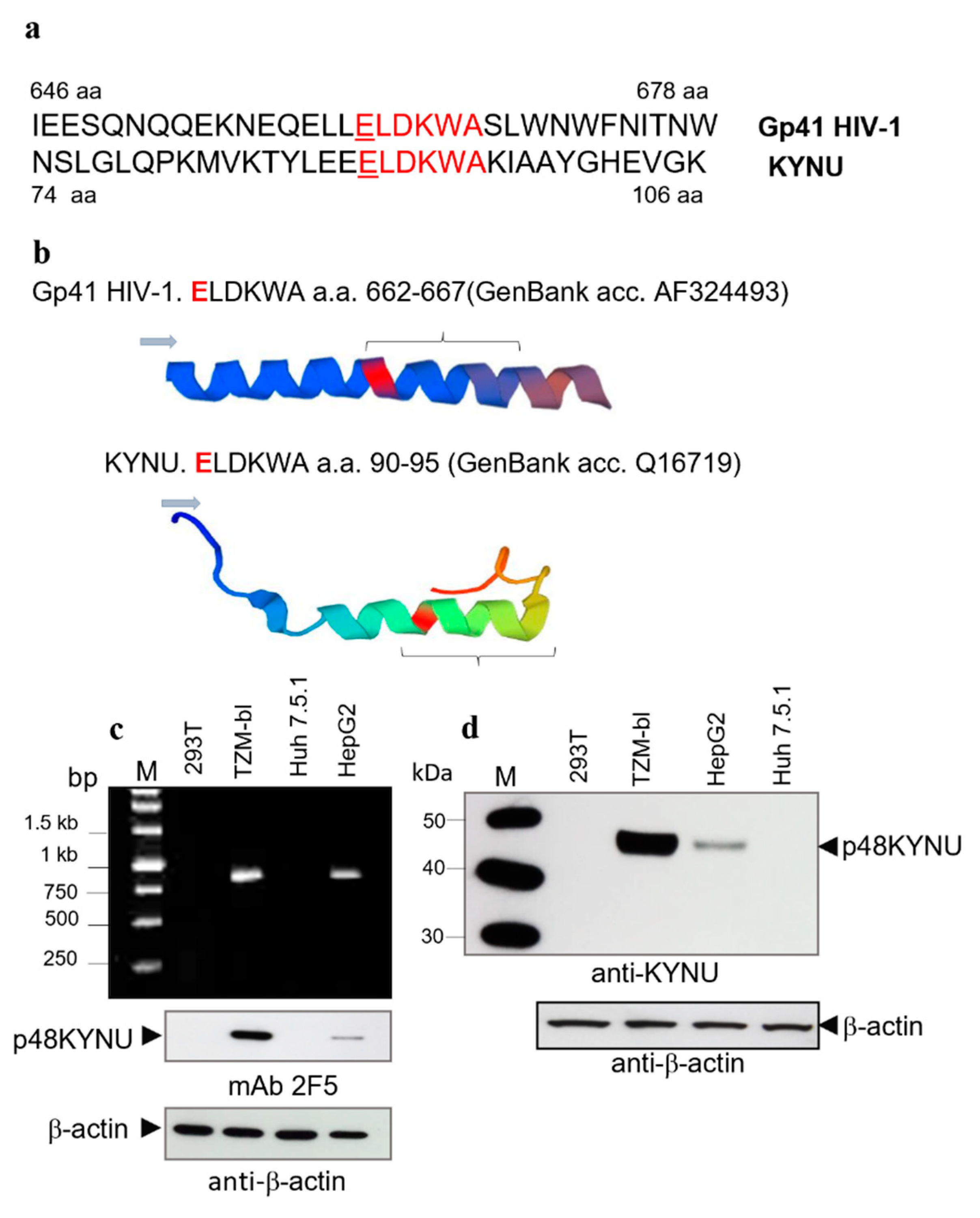
2.3. Identification of p48 as Kynureninase (KYNU)
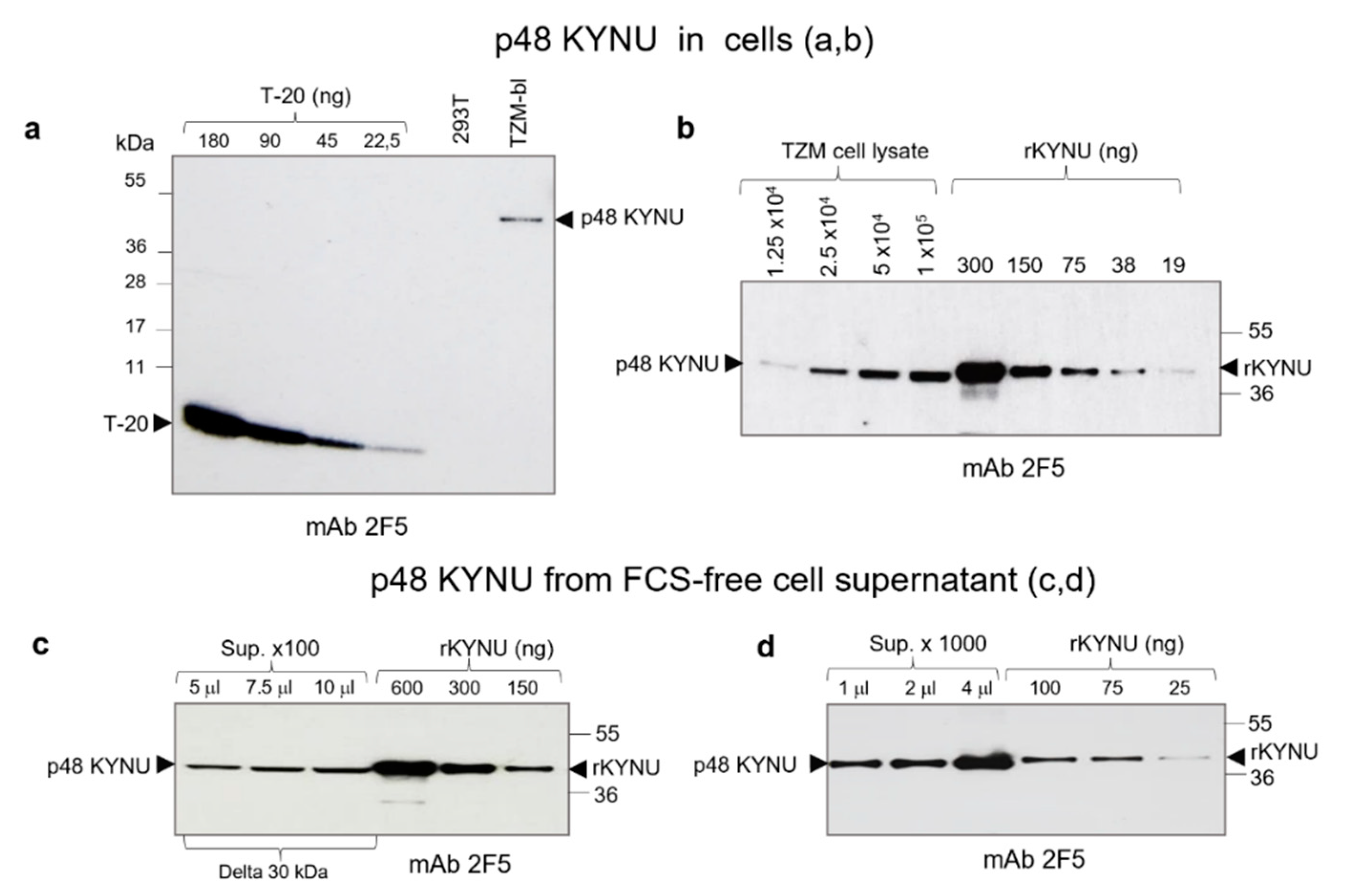
2.4. Estimation of p48 KYNU Amount in TZM-bl Cells and Cell Supernatant
2.5. Evaluation of the p48 KYNU Effect on mAbs 2F5 and 4E10 in NT Assays
3. Discussion
4. Materials and Methods
4.1. Cells and Cellular Lysates
4.2. Monoclonal Antibodies (mAb), Sera, Peptide, and Recombinant Protein
4.3. Protein de-Glycosylation
4.4. SDS-PAGE, Native PAGE, and Western Blot Analyses
4.5. Confocal Microscopy
4.6. Isolation of p48 from Cell-Free Supernatant of TZM-bl Cells
4.7. Extraction of RNA and DNA
4.8. Reverse Transcriptase (RT)-PCR, Cloning, and Duplex Real-Time PCR
4.9. Transfection
4.10. HIV-1pNL4-3 for the Neutralization (NT) Assay
4.11. NT Assay on TZM-bl and Jurkat Cells
4.12. Software
4.13. Statistics
5. Conclusions
- -
- The amount of p48 KYNU in TZM-bl cells is nearly 0.64 pg/cell
- -
- About 0.19 pg/cell of p48 KYNU are released to cell supernatant after 48 h
- -
- From 7.5 ng to 9.35 ng of p48 KYNU can be released after 48 h from cells in one well of 96 well plate at the confluence.
- -
- Released p48 KYNU can interact/inactivate 21–26 ng of 2F5-like antibodies in one well of 96 well plate.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Landry, J.J.; Pyl, P.T.; Rausch, T.; Zichner, T.; Tekkedil, M.M.; Stutz, A.M.; Jauch, A.; Aiyar, R.S.; Pau, G.; Delhomme, N.; et al. The genomic and transcriptomic landscape of a HeLa cell line. G3 Genes Genomes Genet 2013, 3, 1213–1224. [Google Scholar] [CrossRef] [Green Version]
- Platt, E.J.; Wehrly, K.; Kuhmann, S.E.; Chesebro, B.; Kabat, D. Effects of CCR5 and CD4 cell surface concentrations on infections by macrophagetropic isolates of human immunodeficiency virus type 1. J. Virol. 1998, 72, 2855–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappes, J.; Wu, X. Cell-Based Method and Assay for Measuring the Infectivity and Drug Sensitivity of Immuno-Deficiency Virus. U.S. Patent 6,797,4622 B1, 10 February 2005. [Google Scholar]
- Wei, X.; Decker, J.M.; Liu, H.; Zhang, Z.; Arani, R.B.; Kilby, J.M.; Saag, M.S.; Wu, X.; Shaw, G.M.; Kappes, J.C. Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrob. Agents Chemother. 2002, 46, 1896–1905. [Google Scholar] [CrossRef] [Green Version]
- Montefiori, D.C. Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays. Curr. Protoc. Immunol. 2005, 64, 12.11.1–12.11.17. [Google Scholar] [CrossRef]
- Polonis, V.R.; Brown, B.K.; Rosa Borges, A.; Zolla-Pazner, S.; Dimitrov, D.S.; Zhang, M.Y.; Barnett, S.W.; Ruprecht, R.M.; Scarlatti, G.; Fenyo, E.M.; et al. Recent advances in the characterization of HIV-1 neutralization assays for standardized evaluation of the antibody response to infection and vaccination. Virology 2008, 375, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Montefiori, D.C. Measuring HIV neutralization in a luciferase reporter gene assay. Methods Mol. Biol. 2009, 485, 395–405. [Google Scholar] [CrossRef]
- Willey, R.; Nason, M.C.; Nishimura, Y.; Follmann, D.A.; Martin, M.A. Neutralizing antibody titers conferring protection to macaques from a simian/human immunodeficiency virus challenge using the TZM-bl assay. AIDS Res. Hum. Retrovir. 2010, 26, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Morozov, V.A.; Morozov, A.V.; Lagaye, S. Short communication: Simultaneous substitutions of V38M and N43T-N44K in the gp41 heptad repeat 1 (HR1) disrupt HIV type 1 gPr160 endoproteolytic cleavage. AIDS Res. Hum. Retrovir. 2010, 26, 73–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, W.W.; Rao, S.S.; Lim, S.Y.; Zhang, J.; Hraber, P.T.; Brassard, L.M.; Luedemann, C.; Todd, J.P.; Dodson, A.; Shen, L.; et al. The TRIM5 gene modulates penile mucosal acquisition of simian immunodeficiency virus in rhesus monkeys. J. Virol. 2011, 85, 10389–10398. [Google Scholar] [CrossRef] [Green Version]
- Montefiori, D.C.; Karnasuta, C.; Huang, Y.; Ahmed, H.; Gilbert, P.; de Souza, M.S.; McLinden, R.; Tovanabutra, S.; Laurence-Chenine, A.; Sanders-Buell, E.; et al. Magnitude and breadth of the neutralizing antibody response in the RV144 and Vax003 HIV-1 vaccine efficacy trials. J. Infect. Dis. 2012, 206, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Kramer, V.G.; Varsaneux, O.; Oliviera, M.; Colby-Germinario, S.P.; Mesplede, T.; Wainberg, M.A. Exposure to entry inhibitors alters HIV infectiousness and sensitivity to broadly neutralizing monoclonal antibodies. J. Acquir. Immune Defic. Syndr. 2014, 67, 7–14. [Google Scholar] [CrossRef]
- Lai, R.P.; Hock, M.; Radzimanowski, J.; Tonks, P.; Hulsik, D.L.; Effantin, G.; Seilly, D.J.; Dreja, H.; Kliche, A.; Wagner, R.; et al. A fusion intermediate gp41 immunogen elicits neutralizing antibodies to HIV-1. J. Biol. Chem. 2014, 289, 29912–29926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gombos, R.B.; Kolodkin-Gal, D.; Eslamizar, L.; Owuor, J.O.; Mazzola, E.; Gonzalez, A.M.; Korioth-Schmitz, B.; Gelman, R.S.; Montefiori, D.C.; Haynes, B.F.; et al. Inhibitory Effect of Individual or Combinations of Broadly Neutralizing Antibodies and Antiviral Reagents against Cell-Free and Cell-to-Cell HIV-1 Transmission. J. Virol. 2015, 89, 7813–7828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santra, S.; Tomaras, G.D.; Warrier, R.; Nicely, N.I.; Liao, H.X.; Pollara, J.; Liu, P.; Alam, S.M.; Zhang, R.; Cocklin, S.L.; et al. Human Non-neutralizing HIV-1 Envelope Monoclonal Antibodies Limit the Number of Founder Viruses during SHIV Mucosal Infection in Rhesus Macaques. PLoS Pathog. 2015, 11, e1005042. [Google Scholar] [CrossRef]
- Molinos-Albert, L.M.; Carrillo, J.; Curriu, M.; Rodriguez de la Concepcion, M.L.; Marfil, S.; Garcia, E.; Clotet, B.; Blanco, J. Anti-MPER antibodies with heterogeneous neutralization capacity are detectable in most untreated HIV-1 infected individuals. Retrovirology 2014, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gludish, D.W.; Boliar, S.; Caldwell, S.; Tembo, D.L.; Chimbayo, E.T.; Jambo, K.C.; Mwandumba, H.C.; Russell, D.G. TZM-gfp cells: A tractable fluorescent tool for analysis of rare and early HIV-1 infection. Sci Rep. 2020, 10, 19900. [Google Scholar] [CrossRef]
- Rindler, A.E.; Kuster, H.; Neumann, K.; Leemann, C.; Braun, D.L.; Metzner, K.J.; Günthard, H.F. A Novel High Throughput, Parallel Infection Assay for Determining the Replication Capacities of 346 Primary HIV-1 Isolates of the Zurich Primary HIV-1 Infection Study in Primary Cells. Viruses 2021, 13, 404. [Google Scholar] [CrossRef]
- Pısıl, Y.; Shida, H.; Miura, T. A Neutralization Assay Based on Pseudo-Typed Lentivirus with SARS CoV-2 Spike Protein in ACE2-Expressing CRFK Cells. Pathogens 2021, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Tang, X.; Li, L.; Chen, Y.; Zhu, Y.; Chong, H.; He, Y. Generation of HIV-resistant cells with a single-domain antibody: Implications for HIV-1 gene therapy. Cell Mol. Immunol. 2021, 18, 660–674. [Google Scholar] [CrossRef]
- Müller, T.G.; Zila, V.; Peters, K.; Schifferdecker, S.; Stanic, M.; Lucic, B.; Laketa, V.; Lusic, M.; Müller, B.; Kräusslich, H.G. HIV-1 uncoating by release of viral cDNA from capsid-like structures in the nucleus of infected cells. eLife 2021, 10, e64776. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; McClure, M.O.; Pizzato, M. Identification of gammaretroviruses constitutively released from cell lines used for human immunodeficiency virus research. J. Virol. 2008, 82, 12585–12588. [Google Scholar] [CrossRef] [Green Version]
- Platt, E.J.; Bilska, M.; Kozak, S.L.; Kabat, D.; Montefiori, D.C. Evidence that ecotropic murine leukemia virus contamination in TZM-bl cells does not affect the outcome of neutralizing antibody assays with human immunodeficiency virus type 1. J. Virol. 2009, 83, 8289–8292. [Google Scholar] [CrossRef] [Green Version]
- Sarzotti-Kelsoe, M.; Bailer, R.T.; Turk, E.; Lin, C.L.; Bilska, M.; Greene, K.M.; Gao, H.; Todd, C.A.; Ozaki, D.A.; Seaman, M.S.; et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 2014, 409, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Muster, T.; Steindl, F.; Purtscher, M.; Trkola, A.; Klima, A.; Himmler, G.; Ruker, F.; Katinger, H. A conserved neutralizing epitope on gp41 of human immunodeficiency virus type 1. J. Virol. 1993, 67, 6642–6647. [Google Scholar] [CrossRef] [Green Version]
- Zwick, M.B.; Wang, M.; Poignard, P.; Stiegler, G.; Katinger, H.; Burton, D.R.; Parren, P.W. Neutralization synergy of human immunodeficiency virus type 1 primary isolates by cocktails of broadly neutralizing antibodies. J. Virol. 2001, 75, 12198–12208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joos, B.; Trkola, A.; Kuster, H.; Aceto, L.; Fischer, M.; Stiegler, G.; Armbruster, C.; Vcelar, B.; Katinger, H.; Gunthard, H.F. Long-term multiple-dose pharmacokinetics of human monoclonal antibodies (MAbs) against human immunodeficiency virus type 1 envelope gp120 (MAb 2G12) and gp41 (MAbs 4E10 and 2F5). Antimicrob. Agents Chemother. 2006, 50, 1773–1779. [Google Scholar] [CrossRef] [Green Version]
- Hessell, A.J.; Rakasz, E.G.; Tehrani, D.M.; Huber, M.; Weisgrau, K.L.; Landucci, G.; Forthal, D.N.; Koff, W.C.; Poignard, P.; Watkins, D.I.; et al. Broadly neutralizing monoclonal antibodies 2F5 and 4E10 directed against the human immunodeficiency virus type 1 gp41 membrane-proximal external region protect against mucosal challenge by simian-human immunodeficiency virus SHIVBa-L. J. Virol. 2010, 84, 1302–1313. [Google Scholar] [CrossRef] [Green Version]
- Balazs, A.B.; Chen, J.; Hong, C.M.; Rao, D.S.; Yang, L.; Baltimore, D. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 2011, 481, 81–84. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.S. Structure and mechanism of kynureninase. Arch. Biochem. Biophys. 2014, 544, 69–74. [Google Scholar] [CrossRef]
- Walsh, H.A.; Botting, N.P. Purification and biochemical characterization of some of the properties of recombinant human kynureninase. Eur. J. Biochem. 2002, 269, 2069–2074. [Google Scholar] [CrossRef]
- Lima, S.; Khristoforov, R.; Momany, C.; Phillips, R.S. Crystal structure of Homo sapiens kynureninase. Biochemistry 2007, 46, 2735–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Holl, T.M.; Liu, Y.; Li, Y.; Lu, X.; Nicely, N.I.; Kepler, T.B.; Alam, S.M.; Liao, H.X.; Cain, D.W.; et al. Identification of autoantigens recognized by the 2F5 and 4E10 broadly neutralizing HIV-1 antibodies. J. Exp. Med. 2013, 210, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Tanizawa, K.; Soda, K. Purification and properties of pig liver kynureninase. J. Biochem. 1979, 85, 901–906. [Google Scholar] [CrossRef]
- Asp, L.; Johansson, A.S.; Mann, A.; Owe-Larsson, B.; Urbanska, E.M.; Kocki, T.; Kegel, M.; Engberg, G.; Lundkvist, G.B.; Karlsson, H. Effects of pro-inflammatory cytokines on expression of kynurenine pathway enzymes in human dermal fibroblasts. J. Inflamm. 2011, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Harden, J.L.; Lewis, S.M.; Lish, S.R.; Suarez-Farinas, M.; Gareau, D.; Lentini, T.; Johnson-Huang, L.M.; Krueger, J.G.; Lowes, M.A. The tryptophan metabolism enzyme L-kynureninase is a novel inflammatory factor in psoriasis and other inflammatory diseases. J. Allergy Clin. Immunol. 2016, 137, 1830–1840. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mi, Y.; Mueller, T.; Kreibich, S.; Williams, E.; Van Drogen, A.; Borel, C.; Frank, M.; Germain, P.-L.; Bludau, I.; et al. Multi-omic measurements of heterogeneity in HeLa cells across laboratories. Nat. Biotechnol. 2019, 37, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Rubartelli, A.; Cozzolino, F.; Talio, M.; Sitia, R. A novel secretory pathway for interleukin-1 beta, a protein lacking a signal sequence. EMBO J. 1990, 9, 1503–1510. [Google Scholar] [CrossRef]
- MacKenzie, A.; Wilson, H.L.; Kiss-Toth, E.; Dower, S.K.; North, R.A.; Surprenant, A. Rapid secretion of interleukin-1beta by microvesicle shedding. Immunity 2001, 15, 825–835. [Google Scholar] [CrossRef]
- Prudovsky, I.; Mandinova, A.; Soldi, R.; Bagala, C.; Graziani, I.; Landriscina, M.; Tarantini, F.; Duarte, M.; Bellum, S.; Doherty, H.; et al. The non-classical export routes: FGF1 and IL-1alpha point the way. J. Cell Sci. 2003, 116, 4871–4881. [Google Scholar] [CrossRef] [Green Version]
- Burton, D.R.; Desrosiers, R.C.; Doms, R.W.; Koff, W.C.; Kwong, P.D.; Moore, J.P.; Nabel, G.J.; Sodroski, J.; Wilson, I.A.; Wyatt, R.T. HIV vaccine design and the neutralizing antibody problem. Nat. Immunol. 2004, 5, 233–236. [Google Scholar] [CrossRef]
- Gallaher, W.R.; Ball, J.M.; Garry, R.F.; Griffin, M.C.; Montelaro, R.C. A general model for the transmembrane proteins of HIV and other retroviruses. AIDS Res. Hum. Retrovir. 1989, 5, 431–440. [Google Scholar] [CrossRef]
- Earl, P.L.; Broder, C.C.; Doms, R.W.; Moss, B. Epitope map of human immunodeficiency virus type 1 gp41 derived from 47 monoclonal antibodies produced by immunization with oligomeric envelope protein. J. Virol. 1997, 71, 2674–2684. [Google Scholar] [CrossRef] [Green Version]
- Pietzsch, J.; Scheid, J.F.; Mouquet, H.; Klein, F.; Seaman, M.S.; Jankovic, M.; Corti, D.; Lanzavecchia, A.; Nussenzweig, M.C. Human anti-HIV-neutralizing antibodies frequently target a conserved epitope essential for viral fitness. J. Exp. Med. 2010, 207, 1995–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opalka, D.; Pessi, A.; Bianchi, E.; Ciliberto, G.; Schleif, W.; McElhaugh, M.; Danzeisen, R.; Geleziunas, R.; Miller, M.; Eckert, D.M.; et al. Analysis of the HIV-1 gp41 specific immune response using a multiplexed antibody detection assay. J. Immunol. Methods 2004, 287, 49–65. [Google Scholar] [CrossRef]
- Zolla-Pazner, S. Identifying epitopes of HIV-1 that induce protective antibodies. Nat. Rev. Immunol. 2004, 4, 199–210. [Google Scholar] [CrossRef]
- Montero, M.; van Houten, N.E.; Wang, X.; Scott, J.K. The membrane-proximal external region of the human immunodeficiency virus type 1 envelope: Dominant site of antibody neutralization and target for vaccine design. Microbiol. Mol. Biol. Rev. MMBR 2008, 72, 54–84. [Google Scholar] [CrossRef] [Green Version]
- Gray, E.S.; Madiga, M.C.; Moore, P.L.; Mlisana, K.; Abdool Karim, S.S.; Binley, J.M.; Shaw, G.M.; Mascola, J.R.; Morris, L. Broad neutralization of human immunodeficiency virus type 1 mediated by plasma antibodies against the gp41 membrane proximal external region. J. Virol. 2009, 83, 11265–11274. [Google Scholar] [CrossRef] [Green Version]
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute HIV-1 infection: Clues for vaccine development. Nat. Rev. Immunol. 2010, 10, 11–23. [Google Scholar] [CrossRef]
- Lutje Hulsik, D.; Liu, Y.Y.; Strokappe, N.M.; Battella, S.; El Khattabi, M.; McCoy, L.E.; Sabin, C.; Hinz, A.; Hock, M.; Macheboeuf, P.; et al. A gp41 MPER-specific llama VHH requires a hydrophobic CDR3 for neutralization but not for antigen recognition. PLoS Pathog. 2013, 9, e1003202. [Google Scholar] [CrossRef]
- Frey, G.; Peng, H.; Rits-Volloch, S.; Morelli, M.; Cheng, Y.; Chen, B. A fusion-intermediate state of HIV-1 gp41 targeted by broadly neutralizing antibodies. Proc. Natl. Acad. Sci. USA 2008, 105, 3739–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, G.; Chen, J.; Rits-Volloch, S.; Freeman, M.M.; Zolla-Pazner, S.; Chen, B. Distinct conformational states of HIV-1 gp41 are recognized by neutralizing and non-neutralizing antibodies. Nat. Struct. Mol. Biol. 2010, 17, 1486–1491. [Google Scholar] [CrossRef]
- Vassell, R.; He, Y.; Vennakalanti, P.; Dey, A.K.; Zhuang, M.; Wang, W.; Sun, Y.; Biron-Sorek, Z.; Srivastava, I.K.; LaBranche, C.C.; et al. Immunogens Modeling a Fusion-Intermediate Conformation of gp41 Elicit Antibodies to the Membrane Proximal External Region of the HIV Envelope Glycoprotein. PLoS ONE 2015, 10, e0128562. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, Z.; Cheng, X.; Chen, Y.H. The recombinant immunogen with high-density epitopes of ELDKWA and ELDEWA induced antibodies recognizing both epitopes on HIV-1 gp41. Microbiol. Immunol. 2005, 49, 703–709. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, Y. Characterization of Antibody Responses Against the 2F5 Epitope ELDKWA Using HIV-1 Env-Mediated Membrane Fusion and Neutralization Assays. Tsinghua Sci. Technol. 2010, 15, 447–451. [Google Scholar] [CrossRef]
- Strasz, N.; Morozov, V.A.; Kreutzberger, J.; Keller, M.; Eschricht, M.; Denner, J. Immunization with hybrid proteins containing the membrane proximal external region of HIV-1. AIDS Res. Hum. Retrovir. 2014, 30, 498–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holl, V.; Peressin, M.; Decoville, T.; Schmidt, S.; Zolla-Pazner, S.; Aubertin, A.M.; Moog, C. Nonneutralizing antibodies are able to inhibit human immunodeficiency virus type 1 replication in macrophages and immature dendritic cells. J. Virol. 2006, 80, 6177–6181. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Origin | Cell Type/Morphology |
|---|---|---|
| HeLa | NIH AIDS Reagent Program (#153) | Human cervical carcinoma |
| HeLa CD4+LTR-Luc | Robert Koch Institute | Human cervical carcinoma |
| TZM-bl (HeLa-derived) | NIH AIDS Reagent Program (#8129) | Human cervical carcinoma |
| Huh 7.5.1. | Scripps Res. Inst., San Diego, CA, USA (Prof. Frank Chisari) | Human hepatocytes, epithelial-like |
| HepG2 | ATCC, HB8065 | Human hepatocytes |
| Jurkat | NIH AIDS Reagent Program (#177) | Human lymphocytes |
| Hepa 1-6 | ATCC, CRL-1830 | Mouse hepatoma |
| Vero | NIH AIDS Reagent Program (CCL-81) | Grivet monkey, kidney |
| Name | Sequence 5′-3′ | Nt. Position | Acc. Numbers |
|---|---|---|---|
| HIV env F1 (7531) | 5′-CAGTGGACAAATTAGATGTTCATC | 7531–7554 | AF324493 |
| HIV env F2 (7580) | 5′-AGAGATGGTGGTAATAACAAC | 7580–7600 | AF324493 |
| HIV env R1 (8475) | 5′-GGCTCCGCAGATCGTCCCAGATAAGTG | 8501–8475 | AF324493 |
| HIV env R2 (8347) | 5′-TCCCTGCCTAACTCTATTCACT | 8347–8326 | AF324493 |
| HIV gag F | 5′-ATGGGTGCGAGAGCGTCGGTATT | 790–811 | AF324493 |
| HIV gag R | 5′-GGCTTCCTTGGTGTCTTTTACA | 1089–1068 | AF324493 |
| HIV gag (Probe) | FAM 5′-AATCCTGGCCTTTTAGAGACATCAG-BHQ | 928–950 | AF324493 |
| Kynu F | 5′-ATGGAGCCTTCATCTCTTGAG | 131–151 | NM_003937 |
| Kynu R | 5′-TGCTCCTGCATTTAAATA | 976–959 | NM_003937 |
| GAPDH F | 5′-CCACTCCTCCACCTTTGAC-3’ | 1058–1076 | NM_002046.5 |
| GAPDH R | 5’-ACCCTGTTGCTGTAGCCA-3′ | 1159–1142 | NM_002046.5 |
| GAPDH (Probe) | FAM 5′-TTGCCCTCAACGACCACTTTGTC-BHQ | 1090–1112 | NM_002046.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morozov, V.; Lagaye, S.; Morozov, A. The TZM-bl Reporter Cell Line Expresses Kynureninase That Can Neutralize 2F5-like Antibodies in the HIV-1 Neutralization Assay. Int. J. Mol. Sci. 2022, 23, 641. https://doi.org/10.3390/ijms23020641
Morozov V, Lagaye S, Morozov A. The TZM-bl Reporter Cell Line Expresses Kynureninase That Can Neutralize 2F5-like Antibodies in the HIV-1 Neutralization Assay. International Journal of Molecular Sciences. 2022; 23(2):641. https://doi.org/10.3390/ijms23020641
Chicago/Turabian StyleMorozov, Vladimir, Sylvie Lagaye, and Alexey Morozov. 2022. "The TZM-bl Reporter Cell Line Expresses Kynureninase That Can Neutralize 2F5-like Antibodies in the HIV-1 Neutralization Assay" International Journal of Molecular Sciences 23, no. 2: 641. https://doi.org/10.3390/ijms23020641
APA StyleMorozov, V., Lagaye, S., & Morozov, A. (2022). The TZM-bl Reporter Cell Line Expresses Kynureninase That Can Neutralize 2F5-like Antibodies in the HIV-1 Neutralization Assay. International Journal of Molecular Sciences, 23(2), 641. https://doi.org/10.3390/ijms23020641