

Comparative Genomic and Expression Analysis Insight into Evolutionary Characteristics of PEBP Genes in Cultivated Peanuts and Their Roles in Floral Induction
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of PEBP Proteins
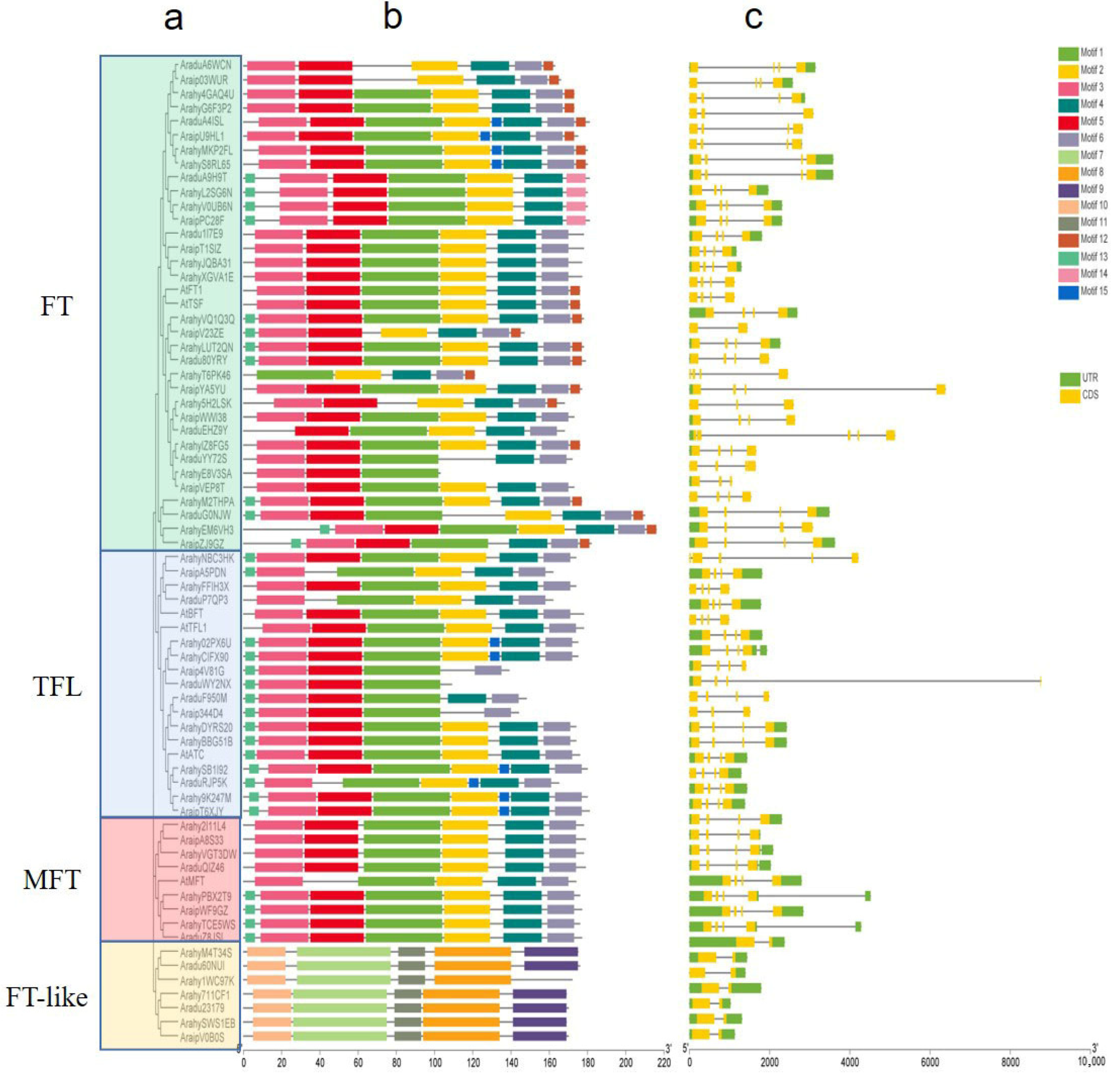
2.2. Structure and Orthologous Relationship Analysis of PEBP Genes
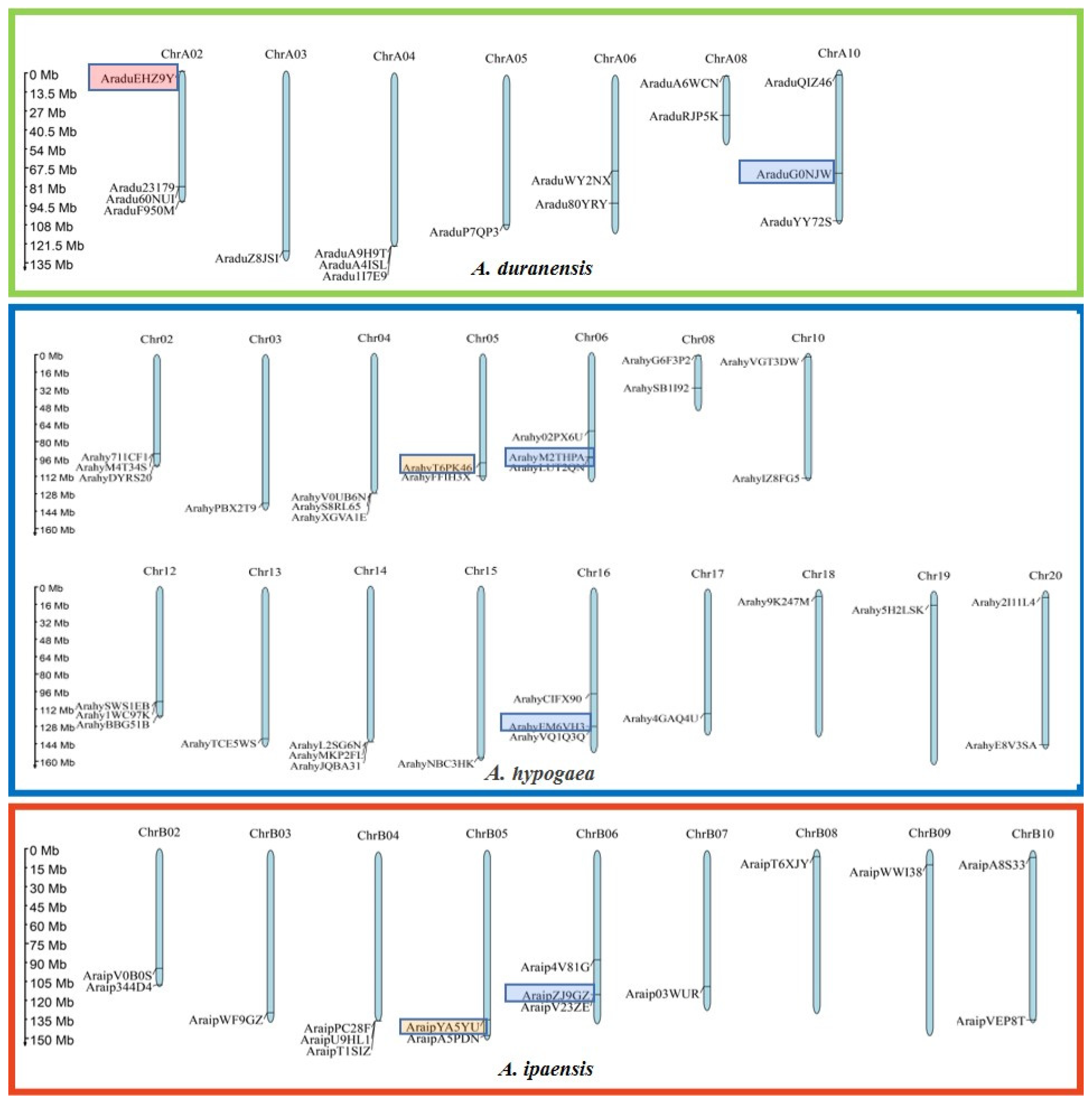
2.3. Distribution on Chromosomes and Synteny Analysis of PEBP Genes
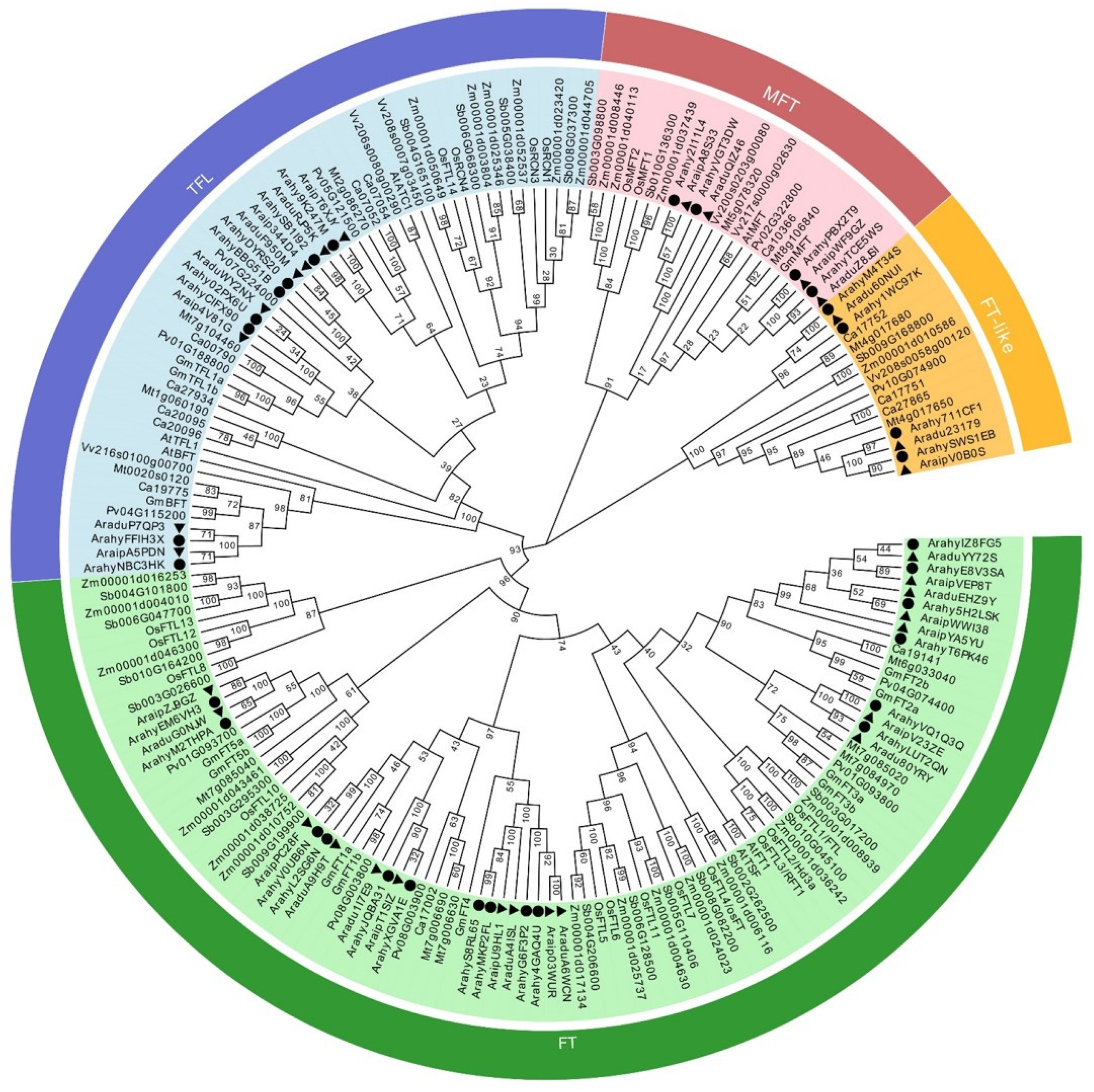
2.4. Phylogenetic Analysis for PEBP Genes among Cultivated Peanust and Other Plant Species
2.5. Cis-Element in the Promoter Region of PEBP Genes
2.6. Phenotypic Identification of Peanut Cultivars with Different Flowering Times in Response to Photoperiod and Temperature
2.7. PEBP Expression Pattern Analysis Responds to Genotypes, Temperature and Photoperiod in Early and Late Flowering Genotypes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Download for Genome Data
4.3. Genome-Wide Identification of PEBP Proteins
4.4. Chromosome Location, Phylogenetic Analysis
4.5. Gene Structure and Conserved Motifs Characterization of PEBP Genes
4.6. Gene Collinearity and Selection Pressure Analysis
4.7. Cis-Element Analysis of the PEBP Genes
4.8. Quantitative RT-PCR Analysis for Gene Expression
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fornara, F.; de Montaigu, A.; Coupland, G. SnapShot: Control of Flowering in Arabidopsis. Cell 2010, 141, 550. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, A.; Richter, R. Genetic and molecular basis of floral induction in Arabidopsis thaliana. J. Exp. Bot. 2020, 71, 2490–2504. [Google Scholar] [CrossRef]
- Zhu, Y.; Klasfeld, S.; Wagner, D. Molecular regulation of plant developmental transitions and plant architecture via PEPB family proteins: An update on mechanism of action. J. Exp. Bot. 2021, 72, 2301–2311. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Nasim, Z.; Susila, H.; Ahn, J.H. Evolution and functional diversification of FLOWERING LOCUS T/TERMINAL FLOWER 1 family genes in plants. Semin. Cell Dev. Biol. 2021, 109, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Putterill, J.; Varkonyi-Gasic, E. FT and florigen long-distance flowering control in plants. Curr. Opin. Plant Biol. 2016, 33, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, T.; Zeng, X.; He, D.; He, Y. Feedback Regulation of FLC by FLOWERING LOCUS T (FT) and FD through a 5’ FLC Promoter Region in Arabidopsis. Mol. Plant 2019, 12, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Kobayashi, Y.; Goto, K.; Abe, M.; Araki, T. TWIN SISTER OF FT (TSF) Acts as a Floral Pathway Integrator Redundantly with FT. Plant Cell Physiol. 2005, 46, 1175–1189. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.-C.; Jane, W.-N.; Chen, J.; Yu, T.-S. Arabidopsis thaliana CENTRORADIALIS homologue (ATC) acts systemically to inhibit floral initiation in Arabidopsis. Plant J. 2012, 72, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.J.; Chung, K.S.; Jung, S.H.; Yoo, S.Y.; Lee, J.S.; Ahn, J.H. BROTHER OF FT AND TFL1 (BFT) has TFL1-like activity and functions redundantly with TFL1 in inflorescence meristem development in Arabidopsis. Plant J. 2010, 63, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, R.; Devisetty, U.K.; Maloof, J.N.; Zuo, Y.; Li, J.; Shen, Y.; Zhao, J.; Bao, M.; Ning, G. The Divergence of Flowering Time Modulated by FT/TFL1 Is Independent to Their Interaction and Binding Activities. Front Plant Sci. 2017, 8, 697. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.Y.; Lee, H.-J.; Seo, P.J.; Jung, J.-H.; Ahn, J.H.; Park, C.-M. The Arabidopsis Floral Repressor BFT Delays Flowering by Competing with FT for FD Binding under High Salinity. Mol. Plant 2014, 7, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Vaistij, F.E.; Barros-Galvão, T.; Cole, A.F.; Gilday, A.D.; He, Z.; Li, Y.; Harvey, D.; Larson, T.R.; Graham, I.A. MOTHER-OF-FT-AND-TFL1 represses seed germination under far-red light by modulating phytohormone responses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2018, 115, 8442–8447. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Liu, H.; Sang, N.; Li, Y.; Zhang, T.; Sun, J.; Huang, X. Identification of cotton MOTHER OF FT AND TFL1 homologs, GhMFT1 and GhMFT2, involved in seed germination. PLoS ONE 2019, 14, e0215771. [Google Scholar] [CrossRef] [Green Version]
- Chardon, F.; Damerval, C. Phylogenomic Analysis of the PEBP Gene Family in Cereals. J. Mol. Evol. 2005, 61, 579–590. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Tamaki, S.; Matsuo, S.; Wong, H.L.; Yokoi, S.; Shimamoto, K. Hd3a protein is a mobile flowering signal in rice. Science 2007, 316, 1033–1036. [Google Scholar] [CrossRef]
- Kong, F.; Liu, B.; Xia, Z.; Sato, S.; Kim, B.M.; Watanabe, S.; Yamada, T.; Tabata, S.; Kanazawa, A.; Harada, K.; et al. Two Coordinately Regulated Homologs of FLOWERING LOCUS T Are Involved in the Control of Photoperiodic Flowering in Soybean. Plant Physiol. 2010, 154, 1220–1231. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Jiang, B.; Ma, L.; Zhang, S.; Zhai, H.; Xu, X.; Hou, W.; Xia, Z.; Wu, C.; Sun, S.; et al. Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation. New Phytol. 2018, 217, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Muszynski, M.G.; Danilevskaya, O.N. The FT-Like ZCN8 Gene Functions as a Floral Activator and Is Involved in Photoperiod Sensitivity in Maize. Plant Cell 2011, 23, 942–960. [Google Scholar] [CrossRef] [Green Version]
- Kaba, J.S.; Kumaga, F.; Ofori, K. Effect of flower production and time of flowering on pod yield of peanut (Arachis hypogaea L) genotypes. IOSR J. Agric. Vet. Sci. 2014, 7, 44–49. [Google Scholar] [CrossRef]
- Bell, M.J.; Bagnall, D.J.; Harch, G. Effects of photoperiod on reproductive development of peanut (Arachis hypogaea L.) in a cool subtropical environment. II. Temperature interactions. Aust. J. Agric. Res. 1991, 42, 1151–1161. [Google Scholar] [CrossRef]
- Nigam, S.N.; Dwivedi, S.L.; Ramraj, V.M.; Chandra, S. Combining Ability of Response to Photoperiod in Peanut. Crop Sci. 1997, 37, 1159–1162. [Google Scholar] [CrossRef]
- Moretzsohn, M.C.; Gouvea, E.G.; Inglis, P.W.; Leal-Bertioli, S.C.M.; Valls, J.F.M.; Bertioli, D.J. A study of the relationships of cultivated peanut (Arachis hypogaea) and its most closely related wild species using intron sequences and microsatellite markers. Ann. Bot. 2013, 111, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Gao, C.; Bian, X.; Zhao, S.; Zhao, C.; Xia, H.; Song, H.; Hou, L.; Wan, S.; Wang, X. Genome-Wide Identification and Comparative Analysis of Cytosine-5 DNA Methyltransferase and Demethylase Families in Wild and Cultivated Peanut. Front Plant Sci. 2016, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Tang, X.; Xing, M.; Zhu, H.; Sui, J.; Cai, C.; Li, S. Molecular and transcriptional characterization of phosphatidyl ethanolamine-binding proteins in wild peanuts Arachis duranensis and Arachis ipaensis. BMC Plant Biol. 2019, 19, 484. [Google Scholar] [CrossRef] [Green Version]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Lu, Q.; Liu, H.; Zhang, J.; Hong, Y.; Lan, H.; Li, H.; Wang, J.; Liu, H.; Li, S.; et al. Sequencing of Cultivated Peanut, Arachis hypogaea, Yields Insights into Genome Evolution and Oil Improvement. Mol. Plant 2019, 12, 920–934. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.-C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef] [Green Version]
- Karlgren, A.; Gyllenstrand, N.; Källman, T.; Sundström, J.F.; Moore, D.; Lascoux, M.; Lagercrantz, U. Evolution of the PEBP Gene Family in Plants: Functional Diversification in Seed Plant Evolution. Plant Physiol. 2011, 156, 1967–1977. [Google Scholar] [CrossRef] [Green Version]
- Tsoy, O.; Mushegian, A. Florigen and its homologs of FT/CETS/PEBP/RKIP/YbhB family may be the enzymes of small molecule metabolism: Review of the evidence. BMC Plant Biol. 2022, 22, 56. [Google Scholar] [CrossRef]
- Wang, M.; Tan, Y.; Cai, C.; Zhang, B. Identification and expression analysis of phosphatidy ethanolamine-binding protein (PEBP) gene family in cotton. Genomics 2019, 111, 1373–1380. [Google Scholar] [CrossRef]
- Dong, L.; Lu, Y.; Liu, S. Genome-wide member identification, phylogeny and expression analysis of PEBP gene family in wheat and its progenitors. PeerJ 2020, 8, e10483. [Google Scholar] [CrossRef]
- Zhang, G.; Jin, X.; Li, X.; Zhang, N.; Li, S.; Si, H.; Rajora, O.P.; Li, X.-Q. Genome-wide identification of PEBP gene family members in potato, their phylogenetic relationships, and expression patterns under heat stress. Mol. Biol. Rep. 2022, 49, 4683–4697. [Google Scholar] [CrossRef]
- Danilevskaya, O.N.; Meng, X.; Hou, Z.; Ananiev, E.V.; Simmons, C.R. A Genomic and Expression Compendium of the Expanded PEBP Gene Family from Maize. Plant Physiol. 2007, 146, 250–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Abe, F.; Kawahigashi, H.; Nakazono, K.; Tagiri, A.; Matsumoto, T.; Utsugi, S.; Ogawa, T.; Handa, H.; Ishida, H.; et al. A wheat homolog of MOTHER OF FT AND TFL1 acts in the regulation of germination. Plant Cell 2011, 23, 3215–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.-B.; Luo, L.; He, L.-L.; Ni, J.; Xu, Z.-F. A promoter analysis of MOTHER OF FT AND TFL11 (JcMFT1), a seed-preferential gene from the biofuel plant Jatropha curcas. J. Plant Res. 2014, 127, 513–524. [Google Scholar] [CrossRef]
- Clevenger, J.; Chu, Y.; Scheffler, B.; Ozias-Akins, P. A Developmental Transcriptome Map for Allotetraploid Arachis hypogaea. Front. Plant Sci. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Liu, C.; Hou, X.; Yu, H. MOTHER OF FT AND TFL1 regulates seed germination through a negative feedback loop modulating ABA signaling in Arabidopsis. Plant Cell 2010, 22, 1733–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunta, S.; Chu, Y.; Levy, Y.; Harel, A.; Abbo, S.; Ozias-Akins, P.; Hovav, R. Identification of a major locus for flowering pattern sheds light on plant architecture diversification in cultivated peanut. Theor. Appl. Genet. 2022, 135, 1767–1777. [Google Scholar] [CrossRef]
- Guo, G.; Xu, K.; Zhang, X.; Zhu, J.; Lu, M.; Chen, F.; Liu, L.; Xi, Z.-Y.; Bachmair, A.; Chen, Q.; et al. Extensive Analysis of GmFTL and GmCOL Expression in Northern Soybean Cultivars in Field Conditions. PLoS ONE 2015, 10, e0136601. [Google Scholar] [CrossRef]
- Zhai, H.; Lü, S.; Liang, S.; Wu, H.; Zhang, X.; Liu, B.; Kong, F.; Yuan, X.; Li, J.; Xia, Z. GmFT4, a Homolog of FLOWERING LOCUS T, Is Positively Regulated by E1 and Functions as a Flowering Repressor in Soybean. PLoS ONE 2014, 9, e89030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- No, D.H.; Baek, D.; Lee, S.H.; Cheong, M.S.; Chun, H.J.; Park, M.S.; Cho, H.M.; Jin, B.J.; Lim, L.H.; Lee, Y.B.; et al. High-Temperature Conditions Promote Soybean Flowering through the Transcriptional Reprograming of Flowering Genes in the Photoperiod Pathway. Int. J. Mol. Sci. 2021, 22, 1314. [Google Scholar] [CrossRef]
- Nigam, S.N.; Rao, R.C.N.; Wynne, J.C.; Williams, J.H.; Fitzner, M.; Nagabhushanam, G.V.S. Effect and interaction of temperature and photoperiod on growth and partitioning in three groundnut (Arachis hypogaea L.) genotypes1. Ann. Appl. Biol. 1994, 125, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Bagnall, D.J.; King, R.W. Response of peanut (Arachis hypogaea) to temperature, photoperiod and irradiance 1. Effect on flowering. Field Crops Res. 1991, 26, 263–277. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, C.; Li, Z.; Cheng, Y.; Zhang, H.; Liu, Y.; Wang, X.; Jiang, C.; Zhao, X.; Zhao, S.; Wang, J.; et al. Comparative Genomic and Expression Analysis Insight into Evolutionary Characteristics of PEBP Genes in Cultivated Peanuts and Their Roles in Floral Induction. Int. J. Mol. Sci. 2022, 23, 12429. https://doi.org/10.3390/ijms232012429
Zhong C, Li Z, Cheng Y, Zhang H, Liu Y, Wang X, Jiang C, Zhao X, Zhao S, Wang J, et al. Comparative Genomic and Expression Analysis Insight into Evolutionary Characteristics of PEBP Genes in Cultivated Peanuts and Their Roles in Floral Induction. International Journal of Molecular Sciences. 2022; 23(20):12429. https://doi.org/10.3390/ijms232012429
Chicago/Turabian StyleZhong, Chao, Zhao Li, Yunlian Cheng, Haina Zhang, Yu Liu, Xiaoguang Wang, Chunji Jiang, Xinhua Zhao, Shuli Zhao, Jing Wang, and et al. 2022. "Comparative Genomic and Expression Analysis Insight into Evolutionary Characteristics of PEBP Genes in Cultivated Peanuts and Their Roles in Floral Induction" International Journal of Molecular Sciences 23, no. 20: 12429. https://doi.org/10.3390/ijms232012429
APA StyleZhong, C., Li, Z., Cheng, Y., Zhang, H., Liu, Y., Wang, X., Jiang, C., Zhao, X., Zhao, S., Wang, J., Zhang, H., Liu, X., & Yu, H. (2022). Comparative Genomic and Expression Analysis Insight into Evolutionary Characteristics of PEBP Genes in Cultivated Peanuts and Their Roles in Floral Induction. International Journal of Molecular Sciences, 23(20), 12429. https://doi.org/10.3390/ijms232012429