
Neurogenesis as a Tool for Spinal Cord Injury



{kind=link}
{kind=link}
Abstract
:1. Introduction
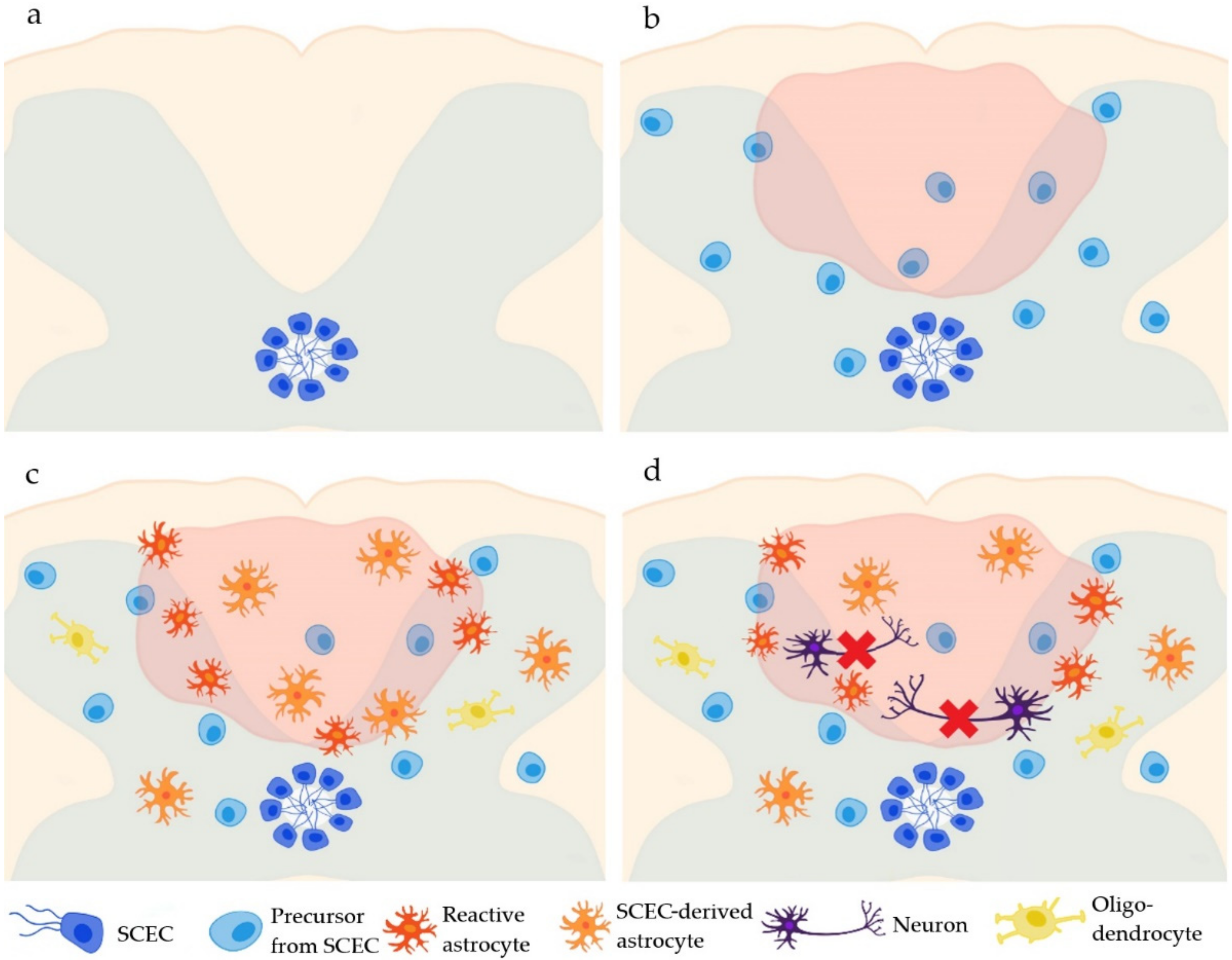
2. Endogenous Neural Stem Cells
SCECs in Spinal Cord Injury
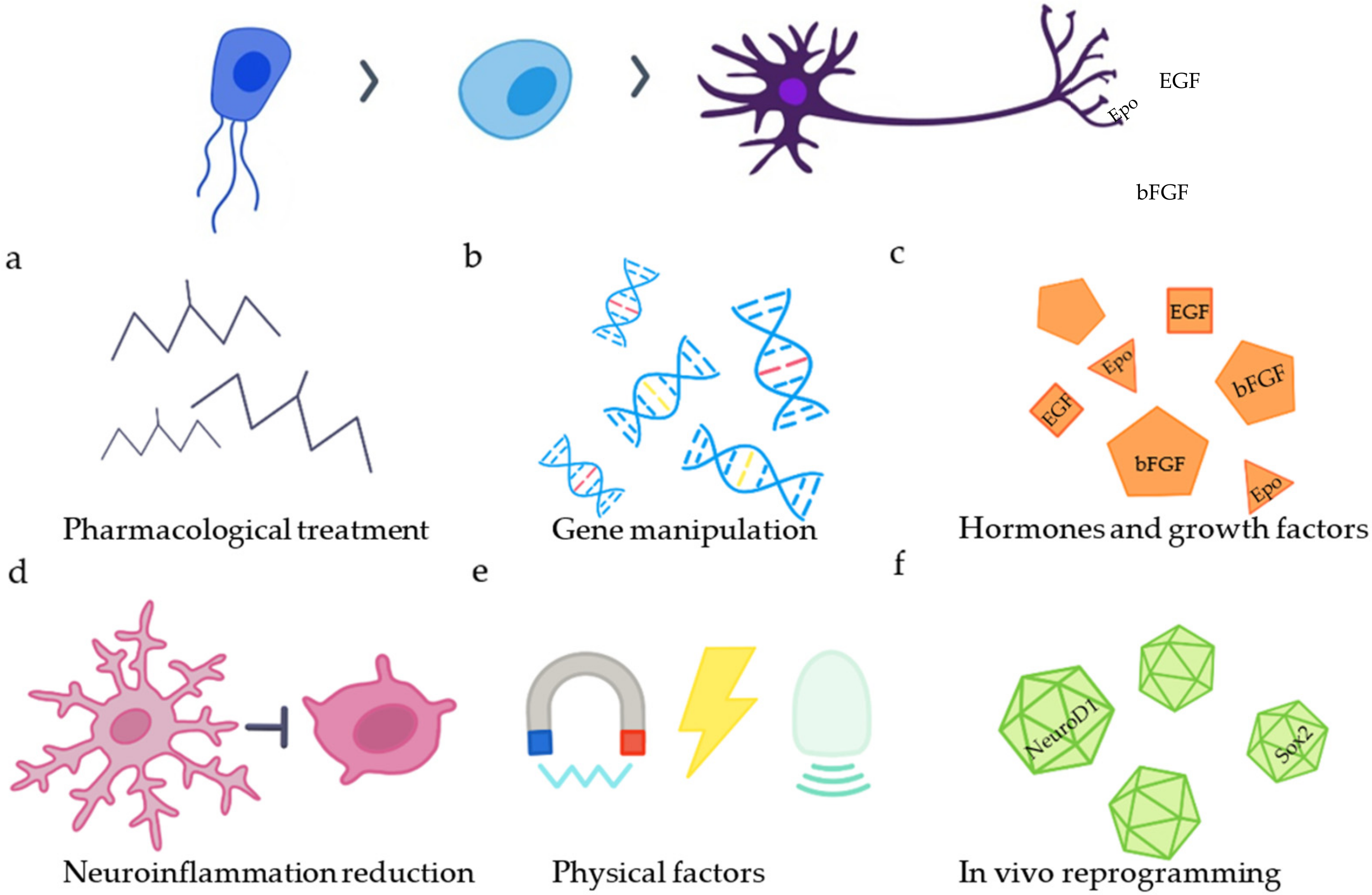
3. Approaches to Promote Neurogenesis
3.1. The Differentiation of SCECs into Neurons
3.2. The Reduction of Neuroinflammation
3.3. Neuromodulation with Physical Factors
3.4. In Vivo Reprogramming
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rowland, J.W.; Hawryluk, G.W.; Kwon, B.; Fehlings, M.G. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef]
- Venkatesh, K.; Ghosh, S.K.; Mullick, M.; Manivasagam, G.; Sen, D. Spinal cord injury: Pathophysiology, treatment strategies, associated challenges, and future implications. Cell Tissue Res. 2019, 377, 125–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.M.; Wu, J.Y.; Li, F.C.; Chen, Q.X. Ion channel blockers and spinal cord injury. J. Neurosci. Res. 2011, 89, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, B.S. Glutamate as a neurotransmitter in the brain: Review of physiology and pathology. J. Nutr. 2000, 130, 1007S–1015S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Zhu, H.; Li, J.; Wang, X.; Misra, H.; Li, Y. Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord 2012, 50, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, S.D.; Comeau, B.; Myers, T.; Sadi, D.; Purdy, M.; Mendez, I. Duration of lipid peroxidation after acute spinal cord injury in rats and the effect of methylprednisolone. Neurosurg. Focus 2008, 25, E5. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Sarkar, C.; Dinizo, M.; Faden, A.I.; Koh, E.Y.; Lipinski, M.M.; Wu, J. Disrupted autophagy after spinal cord injury is associated with er stress and neuronal cell death. Cell Death Dis. 2015, 6, e1582. [Google Scholar] [CrossRef]
- Marsala, J.; Orendacova, J.; Lukacova, N.; Vanicky, I. Traumatic injury of the spinal cord and nitric oxide. Prog. Brain Res. 2007, 161, 171–183. [Google Scholar]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef]
- Han, Q.; Xie, Y.; Ordaz, J.D.; Huh, A.J.; Huang, N.; Wu, W.; Liu, N.; Chamberlain, K.A.; Sheng, Z.H.; Xu, X.M. Restoring cellular energetics promotes axonal regeneration and functional recovery after spinal cord injury. Cell Metab. 2020, 31, 623–641.e8. [Google Scholar] [CrossRef]
- Beattie, M.S.; Hermann, G.E.; Rogers, R.C.; Bresnahan, J.C. Cell death in models of spinal cord injury. Prog. Brain Res. 2002, 137, 37–47. [Google Scholar]
- Oyinbo, C.A. Secondary injury mechanisms in traumatic spinal cord injury: A nugget of this multiply cascade. Acta Neurobiol. Exp. 2011, 71, 281–299. [Google Scholar]
- Rowlands, D.; Sugahara, K.; Kwok, J.C. Glycosaminoglycans and glycomimetics in the central nervous system. Molecules 2015, 20, 3527–3548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal-Galicia, P.; Chavez-Hernandez, M.E.; Mata, F.; Mata-Luevanos, J.; Rodriguez-Serrano, L.M.; Tapia-de-Jesus, A.; Buenrostro-Jauregui, M.H. Adult neurogenesis: A story ranging from controversial new neurogenic areas and human adult neurogenesis to molecular regulation. Int. J. Mol. Sci. 2021, 22, 11489. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, O.; Spalding, K.L.; Frisen, J. Adult neurogenesis in humans. Cold Spring Harb. Perspect. Biol. 2015, 7, a018994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, A.B.; Hoshino, H.S.R.; Ortega, N.C.; Dos Santos, B.G.S.; Teixeira, M.J. Adult human neurogenesis: Early studies clarify recent controversies and go further. Metab. Brain Dis. 2022, 37, 153–172. [Google Scholar] [CrossRef] [PubMed]
- Barnabe-Heider, F.; Goritz, C.; Sabelstrom, H.; Takebayashi, H.; Pfrieger, F.W.; Meletis, K.; Frisen, J. Origin of new glial cells in intact and injured adult spinal cord. Cell Stem Cell 2010, 7, 470–482. [Google Scholar] [CrossRef] [Green Version]
- Adrian, E.K., Jr.; Walker, B.E. Incorporation of thymidine-h3 by cells in normal and injured mouse spinal cord. J. Neuropathol. Exp. Neurol. 1962, 21, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Horner, P.J.; Power, A.E.; Kempermann, G.; Kuhn, H.G.; Palmer, T.D.; Winkler, J.; Thal, L.J.; Gage, F.H. Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J. Neurosci. 2000, 20, 2218–2228. [Google Scholar] [CrossRef]
- Meletis, K.; Barnabe-Heider, F.; Carlen, M.; Evergren, E.; Tomilin, N.; Shupliakov, O.; Frisen, J. Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol. 2008, 6, e182. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Dunne, C.; Hewson, J.; Wohl, C.; Wheatley, M.; Peterson, A.C.; Reynolds, B.A. Multipotent cns stem cells are present in the adult mammalian spinal cord and ventricular neuroaxis. J. Neurosci. 1996, 16, 7599–7609. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Nakagomi, N.; Nakano-Doi, A.; Ishikawa, H.; Tatsumi, Y.; Bando, Y.; Yoshikawa, H.; Matsuyama, T.; Gomi, F.; Nakagomi, T. Potential of adult endogenous neural stem/progenitor cells in the spinal cord to contribute to remyelination in experimental autoimmune encephalomyelitis. Cells 2019, 8, 1025. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lu, Y.; Fang, X.; Zhang, J.; Li, J.; Li, S.; Deng, X.; Yu, Y.; Xu, R. An astrocyte regenerative response from vimentin-containing cells in the spinal cord of amyotrophic lateral sclerosis’s disease-like transgenic (g93a sod1) mice. Neurodegener. Dis. 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Cizkova, D.; Nagyova, M.; Slovinska, L.; Novotna, I.; Radonak, J.; Cizek, M.; Mechirova, E.; Tomori, Z.; Hlucilova, J.; Motlik, J.; et al. Response of ependymal progenitors to spinal cord injury or enhanced physical activity in adult rat. Cell Mol. Neurobiol. 2009, 29, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, B.; Wang, L.; Long, Z.; Li, Y.; Liao, W.; Wu, Y. Endogenous neural stem cells in central canal of adult rats acquired limited ability to differentiate into neurons following mild spinal cord injury. Int. J. Clin. Exp. Pathol. 2015, 8, 3835–3842. [Google Scholar]
- Yang, H.; Lu, P.; McKay, H.M.; Bernot, T.; Keirstead, H.; Steward, O.; Gage, F.H.; Edgerton, V.R.; Tuszynski, M.H. Endogenous neurogenesis replaces oligodendrocytes and astrocytes after primate spinal cord injury. J. Neurosci. 2006, 26, 2157–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhauser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Hamby, M.E.; Sofroniew, M.V. Reactive astrocytes as therapeutic targets for cns disorders. Neurotherapeutics 2010, 7, 494–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabelstrom, H.; Stenudd, M.; Reu, P.; Dias, D.O.; Elfineh, M.; Zdunek, S.; Damberg, P.; Goritz, C.; Frisen, J. Resident neural stem cells restrict tissue damage and neuronal loss after spinal cord injury in mice. Science 2013, 342, 637–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawryluk, G.W.; Mothe, A.J.; Chamankhah, M.; Wang, J.; Tator, C.; Fehlings, M.G. In vitro characterization of trophic factor expression in neural precursor cells. Stem Cells Dev. 2012, 21, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, C.P.; Holmstrom, N.A.; Lilja, J.A.; Schweinhardt, P.; Hao, J.; Spenger, C.; Wiesenfeld-Hallin, Z.; Kurpad, S.N.; Frisen, J.; Olson, L. Allodynia limits the usefulness of intraspinal neural stem cell grafts; directed differentiation improves outcome. Nat. Neurosci. 2005, 8, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Shihabuddin, L.S.; Horner, P.J.; Ray, J.; Gage, F.H. Adult spinal cord stem cells generate neurons after transplantation in the adult dentate gyrus. J. Neurosci. 2000, 20, 8727–8735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abematsu, M.; Tsujimura, K.; Yamano, M.; Saito, M.; Kohno, K.; Kohyama, J.; Namihira, M.; Komiya, S.; Nakashima, K. Neurons derived from transplanted neural stem cells restore disrupted neuronal circuitry in a mouse model of spinal cord injury. J. Clin. Investig. 2010, 120, 3255–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, T.; Zhou, H.; Wang, T.; Lu, L.; Li, F.; Liu, B.; Kong, X.; Feng, S. In vitro characteristics of valproic acid and all-trans-retinoic acid and their combined use in promoting neuronal differentiation while suppressing astrocytic differentiation in neural stem cells. Brain Res. 2015, 1596, 31–47. [Google Scholar] [CrossRef]
- Chu, W.; Yuan, J.; Huang, L.; Xiang, X.; Zhu, H.; Chen, F.; Chen, Y.; Lin, J.; Feng, H. Valproic acid arrests proliferation but promotes neuronal differentiation of adult spinal nspcs from sci rats. Neurochem. Res. 2015, 40, 1472–1486. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Johnson, G.V. The glamour and gloom of glycogen synthase kinase-3. Trends Biochem. Sci. 2004, 29, 95–102. [Google Scholar] [CrossRef]
- Takahashi-Yanaga, F.; Sasaguri, T. The wnt/beta-catenin signaling pathway as a target in drug discovery. J. Pharmacol. Sci. 2007, 104, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.T.; Wang, Z.; Hunsberger, J.G.; Chuang, D.M. Therapeutic potential of mood stabilizers lithium and valproic acid: Beyond bipolar disorder. Pharmacol. Rev. 2013, 65, 105–142. [Google Scholar] [CrossRef] [Green Version]
- Kostrouchova, M.; Kostrouch, Z.; Kostrouchova, M. Valproic acid, a molecular lead to multiple regulatory pathways. Folia Biol. 2007, 53, 37–49. [Google Scholar]
- Chu, T.; Zhou, H.; Lu, L.; Kong, X.; Wang, T.; Pan, B.; Feng, S. Valproic acid-mediated neuroprotection and neurogenesis after spinal cord injury: From mechanism to clinical potential. Regen. Med. 2015, 10, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jimenez, F.J.; Vilches, A.; Perez-Arago, M.A.; Clemente, E.; Roman, R.; Leal, J.; Castro, A.A.; Fustero, S.; Moreno-Manzano, V.; Jendelova, P.; et al. Activation of neurogenesis in multipotent stem cells cultured in vitro and in the spinal cord tissue after severe injury by inhibition of glycogen synthase kinase-3. Neurotherapeutics 2021, 18, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, G.; Ye, J.; Lu, D.; Chen, Z.; Xiang, A.P.; Jiang, M.H. Substance p enhances endogenous neurogenesis to improve functional recovery after spinal cord injury. Int. J. Biochem. Cell Biol. 2017, 89, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Pal, R.; Bondar, V.V.; Adamski, C.J.; Rodney, G.G.; Sardiello, M. Inhibition of erk1/2 restores gsk3beta activity and protein synthesis levels in a model of tuberous sclerosis. Sci. Rep. 2017, 7, 4174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, Y.; Wang, X.; Evers, B.M. Glycogen synthase kinase-3 is a negative regulator of extracellular signal-regulated kinase. Oncogene 2006, 25, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Chao, X.J.; Wu, J.F.; Cheng, B.C.; Su, T.; Fu, X.Q.; Li, T.; Guo, H.; Tse, A.K.; Kwan, H.Y.; et al. Erk/gsk3beta signaling is involved in atractylenolide i-induced apoptosis and cell cycle arrest in melanoma cells. Oncol. Rep. 2015, 34, 1543–1548. [Google Scholar] [CrossRef]
- Tan, B.T.; Wang, L.; Li, S.; Long, Z.Y.; Wu, Y.M.; Liu, Y. Retinoic acid induced the differentiation of neural stem cells from embryonic spinal cord into functional neurons in vitro. Int. J. Clin. Exp. Pathol. 2015, 8, 8129–8135. [Google Scholar]
- Zhao, H.; Zuo, X.; Ren, L.; Li, Y.; Tai, H.; Du, J.; Xie, X.; Zhang, X.; Han, Y.; Wu, Y.; et al. Combined use of bfgf/egf and all-trans-retinoic acid cooperatively promotes neuronal differentiation and neurite outgrowth in neural stem cells. Neurosci. Lett 2019, 690, 61–68. [Google Scholar] [CrossRef]
- Liu, C.; Sun, R.; Huang, J.; Zhang, D.; Huang, D.; Qi, W.; Wang, S.; Xie, F.; Shen, Y.; Shen, C. The baf45d protein is preferentially expressed in adult neurogenic zones and in neurons and may be required for retinoid acid induced pax6 expression. Front. Neuroanat. 2017, 11, 94. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, J.; Liu, C.; Liu, L.; Shen, Y.; Shen, C.; Liu, C. Baf45d downregulation in spinal cord ependymal cells following spinal cord injury in adult rats and its potential role in the development of neuronal lesions. Front. Neurosci. 2019, 13, 1151. [Google Scholar] [CrossRef] [PubMed]
- Fabbiani, G.; Reali, C.; Valentin-Kahan, A.; Rehermann, M.I.; Fagetti, J.; Falco, M.V.; Russo, R.E. Connexin signaling is involved in the reactivation of a latent stem cell niche after spinal cord injury. J. Neurosci. 2020, 40, 2246–2258. [Google Scholar] [CrossRef]
- Rodriguez-Jimenez, F.J.; Alastrue, A.; Stojkovic, M.; Erceg, S.; Moreno-Manzano, V. Connexin 50 modulates sox2 expression in spinal-cord-derived ependymal stem/progenitor cells. Cell Tissue Res. 2016, 365, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Niu, W.; Liu, M.L.; Zou, Y.; Zhang, C.L. In vivo conversion of astrocytes to neurons in the injured adult spinal cord. Nat Commun. 2014, 5, 3338. [Google Scholar] [CrossRef] [PubMed]
- Nekoui, A.; Blaise, G. Erythropoietin and nonhematopoietic effects. Am. J. Med. Sci. 2017, 353, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Simon, F.H.; Erhart, P.; Vcelar, B.; Scheuerle, A.; Schelzig, H.; Oberhuber, A. Erythropoietin preconditioning improves clinical and histologic outcome in an acute spinal cord ischemia and reperfusion rabbit model. J. Vasc Surg. 2016, 64, 1797–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassouna, I.; Ott, C.; Wustefeld, L.; Offen, N.; Neher, R.A.; Mitkovski, M.; Winkler, D.; Sperling, S.; Fries, L.; Goebbels, S.; et al. Revisiting adult neurogenesis and the role of erythropoietin for neuronal and oligodendroglial differentiation in the hippocampus. Mol. Psychiatry 2016, 21, 1752–1767. [Google Scholar] [CrossRef]
- Zhang, H.; Fang, X.; Huang, D.; Luo, Q.; Zheng, M.; Wang, K.; Cao, L.; Yin, Z. Erythropoietin signaling increases neurogenesis and oligodendrogenesis of endogenous neural stem cells following spinal cord injury both in vivo and in vitro. Mol. Med. Rep. 2018, 17, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting neurotrophins to specific populations of neurons: Ngf, bdnf, and nt-3 and their relevance for treatment of spinal cord injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, F.; Cheng, F.; Ying, L.; Wang, C.; Shi, K.; Wang, J.; Xia, K.; Gong, Z.; Huang, X.; et al. Strategies and prospects of effective neural circuits reconstruction after spinal cord injury. Cell Death Dis. 2020, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- Kanakasabai, S.; Pestereva, E.; Chearwae, W.; Gupta, S.K.; Ansari, S.; Bright, J.J. Ppargamma agonists promote oligodendrocyte differentiation of neural stem cells by modulating stemness and differentiation genes. PLoS ONE 2012, 7, e50500. [Google Scholar] [CrossRef] [PubMed]
- Jimenez Hamann, M.C.; Tator, C.H.; Shoichet, M.S. Injectable intrathecal delivery system for localized administration of egf and fgf-2 to the injured rat spinal cord. Exp. Neurol. 2005, 194, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Kojima, A.; Tator, C.H. Epidermal growth factor and fibroblast growth factor 2 cause proliferation of ependymal precursor cells in the adult rat spinal cord in vivo. J. Neuropathol. Exp. Neurol. 2000, 59, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Kojima, A.; Tator, C.H. Intrathecal administration of epidermal growth factor and fibroblast growth factor 2 promotes ependymal proliferation and functional recovery after spinal cord injury in adult rats. J. Neurotrauma 2002, 19, 223–238. [Google Scholar] [CrossRef]
- Ohori, Y.; Yamamoto, S.; Nagao, M.; Sugimori, M.; Yamamoto, N.; Nakamura, K.; Nakafuku, M. Growth factor treatment and genetic manipulation stimulate neurogenesis and oligodendrogenesis by endogenous neural progenitors in the injured adult spinal cord. J. Neurosci. 2006, 26, 11948–11960. [Google Scholar] [CrossRef] [Green Version]
- Oudega, M.; Hao, P.; Shang, J.; Haggerty, A.E.; Wang, Z.; Sun, J.; Liebl, D.J.; Shi, Y.; Cheng, L.; Duan, H.; et al. Validation study of neurotrophin-3-releasing chitosan facilitation of neural tissue generation in the severely injured adult rat spinal cord. Exp. Neurol. 2019, 312, 51–62. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, A.; Duan, H.; Zhang, S.; Hao, P.; Ye, K.; Sun, Y.E.; Li, X. Nt3-chitosan elicits robust endogenous neurogenesis to enable functional recovery after spinal cord injury. Proc. Natl. Acad. Sci. USA 2015, 112, 13354–13359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, J.S.; Zhao, C.; Zhang, A.; Duan, H.; Hao, P.; Wei, R.H.; Shang, J.; Zhao, W.; Liu, Z.; Yu, J.; et al. Nt3-chitosan enables de novo regeneration and functional recovery in monkeys after spinal cord injury. Proc. Natl. Acad. Sci. USA 2018, 115, E5595–E5604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Song, W.; Zhao, W.; Gao, Y.; Shang, J.; Hao, P.; Yang, Z.; Duan, H.; Li, X. Application of the sodium hyaluronate-cntf scaffolds in repairing adult rat spinal cord injury and facilitating neural network formation. Sci. China Life Sci. 2018, 61, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, C.; Haggerty, A.E.; Yan, J.; Lan, M.; Seu, M.; Yang, M.; Marlow, M.M.; Maldonado-Lasuncion, I.; Cho, B.; et al. The effect of a nanofiber-hydrogel composite on neural tissue repair and regeneration in the contused spinal cord. Biomaterials 2020, 245, 119978. [Google Scholar] [CrossRef]
- Liu, H.; Xu, X.; Tu, Y.; Chen, K.; Song, L.; Zhai, J.; Chen, S.; Rong, L.; Zhou, L.; Wu, W.; et al. Engineering microenvironment for endogenous neural regeneration after spinal cord injury by reassembling extracellular matrix. ACS Appl. Mater. Interfaces 2020, 12, 17207–17219. [Google Scholar] [CrossRef] [PubMed]
- Koprivica, V.; Cho, K.S.; Park, J.B.; Yiu, G.; Atwal, J.; Gore, B.; Kim, J.A.; Lin, E.; Tessier-Lavigne, M.; Chen, D.F.; et al. Egfr activation mediates inhibition of axon regeneration by myelin and chondroitin sulfate proteoglycans. Science 2005, 310, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xiao, Z.; Chen, B.; Dai, J. The neuronal differentiation microenvironment is essential for spinal cord injury repair. Organogenesis 2017, 13, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Li, X.; Xiao, Z.; Zhao, Y.; Liang, H.; Wang, B.; Han, S.; Li, X.; Xu, B.; Wang, N.; et al. A modified collagen scaffold facilitates endogenous neurogenesis for acute spinal cord injury repair. Acta Biomater. 2017, 51, 304–316. [Google Scholar] [CrossRef]
- Fan, C.; Li, X.; Zhao, Y.; Xiao, Z.; Xue, W.; Sun, J.; Li, X.; Zhuang, Y.; Chen, Y.; Dai, J. Cetuximab and taxol co-modified collagen scaffolds show combination effects for the repair of acute spinal cord injury. Biomater. Sci. 2018, 6, 1723–1734. [Google Scholar] [CrossRef]
- Li, X.; Xiao, Z.; Han, J.; Chen, L.; Xiao, H.; Ma, F.; Hou, X.; Li, X.; Sun, J.; Ding, W.; et al. Promotion of neuronal differentiation of neural progenitor cells by using egfr antibody functionalized collagen scaffolds for spinal cord injury repair. Biomaterials 2013, 34, 5107–5116. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, Y.; Cheng, S.; Han, S.; Shu, M.; Chen, B.; Chen, X.; Tang, F.; Wang, N.; Tu, Y.; et al. Cetuximab modified collagen scaffold directs neurogenesis of injury-activated endogenous neural stem cells for acute spinal cord injury repair. Biomaterials 2017, 137, 73–86. [Google Scholar] [CrossRef]
- Lu, T.M.; Houghton, S.; Magdeldin, T.; Durán, J.G.B.; Minotti, A.P.; Snead, A.; Sproul, A.; Nguyen, D.T.; Xiang, J.; Fine, H.A.; et al. Pluripotent stem cell-derived epithelium misidentified as brain microvascular endothelium requires ETS factors to acquire vascular fate. Proc Natl Acad Sci USA 2021, 118, e2016950118. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Ao, Y.; O’Shea, T.M.; Burda, J.E.; Bernstein, A.M.; Brumm, A.J.; Muthusamy, N.; Ghashghaei, H.T.; Carmichael, S.T.; Cheng, L.; et al. Ependymal cell contribution to scar formation after spinal cord injury is minimal, local and dependent on direct ependymal injury. Sci. Rep. 2017, 7, 41122. [Google Scholar] [CrossRef] [Green Version]
- Shah, P.T.; Stratton, J.A.; Stykel, M.G.; Abbasi, S.; Sharma, S.; Mayr, K.A.; Koblinger, K.; Whelan, P.J.; Biernaskie, J. Single-cell transcriptomics and fate mapping of ependymal cells reveals an absence of neural stem cell function. Cell 2018, 173, 1045–1057.e1049. [Google Scholar] [CrossRef] [Green Version]
- Cawsey, T.; Duflou, J.; Weickert, C.S.; Gorrie, C.A. Nestin-positive ependymal cells are increased in the human spinal cord after traumatic central nervous system injury. J. Neurotrauma 2015, 32, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.P.; Coulter, M.; Miotke, J.; Meyer, R.L.; Takemaru, K.; Levine, J.M. Abrogation of beta-catenin signaling in oligodendrocyte precursor cells reduces glial scarring and promotes axon regeneration after cns injury. J. Neurosci. 2014, 34, 10285–10297. [Google Scholar] [CrossRef] [Green Version]
- Lukovic, D.; Stojkovic, M.; Moreno-Manzano, V.; Jendelova, P.; Sykova, E.; Bhattacharya, S.S.; Erceg, S. Concise review: Reactive astrocytes and stem cells in spinal cord injury: Good guys or bad guys? Stem Cells 2015, 33, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, W.S.; Abdel-Hamid, G.A.; Al-Karim, S.; Zakar, N.; Elassouli, M.Z. Neuroectodermal stem cells: A remyelinating potential in acute compressed spinal cord injury in rat model. J. Biosci. 2018, 43, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Deng, M.; Zhao, X.Q.; Liu, M. Alternatively polarized macrophages regulate the growth and differentiation of ependymal stem cells through the sirt2 pathway. Exp. Neurobiol. 2020, 29, 150–163. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, P.; Liu, X.; Li, J.; Wan, Y.; Liu, S.; Wei, F. Overexpression of rictor in the injured spinal cord promotes functional recovery in a rat model of spinal cord injury. FASEB J. 2020, 34, 6984–6998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Barrera, R.; Flores-Romero, A.; Garcia, E.; Fernandez-Presas, A.M.; Incontri-Abraham, D.; Navarro-Torres, L.; Garcia-Sanchez, J.; Juarez-Vignon Whaley, J.J.; Madrazo, I.; Ibarra, A. Immunization with neural-derived peptides increases neurogenesis in rats with chronic spinal cord injury. CNS Neurosci. Ther. 2020, 26, 650–658. [Google Scholar] [CrossRef]
- Ye, J.; Qin, Y.; Tang, Y.; Ma, M.; Wang, P.; Huang, L.; Yang, R.; Chen, K.; Chai, C.; Wu, Y.; et al. Methylprednisolone inhibits the proliferation of endogenous neural stem cells in nonhuman primates with spinal cord injury. J. Neurosurg. Spine 2018, 29, 199–207. [Google Scholar] [CrossRef]
- Zheng, Y.; Mao, Y.R.; Yuan, T.F.; Xu, D.S.; Cheng, L.M. Multimodal treatment for spinal cord injury: A sword of neuroregeneration upon neuromodulation. Neural Regen. Res. 2020, 15, 1437–1450. [Google Scholar] [PubMed]
- Blackmore, J.; Shrivastava, S.; Sallet, J.; Butler, C.R.; Cleveland, R.O. Ultrasound neuromodulation: A review of results, mechanisms and safety. Ultrasound Med. Biol. 2019, 45, 1509–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Ge, H.; Zhao, H.; Zou, Y.; Chen, Y.; Feng, H. Electromagnetic fields for the regulation of neural stem cells. Stem Cells Int. 2017, 2017, 9898439. [Google Scholar] [CrossRef] [Green Version]
- Abbasnia, K.; Ghanbari, A.; Abedian, M.; Ghanbari, A.; Sharififar, S.; Azari, H. The effects of repetitive transcranial magnetic stimulation on proliferation and differentiation of neural stem cells. Anat. Cell Biol. 2015, 48, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Liu, X.; Yang, T.; Mei, Y.A.; Hu, C. Exposure to extremely low-frequency electromagnetic fields inhibits t-type calcium channels via aa/lte4 signaling pathway. Cell Calcium 2014, 55, 48–58. [Google Scholar] [CrossRef] [Green Version]
- He, Y.L.; Liu, D.D.; Fang, Y.J.; Zhan, X.Q.; Yao, J.J.; Mei, Y.A. Exposure to extremely low-frequency electromagnetic fields modulates na+ currents in rat cerebellar granule cells through increase of aa/pge2 and ep receptor-mediated camp/pka pathway. PLoS ONE 2013, 8, e54376. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.C.; Ha, K.Y.; Kim, Y.H.; Kim, J.W.; Cho, Y.K.; Kim, S.I. Induction of endogenous neural stem cells by extracorporeal shock waves after spinal cord injury. Spine 2018, 43, E200–E207. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H. Neuronal reprogramming in treating spinal cord injury. Neural Regen. Res. 2022, 17, 1440–1445. [Google Scholar] [PubMed]
- Tai, W.; Wu, W.; Wang, L.L.; Ni, H.; Chen, C.; Yang, J.; Zang, T.; Zou, Y.; Xu, X.M.; Zhang, C.L. In vivo reprogramming of ng2 glia enables adult neurogenesis and functional recovery following spinal cord injury. Cell Stem Cell 2021, 28, 923–937.e4. [Google Scholar] [CrossRef] [PubMed]
- Puls, B.; Ding, Y.; Zhang, F.; Pan, M.; Lei, Z.; Pei, Z.; Jiang, M.; Bai, Y.; Forsyth, C.; Metzger, M.; et al. Regeneration of functional neurons after spinal cord injury via in situ neurod1-mediated astrocyte-to-neuron conversion. Front. Cell Dev. Biol. 2020, 8, 591883. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Su, Z.; Tai, W.; Zou, Y.; Xu, X.M.; Zhang, C.L. The p53 pathway controls sox2-mediated reprogramming in the adult mouse spinal cord. Cell Rep. 2016, 17, 891–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Tao, X.; Sui, M.; Cui, M.; Liu, D.; Wang, B.; Wang, T.; Zheng, Y.; Luo, J.; Mu, Y.; et al. Reprogramming astrocytes to motor neurons by activation of endogenous ngn2 and isl1. Stem Cell Rep. 2021, 16, 1777–1791. [Google Scholar] [CrossRef]
- Paniagua-Torija, B.; Norenberg, M.; Arevalo-Martin, A.; Carballosa-Gautam, M.M.; Campos-Martin, Y.; Molina-Holgado, E.; Garcia-Ovejero, D. Cells in the adult human spinal cord ependymal region do not proliferate after injury. J. Pathol. 2018, 246, 415–421. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havelikova, K.; Smejkalova, B.; Jendelova, P. Neurogenesis as a Tool for Spinal Cord Injury. Int. J. Mol. Sci. 2022, 23, 3728. https://doi.org/10.3390/ijms23073728
Havelikova K, Smejkalova B, Jendelova P. Neurogenesis as a Tool for Spinal Cord Injury. International Journal of Molecular Sciences. 2022; 23(7):3728. https://doi.org/10.3390/ijms23073728
Chicago/Turabian StyleHavelikova, Katerina, Barbora Smejkalova, and Pavla Jendelova. 2022. "Neurogenesis as a Tool for Spinal Cord Injury" International Journal of Molecular Sciences 23, no. 7: 3728. https://doi.org/10.3390/ijms23073728
APA StyleHavelikova, K., Smejkalova, B., & Jendelova, P. (2022). Neurogenesis as a Tool for Spinal Cord Injury. International Journal of Molecular Sciences, 23(7), 3728. https://doi.org/10.3390/ijms23073728