

The Role of the Notch Signaling Pathway in Recovery of Cardiac Function after Myocardial Infarction


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
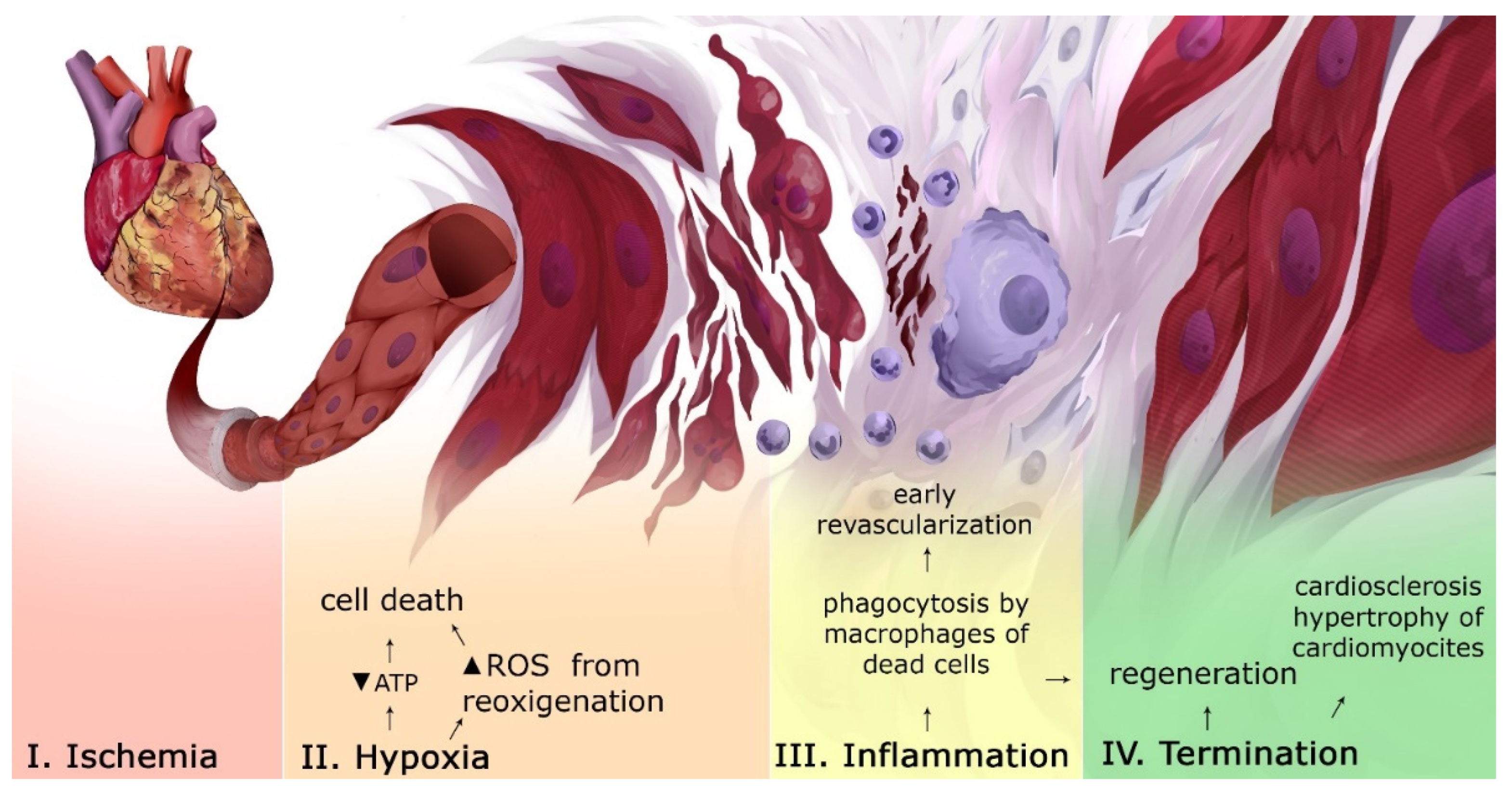
2. Pathogenesis of Myocardial Infarction
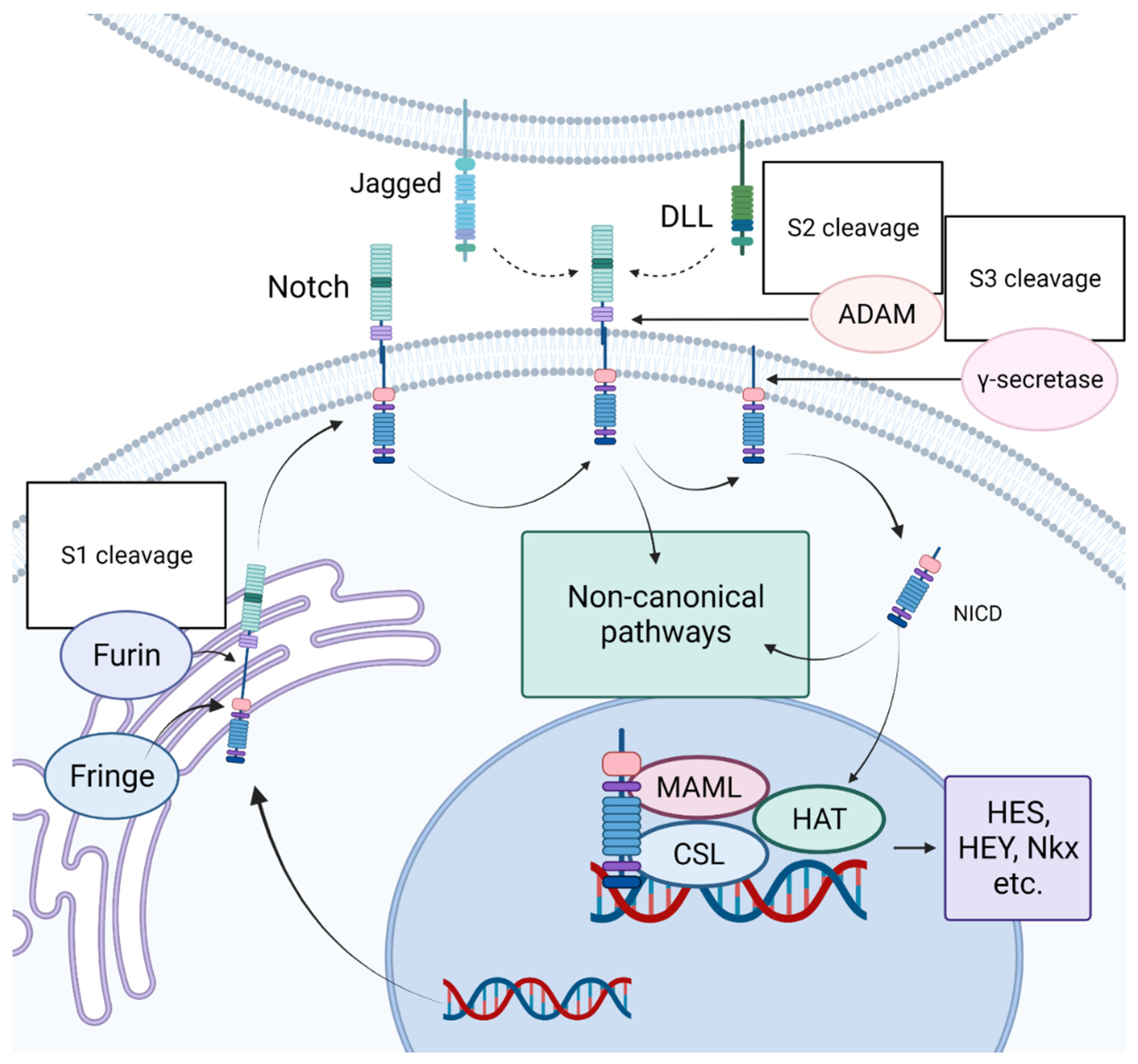
3. Notch Signaling Pathway
4. Role of the Notch Signaling Pathway in Myocardial Infarction
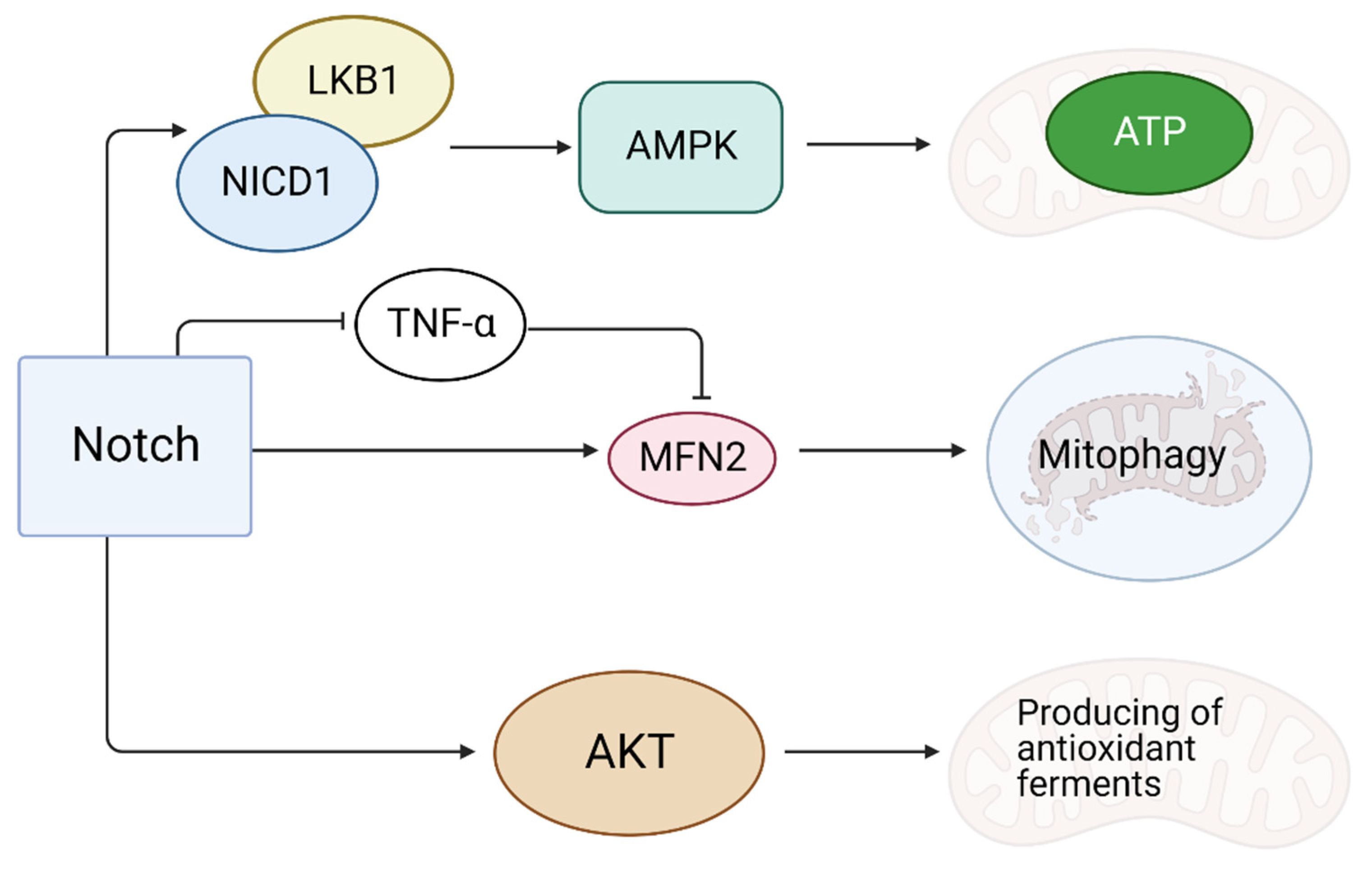
4.1. Notch Reduces Oxidative Stress
4.2. Notch Prevents Apoptosis of Cardiomyocytes
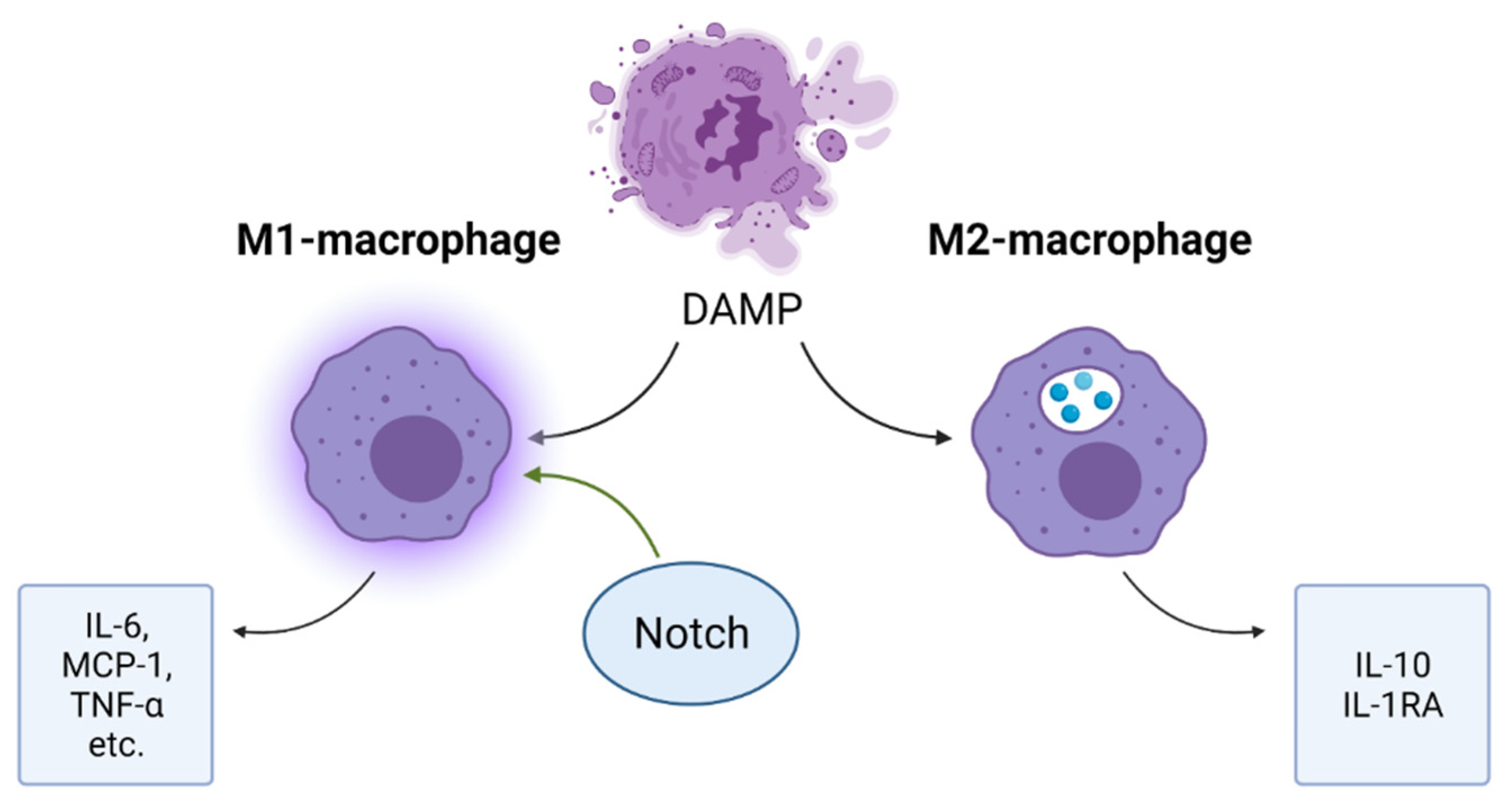
4.3. Notch and Inflammation
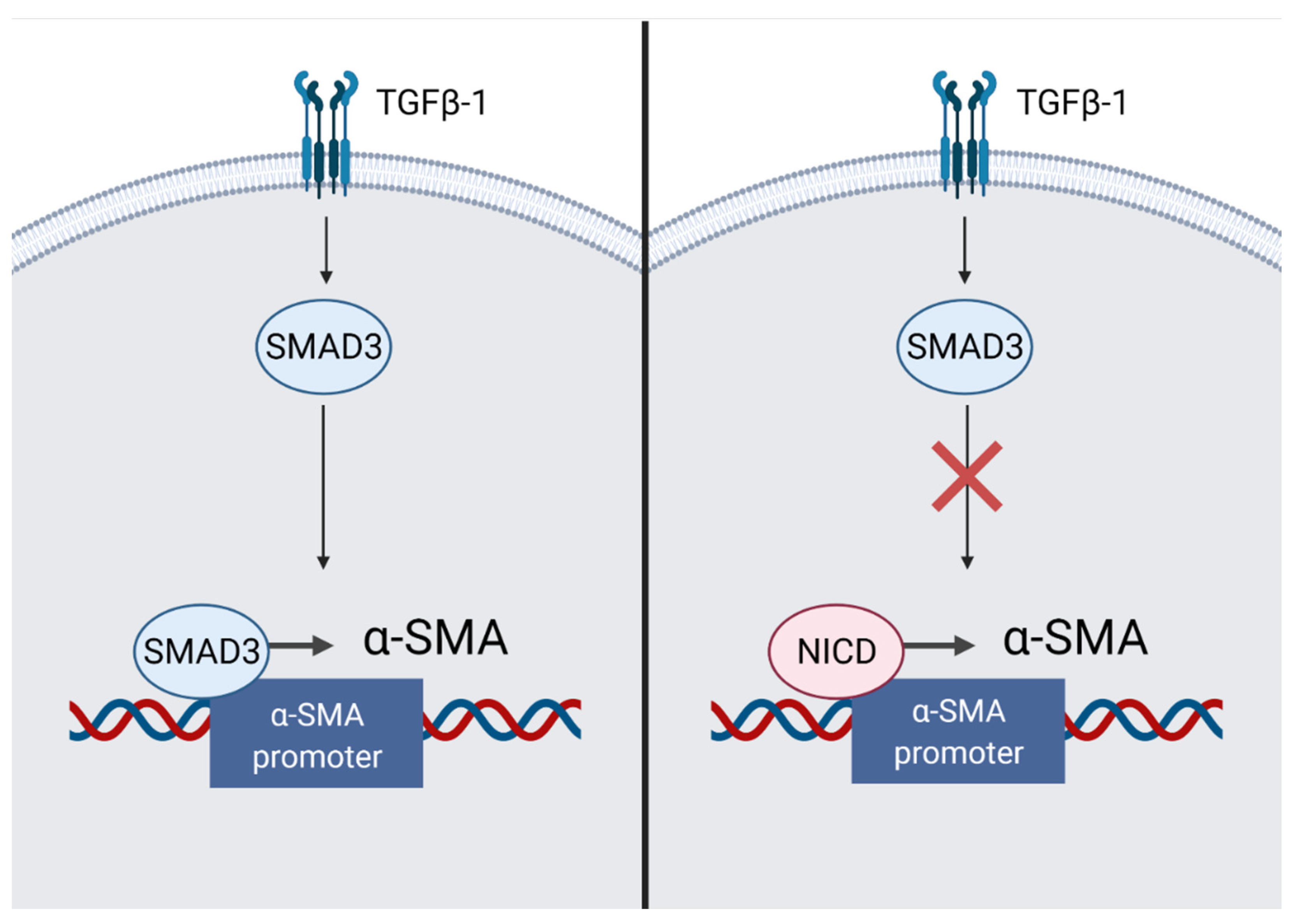
4.4. Notch Signaling Pathway Reduces Myocardial Fibrosis
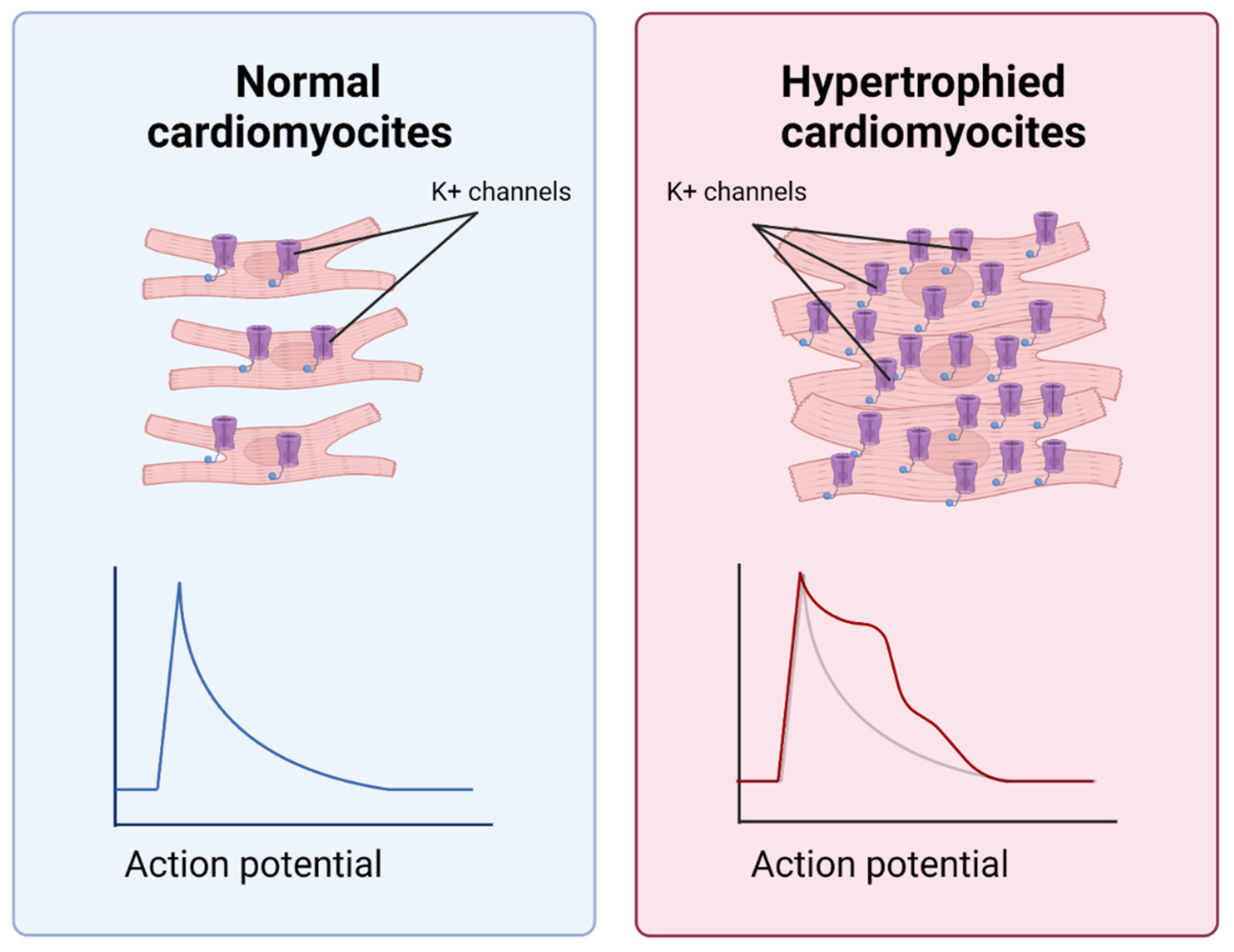
4.5. Notch Negatively Affects Cardiomyocyte Hypertrophy
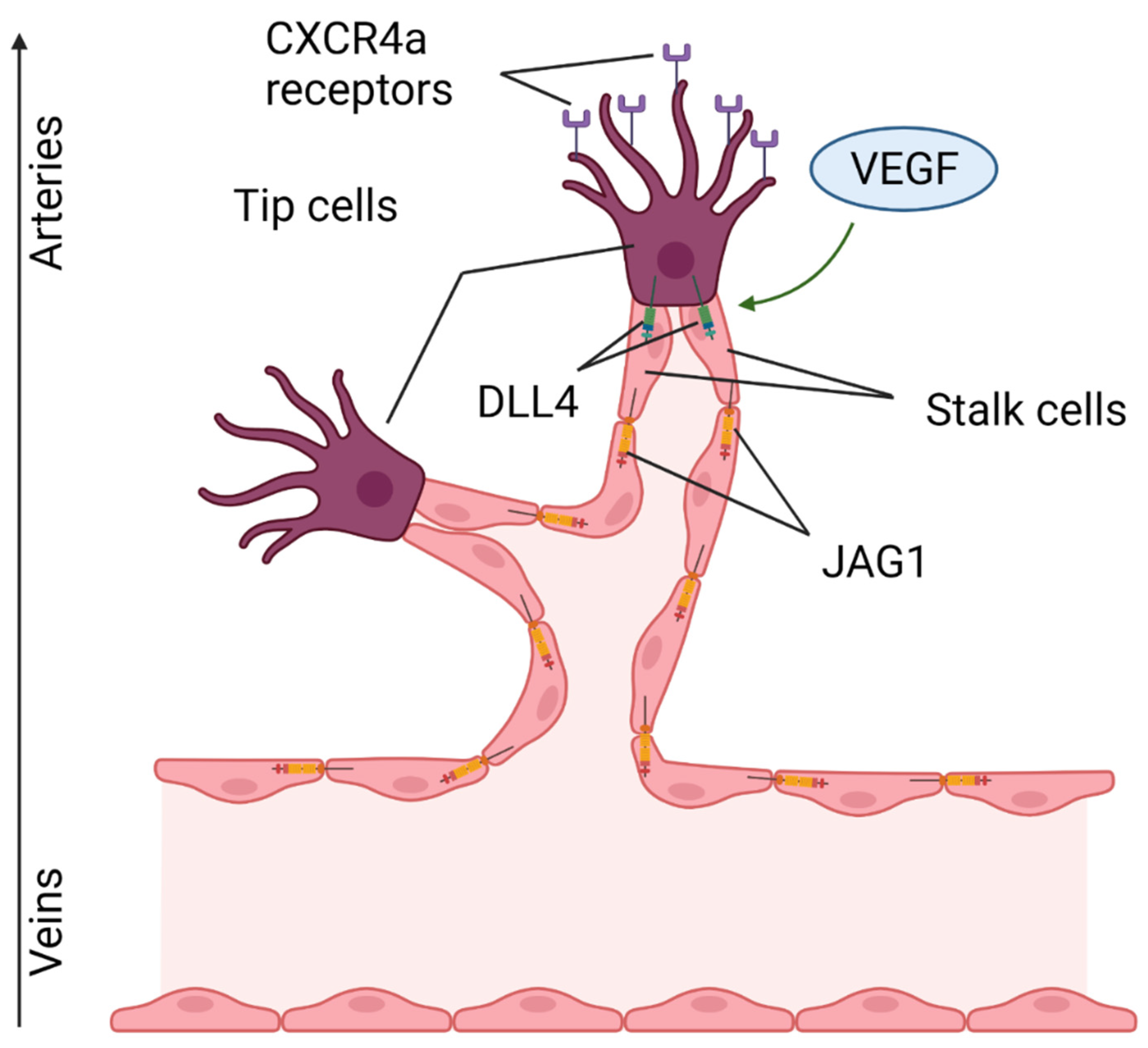
4.6. Notch Stimulates Angiogenesis
4.7. Notch Regulates Proliferation and Differentiation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cardiovascular Diseases. Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 11 July 2020).
- Chapman, A.R.; Adamson, P.D.; Shah, A.S.V.; Anand, A.; Strachan, F.E.; Ferry, A.V.; Lee, K.K.; Berry, C.; Findlay, I.; Cruikshank, A.; et al. High-sensitivity cardiac troponin and the universal definition of myocardial infarction. Circulation 2020, 141, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Neri, M.; Riezzo, I.; Pascale, N.; Pomara, C.; Turillazzi, E. Ischemia/Reperfusion Injury following Acute Myocardial Infarction: A Critical Issue for Clinicians and Forensic Pathologists. Mediat. Inflamm. 2017, 2017, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacca, M. Cardiac Regeneration After Myocardial Infarction: An Approachable Goal. Curr. Cardiol. Rep. 2020, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.; Malagù, M.; Biscaglia, S.; Fucili, A.; Rizzo, P. Remodelling after an Infarct: Crosstalk between Life and Death. Cardiology 2016, 135, 68–76. [Google Scholar] [CrossRef]
- Lesnefsky, E.J.; Chen, Q.; Tandler, B.; Hoppel, C.L. Mitochondrial Dysfunction and Myocardial Ischemia-Reperfusion: Implications for Novel Therapies. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 535–565. [Google Scholar] [CrossRef]
- Prabhu, S.D.; Frangogiannis, N.G. The biological basis for cardiac repair after myocardial infarction. Circ. Res. 2016, 119, 91–112. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Cardiac fibrosis: Cell biological mechanisms, molecular pathways and therapeutic opportunities. Mol. Asp. Med. 2019, 65, 70–99. [Google Scholar] [CrossRef]
- Artavanis Tsakonas, S.; Muskavitch, M.A.T.; Yedvobnick, B. Molecular cloning of Notch, a locus affecting neurogenesis in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1983, 80, 1977–1981. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, P.; Bollini, S.; Bertero, E.; Ferrari, R.; Ameri, P. Beyond cardiomyocyte loss: Role of Notch in cardiac aging. J. Cell. Physiol. 2018, 233, 5670–5683. [Google Scholar] [CrossRef]
- Shang, Y.; Smith, S.; Hu, X. Role of Notch signaling in regulating innate immunity and inflammation in health and disease. Protein Cell 2016, 7, 159–174. [Google Scholar] [CrossRef]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Koch, U.; Radtke, F. Dual function of notch signaling in cancer: Oncogene and tumor suppressor. In Targeting Notch in Cancer: From the Fruit Fly to the Clinic; Springer: New York, NY, USA, 2018; pp. 55–86. [Google Scholar] [CrossRef]
- Nistri, S.; Sassoli, C.; Bani, D. Notch Signaling in Ischemic Damage and Fibrosis: Evidence and Clues from the Heart. Front. Pharmacol. 2017, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- MacGrogan, D.; Münch, J.; de la Pompa, J.L. Notch and interacting signalling pathways in cardiac development, disease, and regeneration. Nat. Rev. Cardiol. 2018, 15, 685–704. [Google Scholar] [CrossRef]
- Bray, S.J. Notch signalling in context. Nat. Rev. Mol. Cell Biol. 2016, 17, 722–735. [Google Scholar] [CrossRef]
- Garg, V.; Muth, A.N.; Ransom, J.F.; Schluterman, M.K.; Barnes, R.; King, I.N.; Grossfeld, P.D.; Srivastava, D. Mutations in NOTCH1 cause aortic valve disease. Nature 2005, 437, 270–274. [Google Scholar] [CrossRef]
- Kostina, A.S.; Uspensky, V.; Irtyuga, O.B.; Ignatieva, E.V.; Freylikhman, O.; Gavriliuk, N.D.; Moiseeva, O.M.; Zhuk, S.; Tomilin, A.; Kostareva, А.А.; et al. Notch-dependent EMT is attenuated in patients with aortic aneurysm and bicuspid aortic valve. Biochim. Biophys. Acta Mol. Basis Dis. 2016, 1862, 733–740. [Google Scholar] [CrossRef]
- Irtyuga, O.; Malashicheva, A.; Zhiduleva, E.; Freylikhman, O.; Rotar, O.; Bäck, M.; Tarnovskaya, S.; Kostareva, A.; Moiseeva, O. NOTCH1 Mutations in Aortic Stenosis: Association with Osteoprotegerin/RANK/RANKL. Biomed. Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chiarini, A.; Onorati, F.; Marconi, M.; Pasquali, A.; Patuzzo, C.; Malashicheva, A.; Irtyega, O.; Faggian, G.; Pignatti, P.F.; Trabetti, E.; et al. Studies on sporadic non-syndromic thoracic aortic aneurysms: 1. Deregulation of Jagged/Notch 1 homeostasis and selection of synthetic/secretor phenotype smooth muscle cells. Eur. J. Prev. Cardiol. 2018, 25, 42–50. [Google Scholar] [CrossRef]
- Chiarini, A.; Onorati, F.; Marconi, M.; Pasquali, A.; Patuzzo, C.; Malashicheva, A.; Irtyega, O.; Faggian, G.; Pignatti, P.F.; Trabetti, E.; et al. Studies on sporadic non-syndromic thoracic aortic aneurysms: II. Alterations of extra-cellular matrix components and focal adhesion proteins. Eur. J. Prev. Cardiol. 2018, 25, 51–58. [Google Scholar] [CrossRef]
- Kostina, A.; Bjork, H.; Ignatieva, E.; Irtyuga, O.; Uspensky, V.; Semenova, D.; Maleki, S.; Tomilin, A.; Moiseeva, O.; Franco-Cereceda, A.; et al. Notch, BMP and WNT/β-catenin network is impaired in endothelial cells of the patients with thoracic aortic aneurysm. Atheroscler. Suppl. 2018, 35, e6–e13. [Google Scholar] [CrossRef]
- Kostina, A.; Shishkova, A.; Ignatieva, E.; Irtyuga, O.; Bogdanova, M.; Levchuk, K.; Golovkin, A.; Zhiduleva, E.; Uspenskiy, V.; Moiseeva, O.; et al. Different Notch signaling in cells from calcified bicuspid and tricuspid aortic valves. J. Mol. Cell. Cardiol. 2017, 114, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Kostina, A.; Semenova, D.; Kostina, D.; Uspensky, V.; Kostareva, A.; Malashicheva, A. Human aortic endothelial cells have osteogenic Notch-dependent properties in co-culture with aortic smooth muscle cells. Biochem. Biophys. Res. Commun. 2019, 514, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Kozyrev, I.; Docshin, P.; Kostina, A.; Kiselev, A.M.; Ignatieva, E.; Golovkin, A.; Pervunina, T.; Grekhov, E.; Gordeev, M.; Kostareva, A.; et al. Dysregulation of Notch signaling in cardiac mesenchymal cells of patients with tetralogy of Fallot. Pediatr. Res. 2020, 88, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Malashicheva, A.; Kostina, A.; Kostareva, A.; Irtyuga, O.; Gordeev, M.; Uspenskiy, V. Notch signaling in the pathogenesis of thoracic aortic aneurysms: A bridge between embryonic and adult states. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165631. [Google Scholar] [CrossRef]
- Theodoris, C.V.; Zhou, P.; Liu, L.; Zhang, Y.; Nishino, T.; Huang, Y.; Kostina, A.; Ranade, S.S.; Gifford, C.A. Network-based screen in iPSC-derived cells reveals therapeutic candidate for heart valve disease. Science 2021, 371, eabd0724. [Google Scholar] [CrossRef]
- Lochner, A.; Marais, E.; Huisamen, B. Melatonin and cardioprotection against ischaemia/reperfusion injury: What’s new? A review. J. Pineal Res. 2018, 65, e12490. [Google Scholar] [CrossRef] [Green Version]
- Pei, H.; Du, J.; Song, X.; He, L.; Zhang, Y.; Li, X.; Qiu, C.; Zhang, Y.; Hou, J.; Feng, J.; et al. Melatonin prevents adverse myocardial infarction remodeling via Notch1/Mfn2 pathway. Free Radic. Biol. Med. 2016, 97, 408–417. [Google Scholar] [CrossRef]
- Dai, S.-H.; Wu, Q.-C.; Zhu, R.-R.; Wan, X.-M.; Zhou, X.-L. Notch1 protects against myocardial ischaemia-reperfusion injury via regulating mitochondrial fusion and function. J. Cell. Mol. Med. 2020, 24, 3183–3191. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Li, J.; Kou, B.; Yi, Q.; Shi, T. MicroRNA-30e protects the heart against ischemia and reperfusion injury through autophagy and the Notch1/Hes1/Akt signaling pathway. Int. J. Mol. Med. 2018, 41, 3221–3230. [Google Scholar] [CrossRef]
- Yang, H.; Sun, W.; Quan, N.; Wang, L.; Chu, D.; Cates, C.; Liu, Q.; Zheng, Y.; Li, J. Cardioprotective actions of Notch1 against myocardial infarction via LKB1-dependent AMPK signaling pathway. Biochem. Pharmacol. 2016, 108, 47–57. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Pang, S.; Huang, J.; Zhu, K.; Tong, J.; Tang, Y.; Ma, G.; Chen, L. Blockade of RBP-J-Mediated Notch Signaling Pathway Exacerbates Cardiac Remodeling after Infarction by Increasing Apoptosis in Mice. BioMed Res. Int. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Villarejo-Zori, B.; Jiménez-Loygorri, J.I.; Boya, P. HIF1α or mitophagy: Which drives cardiomyocyte differentiation? Cell Stress 2020, 4, 95–98. [Google Scholar] [CrossRef]
- Kong, L.Y.; Xi, Z.; Ma, W.T.; Yang, F.Y.; Niu, L.D.; Shi, J.H. Effects of Notch signal on the expressions of HIF-α and autophagy- related genes Beclin1, LC3I, LC3II in oxygen-glucose deprivation induced myocardial cell injury. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2019, 35, 165–168. [Google Scholar] [CrossRef]
- Wang, Y.; Smith, W.; Hao, D.; He, B.; Kong, L. M1 and M2 macrophage polarization and potentially therapeutic naturally occurring compounds. Int. Immunopharmacol. 2019, 70, 459–466. [Google Scholar] [CrossRef]
- Singla, D.K.; Wang, J.; Singla, R. Primary human monocytes differentiate into M2 macrophages and involve Notch-1 pathway. Can. J. Physiol. Pharmacol. 2017, 95, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Pabois, A.; Pagie, S.; Gérard, N.; Laboisse, C.; Pattier, S.; Hulin, P.; Nedellec, S.; Toquet, C.; Charreau, B. Notch signaling mediates crosstalk between endothelial cells and macrophages via Dll4 and IL6 in cardiac microvascular inflammation. Biochem. Pharmacol. 2016, 104, 95–107. [Google Scholar] [CrossRef]
- Nattel, S. Molecular and Cellular Mechanisms of Atrial Fibrosis in Atrial Fibrillation. JACC Clin. Electrophysiol. 2017, 3, 425–435. [Google Scholar] [CrossRef]
- Fan, Z.; Guan, J. Antifibrotic therapies to control cardiac fibrosis. Biomater. Res. 2016, 20, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Fang, Y.; Wan, L.; Xu, Q.; Huang, H.; Zhu, R.; Wu, Q.; Liu, J. Notch signaling inhibits cardiac fibroblast to myofibroblast transformation by antagonizing TGF-β1/Smad3 signaling. J. Cell. Physiol. 2019, 234, 8834–8845. [Google Scholar] [CrossRef]
- Zmorzyński, S.; Styk, W.; Filip, A.A.; Krasowska, D. The significance of Notch pathway in the development of fibrosis in systemic sclerosis. Ann. Dermatol. 2019, 31, 365–371. [Google Scholar] [CrossRef]
- Chen, X.; Qin, M.; Jiang, W.; Zhang, Y.; Liu, X. Electrophysiological characteristics of pressure overload-induced cardiac hypertrophy and its influence on ventricular arrhythmias. PLoS ONE 2017, 12, e0183671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghetti, G.; Eisenberg, C.A.; Signore, S.; Sorrentino, A.; Kaur, K.; Andrade-Vicenty, A.; Edwards, J.G.; Nerkar, M.; Qanud, K.; Sun, N.; et al. Notch signaling modulates the electrical behavior of cardiomyocytes. Am. J. Physiol.-Heart Circ. Physiol. 2018, 314, H68–H81. [Google Scholar] [CrossRef]
- Zhu, R.; Chen, Q.; Liu, Z.; Ruan, H.; Wu, Q.; Zhou, X. Inhibition of the Notch1 pathway induces peripartum cardiomyopathy. J. Cell Mol. Med. 2020, 24, 7907–7914. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Mei, S.; Zhang, S.; Qin, Q.; Li, H.; Liao, Y.; Fan, H.; Liu, Z.; Zhu, H. Cardio-renal exosomes in myocardial infarction serum regulate proangiogenic paracrine signaling in adipose mesenchymal stem cells. Theranostics 2020, 10, 1060–1073. [Google Scholar] [CrossRef]
- Blanco, R.; Gerhardt, H. VEGF and Notch in Tip and Stalk Cell Selection. Cold Spring Harb. Perspect. Med. 2013, 3, a006569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.S.; Tsaryk, R.; Lange, M.; Wisniewski, L.; Moore, J.C.; Lawson, N.; Wojciechowska, K.; Schnittler, H.; Siekmann, A.F. Endothelial Notch signalling limits angiogenesis via control of artery formation. Nat. Cell Biol. 2017, 19, 928–940. [Google Scholar] [CrossRef] [Green Version]
- Pitulescu, M.E.; Schmidt, I.; Giaimo, B.D.; Antoine, T.; Berkenfeld, F.; Ferrante, F.; Park, H.; Ehling, M.; Biljes, D.; Rocha, S.F.; et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat. Cell Biol. 2017, 19, 915–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciria, M.; García, N.A.; Ontoria-Oviedo, I.; González-King, H.; Carrero, R.; De La Pompa, J.L.; Montero, J.A.; Sepúlveda, P. Mesenchymal Stem Cell Migration and Proliferation Are Mediated by Hypoxia-Inducible Factor-1α Upstream of Notch and SUMO Pathways. Stem Cells Dev. 2017, 26, 973–985. [Google Scholar] [CrossRef]
- Zhou, X.-L.; Zhu, R.-R.; Liu, S.; Xu, H.; Xu, X.; Wu, Q.-C.; Liu, J.-C. Notch signaling promotes angiogenesis and improves cardiac function after myocardial infarction. J. Cell. Biochem. 2018, 119, 7105–7112. [Google Scholar] [CrossRef]
- Cahill, T.J.; Choudhury, R.P.; Riley, P.R. Heart regeneration and repair after myocardial infarction: Translational opportunities for novel therapeutics. Nat. Rev. Drug Discov. 2017, 16, 699–717. [Google Scholar] [CrossRef]
- Eschenhagen, T.; Bolli, R.; Braun, T.; Field, L.J.; Fleischmann, B.K.; Frisén, J.; Giacca, M.; Hare, J.M.; Houser, S.; Lee, R.T.; et al. Cardiomyocyte regeneration: A consensus statement. Circulation 2017, 136, 680–686. [Google Scholar] [CrossRef]
- Chakravarty, T.; Makkar, R.R.; Ascheim, D.D.; Traverse, J.H.; Schatz, R.; DeMaria, A.; Francis, G.S.; Povsic, T.J.; Smith, R.R.; Lima, J.A.; et al. ALLogeneic heart STem cells to achieve myocardial regeneration (ALLSTAR) trial: Rationale and design. Cell Transpl. 2017, 26, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Docshin, P.M.; Karpov, A.A.; Mametov, M.V.; Ivkin, D.Y.; Kostareva, A.A.; Malashicheva, A.B. Mechanisms of Regenerative Potential Activation in Cardiac Mesenchymal Cells. Biomedicines 2022, 10, 1283. [Google Scholar] [CrossRef]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef]
- Flink, I.L. Cell cycle reentry of ventricular and atrial cardiomyocytes and cells within the epicardium following amputation of the ventricular apex in the axolotl, Amblystoma mexicanum: Confocal microscopic immunofluorescent image analysis of bromodeoxyuridine-labeled nuclei. Anat. Embryol. 2002, 205, 235–244. [Google Scholar] [CrossRef]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H.A. Transient regenerative potential of the neonatal mouse heart. Science 2011, 331, 1078–1080. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Fan, L.; Zhao, L.; Su, Y. The interaction of Notch and Wnt signaling pathways in vertebrate regeneration. Cell. Regen. 2021, 10, 1–17. [Google Scholar] [CrossRef]
- Gonzalez-King, H.; García, N.A.; Ontoria-Oviedo, I.; Ciria, M.; Montero, J.A.; Sepúlveda, P. Hypoxia Inducible Factor-1α Potentiates Jagged 1-Mediated Angiogenesis by Mesenchymal Stem Cell-Derived Exosomes. Stem Cells 2017, 35, 1747–1759. [Google Scholar] [CrossRef] [Green Version]
- Gálvez-Santisteban, M.; Chen, D.; Zhang, R.; Serrano, R.; Nguyen, C.; Zhao, L.; Nerb, L.; Masutani, E.M.; Vermot, J.; Burns, C.G.; et al. Hemodynamic-mediated endocardial signaling controls in vivo myocardial reprogramming. eLife 2019, 8, e44816. [Google Scholar] [CrossRef]
- Münch, J.; Grivas, D.; González-Rajal, Á.; Torregrosa-Carrión, R.; de la Pompa, J.L. Notch signalling restricts inflammation and Serpine1 expression in the dynamic endocardium of the regenerating zebrafish heart. Development 2017, 144, 1425–1440. [Google Scholar] [CrossRef] [Green Version]
- González-Rosa, J.M.; Burns, C.E.; Burns, C.G. Zebrafish heart regeneration: 15 years of discoveries. Regeneration 2017, 4, 105–123. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.; Hashimoto, H.; Zhou, H.; Morales, M.G.; Chen, B.; Bassel-Duby, R.; Olson, E.N. Notch Inhibition Enhances Cardiac Reprogramming by Increasing MEF2C Transcriptional Activity. Stem Cell Rep. 2017, 8, 548–560. [Google Scholar] [CrossRef]
- Liang, F.; Cheng, S.-Q.; Jin, B.; Tian, Z.; Zou, C.; Jiang, Z. The Mechanism of Myocardial Differentiation and Angiogenesis Regulated by Notch/Dll4 and VEGF Interactions. 2016. Available online: www.ijcem.com/ (accessed on 23 July 2020).
- Zhao, L.; Borikova, A.L.; Ben-Yair, R.; Guner-Ataman, B.; MacRae, C.A.; Lee, R.T.; Burns, C.G. Notch signaling regulates cardiomyocyte proliferation during zebrafish heart regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lu, Q. Plasma membrane-derived extracellular microvesicles mediate non-canonical intercellular NOTCH signaling. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mauretti, A.; Spaans, S.; Bax, N.A.M.; Sahlgren, C.; Bouten, C.V.C. Cardiac Progenitor Cells and the Interplay with Their Microenvironment. Stem Cells Int. 2017, 2017, 1–20. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachanova, O.; Lobov, A.; Malashicheva, A. The Role of the Notch Signaling Pathway in Recovery of Cardiac Function after Myocardial Infarction. Int. J. Mol. Sci. 2022, 23, 12509. https://doi.org/10.3390/ijms232012509
Kachanova O, Lobov A, Malashicheva A. The Role of the Notch Signaling Pathway in Recovery of Cardiac Function after Myocardial Infarction. International Journal of Molecular Sciences. 2022; 23(20):12509. https://doi.org/10.3390/ijms232012509
Chicago/Turabian StyleKachanova, Olga, Arseniy Lobov, and Anna Malashicheva. 2022. "The Role of the Notch Signaling Pathway in Recovery of Cardiac Function after Myocardial Infarction" International Journal of Molecular Sciences 23, no. 20: 12509. https://doi.org/10.3390/ijms232012509
APA StyleKachanova, O., Lobov, A., & Malashicheva, A. (2022). The Role of the Notch Signaling Pathway in Recovery of Cardiac Function after Myocardial Infarction. International Journal of Molecular Sciences, 23(20), 12509. https://doi.org/10.3390/ijms232012509