
Receptive Field Sizes of Nyxnob Mouse Retinal Ganglion Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
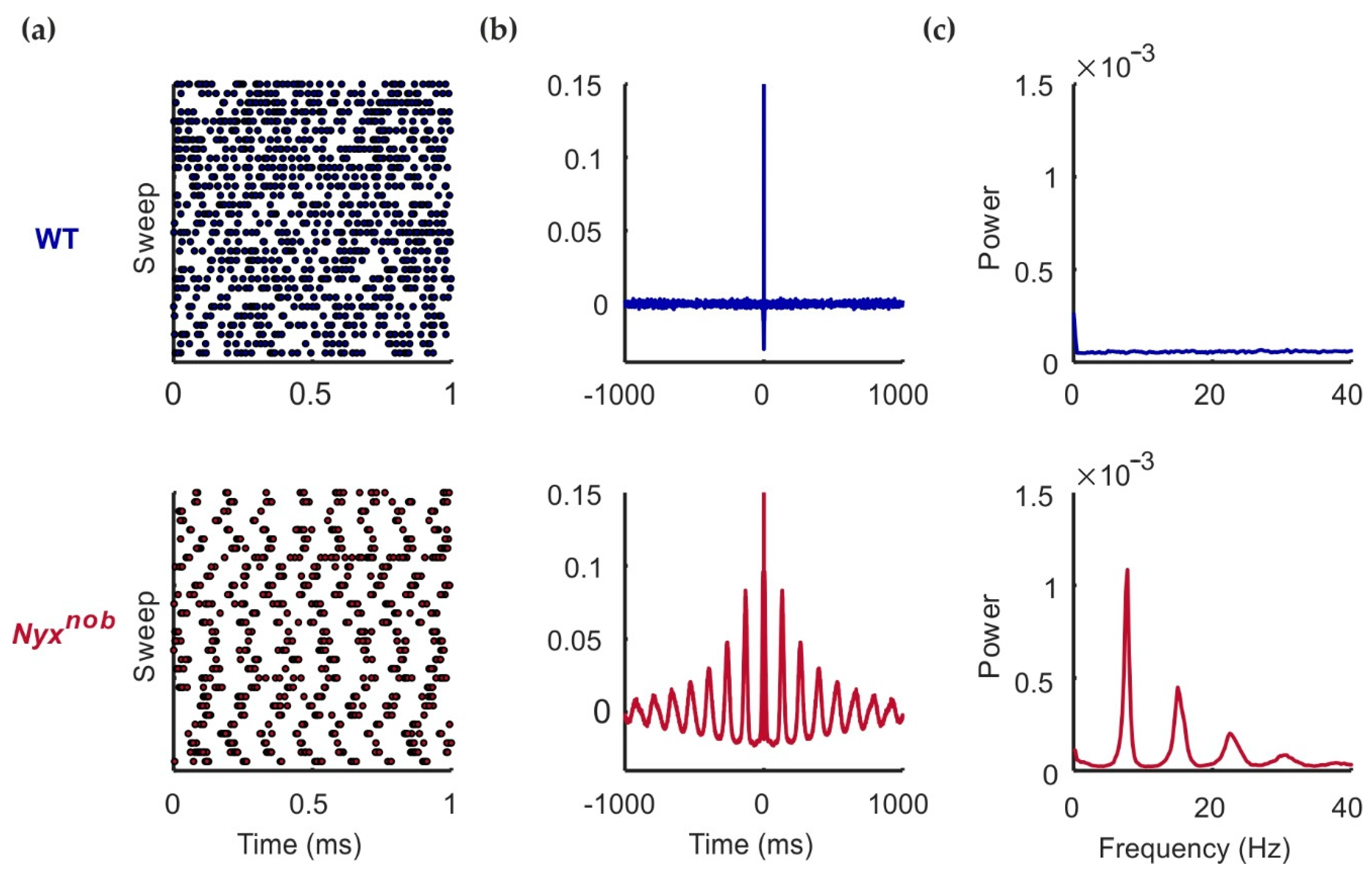
2.1. Nyxnob RGCs Are Spontaneously Oscillating
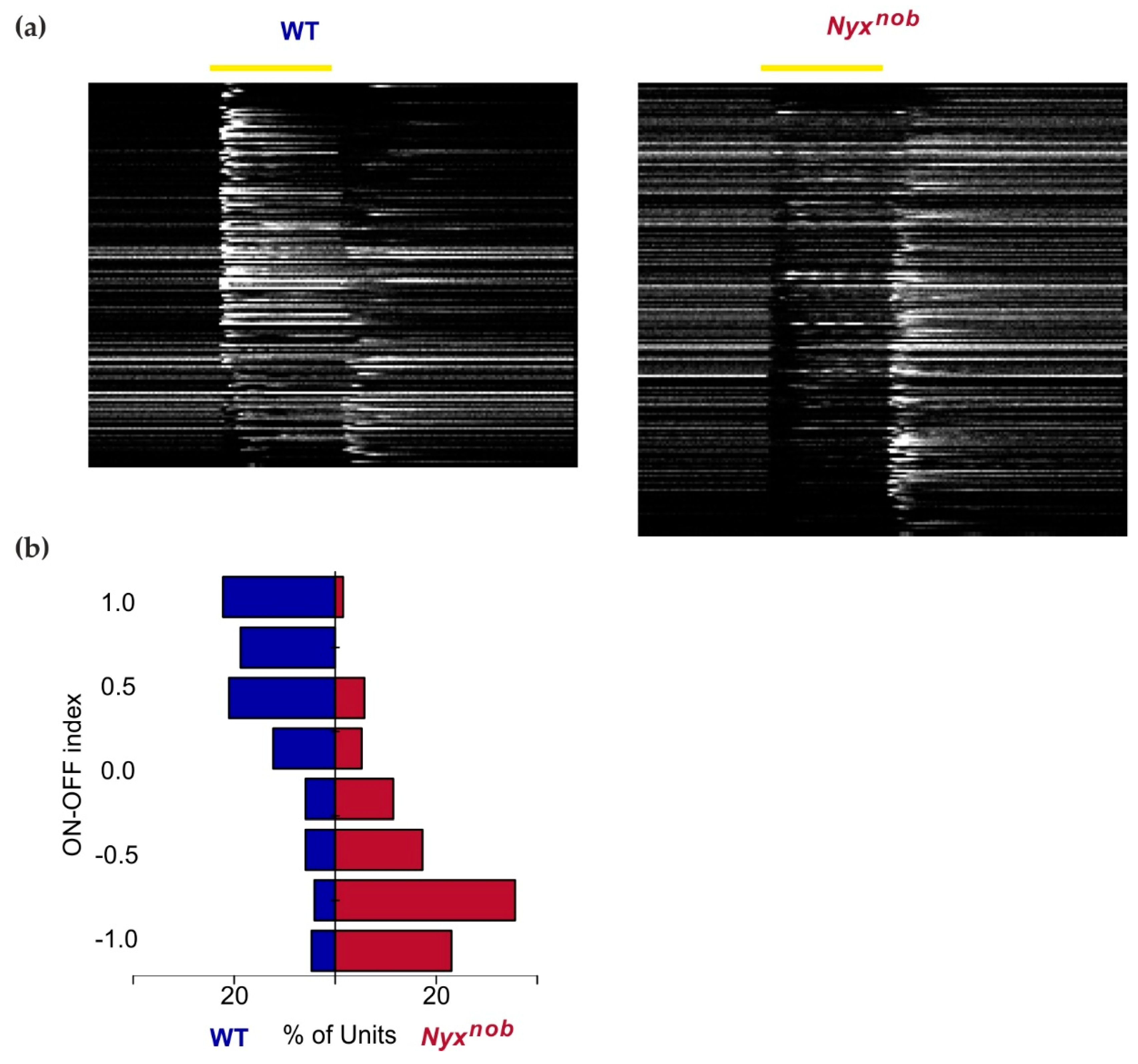
2.2. Nyxnob Retinas Lack Light-Evoked ON-Responses
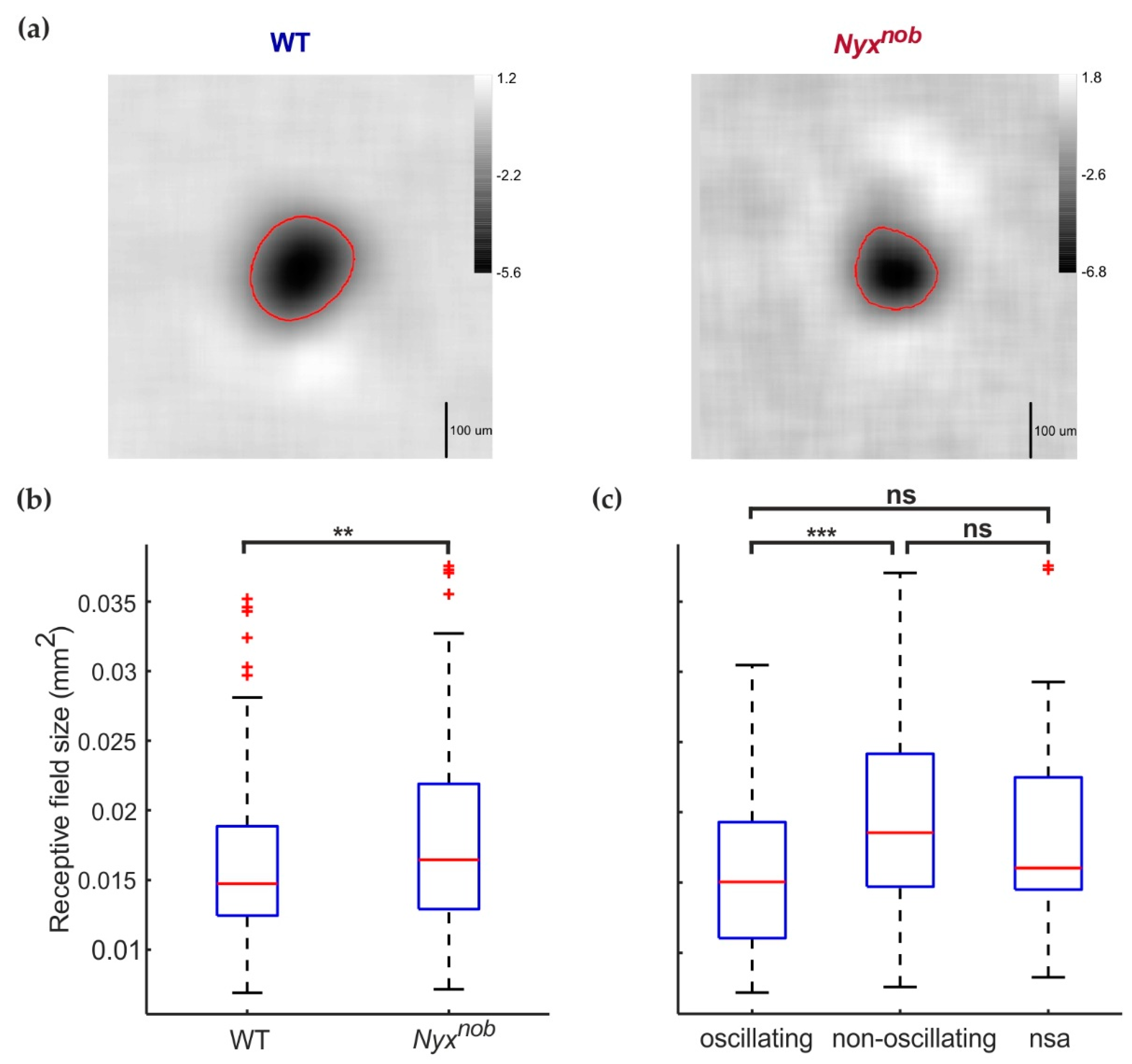
2.3. RGC Receptive Field Sizes Are Similar in WT and Nyxnob
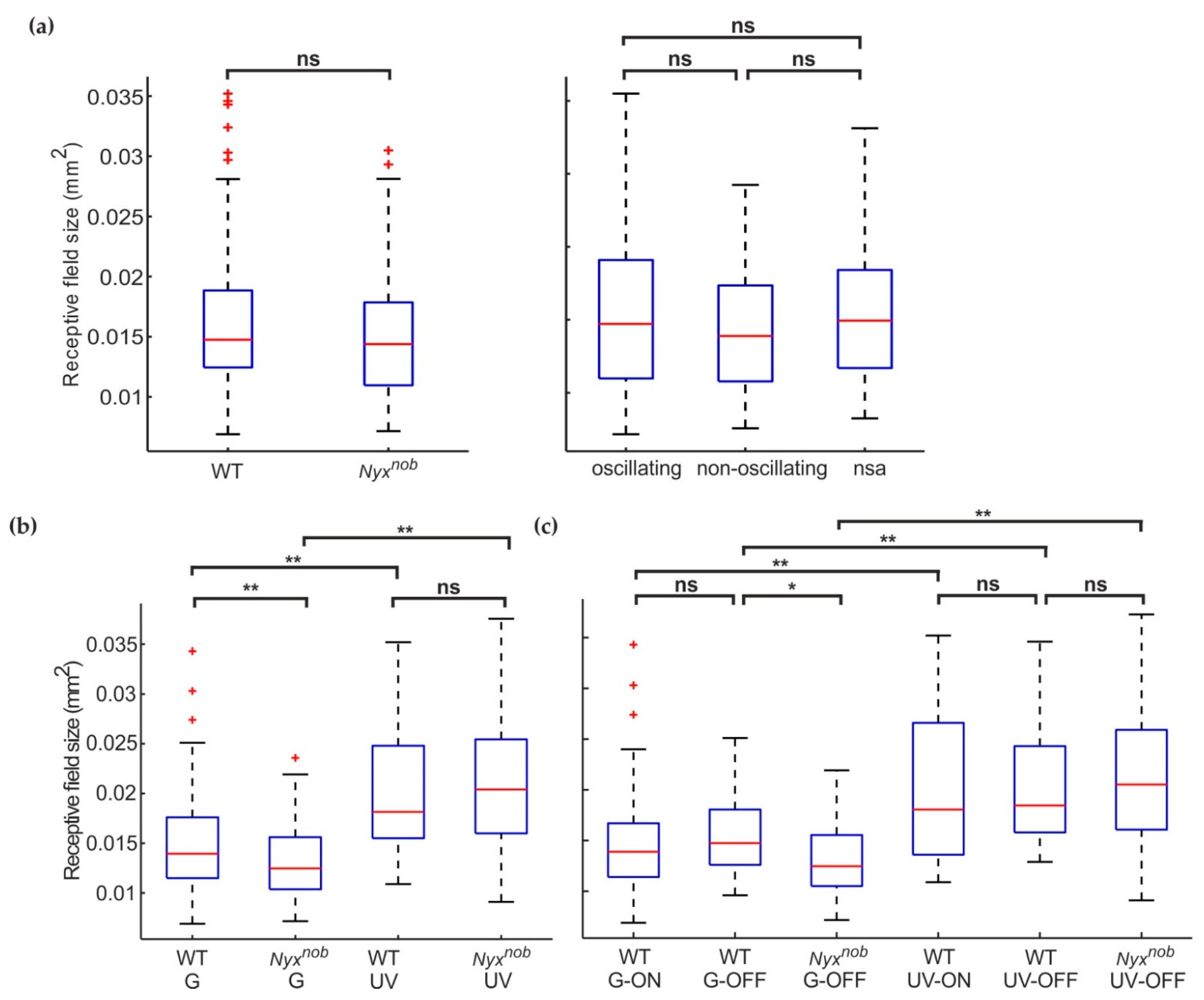
2.4. Green-Light Preferring RGCs Have Smaller Receptive Fields than Do UV-Light Preferring RGCs
2.5. Smaller Receptive Fields of Green-Light Preferring Nyxnob RGCs Compared to WT

2.6. WT ON- and OFF-RGCs Have Similar Receptive Field Sizes
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Retinal Dissection
4.3. Data Acquisition
4.4. Optical Stimulator
4.5. Data Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Optican, L.M.; Zee, D.S. A hypothetical explanation of congenital nystagmus. Biol. Cybern. 1984, 50, 119–134. [Google Scholar] [CrossRef]
- Miyake, Y.; Yagasaki, K.; Horiguchi, M.; Kawase, Y.; Kanda, T. Congenital stationary night blindness with negative electroretinogram: A new classification. Arch. Ophthalmol. 1986, 104, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Bijveld, M.M.C.; Florijn, R.J.; Bergen, A.A.B.; van den Born, L.I.; Kamermans, M.; Prick, L.; Riemslag, F.C.; van Schooneveld, M.J.; Kappers, A.M.; van Genderen, M.M. Genotype and phenotype of 101 dutch patients with congenital stationary night blindness. Ophthalmology 2013, 120, 2072–2081. [Google Scholar] [CrossRef] [PubMed]
- Simonsz, H.J.; Florijn, R.J.; Van Minderhout, H.M.; Bergen, A.A.B.; Kamermans, M. Nightblindness-associated transient tonic downgaze (NATTD) in infant boys with chin-up head posture. Strabismus 2009, 17, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Zubcov, A.A.; Stärk, N.; Weber, A.; Wizov, S.S.; Reinecke, R.D. Improvement of visual acuity after surgery for nystagmus. Ophthalmology 1993, 100, 1488–1497. [Google Scholar] [CrossRef]
- Singh, A.; Ashar, J.; Sharma, P.; Saxena, R.; Menon, V. A prospective evaluation of retroequatorial recession of horizontal rectus muscles and Hertle-Dell’Osso tenotomy procedure in patients with infantile nystagmus with no definite null position. J. Am. Assoc. Pediatric Ophthalmol. Strabismus 2016, 20, 96–99. [Google Scholar] [CrossRef]
- Dell’Osso, L.F.; Flynn, J.T. Congenital nystagmus surgery: A quantitative evaluation of the effects. Arch. Ophthalmol. 1979, 97, 462–469. [Google Scholar] [CrossRef]
- Pardue, M.T.; McCall, M.A.; LaVail, M.M.; Gregg, R.G.; Peachey, N.S. A naturally occurring mouse model of X-linked congenital stationary night blindness. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2443–2449. [Google Scholar]
- Gregg, R.G.; Mukhopadhyay, S.; Candille, S.I.; Ball, S.L.; Pardue, M.T.; McCall, M.A.; Peachey, N.S. Identification of the gene and the mutation responsible for the mouse nob phenotype. Investig. Ophthalmol. Vis. Sci. 2003, 44, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Winkelman, B.H.J.; Howlett, M.H.C.; Hölzel, M.B.; Joling, C.; Fransen, K.H.; Pangeni, G.; Kamermans, S.; Sakuta, H.; Noda, M.; Simonsz, H.J.; et al. Nystagmus in patients with congenital stationary night blindness (CSNB) originates from synchronously firing retinal ganglion cells. PLoS Biol. 2019, 17, e3000174. [Google Scholar] [CrossRef]
- Pusch, C.M.; Zeitz, C.; Brandau, O.; Pesch, K.; Achatz, H.; Feil, S.; Scharfe, C.; Maurer, J.; Jacobi, F.K.; Pinckers, A.; et al. The complete form of X-linked congenital stationary night blindness is caused by mutations in a gene encoding a leucine-rich repeat protein. Nat. Genet. 2000, 26, 324–327. [Google Scholar] [CrossRef]
- Bech-Hansen, N.T.; Naylor, M.J.; Maybaum, T.A.; Sparkes, R.L.; Koop, B.; Birch, D.G.; Bergen, A.A.; Prinsen, C.F.; Polomeno, R.C.; Gal, A.; et al. Mutations in NYX, encoding the leucine-rich proteoglycan nyctalopin, cause X-linked complete congenital stationary night blindness. Nat. Genet. 2000, 26, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Demas, J.; Sagdullaev, B.T.; Green, E.; Jaubert-Miazza, L.; McCall, M.A.; Gregg, R.G.; Wong, R.O.; Guido, W. Failure to maintain eye-specific segregation in nob, a mutant with abnormally patterned retinal activity. Neuron 2006, 50, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Gregg, R.G.; Kamermans, M.; Klooster, J.; Lukasiewicz, P.D.; Peachey, N.S.; Vessey, K.A.; McCall, M.A. Nyctalopin expression in retinal bipolar cells restores visual function in a mouse model of complete X-linked congenital stationary night blindness. J. Neurophysiol. 2007, 98, 3023–3033. [Google Scholar] [CrossRef] [Green Version]
- Koehler, C.L.; Akimov, N.P.; Rentería, R.C. Receptive field center size decreases and firing properties mature in ON and OFF retinal ganglion cells after eye opening in the mouse. J. Neurophysiol. 2011, 106, 895–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantrell, D.R.; Cang, J.; Troy, J.B.; Liu, X. Non-centered spike-triggered covariance analysis reveals neurotrophin-3 as a developmental regulator of receptive field properties of ON-OFF retinal ganglion cells. PLoS Comput. Biol. 2010, 6, e1000967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Xu, H.-P.; Wang, P.; Tian, N. Visual Deprivation Retards the Maturation of Dendritic Fields and Receptive Fields of Mouse Retinal Ganglion Cells. Front. Cell. Neurosci. 2021, 15, 640421. [Google Scholar] [CrossRef]
- Szél, A.; Röhlich, P.; Caffé, A.R.; Juliusson, B.; Aguirre, G.; Van Veen, T. Unique topographic separation of two spectral classes of cones in the mouse retina. J. Comp. Neurol. 1992, 325, 327–342. [Google Scholar] [CrossRef]
- Baden, T.; Schubert, T.; Chang, L.; Wei, T.; Zaichuk, M.; Wissinger, B.; Euler, T. A tale of two retinal domains: Near-optimal sampling of achromatic contrasts in natural scenes through asymmetric photoreceptor distribution. Neuron 2013, 80, 1206–1217. [Google Scholar] [CrossRef] [Green Version]
- Yonehara, K.; Ishikane, H.; Sakuta, H.; Shintani, T.; Nakamura-Yonehara, K.; Kamiji, N.L.; Usui, S.; Noda, M. Identification of retinal ganglion cells and their projections involved in central transmission of information about upward and downward image motion. PLoS ONE 2009, 4, e4320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroga, R.Q.; Nadasdy, Z.; Ben-Shaul, Y. Unsupervised spike detection and sorting with wavelets and superparamagnetic clustering. Neural Comput. 2004, 16, 1661–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hölzel, M.-B.; Howlett, M.H.C.; Kamermans, M. Receptive Field Sizes of Nyxnob Mouse Retinal Ganglion Cells. Int. J. Mol. Sci. 2022, 23, 3202. https://doi.org/10.3390/ijms23063202
Hölzel M-B, Howlett MHC, Kamermans M. Receptive Field Sizes of Nyxnob Mouse Retinal Ganglion Cells. International Journal of Molecular Sciences. 2022; 23(6):3202. https://doi.org/10.3390/ijms23063202
Chicago/Turabian StyleHölzel, Maj-Britt, Marcus H. C. Howlett, and Maarten Kamermans. 2022. "Receptive Field Sizes of Nyxnob Mouse Retinal Ganglion Cells" International Journal of Molecular Sciences 23, no. 6: 3202. https://doi.org/10.3390/ijms23063202
APA StyleHölzel, M. -B., Howlett, M. H. C., & Kamermans, M. (2022). Receptive Field Sizes of Nyxnob Mouse Retinal Ganglion Cells. International Journal of Molecular Sciences, 23(6), 3202. https://doi.org/10.3390/ijms23063202