
Comparison of Carbonic Anhydrases for CO2 Sequestration

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
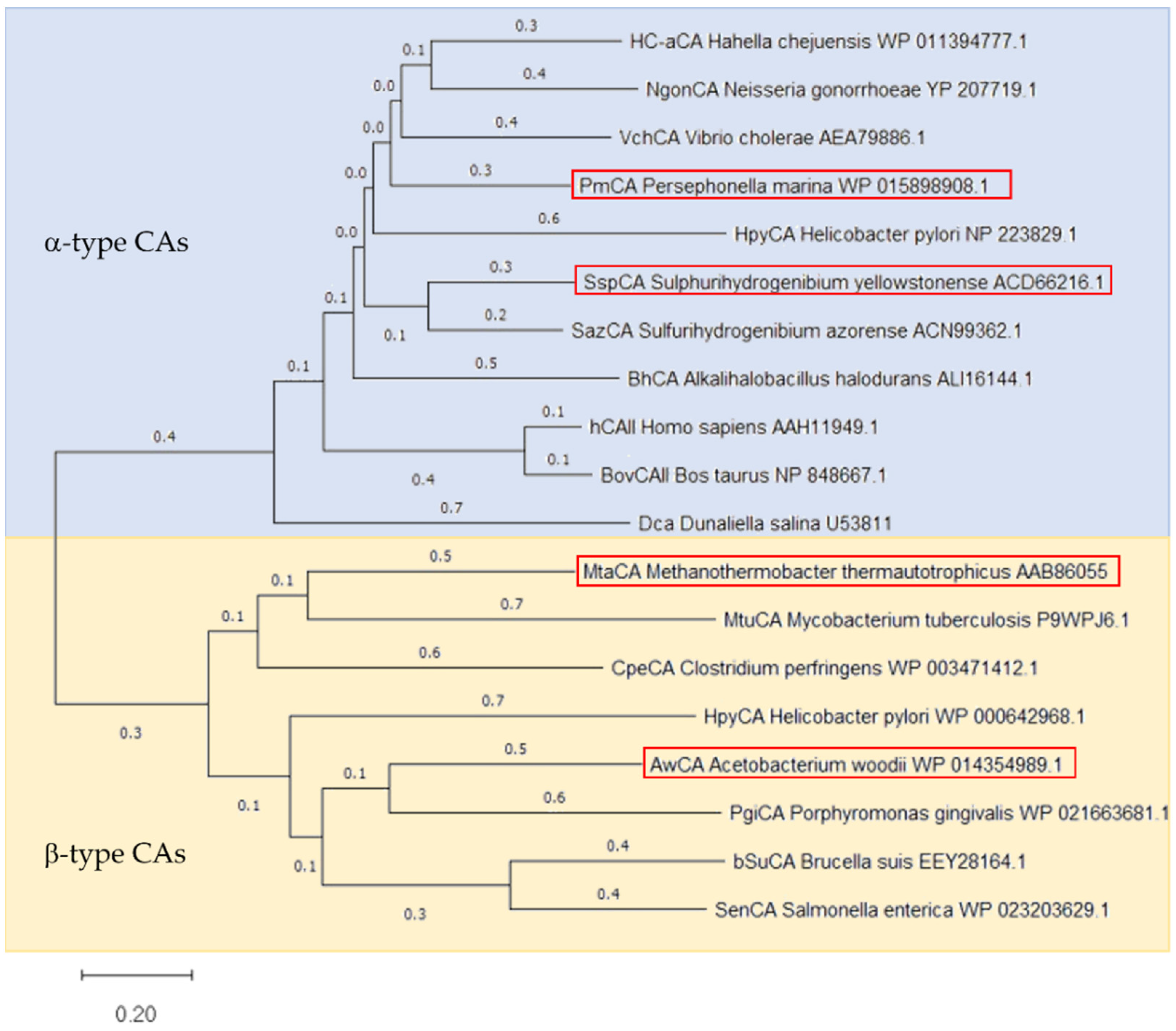
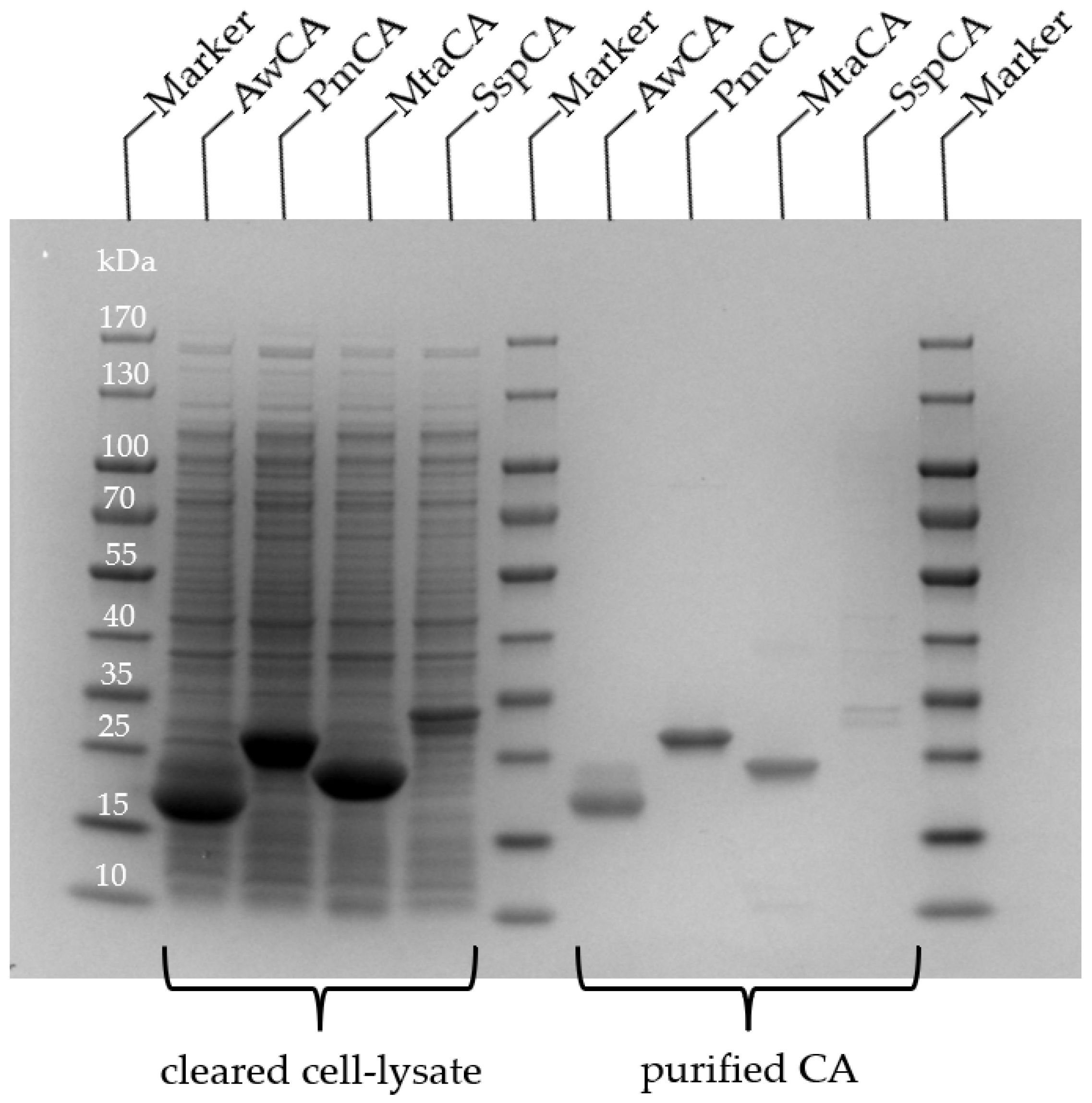
2.1. Selection, Expression and Purification
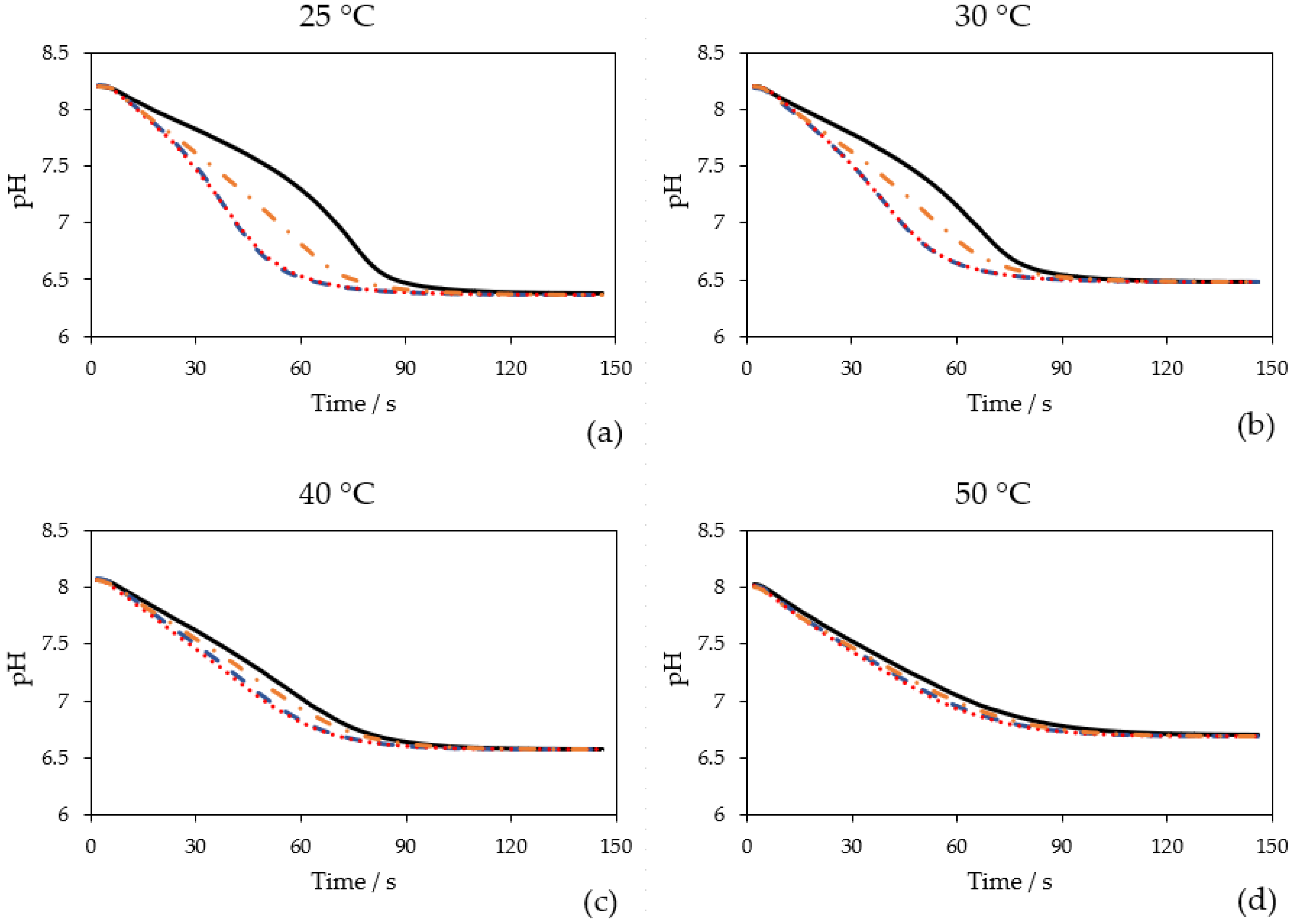
2.2. Effect of CAs on CO2 Hydration at Different Temperatures
2.3. Calculated Hydratase Activities at 25 °C
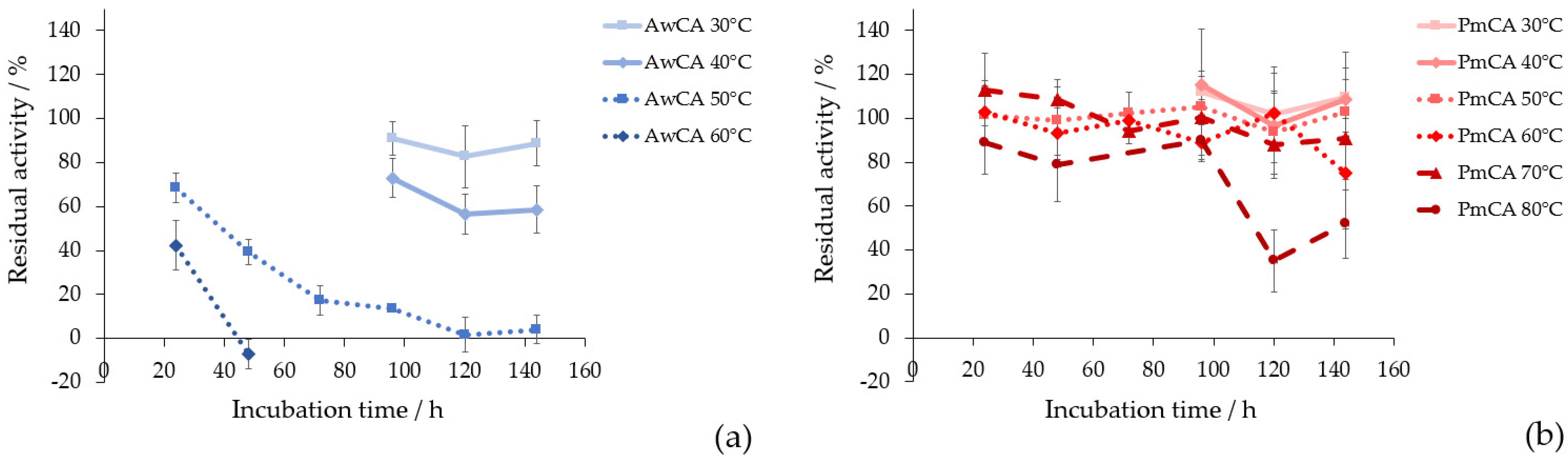
2.4. Thermostability of AwCA and PmCA
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cloning, Expression and Purification
4.3. CO2 Hydration Assay at Different Temperatures
4.4. Calculation of Hydratase Activities at 25 °C
4.5. Determination of Thermostability
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Canadell, J.G.; Le Quéré, C.; Raupach, M.R.; Field, C.B.; Buitenhuis, E.T.; Ciais, P.; Conway, T.J.; Gillett, N.P.; Houghton, R.A.; Marland, G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proc. Natl. Acad. Sci. USA 2007, 104, 18866–18870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Total Greenhous Gas Emission Trends and Projections in Europe. Available online: https://www.eea.europa.eu/data-and-maps/indicators/greenhouse-gas-emission-trends-7/assessment (accessed on 21 December 2020).
- Greenhouse Gas Emissions. Available online: https://www.epa.gov/ghgemissions/overview-greenhouse-gases (accessed on 21 December 2020).
- The Paris Agreement. Available online: https://unfccc.int/process-and-meetings/the-paris-agreement/what-is-the-paris-agreement (accessed on 8 November 2020).
- Benson, S.M.; Surles, T. Carbon dioxide capture and storage: An overview with emphasis on capture and storage in deep geological formations. Proc. IEEE 2006, 94, 1795–1804. [Google Scholar] [CrossRef] [Green Version]
- Kanniche, M.; Gros-Bonnivard, R.; Jaud, P.; Valle-Marcos, J.; Amann, J.-M.; Bouallou, C. Pre-combustion, post-combustion and oxy-combustion in thermal power plant for CO2 capture. Appl. Therm. Eng. 2010, 30, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, T.; Blunt, M.J.; Anthony, E.J.; Park, A.-H.A.; Hughes, R.W.; Webley, P.A.; Yan, J. Advances in carbon capture, utilization and storage. Appl. Energy 2020, 278, 115627. [Google Scholar] [CrossRef]
- Grant, T.; Anderson, C.; Hooper, B. Comparative life cycle assessment of potassium carbonate and monoethanolamine solvents for CO2 capture from post combustion flue gases. Int. J. Greenh. Gas Control 2014, 28, 35–44. [Google Scholar] [CrossRef]
- Mazari, S.A.; Si Ali, B.; Jan, B.M.; Saeed, I.M.; Nizamuddin, S. An overview of solvent management and emissions of amine-based CO2 capture technology. Int. J. Greenh. Gas Control 2015, 34, 129–140. [Google Scholar] [CrossRef]
- Kwak, N.-S.; Lee, J.H.; Lee, I.Y.; Jang, K.R.; Shim, J.-G. A study of the CO2 capture pilot plant by amine absorption. Energy 2012, 47, 41–46. [Google Scholar] [CrossRef]
- Bhown, A.S.; Alto, P. Direction of CO2 capture R&D. In Proceedings of the NETL CO2 Capture Technology Meeting, Pittsburgh, PA, USA, 29 July–1 August 2014; pp. 10–12. [Google Scholar]
- Di Fiore, A.; Alterio, V.; Monti, S.M.; De Simone, G.; D’Ambrosio, K. Thermostable carbonic anhydrases in biotechnological applications. Int. J. Mol. Sci. 2015, 16, 15456–15480. [Google Scholar] [CrossRef]
- Bond, G.M.; Stringer, J.; Brandvold, D.K.; Simsek, F.A.; Medina, M.-G.; Egeland, G. Development of integrated system for biomimetic CO2 sequestration using the enzyme carbonic anhydrase. Energy Fuels 2001, 15, 309–316. [Google Scholar] [CrossRef]
- González, J.M.; Fisher, S.Z. Carbonic anhydrases in industrial applications. In Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications, 1st ed.; Frost, S.C., McKenna, R., Eds.; Springer: Dordrecht, The Netharlands, 2014; pp. 405–426. [Google Scholar] [CrossRef]
- Fradette, L.; Lefebvre, S.; Carley, J. Demonstration results of enzyme-accelerated CO2 capture. Energy Procedia 2017, 114, 1100–1109. [Google Scholar] [CrossRef]
- Savile, C.K.; Lalonde, J.J. Biotechnology for the acceleration of carbon dioxide capture and sequestration. Curr. Opin. Biotechnol. 2011, 22, 818–823. [Google Scholar] [CrossRef]
- Alterio, V.; Monti, S.M.; De Simone, G. Thermal-stable carbonic anhydrases: A structural overview. In Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications, 1st ed.; Frost, S.C., McKenna, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 387–404. [Google Scholar] [CrossRef]
- Ozensoy Guler, O.; Capasso, C.; Supuran, C.T. A magnificent enzyme superfamily: Carbonic anhydrases, their purification and characterization. J. Enzyme Inhib. Med. Chem. 2016, 31, 689–694. [Google Scholar] [CrossRef]
- Boone, C.D.; Pinard, M.; McKenna, R.; Silverman, D. Catalytic mechanism of α-class carbonic anhydrases: CO2 hydration and proton transfer. In Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications; Frost, S.C., McKenna, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 31–52. [Google Scholar] [CrossRef]
- Lindskog, S. Structure and mechanism of Carbonic Anhydrase. Pharmacol. Ther. 1997, 74, 1–20. [Google Scholar] [CrossRef]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Littlechild, J.A. Enzymes from extreme environments and their industrial applications. Front. Bioeng. Biotechnol. 2015, 3, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, D.; Banta, A.; Beveridge, T.J.; Rushdi, A.I.; Simoneit, B.R.T.; Reysenbach, A.-L. Persephonella marina gen. nov., sp. nov. and Persephonella guaymasensis sp. nov., two novel, thermophilic, hydrogen-oxidizing microaerophiles from deep-sea hydrothermal vents. Int. J. Syst. Evol. Microbiol. 2002, 52, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Borchert, M.S. Heat-Stable Persephonella Carbonic Anhydrases and Their Use. U.S. Patent 9909115, 6 March 2018. [Google Scholar]
- Daigle, R.; Madore, É.; Fradette, S. Techniques for CO2 Capture Using Sulfurihydrogenibium Sp. Carbonic Anhydrase. U.S. Patent 9968885, 15 May 2018. [Google Scholar]
- Di Fiore, A.; Capasso, C.; De Luca, V.; Monti, S.M.; Carginale, V.; Supuran, C.T.; Scozzafava, A.; Pedone, C.; Rossi, M.; De Simone, G. X-ray structure of the first ‘extremo-α-carbonic anhydrase’, a dimeric enzyme from the thermophilic bacterium Sulfurihydrogenibium yellowstonense YO3AOP1. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1150–1159. [Google Scholar] [CrossRef]
- Rossi, M. A New Heat-Stable Carbonic Anhydrase and Uses. Thereof. Patent WO 2013064195, 10 May 2013. [Google Scholar]
- Vullo, D.; De Luca, V.; Scozzafava, A.; Carginale, V.; Rossi, M.; Supuran, C.T.; Capasso, C. The first activation study of a bacterial carbonic anhydrase (CA). The thermostable α-CA from Sulfurihydrogenibium yellowstonense YO3AOP1 is highly activated by amino acids and amines. Bioorgan. Med. Chem. Lett. 2012, 22, 6324–6327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanth, B.K.; Jun, S.-Y.; Kumari, S.; Pack, S.P. Highly thermostable carbonic anhydrase from Persephonella marina EX-H1: Its expression and characterization for CO2-sequestration applications. Process Biochem. 2014, 49, 2114–2121. [Google Scholar] [CrossRef]
- Capasso, C.; De Luca, V.; Carginale, V.; Cannio, R.; Rossi, M. Biochemical properties of a novel and highly thermostable bacterial α-carbonic anhydrase from Sulfurihydrogenibium yellowstonense YO3AOP1. J. Enzyme Inhib. Med. Chem. 2012, 27, 892–897. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.S.; Ferry, J.G. A plant-type (β-class) carbonic anhydrase in the thermophilic methanoarchaeon Methanobacterium thermoautotrophicum. J. Bacteriol. 1999, 181, 6247–6253. [Google Scholar] [CrossRef] [Green Version]
- Zeikus, J.G.; Wolfe, R.S. Methanobacterium thermoautotrophicus sp. n., an anaerobic, autotrophic, extreme thermophile. J. Bacteriol. 1972, 109, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Balch, W.E.; Schoberth, S.; Tanner, R.S.; Wolfe, R.S. Acetobacterium, a new genus of hydrogen-oxidizing, carbon dioxide-reducing, anaerobic bacteria. Int. Assoc. Microbiol. Soc. 1977, 27, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braus-Stromeyer, S.A.; Schnappauf, G.; Braus, G.H.; Gößner, A.S.; Drake, H.L. Carbonic anhydrase in Acetobacterium woodii and other acetogenic bacteria. J. Bacteriol. 1997, 179, 7197–7200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial enzyme applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Fuchs, W.; Steger, F.; Reich, J.; Ribitsch, D.; Rittmann, S.K.M.; Bochmann, G. A simple and straightforward method for activity measurement of carbonic anhydrases. Catalysts 2021, 11, 819. [Google Scholar] [CrossRef]
- Wilbur, K.M.; Anderson, N.G. Electrometric and colorimetric determination of carbonic anhydrase. J. Biol. Chem. 1948, 176, 147–154. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzyme Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowlett, R.S. Structure and catalytic mechanism of β-carbonic anhydrases. In Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications; Frost, S.C., McKenna, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 53–76. [Google Scholar] [CrossRef]
- Dodds, W.S.; Stutzman, L.F.; Sollami, B.J. Carbon dioxide solubility in water. Ind. Eng. Chem. Chem. Eng. Data Ser. 1956, 1, 92–95. [Google Scholar] [CrossRef]
- Jo, B.H.; Seo, J.H.; Cha, H.J. Bacterial extremo-α-carbonic anhydrases from deep-sea hydrothermal vents as potential biocatalysts for CO2 sequestration. J. Mol. Catal. B Enzym. 2014, 109, 31–39. [Google Scholar] [CrossRef]
- Smith, K.S.; Jakubzick, C.; Whittam, T.S.; Ferry, J.G. Carbonic anhydrase is an ancient enzyme widespread in prokaryotes. Proc. Natl. Acad. Sci. USA 1999, 96, 15184–15189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T. Carbonic anhydrases—An overview. Curr. Pharm. Des. 2008, 14, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Pander, B.; Harris, G.; Scott, D.J.; Winzer, K.; Köpke, M.; Simpson, S.D.; Minton, N.P.; Henstra, A.M. The carbonic anhydrase of Clostridium autoethanogenum represents a new subclass of β-carbonic anhydrases. Appl. Microbiol. Biotechnol. 2019, 103, 7275–7286. [Google Scholar] [CrossRef] [Green Version]
- Ki, M.R.; Min, K.; Kanth, B.K.; Lee, J.; Pack, S.P. Expression, reconstruction and characterization of codon-optimized carbonic anhydrase from Hahella chejuensis for CO2 sequestration application. Bioprocess Biosyst. Eng. 2013, 36, 375–381. [Google Scholar] [CrossRef]
- Faridi, S.; Satyanarayana, T. Novel alkalistable α-carbonic anhydrase from the polyextremophilic bacterium Bacillus halodurans: Characteristics and applicability in flue gas CO2 sequestration. Environ. Sci. Pollut. Res. 2016, 23, 15236–15249. [Google Scholar] [CrossRef]
- Ramanan, R.; Kannan, K.; Vinayagamoorthy, N.; Ramkumar, K.M.; Sivanesan, S.D.; Chakrabarti, T. Purification and characterization of a novel plant-type carbonic anhydrase from Bacillus subtilis. Biotechnol. Bioprocess Eng. 2009, 14, 32–37. [Google Scholar] [CrossRef]
- Premkumar, L.; Bageshwar, U.K.; Gokhman, I.; Zamir, A.; Sussman, J.L. An unusual halotolerant α-type carbonic anhydrase from the alga Dunaliella salina functionally expressed in Escherichia coli. Protein Expr. Purif. 2003, 28, 151–157. [Google Scholar] [CrossRef]
- Pavičić-Hamer, D.; Baričević, A.; Gerdol, M.; Hamer, B. Mytilus galloprovincialis carbonic anhydrase II: Activity and cDNA sequence analysis. Key Eng. Mater. 2015, 672, 137–150. [Google Scholar] [CrossRef]
- Cardoso, J.C.R.; Ferreira, V.; Zhang, X.; Anjos, L.; Félix, R.C.; Batista, F.M.; Power, D.M. Evolution and diversity of alpha-carbonic anhydrases in the mantle of the Mediterranean mussel (Mytilus galloprovincialis). Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Jensen, E.L.; Clement, R.; Kosta, A.; Maberly, S.C.; Gontero, B. A new widespread subclass of carbonic anhydrase in marine phytoplankton. ISME J. 2019, 13, 2094–2106. [Google Scholar] [CrossRef] [Green Version]
- Karakostis, K.; Costa, C.; Zito, F.; Brümmer, F.; Matranga, V. Characterization of an alpha type carbonic anhydrase from Paracentrotus lividus sea urchin embryos. Mar. Biotechnol. 2016, 18, 384–395. [Google Scholar] [CrossRef]
- Wang, L.; Liang, J.; Zhou, Y.; Tian, T.; Zhang, B.; Duanmu, D. Molecular characterization of carbonic anhydrase genes in Lotus japonicus and their potential roles in symbiotic nitrogen fixation. Int. J. Mol. Sci. 2021, 22, 7766. [Google Scholar] [CrossRef]
- Chatterjee, J.; Coe, R.A.; Acebron, K.; Thakur, V.; Yennamalli, R.M.; Danila, F.; Lin, H.-C.; Balahadia, C.P.; Bagunu, E.; Padhma, P.P.O.S.; et al. A low CO2-responsive mutant of Setaria viridis reveals that reduced carbonic anhydrase limits C4 photosynthesis. J. Exp. Bot. 2021, 72, 3122–3136. [Google Scholar] [CrossRef]
- Da Costa Ores, J.; Sala, L.; Cerveira, G.P.; Kalil, S.J. Purification of carbonic anhydrase from bovine erythrocytes and its application in the enzymic capture of carbon dioxide. Chemosphere 2012, 88, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Ekinci, D.; Beydemir, Ş.; Alim, Z. Some drugs inhibit in vitro hydratase and esterase activities of human carbonic anhydrase-I and II. Pharmacol. Reports 2007, 59, 580–587. [Google Scholar]
- Kim, S.; Sung, J.; Yeon, J.; Choi, S.H.; Jin, M.S. Crystal structure of a highly thermostable α-carbonic anhydrase from Persephonella marina EX-H1. Mol. Cells 2019, 42, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.E.; Olivieri, G.; Capasso, C.; De Luca, V.; Marzocchella, A.; Salatino, P.; Rossi, M. Kinetic study of a novel thermo-stable α-carbonic anhydrase for biomimetic CO2 capture. Enzyme Microb. Technol. 2013, 53, 271–277. [Google Scholar] [CrossRef]
- Sharma, A.; Bhattacharya, A.; Shrivastava, A. Biomimetic CO2 sequestration using purified carbonic anhydrase from indigenous bacterial strains immobilized on biopolymeric materials. Enzyme Microb. Technol. 2011, 48, 416–426. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Total Turnover Rate in µmol s−1 L−1 | Turnover Rate Minus Blank in µmol s−1 L−1 | Turnover Rate per mg Enzyme in µmol s−1 mg−1 | WAU per mg Enzyme |
|---|---|---|---|---|
| Blank | 793 ± 16 | 0 | n. a. | n. a. |
| AwCA | 1502 ± 45 | 710 ± 48 | 2839 ± 97 | 1814 ± 228 |
| MtaCA | 1204 ± 32 | 411 ± 36 | 1646 ± 73 | 580 ± 102 |
| PmCA | 1490 ± 13 | 698 ± 21 | 2790 ± 42 | 1748 ± 63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steger, F.; Reich, J.; Fuchs, W.; Rittmann, S.K.-M.R.; Gübitz, G.M.; Ribitsch, D.; Bochmann, G. Comparison of Carbonic Anhydrases for CO2 Sequestration. Int. J. Mol. Sci. 2022, 23, 957. https://doi.org/10.3390/ijms23020957
Steger F, Reich J, Fuchs W, Rittmann SK-MR, Gübitz GM, Ribitsch D, Bochmann G. Comparison of Carbonic Anhydrases for CO2 Sequestration. International Journal of Molecular Sciences. 2022; 23(2):957. https://doi.org/10.3390/ijms23020957
Chicago/Turabian StyleSteger, Franziska, Johanna Reich, Werner Fuchs, Simon K.-M. R. Rittmann, Georg M. Gübitz, Doris Ribitsch, and Günther Bochmann. 2022. "Comparison of Carbonic Anhydrases for CO2 Sequestration" International Journal of Molecular Sciences 23, no. 2: 957. https://doi.org/10.3390/ijms23020957
APA StyleSteger, F., Reich, J., Fuchs, W., Rittmann, S. K. -M. R., Gübitz, G. M., Ribitsch, D., & Bochmann, G. (2022). Comparison of Carbonic Anhydrases for CO2 Sequestration. International Journal of Molecular Sciences, 23(2), 957. https://doi.org/10.3390/ijms23020957